


Cnida Morphology as Taxonomic Tools within Tube-Dwelling Anemones (Ceriantharia, Cnidaria)



Abstract
:1. Introduction
2. Materials and Methods
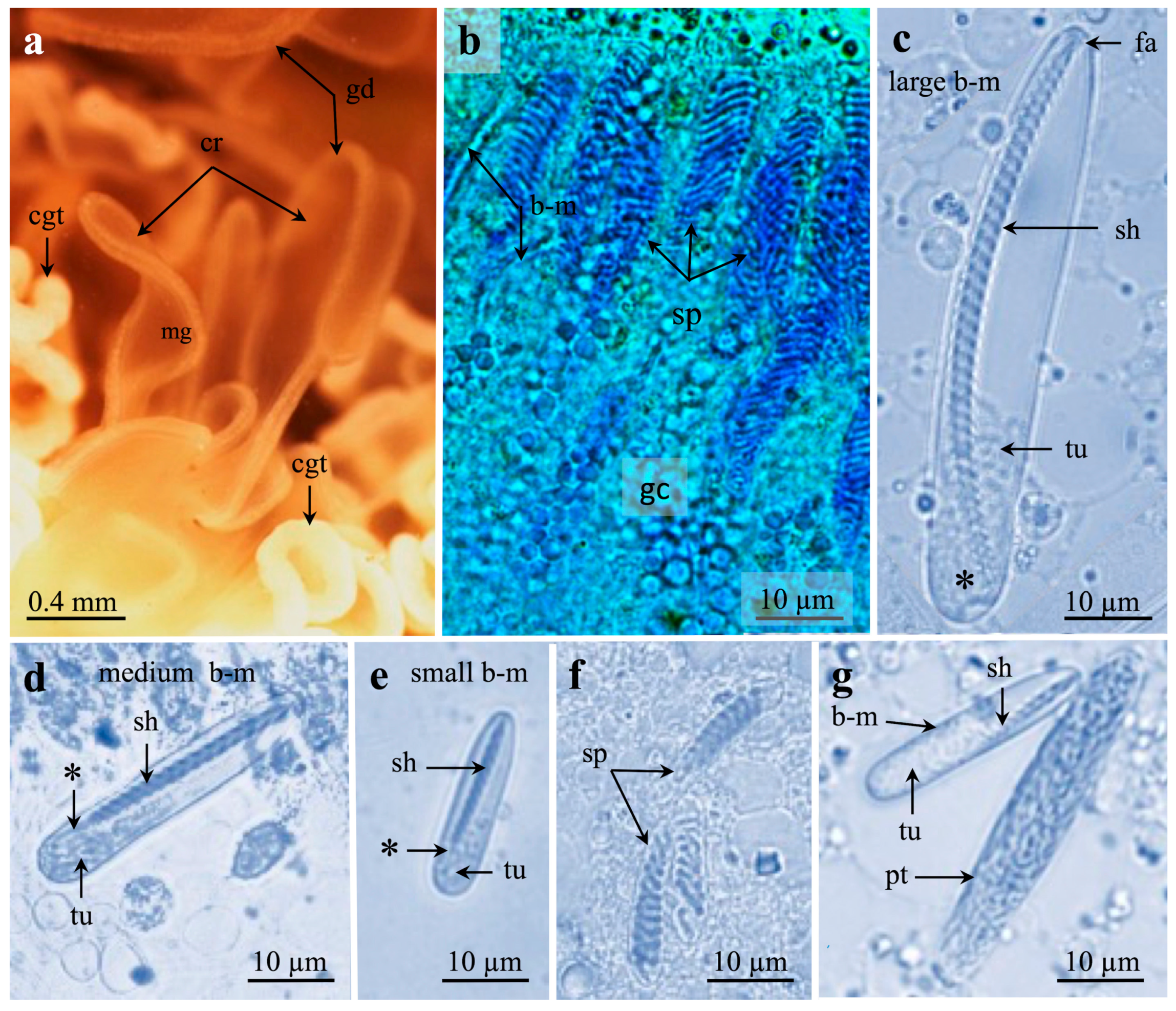
2.1. Investigated Species and Tissue Preparations
2.2. Light Microscopic (LM) Examination
2.3. Scanning Electron Microscopic (SEM) Examination
2.4. Transmission Electron Microscopic (TEM) Examination
2.5. Cnida Types among Anthozoa
3. Results
3.1. Cnida Types and Distribution
3.1.1. b-Mastigophores
3.1.2. Isorhizas
3.1.3. Spirocysts
3.1.4. Ptychocysts
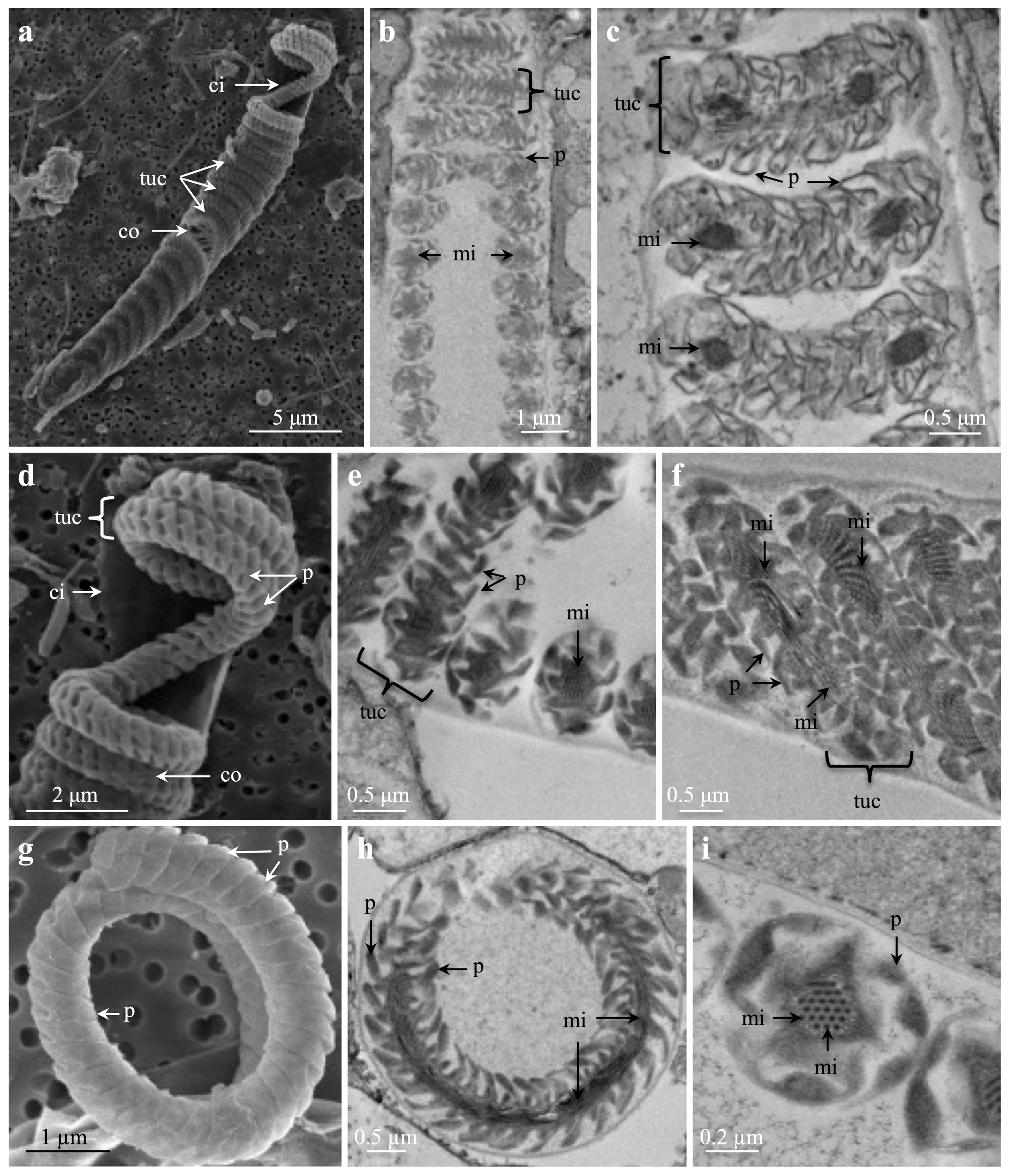
3.2. Specific Cnida Structures
3.3. Cnida Distribution and Comparison between the Recorded Measurments
3.3.1. Spirocysts
3.3.2. b-Mastigophores
3.3.3. Ptychocysts
3.3.4. Isorhizas
4. Discussion
4.1. Cnida Classification and Terminology
4.2. Cnida Subtypes, Their Variations, and Taxonomic Value
4.2.1. b-Mastigophores
4.2.2. Isorhizas
4.2.3. Spirocysts
4.2.4. Ptychocysts
4.3. Specific Cnida Structures
4.4. Spirularia–Spirula and Penicillaria–Penicillus, Terms to Be Retained or Deleted within the Ceriantharia Classification
4.5. Cnida Diversity and Evolution
5. Conclusions
- The cnidae b-mastigophores, spirocysts, and ptychocysts are placed into different morphological subtypes and size groups, some of which are species-specific or specific to certain structures. Other subtypes are common in most structures and are of less taxonomic value. For species determination, detailed cnida surveys within Ceriantharia are valuable;
- The different patterns of the “faltstück” within b-mastigophores may be species-specific features;
- To highlight the similarities and differences between spirulae and penicilli [18] and hexacorallian b-mastigophores and p-mastigophores/p-amastigophores, we propose that spirulae and penicilli can be referred to as b-mastigophores/spirulae subtype and b-mastigophores/penicillin subtype, respectively, until the terms spirulae/Spirularia and penicilli/Penicillaria have been changed. To verify the validity of the penicilli-v-notch, cnida investigations of more Arachnactidae species are desirable;
- The following question is raised: Can the minor morphological differences between spirulae and penicilli nematocysts justify the names Spirularia and Penicillaria of the suborders?
- The occurrence/absence of the less common medium to very large isorhizas are valuable species characteristics;
- The pattern of the ptychocyst tubule is unique. Its function is tube-building. The inverted longitudinally and transversely folded tubules differ from the helically twisted and coiled nematocyst and spirocyst tubules;
- The occurrence of the regular striation pattern on the inner ptychocyst capsule wall in certain Ceriantharia species is a valuable species characteristic;
- Cnidomes combined with morphology and molecular data can be useful for phylogenetic analysis;
- The occurrence of an operculum, tripartite opercular flaps, or apical capsular cap shows possible evolutionary relationships and development within the Cnidaria.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Forero Mejia, A.; Molodtsova, T.; Östman, C.S.; Bavestrello, G.; Rouse, G.W. Molecular phylogeny of Ceriantharia (Anthozoa, Cnidaria) reveals non-monophyly of traditionally accepted families. Zool. J. Linn. Soc. 2020, 190, 397–416. [Google Scholar] [CrossRef]
- Quattrini, A.M.; Snyder, K.E.; Purow-Ruderman, R.; Seiblitz, I.G.L.; Hoang, J.; Floerke, N.; Ramos, N.I.; Wirshing, H.H.; Rodriguez, E.; McFadden, C.S. Mito-nuclear discordance within Anthozoa, with notes on unique properties of their mitochondrial genomes. Sci. Rep. 2023, 13, 7443. [Google Scholar] [CrossRef] [PubMed]
- DeLeo, D.M.; Bessho-Uehara, M.; Haddock, S.H.D.; McFadden, C.S.; Quattrini, A.M. Evolution of bioluminescence in Anthozoa with emphasis on Octocorallia. Proc. R. Soc. B 2024, 291, 20232626. [Google Scholar] [CrossRef] [PubMed]
- DeBiasse, M.B.; Buckenmeyer, A.; Macrander, J.; Babonis, L.S.; Bentlage, B.; Cartwright, P.; Prada, C.; Reitzel, A.M.; Stampar, S.N.; Collins, A.G.; et al. A cnidarian phylogenomic tree fitted with hundreds of 18S leaves. Bull. Soc. Syst. Biol. 2024, 3, 2. [Google Scholar] [CrossRef]
- Molodtsova, T. Ceriantharia (Cnidaria) of the Gulf of Mexico. In Gulf of Mexico Origin, Waters, and Biota. Volume 1: Biodiversity; Felder, D.L., Camp, D.K., Eds.; Texas A&M University Press: College Station, TX, USA, 2009; pp. 365–367. [Google Scholar]
- Stampar, S.N.; Beneti, J.S.; Acuña, F.H.; Morandini, A.C. Ultrastructure and tube formation in Ceriantharia (Cnidaria, Anthozoa). Zool. Anz. 2015, 254, 67–71. [Google Scholar] [CrossRef]
- Mariscal, R.N.; Conklin, B.J.; Bigger, C.H. The ptychocyst, a major new category of cnida used in tube constraction by cerianthid anemone. Biol. Bull. 1977, 152, 392–405. [Google Scholar] [CrossRef]
- Calgren, O. A contribution to the knowledge of structure and distribution of cnidae in the Anthozoa. Acta Univ. Lundensis 1940, 36, 1–62. [Google Scholar]
- Weill, R. Contribution a l’étude des Cnidaires et leur nématocystes. I. Recherches sur les nématocystes. II. Valeur taxonomique du cnidom. In Travaux de la Station Zoologique de Wimereux; Biodiversity Heritage Library: Wimereux, France, 1934; Volume 10, pp. 1–701. [Google Scholar]
- Schmidt, H. Die Nesslkapseln der Aktinien und ihre differentialdiagnostische Bedeutung. Helgoländer Wiss. Meeresunters 1969, 19, 284–317. [Google Scholar] [CrossRef]
- Östman, C.; Roat, C.; Roat Kultima, J.; Rundblom, K. Acontia and mesentery nematocysts of the sea anemone Metridium senile (Linnaeus, 1761) (Cnidaria, Anthozoa). Sci. Mar. 2010, 74, 483–497. [Google Scholar] [CrossRef]
- Schmidt, H. Die nesselkapseln der Anthozoen und irhe Bedeutung fur die phylogenetische Systematik. Helgoländer Wiss. Meeresunters 1972, 23, 422–458. [Google Scholar] [CrossRef]
- Schmidt, H. On evolution in the Anthozoa. In Proceedings of the Second International Coral Reef Symposium, Great Barrier Reef Committee, Queensland, Australasia, 22 June–2 July 1974; Volume 1, pp. 533–560. [Google Scholar]
- Cutress, C.E. An interpretation of the structure and distribution of cnidae in Anthozoa. Syst. Zool. 1955, 4, 120–137. [Google Scholar] [CrossRef]
- Mariscal, R.N. Nematocysts. In Coelenterate Biology: Reviews and New Perspectives; Muscatine, L., Lenhoff, H.M., Eds.; Academic Press: New York, NY, USA, 1974; pp. 129–178. [Google Scholar]
- England, K.W. Nematocysts of sea anemones (Actinaria, Ceriantharia and Corralimorpharia: Cnidaria): Nomenclature. In Coelenterate Biology: Recent Research on Cnidaria and Ctenophora. Developments in Hydrobiology; Williams, R.B., Cornelius, P.F.S., Hughes, R.G., Robson, E.A., Eds.; Springer: Dordrecht, The Netherlands, 1991; Volume 66, pp. 607–691. [Google Scholar] [CrossRef]
- Östman, C. A guideline to nematocyst nomenclature and classification, and some notes on the systematic value of nematocysts. Sci. Mar. 2000, 64, 31–46. [Google Scholar] [CrossRef]
- den Hartog, J.C. Description of two new Ceriantharia from the Caribbean region, Pachyceriantus curacaoensis n. sp. and Arachnanthus nocturnus n. sp. with discussion of the cnidom and of the classification of the Ceriantharia. Zool. Meded. 1977, 51, 211–242. [Google Scholar]
- Stephenson, T.A. The British Sea Anemones, Dulau & Co., Ltd: London, UK, 1928; Volume 1.
- Kayal, E.; Bastian, B.; Pankey, M.S.; Ohdeta, A.H.; Medina, M.; Placketzki, D.C.; Collins, A.G.; Ryan, J.F. Phylogenomics provides a robust topology of the major cnidarian lineages and insights on the origins of key organismal traits. BMC Evol. Biol. 2018, 18, 68. [Google Scholar] [CrossRef]
- Carlgren, O. Ceriantharia. Dan. Ingolf-Exped. 1912, 5, 1–79. [Google Scholar]
- Arai, M.N. A new species of Pachycerianthus, with a discussion of the genus and an appended glossary. Pac. Sci. 1965, 19, 205–218. [Google Scholar]
- Molodtsova, T.N.; Griffiths, C.L.; Acuña, F.H. A new species of shallow-water cerianthid (Cnidaria: Anthozoa) from South Africa, with remarks on the genus Ceriantheopsis. Afr. Nat. Hist. 2011, 7, 2–8. [Google Scholar]
- Stampar, S.N.; Morandini, A.C.; da Silveira, F.L. A new species of Pachycerianthus (Cnidaria, Anthozoa, Ceriantharia) from Tropical Southwestern Atlantic. Zootaxa 2014, 3827, 343–354. [Google Scholar] [CrossRef] [PubMed]
- Stampar, S.N.; Scarabino, F.; Pastorino, F.; Morandini, A.C. A new species of tube-dwelling anemone (Cnidaria, Anthozoa, Cerinathaira, Ceriantheopsis) from the warm temperate South-western Atlantic. J. Mar. Biol. Assoc. U. K. 2015, 96, 1475–1481. [Google Scholar] [CrossRef]
- Stampar, S.N.; Sadie, V.; Keable, S. Ceriantharia (Cnidaria) from Australia, New Zealand and Antarctica with descriptions of four New Species. Rec. Aust. Mus. 2020, 72, 81–100. [Google Scholar] [CrossRef]
- Lopes, C.S.; Ceriello, H.; Morandini, A.C.; Stampar, N. Revision of the genus Ceriantheomorphe (Cnidaria, Anthozoa, Ceriantharia) with description of a new species form the Gulf of Mexico and northwestern Atlantic. Zookeys 2019, 874, 127–148. [Google Scholar] [CrossRef] [PubMed]
- Strömberg, S.M.; Östman, C. The cnidome and internal morphology of Lophelia pertusa (Linnaeus, 1755) (Cnidaria, Anthozoa). Acta Zool. 2016, 98, 191–213. [Google Scholar] [CrossRef] [PubMed]
- Carlgren, O. A survey of the ptychodactiaria, Corallimorpharia and Actiniaria. Kungl. Svenka Vetenskapsakademiens Handlingar 1949, 1, 1–121. [Google Scholar]
- Mariscal, R.N.; McLean, R.B.; Hand, C. The form and function of cnidarian spirocysts. 3. Ultrastructure of the thread and the function of spirocysts. Cell Tissue Res. 1977, 178, 427–433. [Google Scholar] [CrossRef] [PubMed]
- Watson, G.M.; Wood, R.L. Colloquium on the terminology. In The Biology of Nematocysts; Heissinger, D.A., Lenhoff, H.M., Eds.; Academic Press: San Diego, CA, USA, 1988; pp. 21–23. [Google Scholar]
- Acuña, F.H.; Excoffon, A.C.; Zamponi, M.O.; Ricci, L. Importance of nematocysts in taxonomy of acontiarian sea anemones (Cnidaria, Actiniaria): A statistical comparative study. Zool. Anz. 2003, 242, 75–81. [Google Scholar] [CrossRef]
- Garese, A.; Correa, F.G.; Acuña, F.H.; Stampar, S.N. Cnidom in Ceriantharia (Cnidaria, Anthozoa): New findings in the composition and micrometric variations of Cnidocysts. PeerJ 2023, 11, e15549. [Google Scholar] [CrossRef] [PubMed]
- Molodtsova, T.N.; Moskalenko, V.N.; Lipukhin, E.V.; Antokhina, T.I.; Ananeva, M.S.; Simakova, U.V. Cerianthus lloydii (Ceriantharia: Anthozoa: Cnidaria): New status and new perspectives. Biology 2023, 12, 1167. [Google Scholar] [CrossRef] [PubMed]
- Gusmão, L.C.; Grajales, A.; Rodríguez, E. Sea anemones through X-rays: Visualization of two species of Diadumene (Cnidaria, Actiniaria) using micro-CT. Am. Mus. Novit. 2018, 2018, 1–47. [Google Scholar] [CrossRef]
- Sanamyan, N.P.; Sanamyan, K.E.; Tabachnick, K.R. The first species of Actiniaria, Spongiactis japonica gen.n., sp.n. (Cnidaria: Anthozoa), an obligate symbiont of a glass sponge. Invert. Zool. 2012, 9, 127–141. [Google Scholar] [CrossRef]
- Zamponi, M.O.; Acuña, F.H. La variabilidad de los cnidocistos y su importancia en la determinación de clines. Physis 1991, 49, 7–18. [Google Scholar]
- Östman, C.; Møller, F.L. Gross morphology and cnidae of Edwardsiella anemones and larvae (Anthozoa, Edwardsiidae) from the Swedish West Coast. Acta Zool. 2021, 102, 467–482. [Google Scholar] [CrossRef]
- Östman, C.; Borg, F.; Roat, C.; Kultima, J.; Wong, S.Y. Cnidae in the sea anemone Sagartiogeton viduatus (Muller, 1776) (Cnidaria, Anthozoa); A comparison to cnidae in the sea anemone Metridium senile (linnaeus, 1761) (Cnidaria, Anthozoa). Acta Zool. 2012, 94, 392–409. [Google Scholar] [CrossRef]
- Östman, C.; Hydman, J. Nematocyst analysis of Cyanea capillata and Cyanea lamarckii (Scyphozoa, Cnidaria). Sci. Mar. 1997, 61, 313–344. [Google Scholar]
- Williams, R.B. Measurements of cnidae from sea anemones (Cnidaria: Actiniaria). Sci. Mar. 1996, 60, 339–351. [Google Scholar]
- Williams, R.B. Measurements of cnidae from sea anemones (Cnidaria: Actinaria), II: Further studies of differences amongst sample means and their taxonomic relevance. Sci. Mar. 1998, 62, 361–372. [Google Scholar] [CrossRef]
- Williams, R.B. Measurements of cnidae from sea anemones (Cnidaria: Actiniaria), III: Ranges and other measures of statistical dispersion, their interrelations and taxonomic relevance. Sci. Mar. 2000, 64, 49–68. [Google Scholar] [CrossRef]
- Häussermann, V. Identification and taxonomy of soft-bodied hexacorals exemplified by Chilean sea anemones; including guidelines for sampling, preservation and examination. J. Mar. Biol. Assoc. U. K. 2004, 84, 931–936. [Google Scholar] [CrossRef]
- Westfall, J.A. Nematocyst of the sea anemone Metridium. Am. Zool. 1965, 5, 377–393. [Google Scholar] [CrossRef] [PubMed]
- Westfall, J.; Landers, D.; McCallum, J. Ultrastructure of neuro-spirocyte synapses in the sea anemone Aiptasia pallida (Cnidaria, Anthozoa, Zoantharia). J. Morphol. 1999, 241, 165–173. [Google Scholar] [CrossRef]
- Rifkin, J.F. A study of the spirocytes from the Ceriantharia and Actiniaria (Cnidaria: Anthozoa). Cell Tissue Res. 1991, 266, 365–373. [Google Scholar] [CrossRef]
- Williams, R.B. A redescription of the brackish-water sea anemone Nematostella vectensis Stephenson, with an appraisal of congeneric species. J. Nat. Hist. 1975, 9, 51–64. [Google Scholar] [CrossRef]
- Babonis, L.S.; Martindale, M.Q.; Ryan, J.F. Do novel genes drive morphological novelty? An investigation of the nematosomes in the sea anemone Nemastostella vectensis. BMC Evol. Biol. 2016, 16, 114. [Google Scholar] [CrossRef] [PubMed]
- Carlgren, O. The actinian fauna of the Gulf of California. Proc. U. S. Natl. Mus. 1951, 101, 415–449. [Google Scholar] [CrossRef]
- Stampar, S.N.; Maronna, M.M.; Kitahara, M.V.; Reimer, J.D.; Beneti, J.S.; Morandini, A.C. Ceriantharia in current systematics: Life cycles, morphology and genetics. In The Cnidaria, Past, Present and Future; Goffredo, S., Dubinsky, Z., Eds.; Springer: Cham, Switzerland, 2016. [Google Scholar] [CrossRef]
- Stampar, S.N.; Maronna, M.M.; Vermeij, M.J.A.; Silveira, F.L.D.; Morandini, A.C. Evolutionary diversification of Banded tube-dwelling anemones (Cnidaria; Ceriantharia; Isarachnanthus) in the Atlantic Ocean. PLoS ONE 2012, 7, e41091. [Google Scholar] [CrossRef] [PubMed]
- Kise, H.; Fujii, T.; Masucci, G.D.; Biondi, P.; Reimer, J.D. Three new species and the molecular phylogeny of Antipathozoanthus from the Indo-Pacific Ocean (Anthozoa, Hexacorallia, Zoantharia). Zookeys 2017, 725, 97–122. [Google Scholar] [CrossRef] [PubMed]
- Daly, M.; Fautin, D.G.; Cappola, V.A. Systematics of the hexacorallia (Cnidaria: Anthozoa). Zool. J. Linn. Soc. 2003, 139, 419–437. [Google Scholar] [CrossRef]
- Westfall, J.A.; Hand, C. Fine structure of nematocysts in a sea anemone. In Electron Microscopy, Proceedings of the 5th International Congress, 29 August–5 September; Sydney, S., Breese, S.S., Jr., Eds.; Academic Press: Philadelphia, PA, USA, 1962. [Google Scholar]
- Mariscal, R.N. Cnidaria: Cnidae. Biology of the integument. In Invertebrates; Bereiter-Hahn, J., Matoltsy, A.J., Richards, K.S., Eds.; Springer: Berlin/Heidelberg, Germany, 1984; Volume 1, pp. 57–68. [Google Scholar]
- Reft, A.J.; Westfall, J.A.; Fautin, D.G. Formation of the apical flaps in nematocysts of sea anemones (Cnidaria: Actiniaria). Biol. Bull. 2009, 217, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Reft, A.J.; Daly, M. Morphology, distribution, and evolution of apical structure of nematocysts in hexacorallia. J. Morphol. 2012, 273, 121–136. [Google Scholar] [CrossRef] [PubMed]
- Mariscal, R.N.; McLean, R.B. The form and function of cnidarian spirocysts: 2. Ultrastructure of the capsule tip and wall and mechanism of discharge. Cell Tissue Res. 1976, 169, 313–321. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CNIDA TYPES and Subtypes | n | Abundance | Mean (Range Length) × Mean (Range Width) in µm | Length/Width Mean in µm | Carlgren 1940 Recorded cnidae Pachycerianthus multiplicatus | den Hartog 1977 Recorded cnidae Pachycerianthus curacaoensis |
|---|---|---|---|---|---|---|
| b-MASTIGOPHORES | abundant | spirulae, spirulae, 1.2 numerous. common | ||||
| Small, broad | 23 | x | 18.9 (12.5–24.0) × 4.6 (3.5–5.5) | 4.1 | 21.0–25.0 × 4.0 | spirulae 1, 25.7 (19.8–29.7) × 4.4 (3.3–5.3) |
| Small, narrow | 8 | se | 18.5 (10.0–24.0) × 3.5 (3.0–4.5) | 5.3 | ||
| Medium, broad, elongate | 67 | x–xxx | 33.9 (25.6–49.5) × 6.5 (4.5–8.5) | 5.3 | 34.0–38.0 × 3.0–4.5 | spirulae 2, 36.5 (33.0–38.9) × 9.1 (7.3–10.6) |
| Medium, narrow, elongate | 72 | x–xxx | 31.4 (25.0–39.0) × 4.6 (3.5–6.5) | 6.9 | 21.0–25.0 × 4.0 | spirulae, 27.9 (24.4–29.7) ×4.5 (4.0–4.6) |
| Large, elongate with “Faltstück” | 164 | xxx | 55.0–65.0 × about 10.0 | not mentioned | ||
| “Faltstück”, straight * | 34 | se–x | 60.6 (52.0–72.0) × 11.6 (9.50–14.5) | 5.3 | not mentioned | |
| “Faltstück”, undulating * | 66 | xx–xxx | 58.5 (51.0–69.5) × 12.1 (10.0–14.5) | 4.9 | present | |
| “Faltstück”, coiled * | 64 | xx–xxx | 62.8 (55.5–72.0) × 12.3 (9.5–14.0) | 5.2 | present | |
| ISORHIZAS | 0 | not found | no information | not mentioned | not mentioned | |
| SPIROCYSTS | dominant | numerous, up to 67.0 × 10.0 | spirocysts, numerous | |||
| Small narrow | 77 | xx | 21.9 (16.5–25.5) × 3.3 (2.3–4.0) | 7.1 | ||
| Small, narrow, densely coiled | 66 | xxxx | 22.2 (16.5–25.5) × 3.2 (2.5–3.8) | 7.1 | ||
| Medium, broad | 62 | xxx | 43.1 (31.0–49.5) × 6.6 (5.0–11.5) | 6.6 | ||
| Medium, elongate | 90 | xxxx | 37.2 (26.0–50.0) × 4.4 (2.5–6.0) | 8.7 | 31.3 (19.8–39.6) × 4.3 (2.0–5.9) | |
| Large, broad | 82 | xxx–xxxx | 59.8 (49.0–71.5) × 8.9 (6.0–11.5) | 6.9 | numerous, up to 67.0 × 10.0 | |
| Large, elongate | 68 | se–xxxx | 57.7 (50.0–68.5) × 7.4 (4.0–9.0) | 7.9 | ||
| PTYCHOCYSTS | present | not mentioned | not mentioned | |||
| Small, broad, apically bent | 1 | se | 24.8 × 7.5 | 3.2 | ||
| Medium, elongate | 36 | x | 45.8 (39.0–49.5) × 7.6 (5.0–10.0) | 6.1 | ||
| Large, elongate | 5 | se | 56.8 (51.0–59.0) × 7.0 (5.0–9.5) | 8.6 |
| CNIDA TYPES and Subtypes | n | Abundance | Mean (Range Length) × Mean (Range Width) in µm | Length/Width mean in µm | Carlgren 1940 Recorded cnidae Pachycerianthus multiplicatus | den Hartog 1977 Recorded cnidae Pachycerianthus curacaoensis |
|---|---|---|---|---|---|---|
| b-MASTIGOPHORES | present | spirulae, common shaft short and long | ||||
| Small, broad | 19 | se | 20.1 (12.5–24.7) × 4.5 (3.7–6.0) | 4.5 | 23.1 (21.1–24.4) × 3.2 (2.6–3.3) | |
| Small, narrow ok | 1 | Se | 23.8 × 4.3 | 5.5 | 20.3 (17.8–24.4) × 2.9 (2.6–3.3) | |
| Medium, broad, elongate | 65 | xxx | 35.7 (27.0–46.5) × 5.7 (5.0–10.0) | 5.4 | short shaft, 3 spine-rows, 36.0–41.0 × 5.0–5.5 | 34.4 (29.7–37.6) × 5.8 (5.3–7.3) |
| Medium, narrow, elongate | 64 | xx | 31.5 (26.0–39.0) × 4.8 (3.5–7.5) | 6.7 | numerous, 27.5–31.0 × 3.0–4.0 | |
| Large, elongate, tiny “Faltstück” | 96 | xxx | 55.3 (46.0–63.0) × 9.7 (6.5–14.5) | 5.9 | no Faltstück mentioned, 58.0–67.0 × 7.5–8.5 | not identified |
| ISORHIZAS | seldom | |||||
| Large, very large, broad | 3 | se | 78.6 (67.0–82.0) × 15.5 (13.0–18.5) | not identified | not identified | |
| SPIROCYSTS | dominant | spirocysts, numerous, no measurements | spirocysts, very numerous | |||
| Small, elongate, pointed end | 18 | x | 23.7 (21.5–25.5) × 3.2 (3.0 4.0) | 6.6 | ||
| Small, elongate rounded end | 10 | x | 23.8 (22.5–25.5) × 3.6 (3.0–4.0) | 6.6 | ||
| Small to medium, densely coiled | 60 | xxxx | 24.7 (19.0–32.5) × 3.6 (3.0–4.5) | 9.9 | ||
| Medium, broad rounded end | 72 | xxxx | 44.7 (35.0–49.5) × 7.0 (4.5–9.5) | 6.4 | 32.4 (26.4–39.6) × 4.7 (3.6–7.3) | |
| Medium, elongate, pointed end | 31 | x | 43.6 (38.5–49.0) × 5.8 (3.5–7.0) | 7.8 | 32.4 (26.4–39.6) × 4.7 (3.6–7.3) | |
| Large, broad | 55 | xxx | 55.4 (50.0–59.5) × 8.0 (6.5–12.0) | 6.9 | ||
| Large, elongate | 62 | xxxx | 53.9 (50.5–57.5) × 7.5 (6.0–8.5) | 7.3 | ||
| PTYCHOCYSTS | present | atrichs, not rare, of about same width | atrichs, common | |||
| Small, broad, apically bent | 1 | se | 28.6 × 7.7 | 3.7 | ||
| Medium broad straight | 3 | se | 46.8 (44.0–47.5) × 9.3 (7.5–10.5) | 5.1 | ||
| Medium, elongate, straight | 74 | xx | 43.8 (33.6–49.9) × 7.0 (5.4–9.5) | 6.3 | 40.0–48.0 × 5.5–7.0 | 30.0 (27.1–33.0) × 5.5 (5.3–5.9) |
| Large, elongate, straight | 15 | se | 53.4 (50.0–54.5) × 7.3 (7.5–10.0) | 7.4 |
| CNIDA TYPES and Subtypes | n | Abundance | Mean (Range Length × Mean (Range Width) in µm | Length/Width Mean in µm | Carlgren 1940 Recorded cnidae Pachycerianthus multiplicatus | den Hartog 1977 Recorded cnidae Pachycerianthus curacaoensis |
|---|---|---|---|---|---|---|
| b-MASTIGOPHORES | present | no information | no information | |||
| Small, broad | 2 | se | 21.5–23.0 × 6.5–7.5 | 3.3 | ||
| Small, narrow | 1 | se | 23.0 × 3.0 | 7.7 | ||
| Medium, broad, elongate | 21 | x | 41.1 (30.5–46.0) × 7.3 (5.5–9.5) | 5.8 | ||
| Medium, broad, oval | 1 | se | 26.0 × 7.9 | 3.3 | ||
| Medium, narrow, elongate | 2 | xx | 29.1 (29.0–29.5) × 4.7 (4.5–5.0) | 6.2 | ||
| Large, broad | 1 | se | 51.0 × 9.0 | 5.7 | ||
| Large, elongate, tiny “Faltstück” | 3 | se | 61.6 (50.0–74.7) × 10.0 (7.5–11.5) | 6.6 | ||
| ISORHIZAS | present | no information | no information | |||
| Medium, broad | 3 | se | 45.8 (44.0–47.5) × 8.8 (8.5–9.0) | 5.2 | ||
| Large, broad | 8 | se | 71.8 (58.0–80.0) × 15.7 (12.0–20.0) | 4.7 | ||
| Very large | 6 | se | 85.2 (80.5–90.2) × 16.6 (15.5–18.5) | 4.9 | ||
| SPIROCYSTS | present | |||||
| Small to medium densely coiled | 3 | se | 25.7 (22.0–30.5) × 2.8 (2.5–3.5) | 9.5 | ||
| Medium, elongate | 11 | se | 38.8 (27.0–49.0) × 5.4 (3.0–8.0) | 7.7 | ||
| Large, elongate | 13 | x | 54.4 (50.5–60.0) × 6.4 (5.5–8.0) | 8.5 | ||
| PTYCHOCYSTS | dominant | no information | no information | |||
| Tiny, straight | 1 | se | 8.2 × 1.8 | 4.5 | ||
| Small, elongate | 1 | se | 26.6 × 6.8 | 3.9 | ||
| Small, drop-shaped | 3 | se | 25.2 (25.0–25.5) × 8.2 (8.0–8.5) | 3.1 | ||
| Medium, drop-shaped ok | 8 | se | 36.5 (27.5–48.0) × 10.3 (7.5–14.0) | 3.6 | ||
| Medium, elongate, broad | 40 | xxx | 41.9 (33.0–49.0) × 11.7 (10.0–19.0) | 3.6 | ||
| Medium, elongate, narrow | 32 | xx | 40.9 (30.0–50.0) × 8.9 (6.0–10.5) | 4.6 | ||
| Medium, elongate, apically bent | 30 | xx | 44.7 (27.0–50.0) × 12.8 (8.5–20.0) | 3.6 | ||
| Large, drop-shaped broad | 87 | xxx | 71.9 (50.0–80.0) × 24.7 (9.0–35.5) | 3.1 | ||
| Large, elongate broad | 30 | x | 62.6 (50.5–79.5) × 13.9 (10.5–23.0) | 4.7 | ||
| Large elongate, narrow | 32 | se | 56.9 (50.5–64.0) × 9.5 (7.0–10.5) | 6.1 | ||
| Large, apically bent, elongate, broad | 145 | xxxx | 62.4 (50.0–79.5) × 15.4 (10.0–21.5) | 4.1 | ||
| Large, apically bent elongate narrow | 2 | se | 63.3 (62.5–63.5) × 10.1 (10.0–10.5) | 6.3 | ||
| Very large, drop-shaped | 66 | absent, xxx | 90.4 (80.0–106.0) × 28,9 (17.5–38.5) | 3.2 | ||
| Very large oval | 3 | se | 83.8 (82.5–85.0) × 36.8 (35.0–41.0) | 2.2 | ||
| Very large, elongate, broad | 8 | se | 92.0 (81.5–102.5) × 27.3 (20.0–33.0) | 3.0 | ||
| Very large, apically bent | 1 | se | 89.0 × 17.0 | 5.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Forero-Mejia, A.C.; Duchatelet, L.; Östman, C. Cnida Morphology as Taxonomic Tools within Tube-Dwelling Anemones (Ceriantharia, Cnidaria). Oceans 2024, 5, 491-521. https://doi.org/10.3390/oceans5030029
Forero-Mejia AC, Duchatelet L, Östman C. Cnida Morphology as Taxonomic Tools within Tube-Dwelling Anemones (Ceriantharia, Cnidaria). Oceans. 2024; 5(3):491-521. https://doi.org/10.3390/oceans5030029
Chicago/Turabian StyleForero-Mejia, Anny C., Laurent Duchatelet, and Carina Östman. 2024. "Cnida Morphology as Taxonomic Tools within Tube-Dwelling Anemones (Ceriantharia, Cnidaria)" Oceans 5, no. 3: 491-521. https://doi.org/10.3390/oceans5030029
APA StyleForero-Mejia, A. C., Duchatelet, L., & Östman, C. (2024). Cnida Morphology as Taxonomic Tools within Tube-Dwelling Anemones (Ceriantharia, Cnidaria). Oceans, 5(3), 491-521. https://doi.org/10.3390/oceans5030029