




Unraveling a Historical Mystery: Identification of a Lichen Dye Source in a Fifteenth Century Medieval Tapestry

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Julius Caesar (from the Heroes Tapestries), Accession Number 47.101.3
2.2. Materials
2.3. Reference Material
2.4. On-Fiber Dye Extraction Methodology
2.5. Preparation of Lecanora sulphurata for HPLC-ESI-qToF-MS Analysis
2.6. High-Performance Liquid Chromatography Electrospray Ionization, Quadrupole Time-of-Flight Mass Spectrometry (HPLC-ESI-qToF-MS)
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cardon, D. Natural Dyes: Sources, Tradition, Technology and Science; Archetype Publications: London, UK, 2007. [Google Scholar]
- Hofenk de Graaff, J.H.; Roelofs, W.G.; Van Bommel, M. The Colourful Past: Origins, Chemistry, and Identification of Natural Dyestuffs; Archetype Publications: London, UK, 2004. [Google Scholar]
- Perkins, P. Ecology, Beauty, Profits: Trade in Lichen-Based Dyestuffs through Western History. J. Soc. Dye. Colour. 1986, 102, 221–227. [Google Scholar] [CrossRef]
- Lindsay, W.L. Experiments on the Dyeing Properties of Lichens. Edinb. New Philos. J. 1854, 57, 228–249. [Google Scholar]
- Horwood, A.R. Lichen Dyeing To-Day: The Revival of an Ancient Industry. Sci. Prog. Twent. Century (1919–1933) 1928, 23, 279–283. [Google Scholar]
- Brough, S.G. Navajo Lichen Dyes. Lichenologist 1988, 20, 279–290. [Google Scholar] [CrossRef]
- Westring, J.P. Svenska Lafvarnas Färghistoria: Eller Sättet Att Använda Dem till Färgning Och Annan Hushållsnytta; Tryckt hos C. Delén: Stockholm, Sweden, 1805. [Google Scholar]
- Casselman, K.D. Lichen Dyes: The New Source Book, 2nd ed.; Dover Publications, Inc.: Mineola, NY, USA, 2001. [Google Scholar]
- Dalip, K.U.; Divakar, P.K.; Nayaka, S. Commercial and Ethnic Use of Lichens in India. Econ. Bot. 2005, 59, 269–273. [Google Scholar]
- Michael, M.E. Contentious Cures: The Rise and Decline of Lichens as European Materia Medica. Pharm. Hist. 2015, 57, 55–66. [Google Scholar] [CrossRef]
- Lawrey, J.D. Biological Role of Lichen Substances. Bryologist 1986, 89, 111–122. [Google Scholar] [CrossRef]
- Molnár, K.; Farkas, E. Current Results on Biological Activities of Lichen Secondary Metabolites: A Review. Z. Naturforschung C 2010, 65, 157–173. [Google Scholar] [CrossRef] [PubMed]
- Olivier-Jimenez, D.; Chollet-Krugler, M.; Rondeau, D.; Beniddir, M.A.; Ferron, S.; Delhaye, T.; Allard, P.-M.; Wolfender, J.-L.; Sipman, H.J.M.; Lücking, R.; et al. A Database of High-Resolution MS/MS Spectra for Lichen Metabolites. Sci. Data 2019, 6, 294. [Google Scholar] [CrossRef]
- Elix, J.A. A Catalogue of Standardized Chromatographic Data and Biosynthetic Relationships for Lichen Substances, 4th ed.; The Author: Canberra, Australia, 2018. [Google Scholar]
- Kranner, I.; Beckett, R.; Hochman, A.; Nash, T.H. Desiccation-Tolerance in Lichens: A Review. Bryologist 2008, 111, 576–593. [Google Scholar]
- Nguyen, K.-H.; Chollet-Krugler, M.; Gouault, N.; Tomasi, S. UV-Protectant Metabolites from Lichens and Their Symbiotic Partners. Nat. Prod. Rep. 2013, 30, 1490–1508. [Google Scholar] [CrossRef] [PubMed]
- Casselman, K.L. Lichen Dyes: Preparation and Dyeing. Maine Nat. 1994, 2, 105–110. [Google Scholar] [CrossRef]
- Rosi, F.; Clementi, C.; Paolantoni, M.; Romani, A.; Pellegrino, R.; Brunetti, B.G.; Nowik, W.; Miliani, C. Study of Raman Scattering and Luminescence Properties of Orchil Dye for Its Nondestructive Identification on Artworks. J. Raman Spectrosc. 2013, 44, 1451–1456. [Google Scholar] [CrossRef]
- Doherty, B.; Gabrieli, F.; Clementi, C.; Cardon, D.; Sgamellotti, A.; Brunetti, B.; Miliani, C. Surface Enhanced Raman Spectroscopic Investigation of Orchil Dyed Wool from Roccella tinctoria and Lasallia pustulata. J. Raman Spectrosc. 2014, 45, 723–729. [Google Scholar] [CrossRef]
- Clementi, C.; Miliani, C.; Romani, A.; Favaro, G. In Situ Fluorimetry: A Powerful Non-Invasive Diagnostic Technique for Natural Dyes Used in Artefacts: Part I. Spectral Characterization of Orcein in Solution, on Silk and Wool Laboratory-Standards and a Fragment of Renaissance Tapestry. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2006, 64, 906–912. [Google Scholar] [CrossRef] [PubMed]
- Aceto, M.; Arrais, A.; Marsano, F.; Agostino, A.; Fenoglio, G.; Idone, A.; Gulmini, M. A Diagnostic Study on Folium and Orchil Dyes with Non-Invasive and Micro-Destructive Methods. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2015, 142, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Witkowski, B.; Ganeczko, M.; Hryszko, H.; Stachurska, M.; Gierczak, T.; Biesaga, M. Identification of Orcein and Selected Natural Dyes in 14th and 15th Century Liturgical Paraments with High-Performance Liquid Chromatography Coupled to the Electrospray Ionization Tandem Mass Spectrometry (HPLC-ESI/MS/MS). Microchem. J. 2017, 133, 370–379. [Google Scholar] [CrossRef]
- Calà, E.; Benzi, M.; Gosetti, F.; Zanin, A.; Gulmini, M.; Idone, A.; Serafini, I.; Ciccola, A.; Curini, R.; Whitworth, I.; et al. Towards the Identification of the Lichen Species in Historical Orchil Dyes by HPLC-MS/MS. Microchem. J. 2019, 150, 104140. [Google Scholar] [CrossRef]
- Chen, V.J.; Smith, G.D.; Holden, A.; Arft, S. Dye Analysis of a 17th-Century Mattens Workshop Tapestry Based on Raphael’s Drawing, The Miraculous Draught of Fishes. Heritage 2024, 7, 1221–1236. [Google Scholar] [CrossRef]
- Rorimer, J.J.; Freeman, M.B. The Nine Heroes Tapestries at the Cloisters; Metropolitan Museum of Art: New York, NY, USA, 1960. [Google Scholar]
- Carò, F.; Chiostrini, G.; Cleland, E.; Shibayama, N. Redeeming Pieter Coecke van Aelst’s Gluttony Tapestry: Learning from Scientific Analysis. Metrop. Mus. J. 2014, 49, 151–164. [Google Scholar] [CrossRef]
- Vlachou-Mogire, C.; Danskin, J.; Gilchrist, J.R.; Hallett, K. Mapping Materials and Dyes on Historic Tapestries Using Hyperspectral Imaging. Heritage 2023, 6, 3159–3182. [Google Scholar] [CrossRef]
- Claisse, P.; de La Codre, H.; Nowik, W.; Daniel, F.; Chapoulie, R.; Dallel, M.; Mounier, A. Original and Restored Materials Revealed by Contactless and Micro-Invasive Methods of the Lady and the Unicorn Tapestry “Mon Seul Désir”. Eur. Phys. J. Plus 2023, 138, 832. [Google Scholar] [CrossRef]
- Chavanne, C.; Verney, A.; Paquier-Berthelot, C.; Bostal, M.; Buléon, P.; Walter, P. Bayeux Tapestry: First Use of Early Synthetic Dyes for the Restoration of a Masterpiece. Dye Pigment. 2023, 208, 110798. [Google Scholar] [CrossRef]
- Delaney, J.K.; Ricciardi, P.; Glinsman, L.; Palmer, M.; Burke, J. Use of near Infrared Reflectance Imaging Spectroscopy to Map Wool and Silk Fibres in Historic Tapestries. Anal. Methods 2016, 8, 7886–7890. [Google Scholar] [CrossRef]
- de La Codre, H.; Marembert, C.; Claisse, P.; Daniel, F.; Chapoulie, R.; Servant, L.; Mounier, A. Non-Invasive Characterization of Yellow Dyes in Tapestries of the 18th Century: Influence of Composition on Degradation. Color Res. Appl. 2021, 46, 613–622. [Google Scholar] [CrossRef]
- Colburn, K. Three Fragments of the “Mystic Capture of the Unicorn” Tapestry. Metrop. Mus. J. 2010, 45, 97–106. [Google Scholar]
- Lackner, R.M.; Fozi, S.; Suh, K. Julius Caesar from the Heroes Tapestries at The Met Cloisters: Dye Analysis and Molecular Insights. Metrop. Mus. J. 2024. forthcoming. [Google Scholar]
- Hyde, A. The Heroines of Heroes; Met Perspectives: New York, NY, USA, 2022. [Google Scholar]
- Mouri, C.; Laursen, R. Identification and Partial Characterization of C-Glycosylflavone Markers in Asian Plant Dyes Using Liquid Chromatography–Tandem Mass Spectrometry. J. Chromatogr. A 2011, 1218, 7325–7330. [Google Scholar] [CrossRef]
- Gribble, G.W. Naturally Occurring Organohalogen Compounds. Acc. Chem. Res. 1998, 31, 141–152. [Google Scholar] [CrossRef]
- Neumann, C.S.; Fujimori, D.G.; Walsh, C.T. Halogenation Strategies In Natural Product Biosynthesis. Chem. Biol. 2008, 15, 99–109. [Google Scholar] [CrossRef]
- Karapanagiotis, I. A Review on the Archaeological Chemistry of Shellfish Purple. Sustainability 2019, 11, 3595. [Google Scholar] [CrossRef]
- Elix, J.A.; Crook, C.E. The Joint Occurrence of Chloroxanthones in Lichens, and a Further Thirteen New Lichen Xanthones. Bryologist 1992, 95, 52–64. [Google Scholar] [CrossRef]
- Elix, J.A.; Chappell, H.-M.; Jiang, H. Four New Lichen Xanthones. Bryologist 1991, 94, 304–307. [Google Scholar] [CrossRef]
- Dayan, F.E.; Romagni, J.G. Lichens as a Potential Source of Pesticides. Pestic. Outlook 2001, 12, 229–232. [Google Scholar] [CrossRef]
- El-Seedi, R.H.; El-Barbary, A.M.; El-Ghorab, M.H.D.; Bohlin, L.; Borg-Karlson, A.-K.; Goransson, U.; Verpoorte, R. Recent Insights into the Biosynthesis and Biological Activities of Natural Xanthones. Curr. Med. Chem. 2010, 17, 854–901. [Google Scholar] [CrossRef] [PubMed]
- Cacho, R.A.; Chooi, Y.-H.; Zhou, H.; Tang, Y. Complexity Generation in Fungal Polyketide Biosynthesis: A Spirocycle-Forming P450 in the Concise Pathway to the Antifungal Drug Griseofulvin. ACS Chem. Biol. 2013, 8, 2322–2330. [Google Scholar] [CrossRef] [PubMed]
- Le Pogam, P.; Boustie, J. Xanthones of Lichen Source: A 2016 Update. Molecules 2016, 21, 294. [Google Scholar] [CrossRef] [PubMed]
- Dembitsky, V.M.; Tolstikov, G.A. Natural Chlorine-Containing Xanthones. Chem. Sustain. Dev. 2004, 12, 13–18. [Google Scholar]
- Huneck, S.; Yoshimura, I. Identification of Lichen Substances. In Identification of Lichen Substances; Huneck, S., Yoshimura, I., Eds.; Springer: Berlin/Heidelberg, Germany, 1996; pp. 11–123. ISBN 978-3-642-85243-5. [Google Scholar]
- Culberson, C.F.; Culberson, W.L. Chemosyndromic Variation in Lichens. Syst. Bot. 1976, 1, 325–339. [Google Scholar] [CrossRef]
- Egan, R.S. Correlations and Non-Correlations of Chemical Variation Patterns with Lichen Morphology and Geography. Bryologist 1986, 89, 99–110. [Google Scholar] [CrossRef]
- Schweiger, A.H.; Ullmann, G.M.; Nürk, N.M.; Triebel, D.; Schobert, R.; Rambold, G. Chemical Properties of Key Metabolites Determine the Global Distribution of Lichens. Ecol. Lett. 2022, 25, 416–426. [Google Scholar] [CrossRef] [PubMed]
- Mairet, E. A Book on Vegetable Dyes; St. Dominic’s Press: Ditchling, UK, 1916. [Google Scholar]
- Bolton, E.M.; Casselman, K.L.; Holloway, J.B. Lichens for Vegetable Dyeing; Robin & Russ Handweavers: Santa Barbara, CA, USA, 1991; ISBN 978-1-56659-001-3. [Google Scholar]
- Smith, C.W. The Lichens of Great Britain and Ireland; British Lichen Society: London, UK, 2009; ISBN 978-0-9540418-8-5. [Google Scholar]
- Lumbsch, H.T.; Feige, G.B. Comments on the Exsiccat “Lecanoroid Lichens” I. Mycotaxon 1992, 45, 473–488. [Google Scholar]
- Consortium of Lichen Herbaria. Available online: https://lichenportal.org/portal/ (accessed on 1 April 2024).
- Leuckert, C.; Poelt, J. Studien Über Die Lecanora rupicola-Gruppe in Europa (Lecanoraceae). Nova Hedwig. 1989, 49, 121–167. [Google Scholar]
- Tatipamula, V.; Vedula, G.; Sastry, A.V.S. Antarvedisides A-B from Manglicolous Lichen Dirinaria consimilis (Stirton) and Their Pharmacological Profile. Asian J. Chem. 2019, 31, 805–812. [Google Scholar] [CrossRef]
- Grube, M.; Baloch, E.; Arup, U. A Phylogenetic Study of the Lecanora rupicola Group (Lecanoraceae, Ascomycota). Mycol. Res. 2004, 108, 506–514. [Google Scholar] [CrossRef] [PubMed]
- Begora, M.; Fahselt, D. Photolability of Secondary Compounds in Some Lichen Species. Symbiosis 2001, 31, 3–22. [Google Scholar]
- Oborn, E.T. Iron Content of Selected Water and Land Plants; Water Supply Paper; U.S. Government Printing Office: Washington, DC, USA, 1960.
- Fry, E.J. A Suggested Explanation of the Mechanical Action of Lithophytic Lichens on Rocks (Shale). Ann. Bot. 1924, os-38, 175–196. [Google Scholar] [CrossRef]
- Williams, M.E.; Rudolph, E.D. The Role of Lichens and Associated Fungi in the Chemical Weathering of Rock. Mycologia 1974, 66, 648–660. [Google Scholar] [CrossRef]
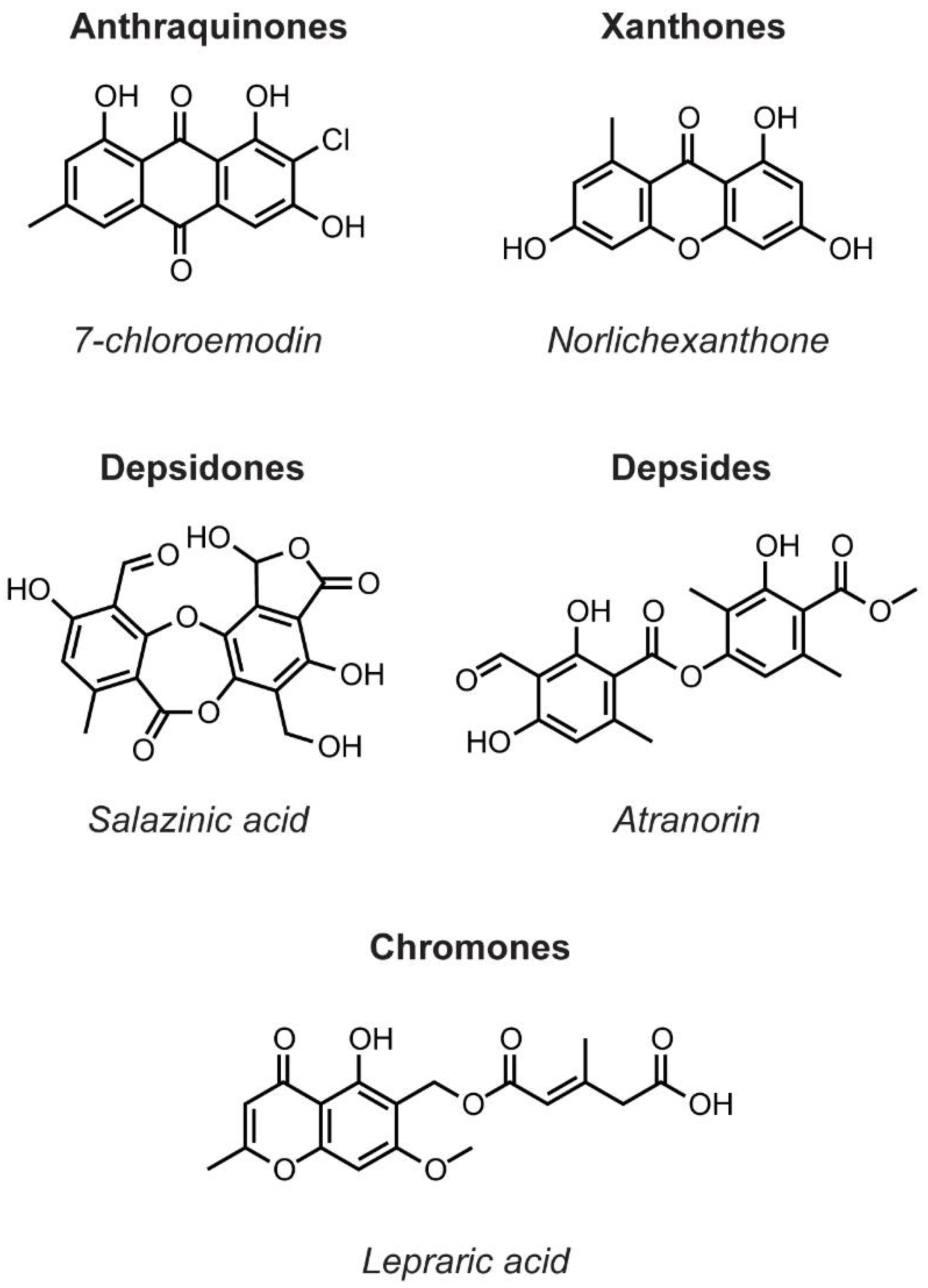


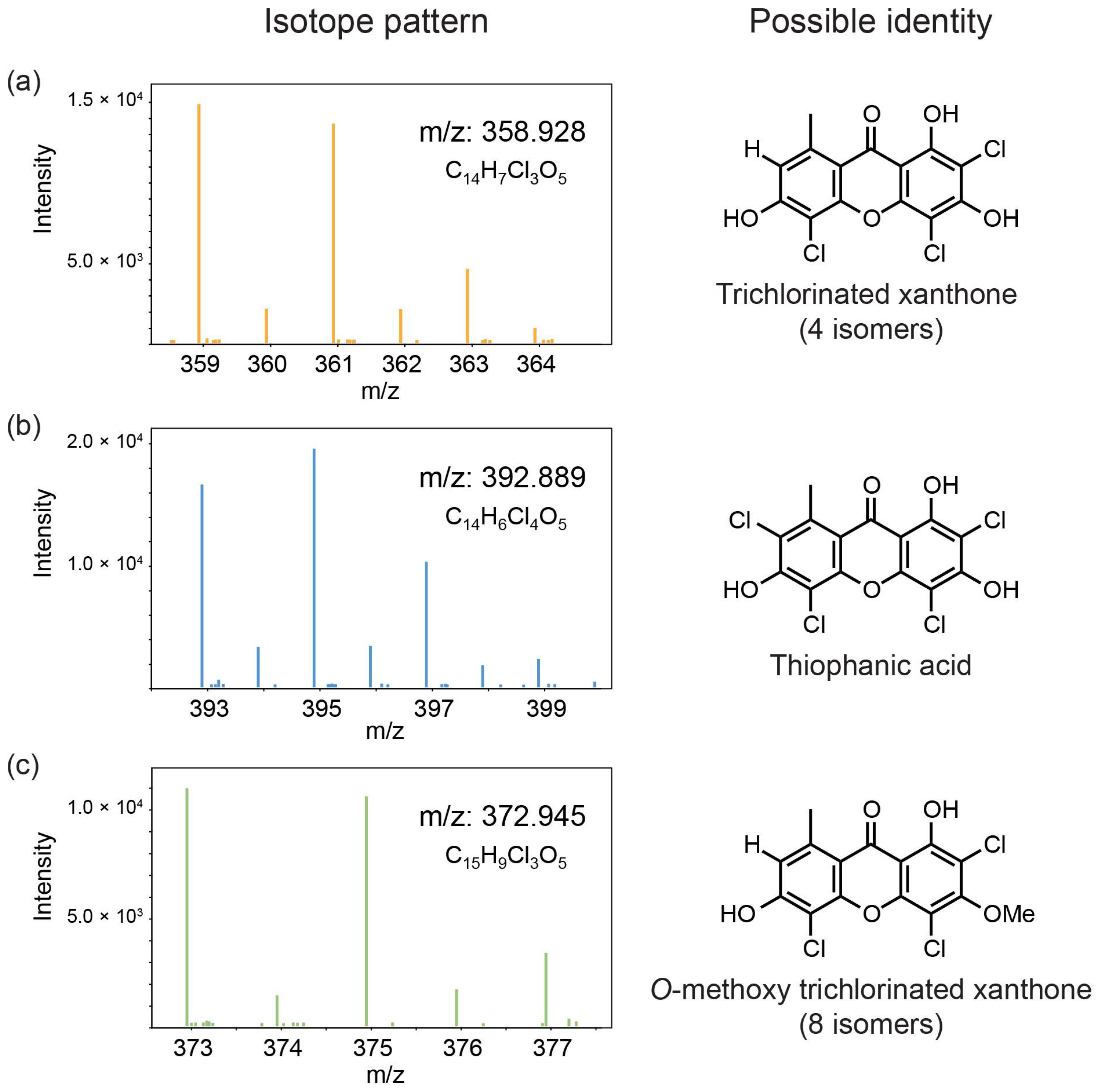
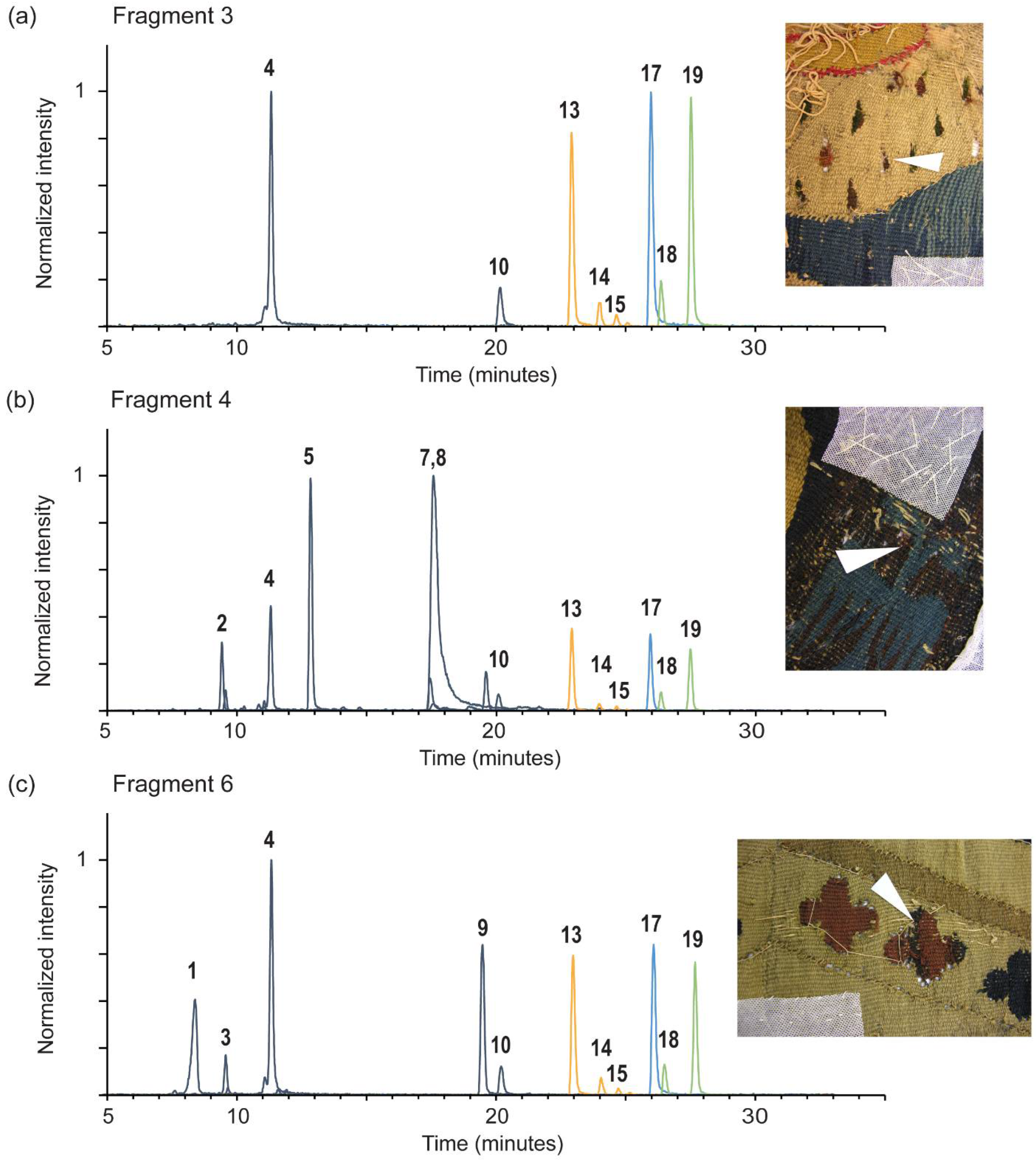
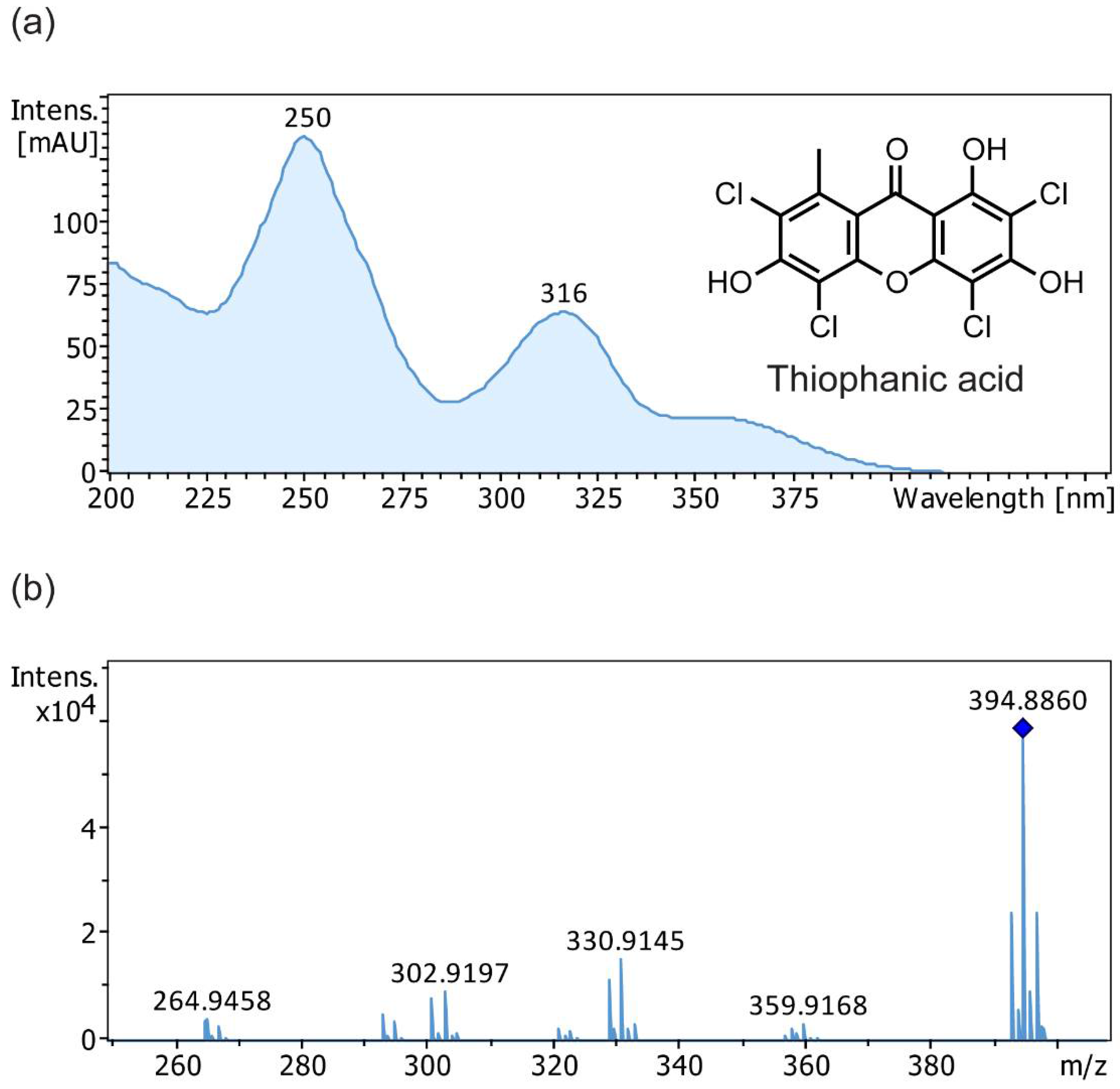

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
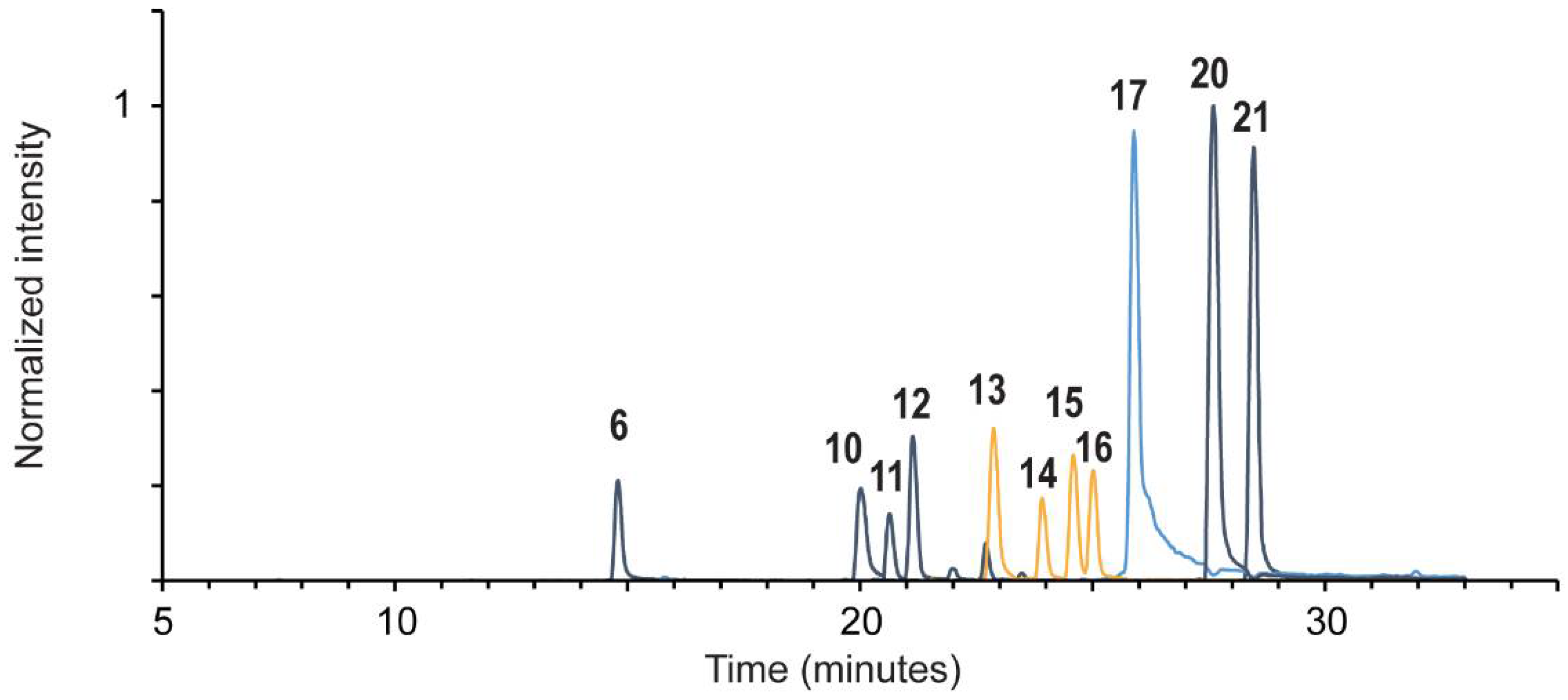
| # | Compound Name | RT (min) | [M − H]−, m/z | Product ions, m/z | λmax, nm a | Frag. 3 | Frag. 4 | Frag. 6 | Lecanora sulphurata Reference |
|---|---|---|---|---|---|---|---|---|---|
| 1 | isatin | 8.4 | 146.024 | 118 | - | - | - | + | - |
| 2 | luteolin 7-O-glucoside | 9.4 | 447.093 | 285 | 197, 253, 346 | - | + | - | - |
| 3 | ellagic acid | 9.6 | 300.999 | 284, 257, 245, 229, 173, 145 | 252, 366 | - | - | + | - |
| 4 | O-methyl ellagic acid | 11.3 | 315.015 | 300 | - | + | + | + | - |
| 5 | luteolin | 12.7 | 285.04 | 241, 199, 175, 151, 133, 107, 83 | 204, 253, 346 | - | + | - | - |
| 6 | C14H4Cl4O7 | 14.8 | 422.863 | 381, 346, 318, 307, 289, 279, 251, 224 | 198, 249, 320, 366 | - | - | - | + |
| 7 | alizarin | 17.5 | 239.035 | 211, 195, 183, 167 | 247, 428 | - | + | - | - |
| 8 | munjistin | 17.6 | 283.025 | 239, 211, 195, 167 | 209, 246, 288, 430 | - | + | - | - |
| 9 | indigotin b | 19.5 | 261.067 | 260, 233, 217, 156 | 200, 239, 284, 609 | - | - | t | - |
| 10 | C14H6Cl4O6 | 20.0 | 408.885 | 393, 381, 357, 329 | 211, 246, 317, 355 | + | + | + | + |
| 11 | 4,5-dichloronolichexanthone | 20.6 | 324.967 | 290, 261, 233, 183 | 211, 246, 317, ~366 | - | - | - | + |
| 12 | 2,5-dichloronorlichexanthone | 21.1 | 324.967 | 290, 261, 233, 183 | 197, 247, 316 | - | - | - | + |
| 13 | 2,4,5-trichloronorlichexanthone (arthothelin) | 22.7 | 358.929 | 323, 295, 267, 259, 231 | 212, 249, 315 | + | + | + | + |
| 14 | 4,5,7-trichloronorlichexanthone (asemone) | 23.8 | 358.929 | 324, 295, 267.0 | 248, 314, 352 (sh) | t | t | + | + |
| 15 | 2,5,7-trichloronorlichexanthone (isoarthothelin) | 24.5 | 358.929 | 324, 295, 267, 181, 163.0 137.1 | 201, 250, 315, 350 (sh) | t | t | t | + |
| 16 | 2,4,7-trichloronorlichexanthone | 25.0 | 358.929 | 323, 295, 287, 267, 259, 231, 177, 149 | 201, 247, 314, 350 (sh) | - | - | - | + |
| 17 | 2,4,5,7-tetrachloronorlichexanthone (thiophanic acid) | 25.8 | 392.89 | 360, 329, 321, 301, 293, 264 303, 265, 149 | 200, 250, 316, 355 (sh) | + | + | + | + |
| 18 | C15H9Cl3O5 A | 26.4 | 372.944 | 358, 330, 294, 266, 97 | - | + | + | + | - |
| 19 | C15H9Cl3O5 B | 27.5 | 372.944 | 358, 340, 294, 266 | - | + | + | + | - |
| 20 | atranorin | 27.5 | 373.093 | 177, 163, 133, 119, 105 | 208, 250, 319 | - | - | - | + |
| 21 | chloroatranorin | 28.5 | 407.054 | 211, 167, 163, 139, 131, 119 | 211, 248, ~350 | - | - | - | + |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lackner, R.M.; Ferron, S.; Boustie, J.; Le Devehat, F.; Lumbsch, H.T.; Shibayama, N. Unraveling a Historical Mystery: Identification of a Lichen Dye Source in a Fifteenth Century Medieval Tapestry. Heritage 2024, 7, 2370-2384. https://doi.org/10.3390/heritage7050112
Lackner RM, Ferron S, Boustie J, Le Devehat F, Lumbsch HT, Shibayama N. Unraveling a Historical Mystery: Identification of a Lichen Dye Source in a Fifteenth Century Medieval Tapestry. Heritage. 2024; 7(5):2370-2384. https://doi.org/10.3390/heritage7050112
Chicago/Turabian StyleLackner, Rachel M., Solenn Ferron, Joël Boustie, Françoise Le Devehat, H. Thorsten Lumbsch, and Nobuko Shibayama. 2024. "Unraveling a Historical Mystery: Identification of a Lichen Dye Source in a Fifteenth Century Medieval Tapestry" Heritage 7, no. 5: 2370-2384. https://doi.org/10.3390/heritage7050112
APA StyleLackner, R. M., Ferron, S., Boustie, J., Le Devehat, F., Lumbsch, H. T., & Shibayama, N. (2024). Unraveling a Historical Mystery: Identification of a Lichen Dye Source in a Fifteenth Century Medieval Tapestry. Heritage, 7(5), 2370-2384. https://doi.org/10.3390/heritage7050112