
Human Peopling and Population Dynamics in Sicily: Preliminary Analysis of the Craniofacial Morphometric Variation from the Paleolithic to the Contemporary Age

Abstract
1. Introduction
1.1. Human Biodiversity in Sicily
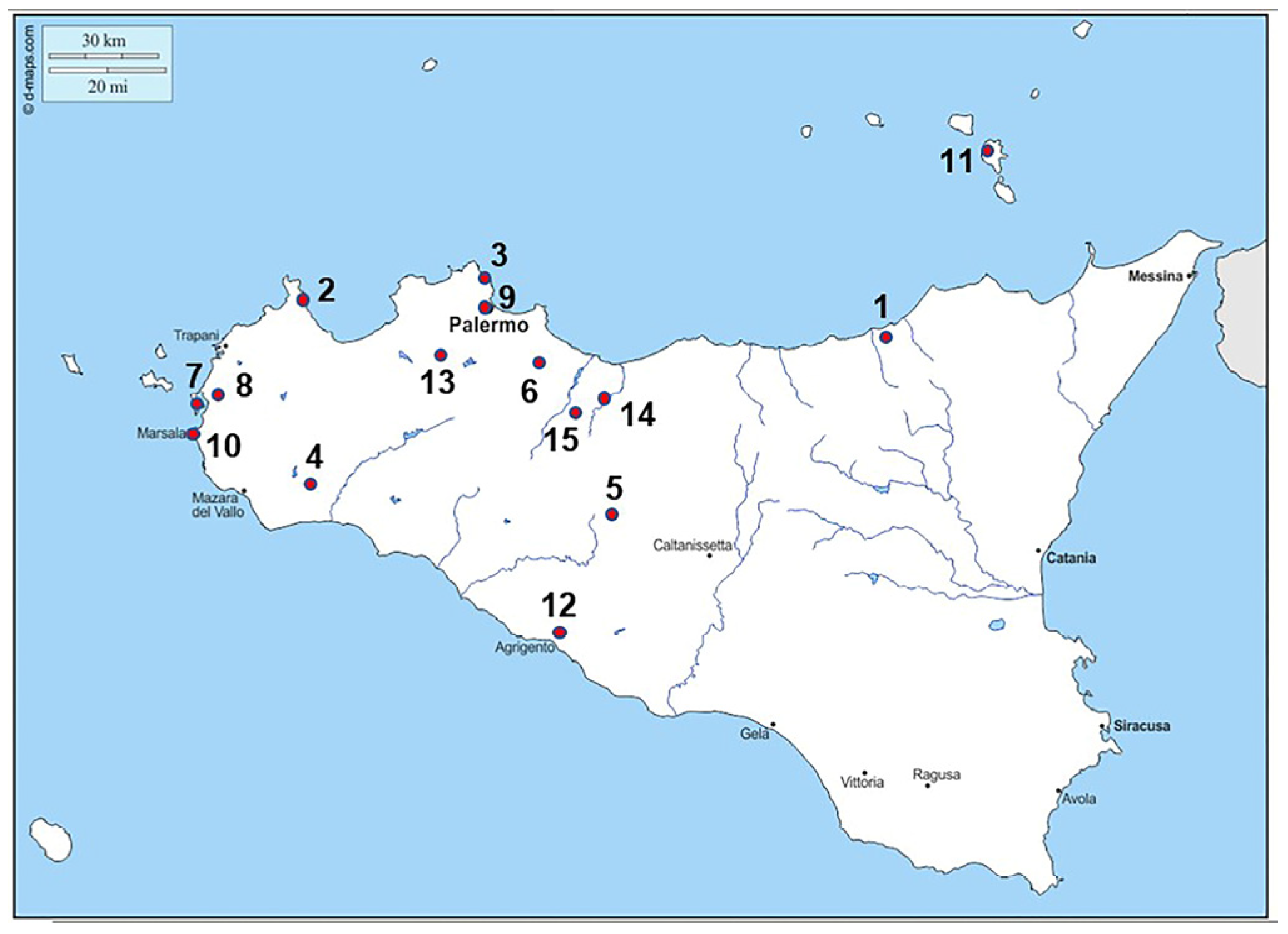
1.2. Background Studies and Sample Recognition

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Place | ||
| San Teodoro | Garilli et al. [41] Sineo et al. [50] | D’Amore et al. [31] |
| Uzzo | Borgognini & Repetto [27] Borgognini et al. [28] | Mannino et al. [47] Costantini [30] |
| Molara | Borgognini et al. [26] Becker [22] | |
| Marcita | Di Salvo [33,34] La Rocca [46] | Becker [22] |
| Polizzello | De Miro [8] Messina et al. [48] | Hodos [38] |
| Baucina | Castellana & Mallegni [29] Belvedere et al. [5] | Micciche’ et al. [48] |
| Mozia | Becker [21,22,23] Sconzo & Falzone [19] | Lauria et al. [43,44,45] |
| Birgi | Griffo [12,13] Fama’ & Toti [11] | |
| Tukory | Germana’ & Di Salvo [42] Di Stefano [10] | |
| Phoenician/Punic of PA | not published | |
| Lilibeo | Becker [24] Bechtold et al. [22] | |
| Marsala | La Duca [16] Becker [24] | Di Salvo [36] |
| Lipari | Brea & Cavalier [6] Cavalier [7] | |
| Agrigento | Fiorentino et al. [37] | |
| C. San Pietro | Di Salvo [35] Di Stefano [9] | |
| M. Iato | Di Salvo [35] Kistler [14,15] | |
| Caltavuturo | Romana [18] Vassallo [20] | |
| Alia | Mannino [17] | |
| Rotoli | not published |
1.3. Aim of the Study
2. Materials and Methods
2.1. Cranial Samples
2.2. Methods
3. Results
3.1. Shape Variation and Changes in Direction
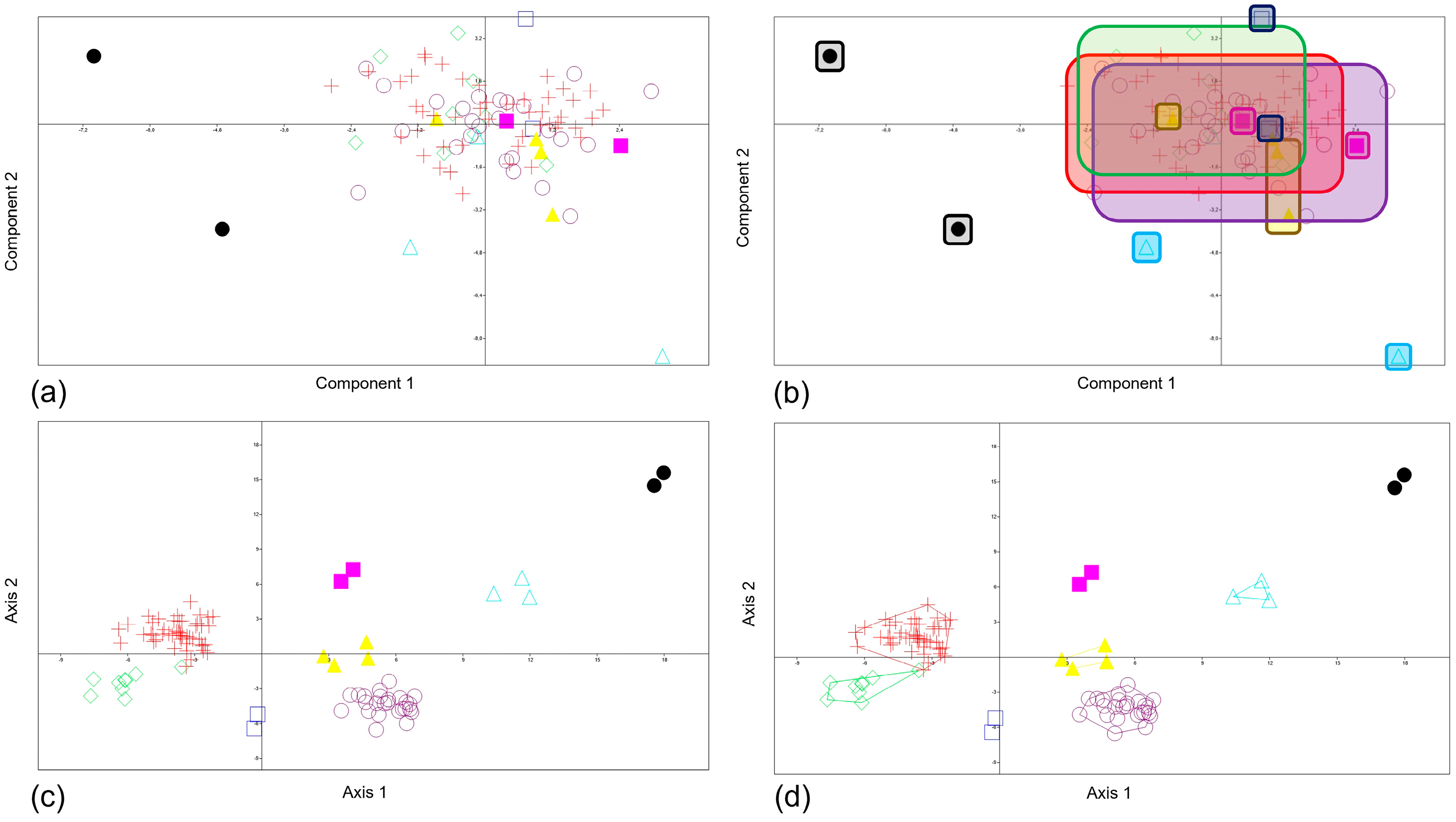
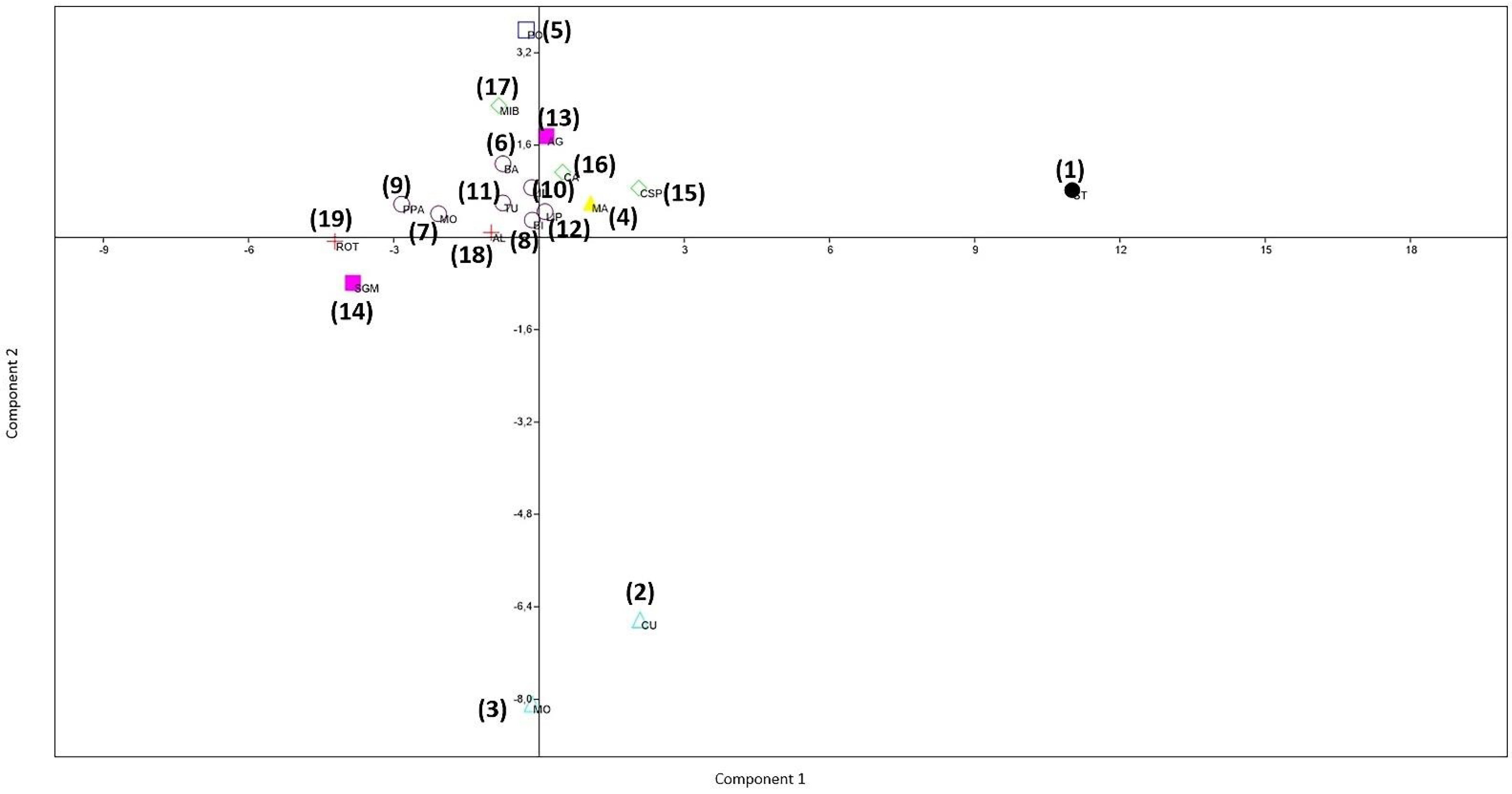
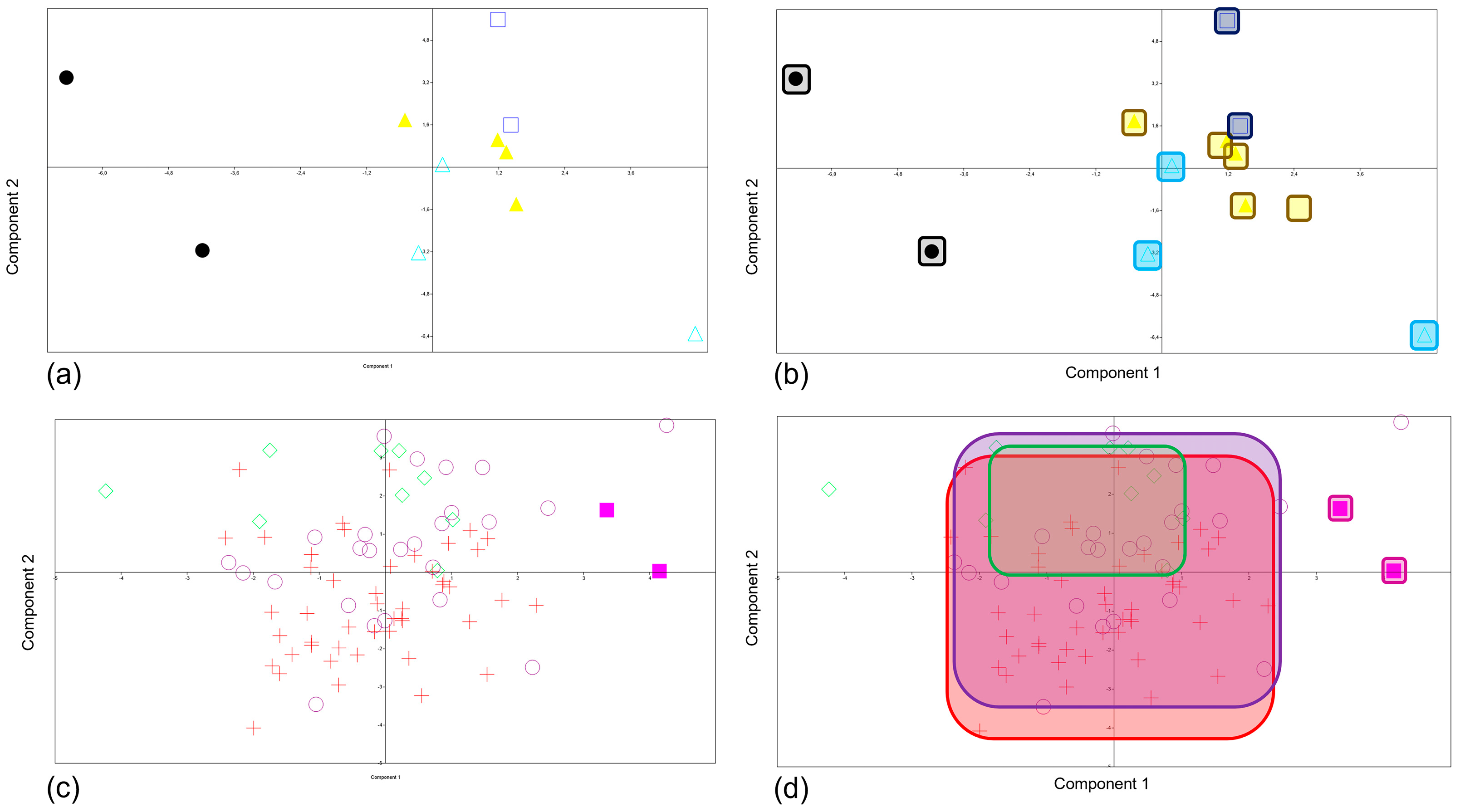
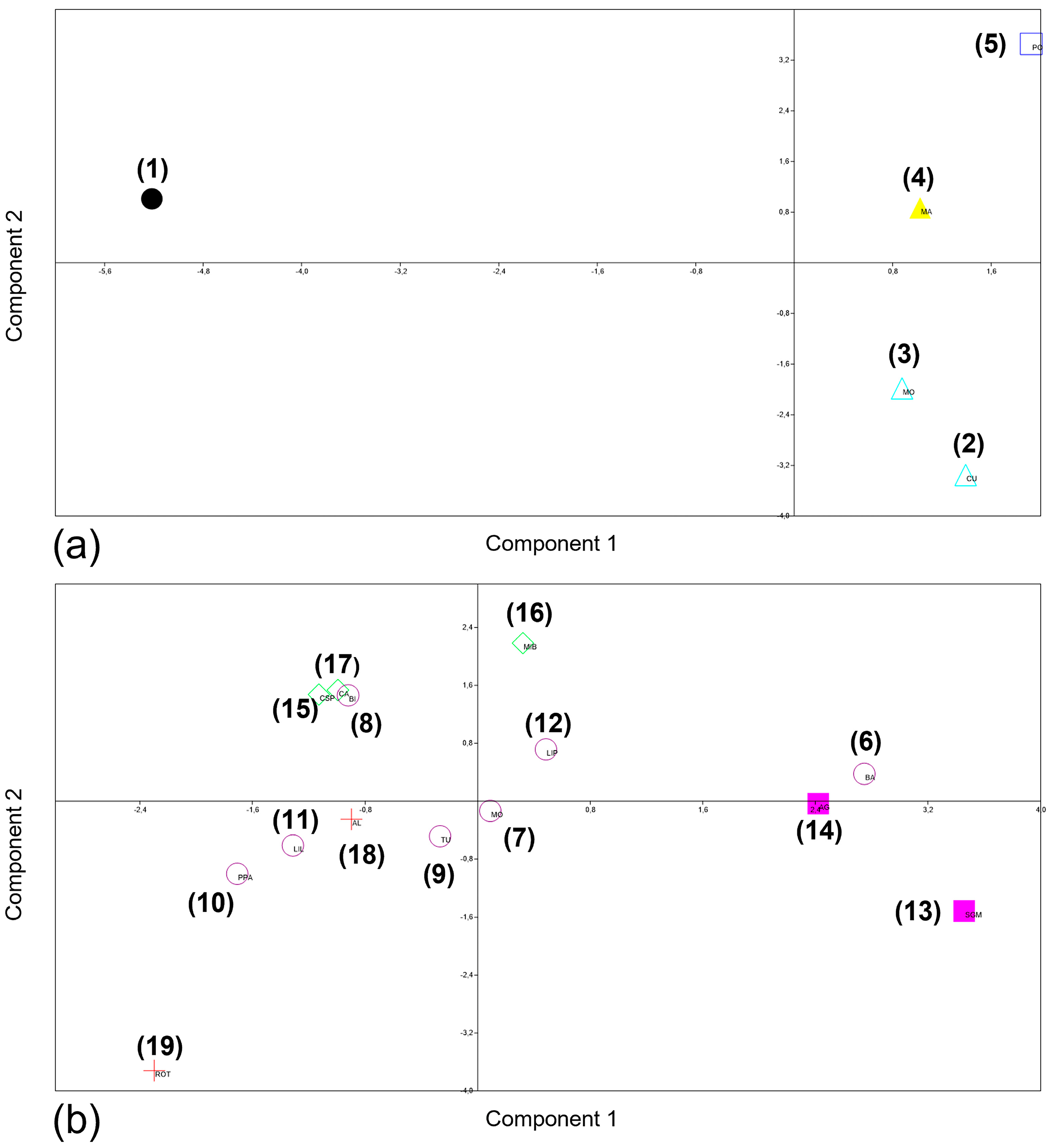
3.2. PCA and MANOVA/CVA Procrustes Coordinates in Shape Space
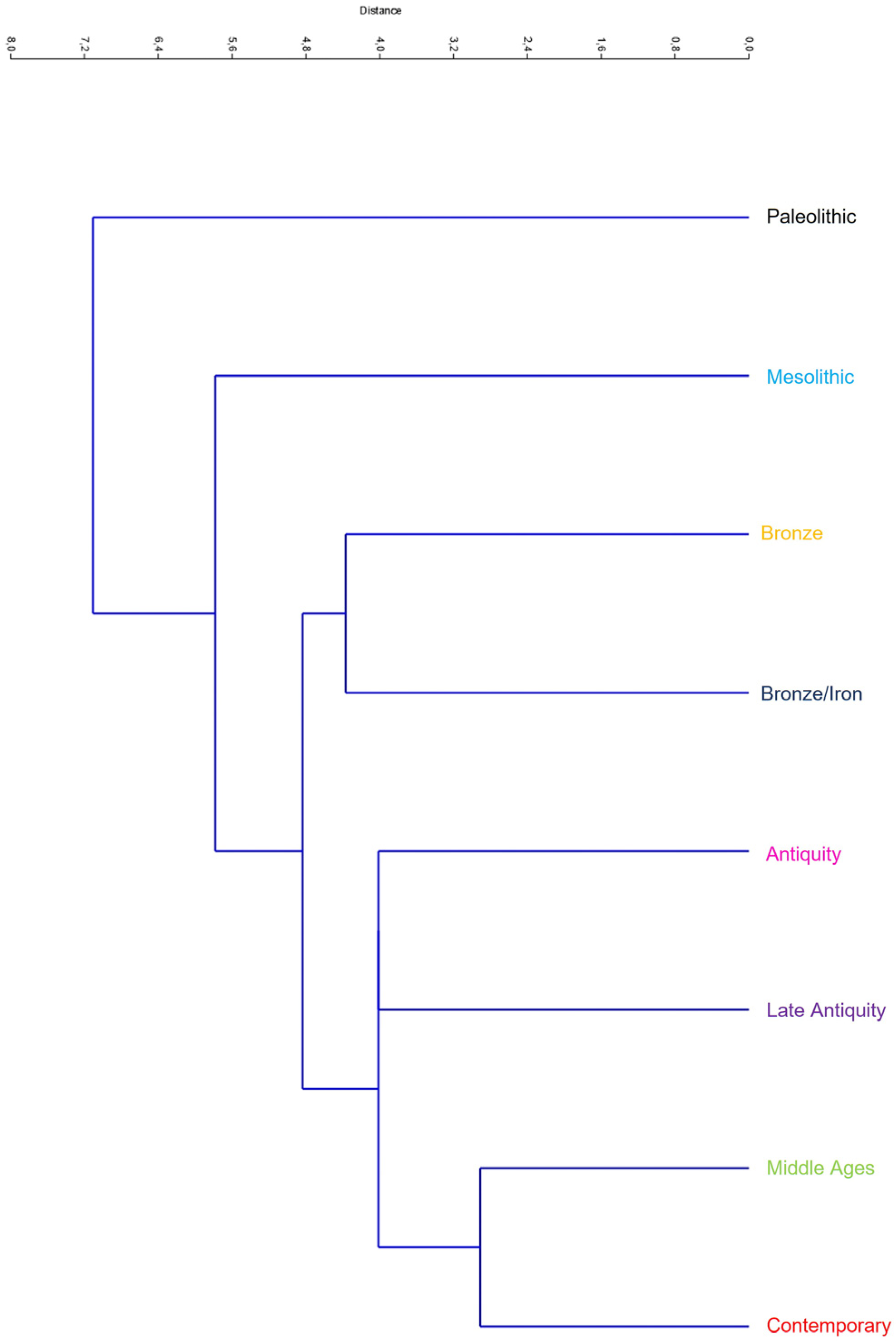
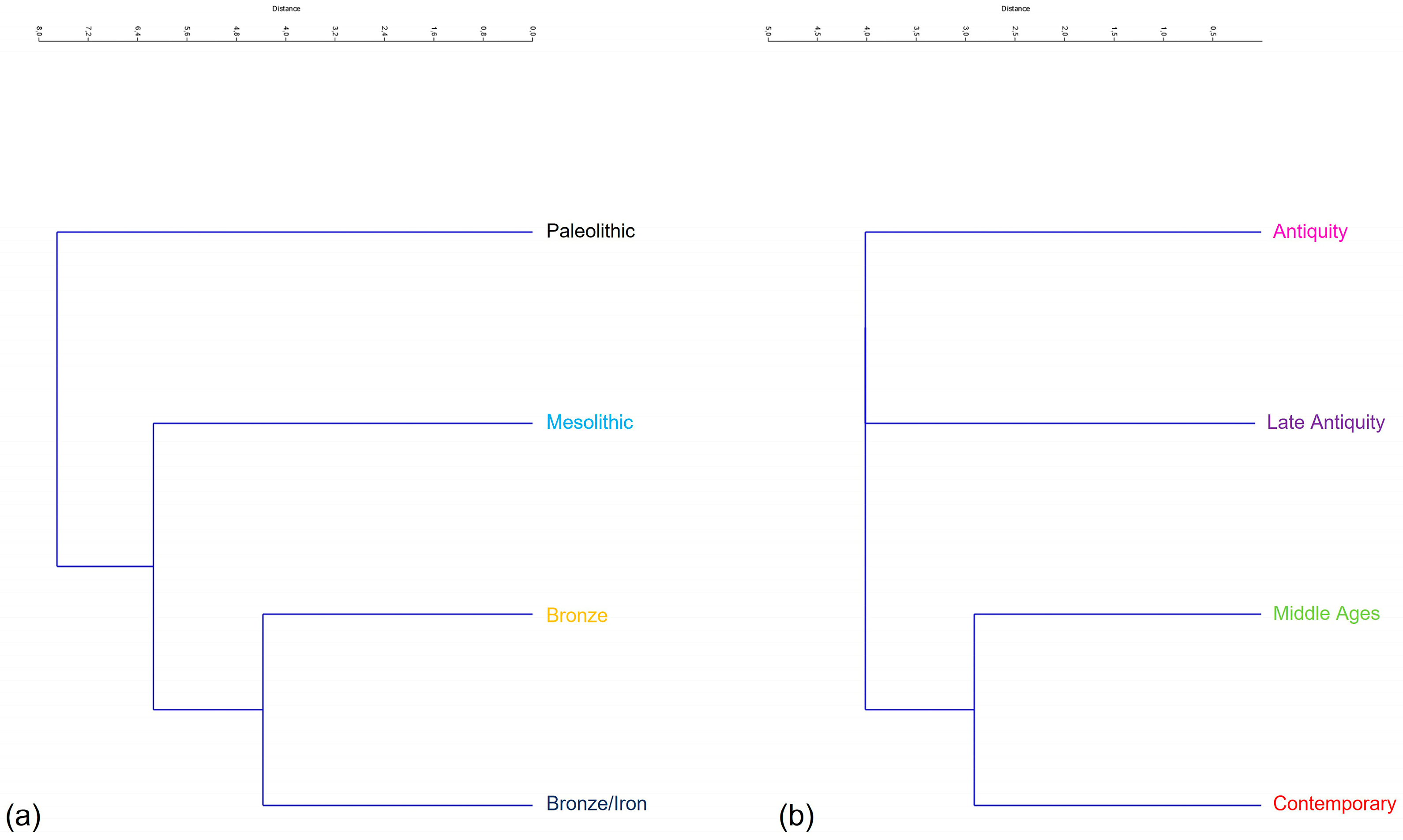
3.3. Neighbour-Joining of Procrustes Coordinates
4. Prehistory and History: Geometric Morphometrics
4.1. Prehistory and History: Shape Variation and Comparison
- Group 1-Prehistory: Upper-Palaeolithic, Mesolithic, Bronze, Iron.
- Group 2-History: Antiquity, Late Antiquity, Middle Ages, Contemporary.
4.2. Prehistory and History Shape Changes
4.3. Neighbor-Joining of Procrustes Coordinates
5. Discussion
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Massa, B.; Sbordoni, V.; Vigna Taglianti, A. In La Biogeografia della Sicilia. Proceeeding of the XXXVII Congresso della Societa’Italiana di Biogeografia, Catania, Italy, 7–1 ottobre 2018. Biogeogr. J. Integr. Biogeogr. 2011, 30, 685–694. [Google Scholar]
- Ruggieri, G. Due parole sulla paleogeografia delle isole minori a Ovest ea Nord della Sicilia. Biogeogr. J. Integr. Biogeogr. 1973, 3, 5–12. [Google Scholar] [CrossRef]
- Pignatti, S. La flora della Sicilia come chiave di lettura per la fitogeografia mediterranea: Una visione autobiografica. Biogeogr. J. Integr. Biogeogr. 2011, 30, 71–94. [Google Scholar] [CrossRef]
- Bechtold, B.; Frey-Kupper, S.; Madella, M.; Brugnone, A. La Necropoli di Lilybaeum; L’Erma di Bretschneider: Trapani, Italy, 1999. [Google Scholar]
- Belvedere, O.; Burgio, A.; Bordonaro, G.; Forgia, V. Baucina (Pa)–Monte Falcone 2014 Indagini nella necropoli. In FOLD&R; FastiOnLine Documents & Research: Roma, Italy, 2017; pp. 1–7. [Google Scholar]
- Brea, L.B.; Cavalier, M. Meligunìs Lipára: La Necropoli Greca e Romana Nella Contrada Diana; Flaccovio: Palermo, Italy, 1995; Volume 2. [Google Scholar]
- Cavalier, M. New Greek and Roman discoveries from Lipari. In Mediterranean Archaeology; Meditarch: Sideny, Australia, 1995; pp. 83–88. [Google Scholar]
- De Miro, E. Polizzello, centro della Sicania. In QuadMess; Università Degli Studi di Messina. Istituto di Archeologia: Messina, Italy, 1988; Volume 3, pp. 25–41. [Google Scholar]
- Di Stefano, C.A.; Cadei, A. Federico e la Sicilia. Dalla Terra Alla Corona; Ediprint: Torino, Italy, 1997. [Google Scholar]
- Di Stefano, C.A. Palermo Punica; Sellerio: Palermo, Italy, 1998. [Google Scholar]
- Famà, M.L.; Toti, M.P. La Necropoli di Birgi: Un Esempio D’interazione Culturale Tra Fenici e Greci Nell’eterno Banchetto. In Nel Mondo di Ade: Ideologie, Spazi e Rituali Funerari Per L’eterno Banchetto (Secoli VIII-IV a.C.), Proceedings of the Convegno Internazionale, Ragusa-Gela, Montirone, Italy, 6–8 June 2010; Collana di Studi Archeologici, Triskeles: Montirone, Italy, 2019; pp. 395–409. [Google Scholar]
- Griffo, M.G. La necropoli diBirgi. In Seconde Giornate Internazionali di Studi Sull’area Elima, Proceedings of the Giornate Internazionali di Studi Sull’area elima, Gibellina, Italy, 22–26 October 1977; Scuola Normale di Pisa: Pisa, Italy, 1997; pp. 909–921. [Google Scholar]
- Griffo, M.G. La necropoli di Birgi. In Lilibeo e il Suo Territorio, Proceeding of Centro Internazionale di Studi Fenici, Punici e Romani Per l’archeologia Marsalese; E. Caruso, A., Ed.; Spanò Giammellaroc: Marsala, Italy, 2008; pp. 169–176. [Google Scholar]
- Kistler, E. Glocal responses from archaic Sicily. Anc. West East 2012, 11, 219–233. [Google Scholar]
- Kistler, E. Monte Iato, Sicily. In The Encyclopedia of Ancient History; John Wiley & Sons: Hoboken, NJ, USA, 2013. [Google Scholar]
- La Duca, R. Storia di Palermo I.I. Dal Tardo-Antico All’Islam; L’Epos: Palermo, Italy, 2000. [Google Scholar]
- Mannino, G. Alia, il complesso rupestre della Gurfa. Notiziario Archeologico della Soprintendenza di Palermo; Regione Siciliana-Assessorato dei Beni Culturali e Dell’identità Siciliana: Palermo, Italy, 2016; Volume 8, pp. 1–39. [Google Scholar]
- Romana, L. Caltavuturo. Atlante dei Beni Culturali; Istituto Poligrafico Europeo: Palermo, Italy, 2009. [Google Scholar]
- Sconzo, P.; Falsone, G. New investigations in the North-East quarter of Motya: The archaic cemetery and Building. In Proceedings of the 8th International Congress of Phoenician and Punic Studies, Carbonia, Italy, 21–26 October 2013; Folia Fenicia, Fabrizio Serra Publisher: Roma, Italy, 2018; pp. 62–69. [Google Scholar]
- Vassallo, S. Indagini preliminari alla Terravecchia di Caltavuturo. In Kokalos; L’Erma di Bretschneider: Palermo, Italy, 2009. [Google Scholar]
- Becker, M.J. Metric and non-metric data from a series of skulls from mozia, sicily and a related site. In Antropologia Contemporanea; Il Mulino: Bologna, Italy, 1985; Volume 8, p. 211. [Google Scholar]
- Becker, M.J. Skeletal studies of Sicilian populations. A survey. J. Accord. Res. Inst. 1995, 6, 83–117. [Google Scholar]
- Becker, M.J. Identifying an 8th–7th century BC Cemetery at Mozia, Sicily: Evaulation of redeposited human skeletal remains to test an archaeological hypothesis. In Sicilia Archeologica; L’Erma di Bretschneider: Palermo, Italy, 1998; Volume 31, pp. 7–12. [Google Scholar]
- Becker, M.J. Skeletal studies of the people of Sicily: An update on research into human remains from archaeological contexts. Int. J. Anthropol. 2000, 15, 191–239. [Google Scholar] [CrossRef]
- Bonfiglio, L.; Marra, A.C.; Masini, F.; Petruso, D. Depositi a vertebrati e ambienti costieri pleistocenici della Sicilia e della Calabria meridionale. Biogeogr. J. Integr. Biogeogr. 2001, 22, 29–43. [Google Scholar]
- Borgognini, S.M.; Elena, R. Dietary patterns in the Mesolithic samples from Uzzo and Molara caves (Sicily): The evidence of teeth. J. Hum. Evol. 1985, 14, 241–254. [Google Scholar]
- Borgognini, S.M.; Repetto, T.E. Skeletal indicators of subsistence patterns and activity régime in the Mesolithic sample from Grotta dell’Uzzo (Trapani, Sicily): A case study. Hum. Evol. 1986, 1, 331–351. [Google Scholar] [CrossRef]
- Borgognini, S.M.; Canci, A.; Piperno, M.; Repetto, E. Dati archeologici e antropologici sulle sepolture mesolitiche della Grotta dell’Uzzo (Trapani). In Bullettino di Paletnologia Italiana; Istituto Poligrafico e Zecca Dello Stato: Roma, Italy, 1993; Volume 84, pp. 85–179. [Google Scholar]
- Castellana, G.; Mallegni, F. The Prehistoric Settlement of Piano Vento in the Territory of Palma di Montechiaro (Agrigento, Italy). Arch. Per L’antropologia E La Etnol. 1986, 116, 61–80. [Google Scholar]
- Costantini, L. Plant exploitation at Grotta dell’Uzzo, Sicily: New evidence for the transition from Mesolithic to Neolithic subsistence in southern Europe. In Foraging and Farming; Routledge: London, UK, 2014; pp. 197–206. [Google Scholar]
- D’Amore, G.; Di Marco, S.; Tartarelli, G.; Bigazzi, R.; Sineo, L. Late Pleistocene human evolution in Sicily: Comparative morphometric analysis of Grotta di San Teodoro craniofacial remains. J. Hum. Evol. 2009, 56, 537–550. [Google Scholar] [CrossRef] [PubMed]
- D’Amore, G.; Di Marco, S.; Di Salvo, R.; Messina, A.; Sineo, L. Early human peopling of Sicily: Evidence from the Mesolithic skeletal remains from Grotta d’Oriente. In Annals of Human Biology; Taylor & Francis: London, UK, 2010; Volume 37. [Google Scholar]
- Di Salvo, R. Tre Resti Cranici da Marcita. Archivio Per L’Antropologia e la Etnologia; Società Italiana per l’Antropologia e la Etnologia: Firenze, Italy, 1991; Volume CXXII, pp. 251–258. [Google Scholar]
- Di Salvo, R.; Germanà, F.; Tusa, S. Uomini e Culture Della Sicilia Preistorica; Gaia Editrice: Milano, Italy, 1998. [Google Scholar]
- Di Salvo, R. I Musulmani della Sicilia occidentale: Aspetti antropologici e paleopatologici. In Mélanges de L’école Française de Rome; Moyen Âge (MEFRM): Roma, Italy, 2004; Volume 116. [Google Scholar]
- Di Salvo, R.; Schimmenti, V.; Messina, A. Nota paleobiologia degli inumati del cimitero sub divo si S. Giovanni – Marsala (Trapani-Sicilia) di età paleocristiana (III-IV sec. D.C.). In Archivio Per L’Antropologia e la Etnologia; Società Italiana per l’Antropologia e la Etnologia: Firenze, Italy, 2008; Volume CXXXVIII, pp. 111–121. [Google Scholar]
- Fiorentino, C.; Miccichè, R.M.; Caminneci, V.; Rizzo, M.S.; Di Giuseppe, Z.; Ficarra, S.; Sineo, L. Caratterizzazione antropologica e paleopatologica delle sepolture antistanti in Tempio della Concordia. In Archivio Per L’Antropologia e la Etnologia; Società Italiana per l’Antropologia e la Etnologia: Firenze, Italy, 2021; Volume CLI, pp. 139–164. [Google Scholar]
- Hodos, T. Globalization and Colonization: A View from Iron Age Sicily. J. Mediterr. Archaeol. 2010, 23, 81–106. [Google Scholar] [CrossRef]
- Incarbona, A.; Zarcone, G.; Agate, M.; Bonomo, S.; Stefano, E.; Masini, F.; Sineo, L. A multidisciplinary approach to reveal the Sicily Climate and Environment over the last 20,000 years. In Open Geosciences; De Gruyter: Berlin, Germany, 2010; Volume 2, pp. 71–82. [Google Scholar]
- Incarbona, A.; Agate, M.; Arisco, G.; Bonomo, S.; Buccheri, G.; Di Patti, C.; Di Stefano, E.; Greco, A.; Madonia, G.; Masini, F.; et al. Ambiente e clima della Sicilia durante gli ultimi 20 mila anni. Alp. Mediterr. Quat. 2010, 23, 21–36. [Google Scholar]
- Garilli, V.; Vita, G.; La Parola, V.; Vraca, M.P.; Giarrusso, R.; Rosina, P.; Sineo, L. First evidence of Pleistocene ochre production from bacteriogenic iron oxides. A case study of the Upper Palaeolithic site at the San Teodoro Cave (Sicily, Italy). J. Archaeol. Sci. 2020, 123, 105221. [Google Scholar] [CrossRef]
- Germanà, F.; Di Salvo, R. Dettagli di paleopatologia in un resto cranico punico dalla Caserma Tukory di Palermo. Arch. Per L’antropologia E La Etnol. 1994, 124, 107–119. [Google Scholar]
- Lauria, G.; Sconzo, P.; Falsone, G.; Sineo, L. Human Remains and Funerary Rites in the Phoenician Necropolis of Motya (Sicily). Int. J. Osteoarchaeol. 2017, 27, 1003–1011. [Google Scholar]
- Lauria, G.; Sineo, L.; Falsone, G.; Sconzo, P. New anthropological data from the archaic necropolis at Motya (2013 excavation season). In Proceedings of the 8th International Congress of Phoenician and Punic Studies, Carbonia, Italy, 21–26 October 2013; Folia Fenicia, Fabrizio Serra Publisher: Roma, Italy, 2018; pp. 250–252. [Google Scholar]
- Lauria, G.; Sconzo, P.; Falsone, G.; Sineo, L. Child inhumations on the island of Motya. New evidence from the archaic cemetery. In Un viaje entre el Oriente y el Occidente del Mediterráneo: A Journey between East and West in the Mediterranean, Proceedings of the 9th International Congress of Phoenician and Punic Studies, Merida, Spain, 22–26 october 2018; MYTRA: Merida, Spain, 2020; pp. 1837–1842. [Google Scholar]
- La Rocca, P. Variabilità Cranimetrica e Distanze Biologiche tra Popolazioni Preistoriche ed Antiche Della Sicilia. Ph.D. Thesis, Universita’degli Studi di Catania, Catania, Italy, 2011. [Google Scholar]
- Mannino, M.A.; Thomas, K.D.; Leng, M.J.; Piperno, M.; Tusa, S.; Tagliacozzo, A. Marine resources in the Mesolithic and Neolithic at the Grotta dell’Uzzo (Sicily): Evidence from isotope analyses of marine shells. In Archaeometry; John Wiley & Sons: Hoboken, NJ, USA, 2017; Volume 49, pp. 117–133. [Google Scholar]
- Messina, A.; Sineo, L.; Schimmenti, V.; Di Salvo, R. Cribra Orbitalia and Enamel Hypoplasia of the Iron Age (IX–VII centuries BC) Human Group of Polizzello (Sicily). J. Palaeopathol. 2008, 20, 53–65. [Google Scholar]
- Miccichè, R.; Carotenuto, G.; Sìneo, L. The utility of 3D medical imaging techniques for obtaining a reliable differential diagnosis of metastatic cancer in an Iron Age skull. Int. J. Paleopathol. 2018, 21, 41–46. [Google Scholar]
- Sineo, L.; Bigazzi, R.; D’Amore, G.; Tartarelli, G.; Di Patti, C.; Berzero, A.; Caramella Crespi, V. I resti umani della Grotta di S. Teodoro (Messina): Datazione assoluta con il metodo della spettrometria gamma diretta (U/Pa). In Antropo; University of the Basque Country: Bilbao, Spain, 2002; Volume 2, pp. 9–16. [Google Scholar]
- Sineo, L.; Petruso, D.; Forgia, V.; Messina, A.; D’Amore, G. Human peopling of Sicily during quaternary. In Quaternary Period; AcademyPublish.org: London, UK, 2015; pp. 25–67. [Google Scholar]
- Galland, M.; D’Amore, G.; Friess, M.; Miccichè, R.; Pinhasi, R.; Sparacello, V.S.; Sineo, L. Morphological variability of Upper Paleolithic and Mesolithic skulls from Sicily. J. Antropol. Sci. 2019, 97, 151–172. [Google Scholar]
- Bruner, E.; Manzi, G. Variability in facial size and shape among North and East African human populations. Ital. J. Zool. 2004, 71, 51–56. [Google Scholar] [CrossRef]
- Bruner, E. Cranial shape and size variation in human evolution: Structural and functional perspectives. In Child’s Nervous System; Springer: Boston, MA, USA, 2007; Volume 23, pp. 1357–1365. [Google Scholar]
- Von Cramon-Taubadel, N.; Weaver, T.D. Insights from a quantitative genetic approach to human morphological evolution. In Evolutionary Anthropology, Reviews; John Wiley & Sons: Hoboken, NJ, USA, 2009; Volume 18, pp. 237–240. [Google Scholar]
- Von Cramon-Taubadel, N. Evolutionary insights into global patterns of human cranial diversity: Population history, climatic and dietary effects. J. Anthropol. Sci. 2014, 92, 43–77. [Google Scholar] [PubMed]
- Matsumura, H.; Shinoda, K.I.; Shimanjuntak, T.; Oktaviana, A.A.; Noerwidi, S.; Sofian, H.O.; Adachi, N. Cranio-morphometric and aDNA corroboration of the Austronesian dispersal model in ancient Island Southeast Asia: Support from Gua Harimau, Indonesia. PLoS ONE 2018, 13, e0198689. [Google Scholar] [CrossRef] [PubMed]
- Buikstra, J.E.; Ubelaker, D.H. Standards for Data Collection from Human Skeletal Remains; Arkansas Archaeological Survey: Fayetteville, AR, USA, 1994; pp. 71–73. [Google Scholar]
- Ubelaker, D.H. Human skeletal remains. Excavation, analysis, interpretation. Am. J. Biol. Antropol. 1989, 32, 249–287. [Google Scholar]
- Lauria, G.; Sineo, L.; Ficarra, S. A detailed method for creating digital 3D models of human crania: An example of close-range photogrammetry based on the use of Structure-from-Motion (SfM) in virtual anthropology. In Archaeological and Anthropological Sciences; Springer: Boston, MA, USA, 2022; p. 14. [Google Scholar]
- Freidline, S.E.; Gunz, P.; Harvati, K.; Hublin, J.J. Middle Pleistocene human facial morphology in an evolutionary and developmental context. J. Hum. Evol. 2012, 63, 723–740. [Google Scholar] [CrossRef]
- Von Cramon-Taubadel, N.; Strauss, A.; Hubbe, M. Evolutionary population history of early Paleoamerican cranial morphology. Sci. Adv. 2017, 3, e1602289. [Google Scholar] [CrossRef] [PubMed]
- Bookstein, F.L. Combining the tools of geometric morphometrics. In Advances in Morphometrics; Springer: Boston, MA, USA, 1996; pp. 131–151. [Google Scholar]
- Slice, D.E. Modern morphometrics. In Modern Morphometrics in Physical Anthropology; Springer: Boston, MA, USA, 2005; pp. 1–45. [Google Scholar]
- Wiley, D.F.; Armenta, N.; Alcantara, D.A.; Ghosh, D.; Kil, Y.J.; Delson, E.; Harcour-Smith, W.H.; Rohlf, F.J.; Hamann, B.; St John, K. Landmark 3.6. In Institute of Data Analysis and Visualization (IDAV); University of California: Davis, CA, USA, 2007. [Google Scholar]
- Bookstein, F.L. Landmark methods for forms without landmarks: Morphometrics of group differences in outline shape. In Medical Image Analysis; Elsevier: Amsterdam, The Netherlands, 1991; Volume 1, pp. 225–243. [Google Scholar]
- Klingenberg, C.P. MorphoJ: An integrated Software Package for Geometric Morphometrics. Mol. Ecol. Resour. 2011, 11, 353–357. [Google Scholar]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. In Palaeontologia Electronica; Coquina Press: Minneapolis, MN, USA, 2001; Volume 4, p. 9. [Google Scholar]
- Dryden, I.L.; Mardia, K.V. Statistical Shape Analysis: With Applications in R; John Wiley & Sons: Hoboken, NJ, USA, 2016; Volume 995. [Google Scholar]
- Gower, J.C.; Payne, R.W. A comparison of different criteria for selecting binary tests in diagnostic keys. In Biometrika; Oxford University Press: Oxford, UK, 1975; Volume 62, pp. 665–672. [Google Scholar]
- Rohlf, F.J.; Slice, D. Extensions of the Procrustes method for the optimal superimposition of landmarks. In Systematic Biology; Oxford University Press: Oxford, UK, 1990; Volume 39, pp. 40–59. [Google Scholar]
- Goodall, C. Procrustes methods in the statistical analysis of shape. J. R. Stat. Soc. Ser. B Methodol. 1991, 53, 285–321. [Google Scholar] [CrossRef]
- Klingenberg, C.P. Cranial integration and modularity: Insights into evolution and development from morphometric data. In Hystrix Ital. J. Mammal.; La Sapienza University: Roma, Italy, 2013; Volume 24, 10. [Google Scholar]
- Hammer, Ø.; Harper, D.A. Paleontological data analysis; John Wiley & Sons: Hoboken, NJ, USA, 2008. [Google Scholar]
- Le Maître, A.; Mitteroecker, P. Multivariate comparison of variance in R. In Methods in Ecology and Evolution; British Ecological Society: London, UK, 2019; Volume 10, pp. 1380–1392. [Google Scholar]
- Bronstein, A.M.; Bronstein, M.M.; Kimmel, R. Generalized Multidimensional Scaling: A Framework for Isometry-Invariant Partial Surface Matching. Proc. Natl. Acad. Sci. USA 2006, 103, 1168–1172. [Google Scholar] [CrossRef]
- Rao, C.R. Linear Statistical Inference and its Applications, 2nd ed.; John Wiley & Sons: Hoboken, NJ, USA, 1973. [Google Scholar]
- Pillai, K.C.S. Upper percentage points of the largest root of a matrix in multivariate analysis. In Biometrika; Oxford University Press: Oxford, UK, 1967. [Google Scholar]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. In Molecular Biology and Evolution; Oxford University Press: Oxford, UK, 1987; Volume 4, pp. 406–425. [Google Scholar]
- D’Amore, G.; Di Marco, S.; Floris, G.; Pacciani, E.; Sanna, E. Craniofacial morphometric variation of the peopling of Sardinia. HOMO J. Comp. Hum. Biol. 2010, 61, 385–412. [Google Scholar]
- Reyes-Centeno, H.; Ghirotto, S.; Harvati, K. Genomic validation of the differential preservation of population history in modern human cranial anatomy. Am. J. Phys. Anthropol. 2017, 162, 170–179. [Google Scholar] [CrossRef] [PubMed]
- Von Cramon-Taubadel, N. The relative efficacy of functional and developmental cranial modules for reconstructing global human population history. Am. J. Phys. Anthropol. 2011, 146, 83–93. [Google Scholar] [CrossRef]
- Relethford, J.H.; Smith, F.H. Cranial measures and ancient DNA both show greater similarity of Neandertals to recent modern Eurasians than to recent modern sub-Saharan Africans. Am. J. Phys. Anthropol. 2018, 166, 170–178. [Google Scholar] [CrossRef] [PubMed]
- Sadori, L.; Zanchetta, G.; Giardini, M. Last Glacial to Holocene palaeoenvironmental evolution at Lago di Pergusa (Sicily, Southern Italy) as inferred by pollen, microcharcoal, and stable isotopes. Quat. Int. 2008, 181, 4–14. [Google Scholar] [CrossRef]
- Manica, A.; Amos, W.; Balloux, F.; Hanihara, T. The effect of ancient population bottlenecks on human phenotypic variation. Nature 2007, 448, 346–348. [Google Scholar] [CrossRef]
- Harvati, K.; Weaver, T.D. Human cranial anatomy and the differential preservation of population history and climate signatures. In The Anatomical Record Part A: Discoveries in Molecular, Cellular, and Evolutionary Biology: An Official Publication of the American Association of Anatomists; Wiley & Sons: Hoboken, NJ, USA, 2006; Volume 288, pp. 1225–1233. [Google Scholar]
- Smith, H.F. The Role of Genetic Drift in Shaping Modern Human Cranial Evolution: A Test Using Microevolutionary Modeling. J. Evol. Biol. 2011, 2011, 145262. [Google Scholar] [CrossRef] [PubMed]
- Betti, L.; Balloux, F.; Amos, W.; Hanihara, T.; Manica, A. Distance from Africa, Not Climate, Explains Within-Population Phenotypic Diversity in Humans. Proc. R. Soc. B Biol. Sci. 2009, 276, 809–814. [Google Scholar]









| Main Sicilian Prehistoric and Historical Periods: | |
|---|---|
| B.C.E. Before Common Era—C.E. Common Era | |
| Prehistory | |
| • Upper Palaeolithic: 38.000–8.000 | |
| • Mesolithic: 8.000–6.000 B.C.E. | |
| • Neolithic: 6.000–4.000 B.C.E. | |
| • Eneolithic/Copper Age: 4.000–2.500 B.C.E. | |
| • Bronze Age: 2.500–1.100 B.C.E. | |
| Early Bronze Age: 2.500–2.000 B.C.E. | |
| Middle Bronze Age: 2.000–1.500 B.C.E. | |
| Late Bronze Age: 1.500–1.100 B.C.E. | |
| • Iron Age: 1.100–700 B.C.E. | |
| History | |
| • Antiquity: 700 B.C.E.–100 C.E. | |
| Colonial Period: 700–600 B.C.E. | |
| Classical Period: 600–400 B.C.E. | |
| Hellenistic (Greek Period): 400–200 B.C.E. | |
| Roman Republic Period: 200 B.C.E.–100 C.E. | |
| • Late Antiquity (Roman Empire Period): 100–476 C.E. | |
| • Middle Ages: 476–1492 C.E. | |
| Byzantine Period: 476–1.000 C.E. | |
| Islamic Period: 1.000–1.300 C.E. | |
| Norman/Swabian Period. 1.300–1.492 C.E. | |
| • Modern Ages: 1.492–1.789 C.E. | |
| • Contemporary: 1.789 C.E. to Nowadays | |
| Site | Specimens | Dating | Periods |
|---|---|---|---|
| San Teodoro | 2 | 14.500 B.C.E. -14C | Upper Paleolithic |
| Uzzo | 2 | 9.000 B.C.E. | Mesolithic |
| Molara | 1 | 9.000 B.C.E. | Mesolithic |
| Marcita | 4 | 2.300–1.100 B.C.E. | Bronze |
| Polizzello | 2 | 900–800 B.C.E. | Iron |
| Baucina | 2 | 755–700 B.C.E. -14C | Antiquity |
| Mozia | 2 | 800–400 B.C.E. | Antiquity |
| Birgi | 5 | 700–100 B.C.E. | Antiquity |
| Tukory | 2 | 600–300 B.C.E. | Antiquity |
| Phoenician/Punic of PA | 4 | 600–300 B.C.E. | Antiquity |
| Lilibeo | 4 | 400.100 B.C.E. | Antiquity |
| Marsala | 6 | 200 C.E. | Antiquity |
| Lipari | 1 | 300–400 C.E. | Late Antiquity |
| Agrigento | 1 | 400–500 C.E. | Late Antiquity |
| C. San Pietro | 2 | 1.000–1.300 C.E. | Middle Ages |
| M. Iato | 3 | 1.000–1.300 C.E. | Middle Ages |
| Caltavuturo | 5 | 1.000–1.500 C.E. | Middle Ages |
| Alia | 45 | 1.800 C.E. | Contemporary |
| Rotoli | 3 | 2.000 C.E. | Contemporary |
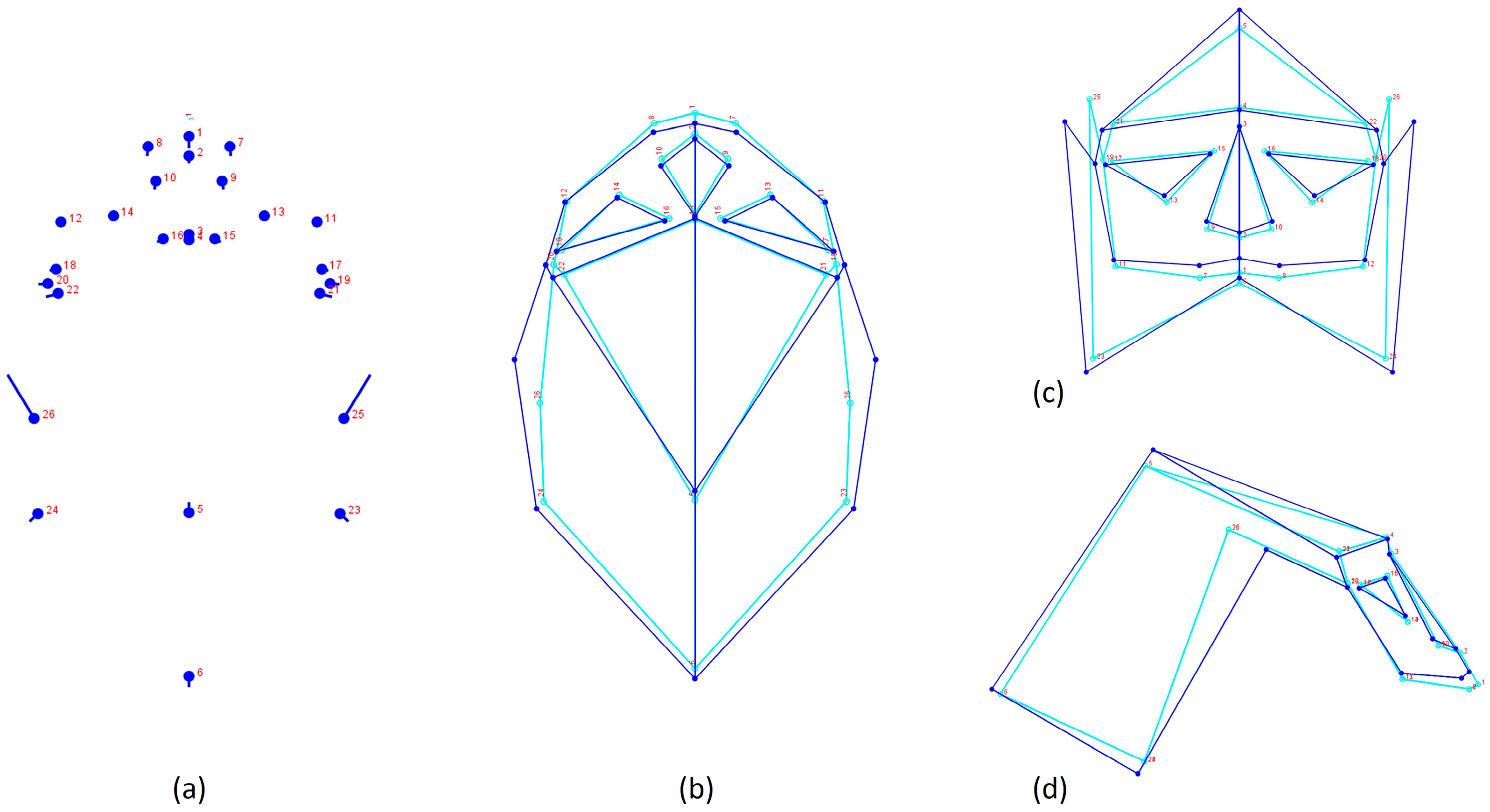
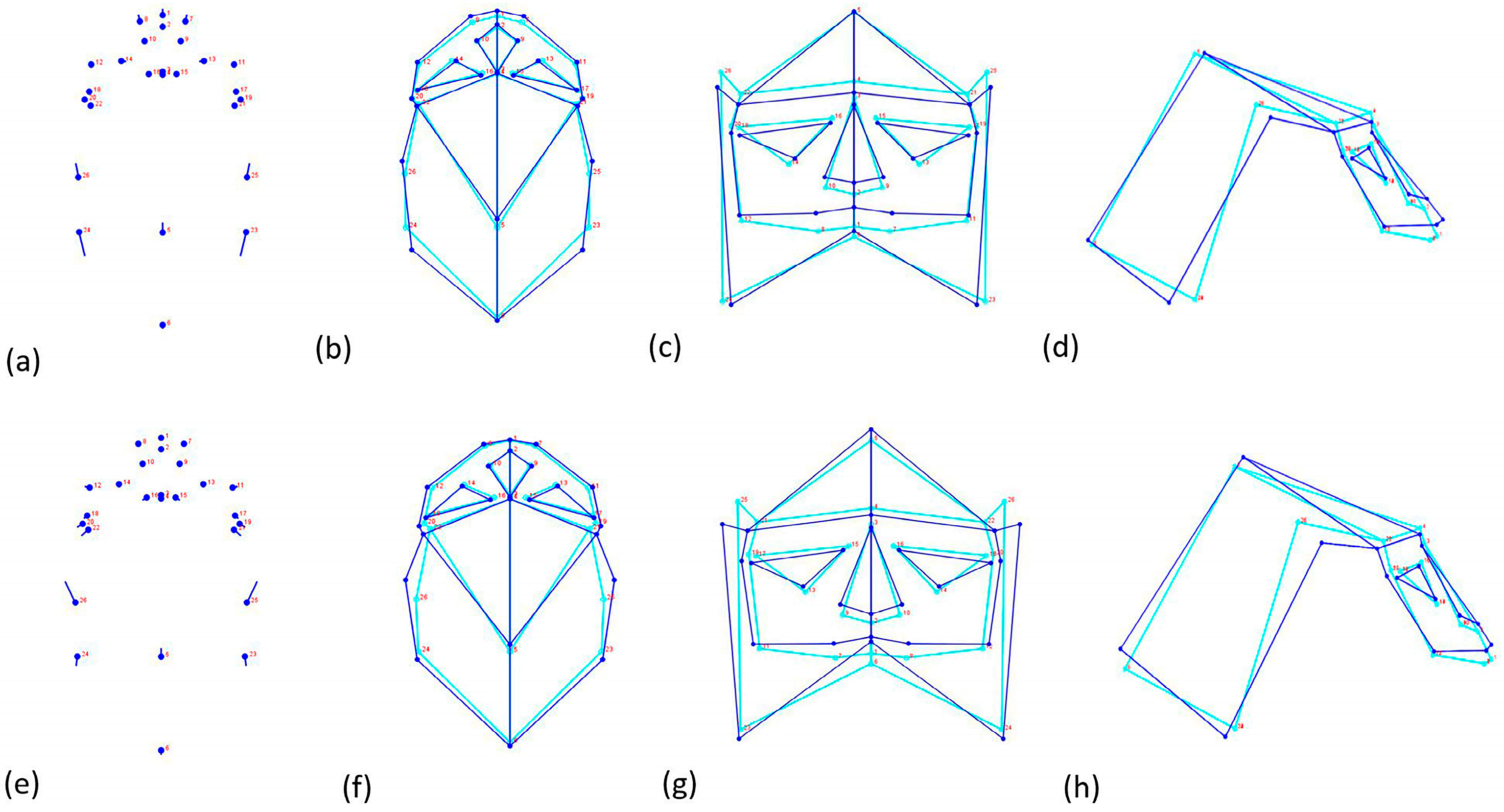
| MoprhpoJ | Landmark3.6 | Landmarks Configuration | Type |
|---|---|---|---|
| 1 | 0 | Prostion | 1 |
| 2 | 1 | Nasospinale | 1 |
| 3 | 2 | Nasion | 1 |
| 4 | 3 | Glabella | 2 |
| 5 | 4 | Bregma | 1 |
| 6 | 5 | Lambda | 1 |
| 6–8 | 6–7 | Point between the dental alveoli I2/C | 1 |
| 9–10 | 8–9 | Alare | 2 |
| 11–12 | 10–11 | Zygomatic-Maxillary suture—lower margin | 1 |
| 13–14 | 12–13 | Zygomatic-Maxillary suture–upper margin | 1 |
| 15–16 | 14–15 | Maxillary-Frontal suture | 1 |
| 17–18 | 16–17 | Ectoconchion | 1 |
| 19–20 | 18–19 | Fronto-Temporal-Malar | 1 |
| 21–22 | 20–21 | Frontotemporal | 1 |
| 23–24 | 22–23 | Occipital—Temporal—Parietal intersection | 1 |
| 25–26 | 24–25 | Stephanion | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lauria, G.; Sineo, L. Human Peopling and Population Dynamics in Sicily: Preliminary Analysis of the Craniofacial Morphometric Variation from the Paleolithic to the Contemporary Age. Heritage 2023, 6, 1187-1208. https://doi.org/10.3390/heritage6020066
Lauria G, Sineo L. Human Peopling and Population Dynamics in Sicily: Preliminary Analysis of the Craniofacial Morphometric Variation from the Paleolithic to the Contemporary Age. Heritage. 2023; 6(2):1187-1208. https://doi.org/10.3390/heritage6020066
Chicago/Turabian StyleLauria, Gabriele, and Luca Sineo. 2023. "Human Peopling and Population Dynamics in Sicily: Preliminary Analysis of the Craniofacial Morphometric Variation from the Paleolithic to the Contemporary Age" Heritage 6, no. 2: 1187-1208. https://doi.org/10.3390/heritage6020066
APA StyleLauria, G., & Sineo, L. (2023). Human Peopling and Population Dynamics in Sicily: Preliminary Analysis of the Craniofacial Morphometric Variation from the Paleolithic to the Contemporary Age. Heritage, 6(2), 1187-1208. https://doi.org/10.3390/heritage6020066