
Adiponectin and Leptin during Pregnancy: A Systematic Review of Their Association with Pregnancy Disorders, Fetal Growth and Placental Function

 , and
, and 
Abstract
1. Introduction
2. Methods
2.1. Data Sources
2.2. Search Strategy
2.3. Data Collection Process
2.4. Data Items
2.5. Assessment of Risk of Bias
3. Results
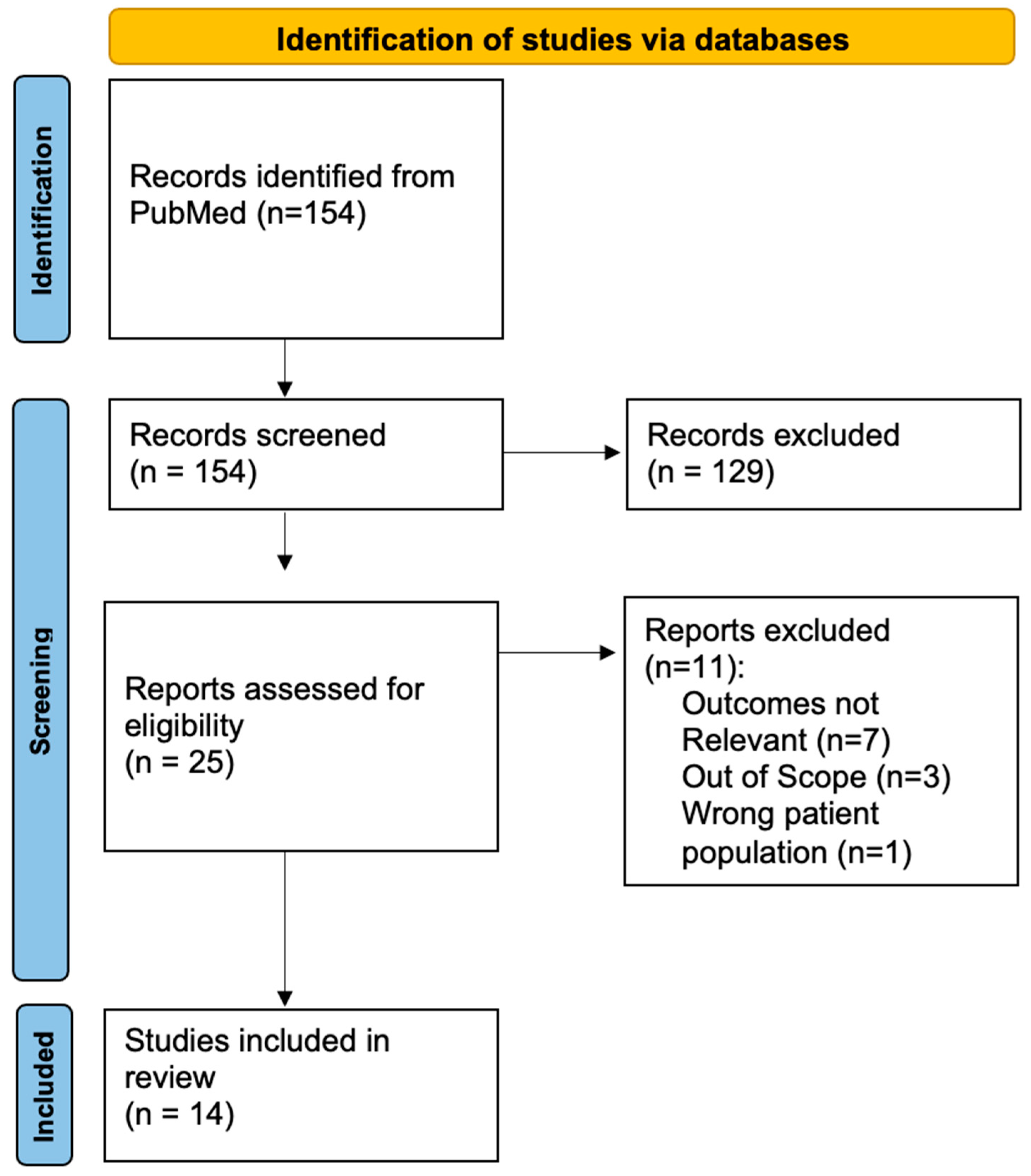
3.1. Search Results
3.2. The Relationship between Maternal Leptin and Adiponectin on Fetal Growth
3.2.1. Association between of Maternal Leptin and Fetal Growth
3.2.2. Association between Maternal Adiponectin and Fetal Growth
3.3. Association between Maternal Obesity or Leptin and Placental Receptors and Nutrient Transporters
3.3.1. Association between Maternal Obesity or Leptin and Placental Expression of Receptors and Nutrient Transporters
3.3.2. Association between Maternal Obesity or Leptin and Protein Expression of Placental Receptors and Nutrient Transporters
3.3.3. Association between Maternal Adiponectin and Placental Expression of Receptors and Nutrient Transporters
3.3.4. Correlation between Maternal Adiponectin and Placental Receptors and Nutrient Transporters
3.4. Association between Maternal Adiponectin and Leptin and Placental Inflammatory Signaling Pathways
3.4.1. Association between Elevated Maternal Leptin and Placental Inflammatory Signaling Pathways
3.4.2. Association between Reduced Adiponectin and Placental Inflammatory Signaling Pathways
3.5. Adiponectin and Leptin in GDM and PE
Association between Elevated Leptin and Reduced Adiponectin in GDM and PE
4. Discussion
4.1. Maternal Leptin and Adiponectin Influences on Fetal Growth
4.2. Maternal Leptin and Adiponectin Influences Placental Nutrient Transporters
4.3. Maternal Leptin and Adiponectin in Gestational Diabetes and Preeclampsia
5. Limitations
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Moyce Gruber, B.L.; Dolinsky, V.W. The Role of Adiponectin during Pregnancy and Gestational Diabetes. Life 2023, 13, 301. [Google Scholar] [CrossRef]
- Zeng, Z.; Liu, F.; Li, S. Metabolic Adaptations in Pregnancy: A Review. Ann. Nutr. Metab. 2017, 70, 59–65. [Google Scholar] [CrossRef]
- Lees, C.C.; Stampalija, T.; Baschat, A.A.; da Silva Costa, F.; Ferrazzi, E.; Figueras, F.; Hecher, K.; Kingdom, J.; Poon, L.C.; Salomon, L.J.; et al. ISUOG Practice Guidelines: Diagnosis and management of small-for-gestational-age fetus and fetal growth restriction. Ultrasound Obstet. Gynecol. 2020, 56, 298–312. [Google Scholar] [CrossRef] [PubMed]
- Sapantzoglou, I.; Vlachos, D.E.; Papageorgiou, D.; Varthaliti, A.; Rodolaki, K.; Daskalaki, M.A.; Psarris, A.; Pergialiotis, V.; Stavros, S.; Daskalakis, G.; et al. Maternal Blood Adipokines and Their Association with Fetal Growth: A Meta-Analysis of the Current Literature. J. Clin. Med. 2024, 13, 1667. [Google Scholar] [CrossRef] [PubMed]
- Kabbani, N.; Blüher, M.; Stepan, H.; Stumvoll, M.; Ebert, T.; Tönjes, A.; Schrey-Petersen, S. Adipokines in Pregnancy: A Systematic Review of Clinical Data. Biomedicines 2023, 11, 1419. [Google Scholar] [CrossRef] [PubMed]
- Kornacki, J.; Gutaj, P.; Kalantarova, A.; Sibiak, R.; Jankowski, M.; Wender-Ozegowska, E. Endothelial Dysfunction in Pregnancy Complications. Biomedicines 2021, 9, 1756. [Google Scholar] [CrossRef]
- Briffa, J.F.; McAinch, A.J.; Romano, T.; Wlodek, M.E.; Hryciw, D.H. Leptin in pregnancy and development: A contributor to adulthood disease? Am. J. Physiol. Endocrinol. Metab. 2015, 308, 16. [Google Scholar] [CrossRef]
- Blüher, M. Adipokines—Removing road blocks to obesity and diabetes therapy. Mol. Metab. 2014, 3, 230–240. [Google Scholar] [CrossRef]
- Howell, K.R.; Powell, T.L. Effects of maternal obesity on placental function and fetal development. Reproduction 2017, 153, R97–R108. [Google Scholar] [CrossRef]
- Huse, O.; Hettiarachchi, J.; Gearon, E.; Nichols, M.; Allender, S.; Peeters, A. Obesity in Australia. Obes. Res. Clin. Pract. 2018, 12, 29–39. [Google Scholar] [CrossRef]
- Segovia, S.A.; Vickers, M.H.; Gray, C.; Reynolds, C.M. Maternal obesity, inflammation, and developmental programming. Biomed. Res. Int. 2014, 2014, 418975. [Google Scholar] [CrossRef] [PubMed]
- Tessier, D.R.; Ferraro, Z.M.; Gruslin, A. Role of leptin in pregnancy: Consequences of maternal obesity. Placenta 2013, 34, 205–211. [Google Scholar] [CrossRef] [PubMed]
- McIntyre, H.D.; Catalano, P.; Zhang, C.; Desoye, G.; Mathiesen, E.R.; Damm, P. Gestational diabetes mellitus. Nat. Rev. Dis. Primers 2019, 5, 47. [Google Scholar] [CrossRef] [PubMed]
- Catalano, P.M.; Shankar, K. Obesity and pregnancy: Mechanisms of short term and long term adverse consequences for mother and child. Br. Med. J. 2017, 356, j1. [Google Scholar] [CrossRef]
- Madan, J.C.; Davis, J.M.; Craig, W.Y.; Collins, M.; Allan, W.; Quinn, R.; Dammann, O. Maternal obesity and markers of inflammation in pregnancy. Cytokine 2009, 47, 61–64. [Google Scholar] [CrossRef]
- Cameo, P.; Bischof, P.; Calvo, J.C. Effect of leptin on progesterone, human chorionic gonadotropin, and interleukin-6 secretion by human term trophoblast cells in culture. Biol. Reprod. 2003, 68, 472–477. [Google Scholar] [CrossRef]
- Lappas, M.; Permezel, M.; Rice, G.E. Leptin and adiponectin stimulate the release of proinflammatory cytokines and prostaglandins from human placenta and maternal adipose tissue via nuclear factor-kappaB, peroxisomal proliferator-activated receptor-gamma and extracellularly regulated kinase 1/2. Endocrinology 2005, 146, 3334–3342. [Google Scholar]
- Rosario, F.J.; Kanai, Y.; Powell, T.L.; Jansson, T. Increased placental nutrient transport in a novel mouse model of maternal obesity with fetal overgrowth. Obesity 2015, 23, 1663–1670. [Google Scholar] [CrossRef]
- RANZCOG. Clinical Guidance Statement: Diagnosis and Management of Suspected Fetal Macrosomia. 2021. Available online: https://ranzcog.edu.au/wp-content/uploads/2022/05/Diagnosis-and-management-of-suspected-fetal-macrosomia.pdf (accessed on 18 July 2024).
- Melamed, N.; Baschat, A.; Yinon, Y.; Athanasiadis, A.; Mecacci, F.; Figueras, F.; Berghella, V.; Nazareth, A.; Tahlak, M.; McIntyre, H.D.; et al. FIGO (international Federation of Gynecology and obstetrics) initiative on fetal growth: Best practice advice for screening, diagnosis, and management of fetal growth restriction. Int. J. Gynaecol. Obstet. 2021, 152 (Suppl. S1), 3–57. [Google Scholar] [CrossRef]
- Eick, S.M.; Goin, D.E.; Chartres, N.; Lam, J.; Woodruff, T.J. Assessing risk of bias in human environmental epidemiology studies using three tools: Different conclusions from different tools. Syst. Rev. 2020, 9, 249. [Google Scholar] [CrossRef]
- Lazo-de-la-Vega-Monroy, M.L.; Mata-Tapia, K.A.; Garcia-Santillan, J.A.; Corona-Figueroa, M.A.; Gonzalez-Dominguez, M.I.; Gomez-Zapata, H.M.; Malacara, J.M.; Daza-Benitez, L.; Barbosa-Sabanero, G. Association of placental nutrient sensing pathways with birth weight. Reproduction 2020, 160, 455–468. [Google Scholar] [CrossRef] [PubMed]
- Lekva, T.; Roland, M.C.P.; Michelsen, A.E.; Friis, C.M.; Aukrust, P.; Bollerslev, J.; Henriksen, T.; Ueland, T. Large Reduction in Adiponectin during Pregnancy Is Associated with Large-for-Gestational-Age Newborns. J. Clin. Endocrinol. Metab. 2017, 102, 2552–2559. [Google Scholar] [CrossRef]
- Shroff, M.R.; Holzman, C.; Tian, Y.; Evans, R.W.; Sikorskii, A. Mid-pregnancy maternal leptin levels, birthweight for gestational age and preterm delivery. Clin. Endocrinol. 2013, 78, 607–613. [Google Scholar] [CrossRef]
- Schoots, M.H.; Bourgonje, M.F.; Bourgonje, A.R.; Prins, J.R.; van Hoorn, E.G.M.; Abdulle, A.E.; Muller Kobold, A.C.; van der Heide, M.; Hillebrands, J.-L.; van Goor, H.; et al. Oxidative stress biomarkers in fetal growth restriction with and without preeclampsia. Placenta 2021, 115, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Stefaniak, M.; Dmoch-Gajzlerska, E. Maternal Serum and Cord Blood Leptin Concentrations at Delivery in Normal Pregnancies and in Pregnancies Complicated by Intrauterine Growth Restriction. Obes. Facts 2022, 15, 62–69. [Google Scholar] [CrossRef]
- Shang, M.; Dong, X.; Hou, L. Correlation of adipokines and markers of oxidative stress in women with gestational diabetes mellitus and their newborns. J. Obstet. Gynaecol. Res. 2018, 44, 637–646. [Google Scholar] [CrossRef]
- Kyriakakou, M.; Malamitsi-Puchner, A.; Militsi, H.; Boutsikou, T.; Margeli, A.; Hassiakos, D.; Kanaka-Gantenbein, C.; Papassotiriou, I.; Mastorakos, G. Leptin and adiponectin concentrations in intrauterine growth restricted and appropriate for gestational age fetuses, neonates, and their mothers. Eur. J. Endocrinol. 2008, 158, 343–348. [Google Scholar] [CrossRef]
- Zamarian, A.C.; Araujo Júnior, E.; Daher, S.; Rolo, L.C.; Moron, A.F.; Nardozza, L.M. Evaluation of biochemical markers combined with uterine artery Doppler parameters in fetuses with growth restriction: A case-control study. Arch. Gynecol. Obstet. 2016, 294, 715–723. [Google Scholar] [CrossRef] [PubMed]
- Kelly, A.C.; Powell, T.L.; Jansson, T. Placental function in maternal obesity. Clin. Sci. 2020, 134, 961–984. [Google Scholar] [CrossRef]
- Nogues, P.; Dos Santos, E.; Couturier-Tarrade, A.; Berveiller, P.; Arnould, L.; Lamy, E.; Grassin-Delyle, S.; Vialard, F.; Dieudonne, M.N. Maternal Obesity Influences Placental Nutrient Transport, Inflammatory Status, and Morphology in Human Term Placenta. J. Clin. Endocrinol. Metab. 2021, 106, e1880–e1896. [Google Scholar] [CrossRef]
- Balachandiran, M.; Bobby, Z.; Dorairajan, G.; Gladwin, V.; Vinayagam, V.; Packirisamy, R.M. Decreased maternal serum adiponectin and increased insulin-like growth factor-1 levels along with increased placental glucose transporter-1 expression in gestational diabetes mellitus: Possible role in fetal overgrowth. Placenta 2021, 104, 71–80. [Google Scholar] [CrossRef]
- Nogues, P.; Dos Santos, E.; Jammes, H.; Berveiller, P.; Arnould, L.; Vialard, F.; Dieudonné, M.N. Maternal obesity influences expression and DNA methylation of the adiponectin and leptin systems in human third-trimester placenta. Clin. Epigenetics 2019, 11, 20. [Google Scholar] [CrossRef]
- Aye, I.L.; Gao, X.; Weintraub, S.T.; Jansson, T.; Powell, T.L. Adiponectin inhibits insulin function in primary trophoblasts by PPARα-mediated ceramide synthesis. Mol. Endocrinol. 2014, 28, 512–524. [Google Scholar] [CrossRef] [PubMed]
- Aye, I.L.; Lager, S.; Ramirez, V.I.; Gaccioli, F.; Dudley, D.J.; Jansson, T.; Powell, T.L. Increasing maternal body mass index is associated with systemic inflammation in the mother and the activation of distinct placental inflammatory pathways. Biol. Reprod. 2014, 90, 129. [Google Scholar] [CrossRef] [PubMed]
- Hogg, K.; Blair, J.D.; von Dadelszen, P.; Robinson, W.P. Hypomethylation of the LEP gene in placenta and elevated maternal leptin concentration in early onset pre-eclampsia. Mol. Cell Endocrinol. 2013, 367, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Domali, E.; Messinis, I.E. Leptin in pregnancy. J. Matern. Fetal Neonatal Med. 2002, 12, 222–230. [Google Scholar] [CrossRef]
- Khant Aung, Z.; Grattan, D.R.; Ladyman, S.R. Pregnancy-induced adaptation of central sensitivity to leptin and insulin. Mol. Cell Endocrinol. 2020, 516, 110933. [Google Scholar] [CrossRef]
- Paganoti, C.F.; Costa, R.A.; Oliveira, A.M.S.S.; Hoshida, M.S.; Francisco, R.P.V. Adiponectin does not improve the prediction of insulin need in pregnant women with gestational diabetes mellitus. Endo Metabol. Sci. 2021, 3, 100095. [Google Scholar] [CrossRef]
- Caselli, C. Role of adiponectin system in insulin resistance. Mol. Genet. Metab. 2014, 113, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Yadav, A.; Kataria, M.A.; Saini, V.; Yadav, A. Role of leptin and adiponectin in insulin resistance. Clin. Chim. Act. 2013, 417, 80–84. [Google Scholar] [CrossRef]
- RObeidat, R.A.; Abdo, N.; Sakee, B.; Alghazo, S.; Jbarah, O.F.; Hazaimeh, E.A.; Albeitawi, S. Maternal and fetal serum leptin levels and their association with maternal and fetal variables and labor: A cross-sectional study. Ann. Med. Surg. 2021, 72, 103050. [Google Scholar] [CrossRef]
- Goto, E. Maternal blood leptin concentration in small for gestational age: A meta-analysis. Eur. J. Pediatr. 2019, 178, 763–770. [Google Scholar] [CrossRef]
- Maymó, J.L.; Pérez Pérez, A.; Gambino, Y.; Calvo, J.C.; Sánchez-Margalet, V.; Varone, C.L. Review: Leptin gene expression in the placenta—Regulation of a key hormone in trophoblast proliferation and survival. Placenta 2011, 32 (Suppl. S2), S146–S153. [Google Scholar] [CrossRef]
- Pérez-Pérez, A.; Guadix, P.; Maymó, J.; Dueñas, J.L.; Varone, C.; Fernández-Sánchez, M.; Sánchez-Margalet, V. Insulin and Leptin Signaling in Placenta from Gestational Diabetic Subjects. Horm. Metab. Res. 2016, 48, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, M.M.; Wermelin, S.; Stener-Victorin, E.; Wernstedt Asterholm, I.; Benrick, A. Adiponectin Deficiency Alters Placenta Function but Does Not Affect Fetal Growth in Mice. Int. J. Mol. Sci. 2022, 23, 4939. [Google Scholar] [CrossRef]
- Aye, I.L.; Powell, T.L.; Jansson, T. Review: Adiponectin—The missing link between maternal adiposity, placental transport and fetal growth? Placenta 2013, 34, S40–S45. [Google Scholar] [CrossRef]
- Gordijn, S.J.; Beune, I.M.; Thilaganathan, B.; Papageorghiou, A.; Baschat, A.A.; Baker, P.N.; Silver, R.M.; Wynia, K.; Ganzevoort, W. Consensus definition of fetal growth restriction: A Delphi procedure. Ultrasound Obstet. Gynecol. 2016, 48, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Perinatal Society of Australia and New Zealand and Centre of Research Excellence in Stillbirth. Position Statement: Detection and Management of Fetal Growth Restriction in Singleton Pregnancies; Centre of Research Excellence in Stillbirth: Brisbane, Australia, 2023. [Google Scholar]
- Miller, S.L.; Huppi, P.S.; Mallard, C. The consequences of fetal growth restriction on brain structure and neurodevelopmental outcome. J. Physiology 2016, 594, 807–823. [Google Scholar] [CrossRef] [PubMed]
- Molina, L.C.G.; Odibo, L.; Zientara, S.; Običan, S.G.; Rodriguez, A.; Stout, M.; Odibo, A.O. Validation of Delphi procedure consensus criteria for defining fetal growth restriction. Ultrasound Obstet. Gynecol. Off. J. Int. Soc. Ultrasound Obstet. Gynecol. 2020, 56, 61–66. [Google Scholar] [CrossRef]
- Obradovic, M.; Sudar-Milovanovic, E.; Soskic, S.; Essack, M.; Arya, S.; Stewart, A.J.; Gojobori, T.; Isenovic, E.R. Leptin and Obesity: Role and Clinical Implication. Front. Endocrinol. 2021, 12, 585887. [Google Scholar] [CrossRef]
- Chen, Y.-Y.; Rosario, F.J.; Abu Shehab, M.; Powell, T.L.; Gupta, M.B.; Jansson, T. Increased ubiquitination and reduced plasma membrane trafficking of placental amino acid transporter SNAT-2 in human IUGR. Clin. Sci. 2015, 129, 1131–1141. [Google Scholar] [CrossRef] [PubMed]
- Maiiendran, D.; Donnai, P.; Glazier, J.D.; D’Souza, S.W.; Boyd, R.D.H.; Sibley, C.P. Amino Acid (System A) Transporter Activity in Microvillous Membrane Vesicles from the Placentas of Appropriate and Small for Gestational Age Babies. Pediatr. Res. 1993, 34, 661–665. [Google Scholar] [CrossRef]
- Farley, D.M.; Choi, J.; Dudley, D.J.; Li, C.; Jenkins, S.L.; Myatt, L.; Nathanielsz, P.W. Placental Amino Acid Transport and Placental Leptin Resistance in Pregnancies Complicated by Maternal Obesity. Placenta 2010, 31, 718–724. [Google Scholar] [CrossRef] [PubMed]
- Shang, M.; Wen, Z. Increased placental IGF-1/mTOR activity in macrosomia born to women with gestational diabetes. Diabetes Res. Clin. Pract. 2018, 146, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Kaufman, M.R.; Brown, T.L. AMPK and Placental Progenitor Cells. In AMP-Activated Protein Kinase; Cordero, M.D., Viollet, B., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 73–79. [Google Scholar]
- Dong, J.; Shin, N.; Chen, S.; Lei, J.; Burd, I.; Wang, X. Is there a definite relationship between placental mTOR signaling and fetal growth? Biol. Reprod. 2020, 103, 471–486. [Google Scholar] [CrossRef]
- Yamauchi, T.; Kamon, J.; Minokoshi, Y.; Ito, Y.; Waki, H.; Uchida, S.; Yamashita, S.; Noda, M.; Kita, S.; Ueki, K.; et al. Adiponectin stimulates glucose utilization and fatty-acid oxidation by activating AMP-activated protein kinase. Nat. Med. 2002, 8, 1288–1295. [Google Scholar] [CrossRef]
- Kumagai, A.; Itakura, A.; Koya, D.; Kanasaki, K. AMP-Activated Protein (AMPK) in Pathophysiology of Pregnancy Complications. Int. J. Mol. Sci. 2018, 19, 3076. [Google Scholar] [CrossRef]
- Xu, J.; Zhao, Y.H.; Chen, Y.P.; Yuan, X.L.; Wang, J.; Zhu, H.; Lu, C.M. Maternal Circulating Concentrations of Tumor Necrosis Factor-Alpha, Leptin, and Adiponectin in Gestational Diabetes Mellitus: A Systematic Review and Meta-Analysis. Sci. World J. 2014, 2014, 926932. [Google Scholar] [CrossRef]
- Stanirowski, P.J.; Szukiewicz, D.; Majewska, A.; Wątroba, M.; Pyzlak, M.; Bomba-Opoń, D.; Wielgoś, M. Placental expression of glucose transporters GLUT-1, GLUT-3, GLUT-8 and GLUT-12 in pregnancies complicated by gestational and type 1 diabetes mellitus. J. Diabetes Investig. 2022, 13, 560–570. [Google Scholar] [CrossRef]
- Kc, K.; Shakya, S.; Zhang, H. Gestational diabetes mellitus and macrosomia: A literature review. Ann. Nutr. Metab. 2015, 66 (Suppl. S2), 14–20. [Google Scholar] [CrossRef]
- Stanirowski, P.J.; Szukiewicz, D.; Pyzlak, M.; Abdalla, N.; Sawicki, W.; Cendrowski, K. Impact of pre-gestational and gestational diabetes mellitus on the expression of glucose transporters GLUT-1, GLUT-4 and GLUT-9 in human term placenta. Endocrine 2017, 55, 799–808. [Google Scholar] [CrossRef] [PubMed]
- Beneventi, F.; Locatelli, E.; De Amici, M.; Cavagnoli, C.; Bellingeri, C.; De Maggio, I.; Ruspini, B.; Spinillo, A. Maternal and fetal Leptin and interleukin 33 concentrations in pregnancy complicated by obesity and preeclampsia. J. Matern.-Fetal Neonatal Med. 2020, 33, 3942–3948. [Google Scholar] [CrossRef] [PubMed]
- Spinelli, M.; Zdanowicz, J.A.; Keller, I.; Nicholson, P.; Raio, L.; Amylidi-Mohr, S.; Mosimann, B.; Surbek, D.; Mueller, M. Hypertensive disorders of pregnancy share common cfDNA methylation profiles. Sci. Rep. 2022, 12, 19837. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Reference | Maternal Serum Concentration Compared to Control | Offspring Birthweight | p-Value |
|---|---|---|---|
| Lazo-de-la-Vega-Monroy et al. [22] | Increased | LGA | 0.028 |
| Lekva et al. [23] | Increased | LGA | 0.047 |
| Shroff et al. [24] | Increased | LGA | <0.01 |
| Shroff et al. [24] | Decreased | SGA | <0.05 |
| Schoots et al. [25] | Increased | FGR | <0.01 |
| Stefaniak et al. [26] | Increased | FGR | 0.01 |
| Shang et al. [27] | Increased | Macrosomia | ns |
| Kyriakakou et al. [28] | Increased | FGR | <0.05 |
| Reference | Maternal Serum Concentration Compared to Control | Offspring Birthweight | p-Value |
|---|---|---|---|
| Lazo-de-la-Vega-Monroy et al. [22] | Increased | SGA | <0.05 |
| Lekva et al. [23] | Decreased | LGA | 0.007 |
| Shang et al. [27] | Decreased | Macrosomia | <0.05 |
| Kyriakakou et al. [28] | Decreased | FGR | <0.05 |
| Zamarian et al. [29] | Decreased | FGR | ns |
| Reference | Participant Characteristics | mRNA | p-Value |
|---|---|---|---|
| Nogues et al. [31] | obese | Glut1 ↓ | 0.0273 |
| Nogues et al. [31] | obese | Snat1 ↓ | 0.0273 |
| Nogues et al. [31] | obese | Snat2 ↓ | 0.0039 |
| Balachandiran et al. [32] | ↑ leptin; GDM | Glut1 ↓ | <0.05 |
| Nogues et al. [33] | ↑ leptin; obese | LepR | ns |
| Reference | Participant Characteristics | Protein | p-Value |
|---|---|---|---|
| Nogues et al. [31] | obese | GLUT1 ↓ | 0.0221 |
| Nogues et al. [31] | obese | SNAT1 ↓ | 0.0283 |
| Lekva et al. [22] | ↑ leptin; LGA | SNAT2 ↓ | 0.01 |
| Balachandiran et al. [32] | ↑ leptin; GDM | GLUT1 ↑ | <0.05 |
| Nogues et al. [33] | ↑ leptin; obese | LEPR ↓ | <0.01 |
| Reference | Participant Characteristics | mRNA | p-Value |
|---|---|---|---|
| Lekva et al. [23] | ↓ adiponectin; LGA | Snat2 ↓ | 0.01 |
| Lekva et al. [23] | ↓ adiponectin; LGA | AdipoR1 ↓ | ns |
| Lekva et al. [23] | ↓ adiponectin; LGA | AdipoR2 ↓ | 0.001 |
| Balachandiran et al. [32] | ↓ adiponectin; GDM | Glut1 ↑ | <0.05 |
| Nogues et al. [33] | ↓ adiponectin; obese | AdipoR1 ↓ | <0.05 |
| Nogues et al. [33] | ↓ adiponectin; obese | AdipoR2 ↓ | <0.05 |
| Reference | Participant Characteristics | Protein | p-Value |
|---|---|---|---|
| Balachandiran et al. [32] | ↓ adiponectin; GDM | GLUT1 ↑ | <0.05 |
| Nogues et al. [33] | ↓ adiponectin; obese | ADIPOR1 ↓ | <0.05 |
| Nogues et al. [33] | ↓ adiponectin; obese | ADIPOR2 ↓ | <0.05 |
| Reference | Participant Characteristics | Protein | p-Value |
|---|---|---|---|
| Lazo-de-la-Vega-Monroy, et al. [22] | ↑ leptin; SGA | pAMPK ↑ | <0.05 |
| Lazo-de-la-Vega-Monroy, et al. [22] | ↑ leptin; SGA | p-mTOR ↓ | <0.02 |
| Lazo-de-la-Vega-Monroy, et al. [22] | ↓ adiponectin; SGA | pAMPK ↑ | <0.05 |
| Lazo-de-la-Vega-Monroy, et al. [22] | ↓ adiponectin; SGA | p-mTOR ↓ | 0.02 |
| Reference | Maternal Serum Concentration | Pathology | p-Value |
|---|---|---|---|
| Shroff et al. [24] | ↑ leptin | PE | <0.05 |
| Hogg et al. [36] | ↑ leptin | PE | <0.05 |
| Hogg et al. [36] | ↑ leptin | GDM | <0.0001 |
| Schoots et al. [25] | ↑ leptin | PE | <0.05 |
| Shang et al. [27] | ↓ adiponectin | GDM | <0.05 |
| Balachandiran et al. [32] | ↓ adiponectin | GDM | <0.05 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Downs, T.; da Silva Costa, F.; de Freitas Paganoti, C.; Holland, O.J.; Hryciw, D.H. Adiponectin and Leptin during Pregnancy: A Systematic Review of Their Association with Pregnancy Disorders, Fetal Growth and Placental Function. Endocrines 2024, 5, 382-394. https://doi.org/10.3390/endocrines5030028
Downs T, da Silva Costa F, de Freitas Paganoti C, Holland OJ, Hryciw DH. Adiponectin and Leptin during Pregnancy: A Systematic Review of Their Association with Pregnancy Disorders, Fetal Growth and Placental Function. Endocrines. 2024; 5(3):382-394. https://doi.org/10.3390/endocrines5030028
Chicago/Turabian StyleDowns, Taylor, Fabricio da Silva Costa, Cristiane de Freitas Paganoti, Olivia J. Holland, and Deanne H. Hryciw. 2024. "Adiponectin and Leptin during Pregnancy: A Systematic Review of Their Association with Pregnancy Disorders, Fetal Growth and Placental Function" Endocrines 5, no. 3: 382-394. https://doi.org/10.3390/endocrines5030028
APA StyleDowns, T., da Silva Costa, F., de Freitas Paganoti, C., Holland, O. J., & Hryciw, D. H. (2024). Adiponectin and Leptin during Pregnancy: A Systematic Review of Their Association with Pregnancy Disorders, Fetal Growth and Placental Function. Endocrines, 5(3), 382-394. https://doi.org/10.3390/endocrines5030028