
The Influence of Burst-Firing EMF on Forskolin-Induced Pheochromocytoma (PC12) Plasma Membrane Extensions



Abstract
1. Introduction
2. Methods
2.1. Cell Culture and Induction of Plasma Membrane Extensions
2.2. Exposure Apparatus
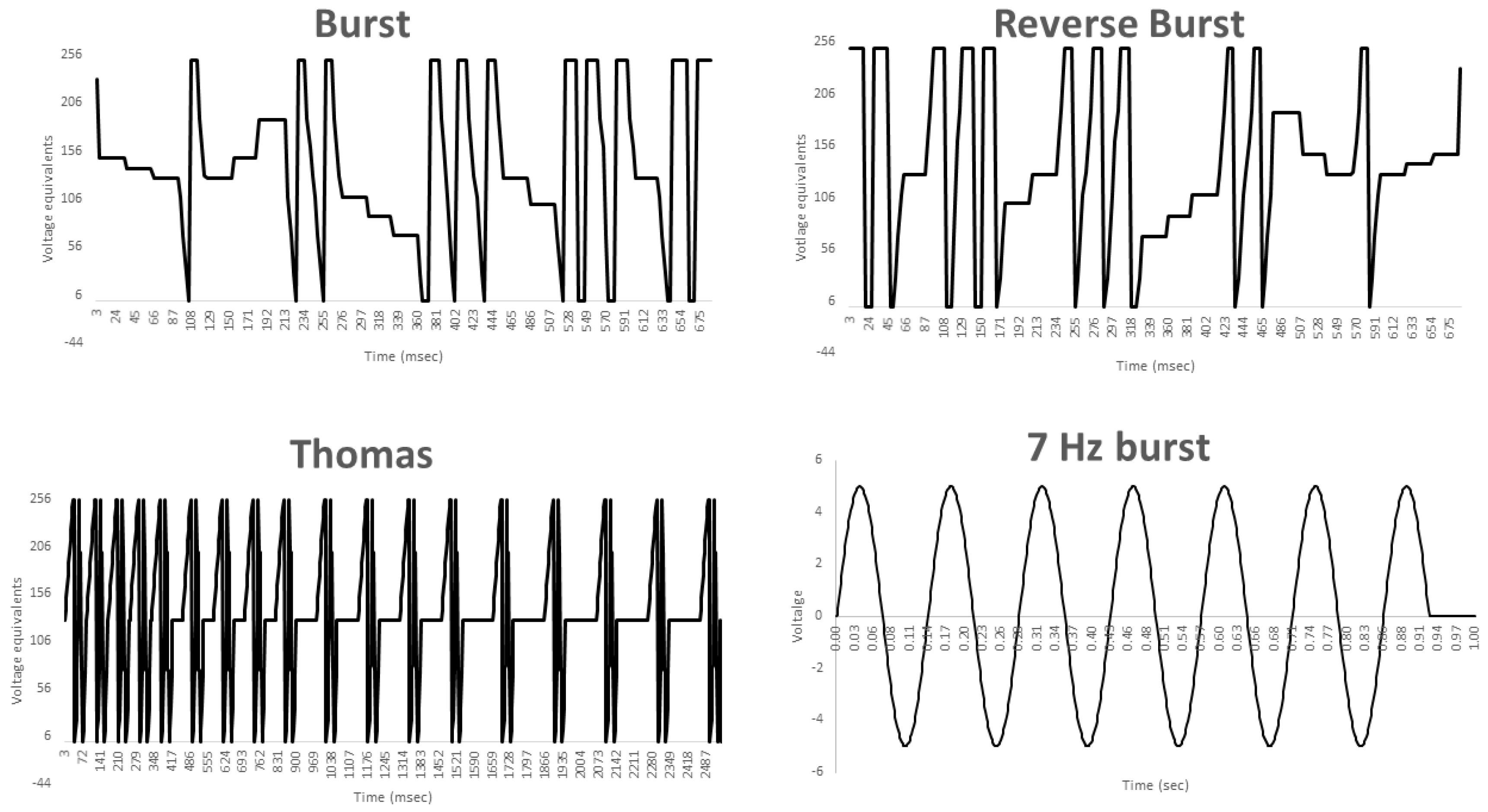
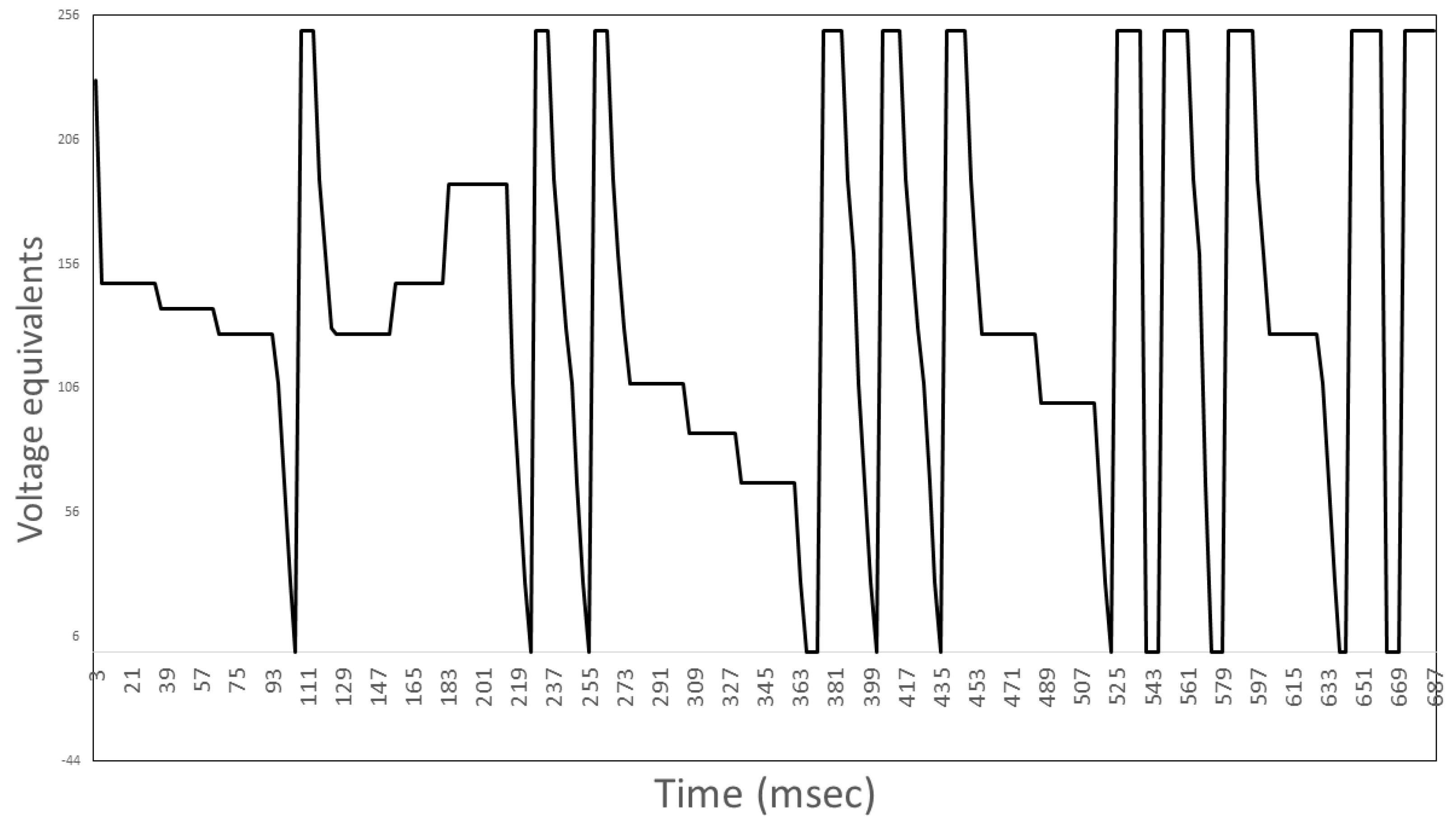
2.3. Exposure Protocols
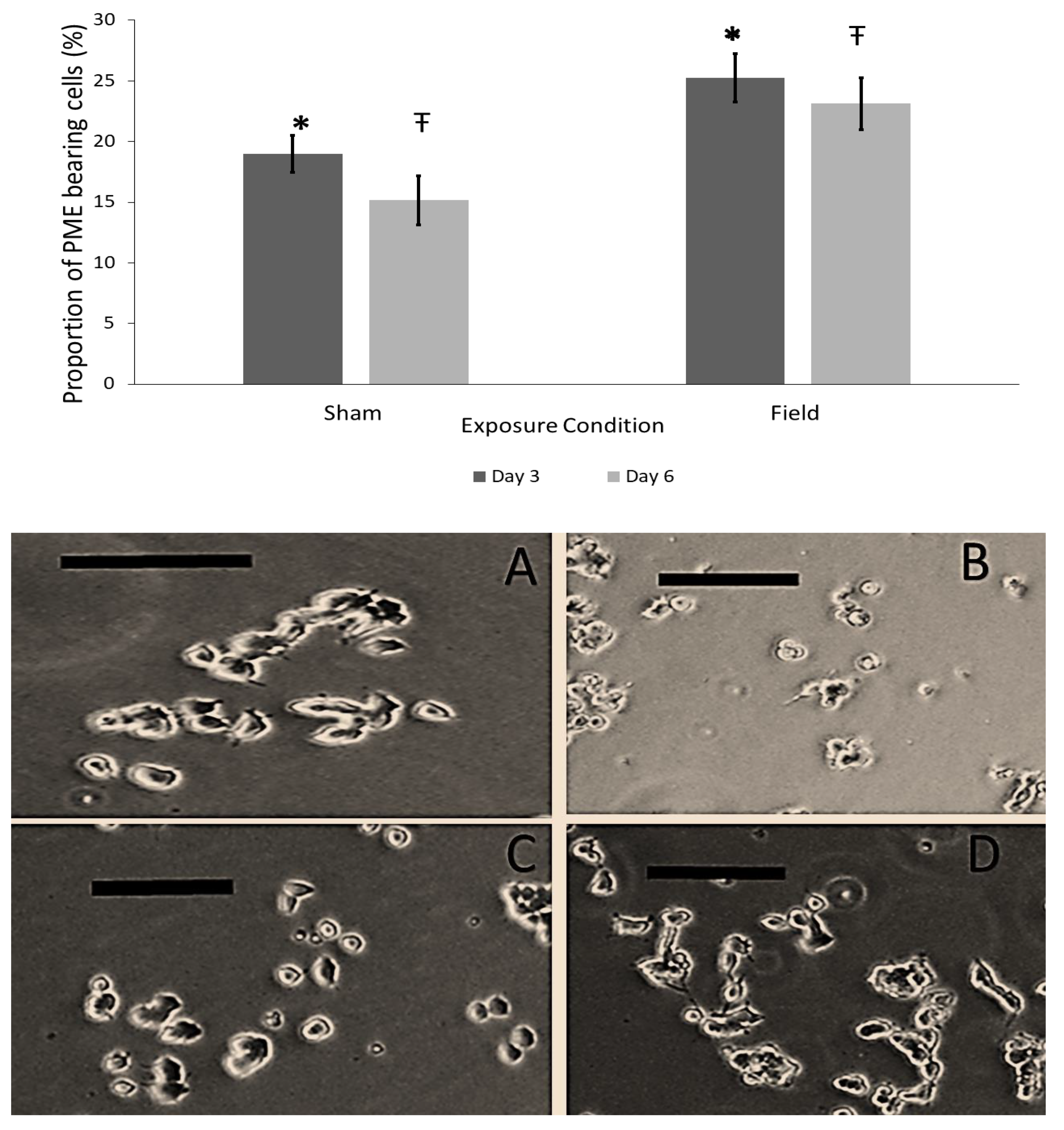
2.4. Experiment #1–Exposure to Burst-Firing EMF for 1 and 3 h
2.5. Experiment #2–Exposure to Different Patterned EMF
2.6. Experiment #3–Identifying the Influence of Burst-Firing EMF Timing on PC12 PME
2.7. Experiment #4–Investigating the Role of Intensity of Burst-Firing EMF on PC12 Cell PME
2.8. Experiment #5–Examining Differences in the Proportion of PC12 PME between Changing and Constant EMF Exposure
2.9. Cell Morphological Measures
2.10. Statistical Methods
3. Results
3.1. Descriptive Statistics
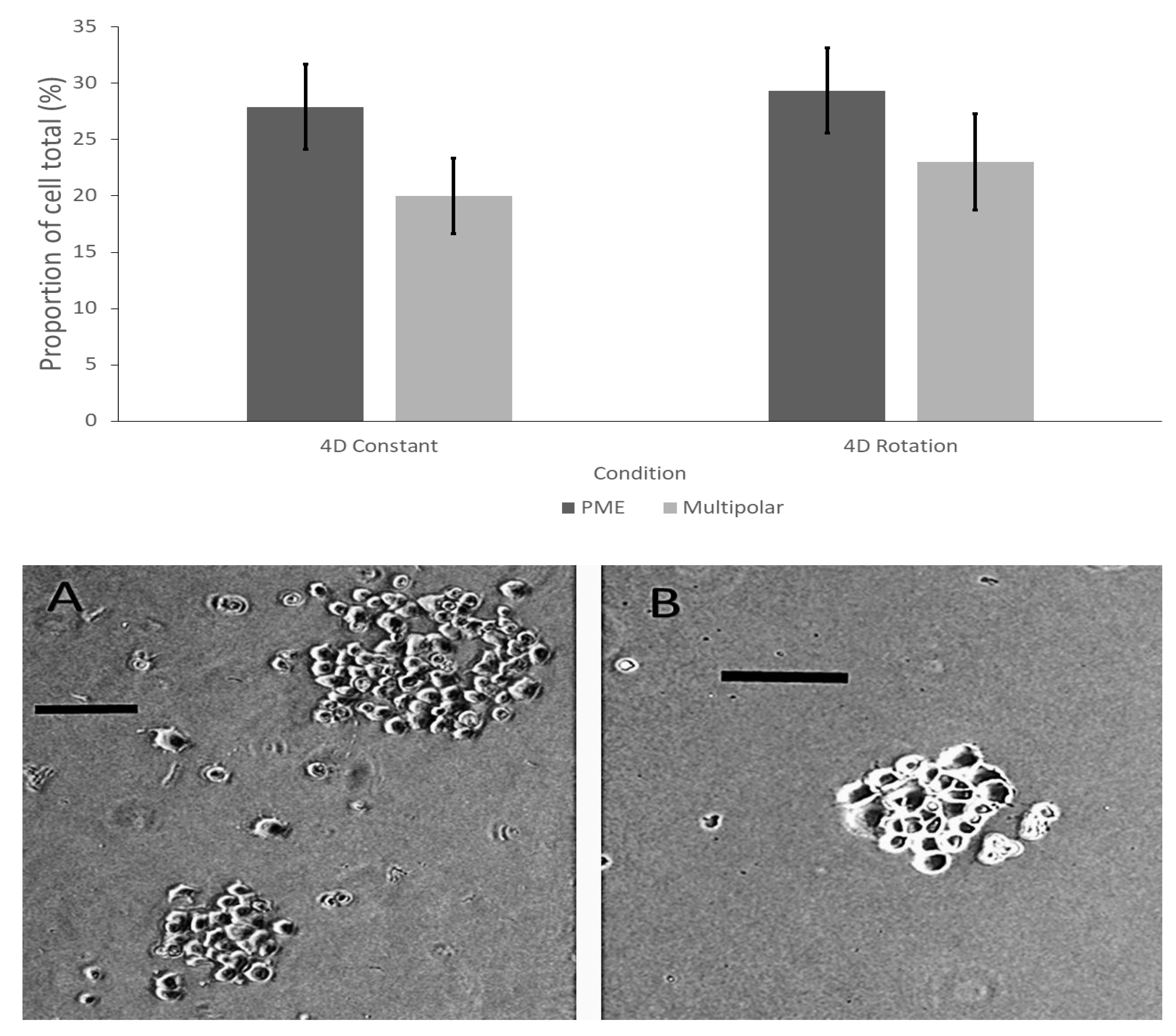
3.2. Results of Experiment #5–Determining the Difference between Rotational or Constant Application of Burst-Firing EMF Generated through the 4-D Exposure Application
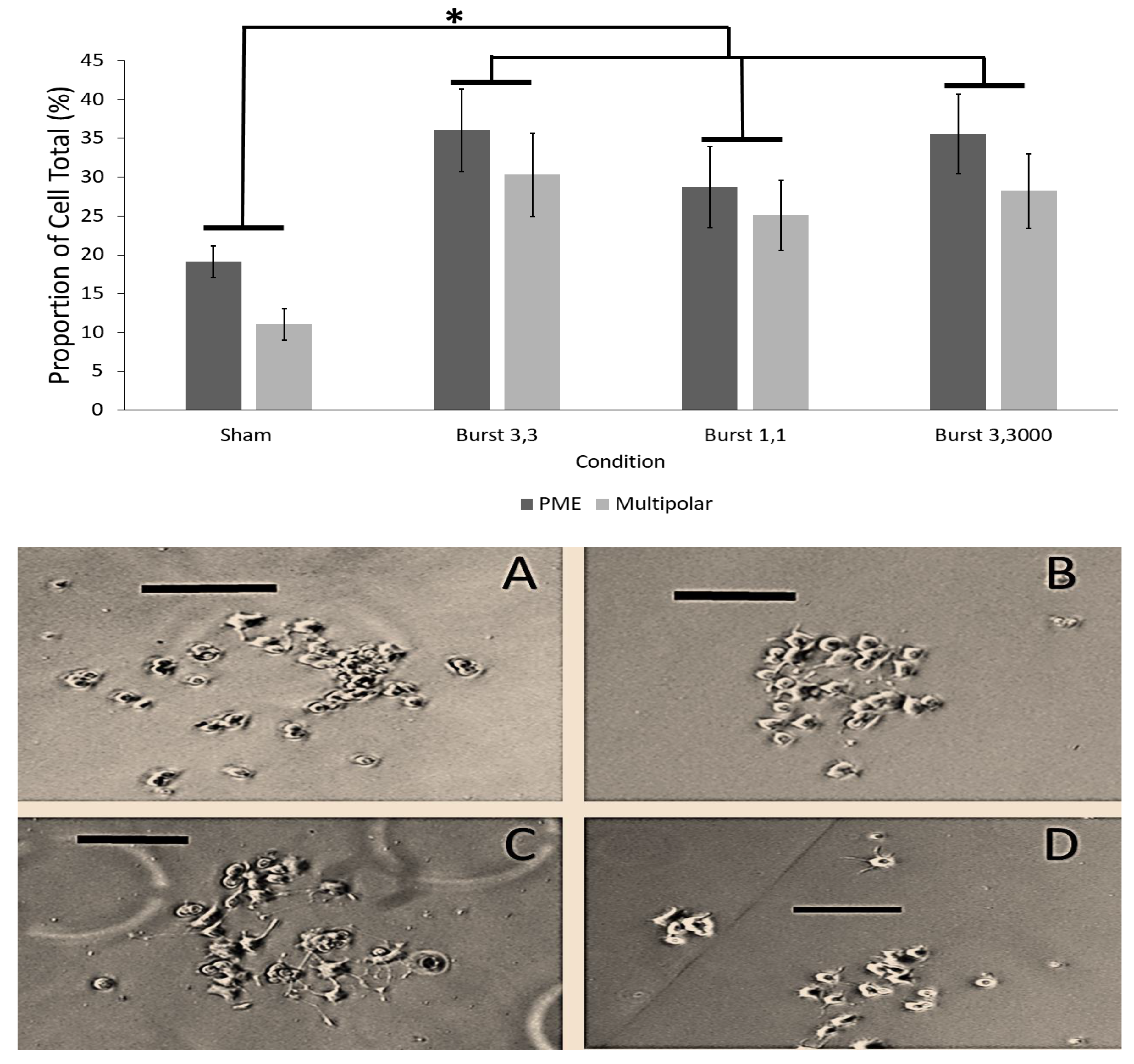
3.3. Results of Experiment #3–The Influence of Point Duration of Burst-Firing EMF on PC12 Plasma Membrane Extensions
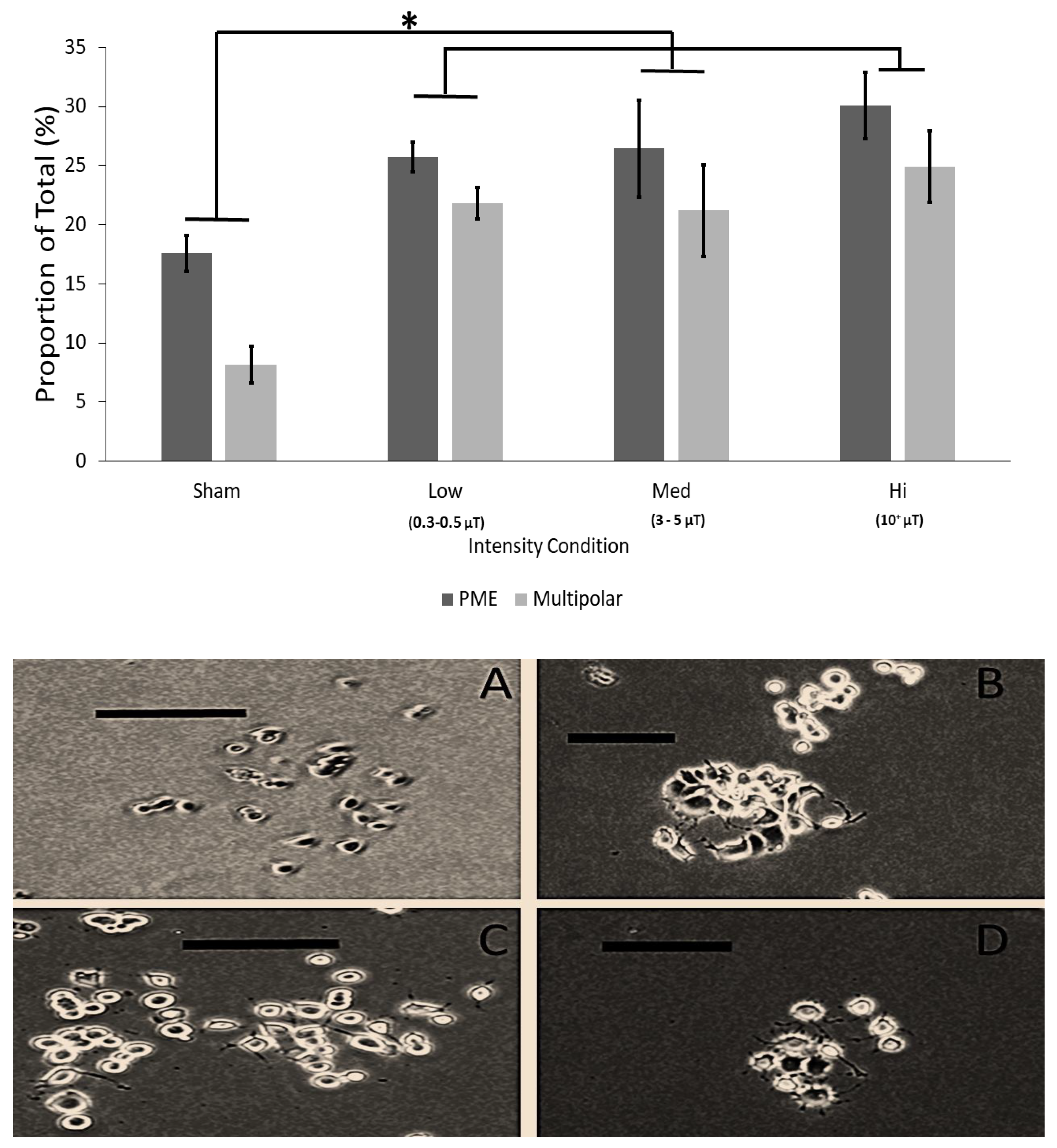
3.4. Results of Experiment #4–Effect of Intensity of Burst-Firing EMF on Plasma Membrane Extensions in PC12 Cultures
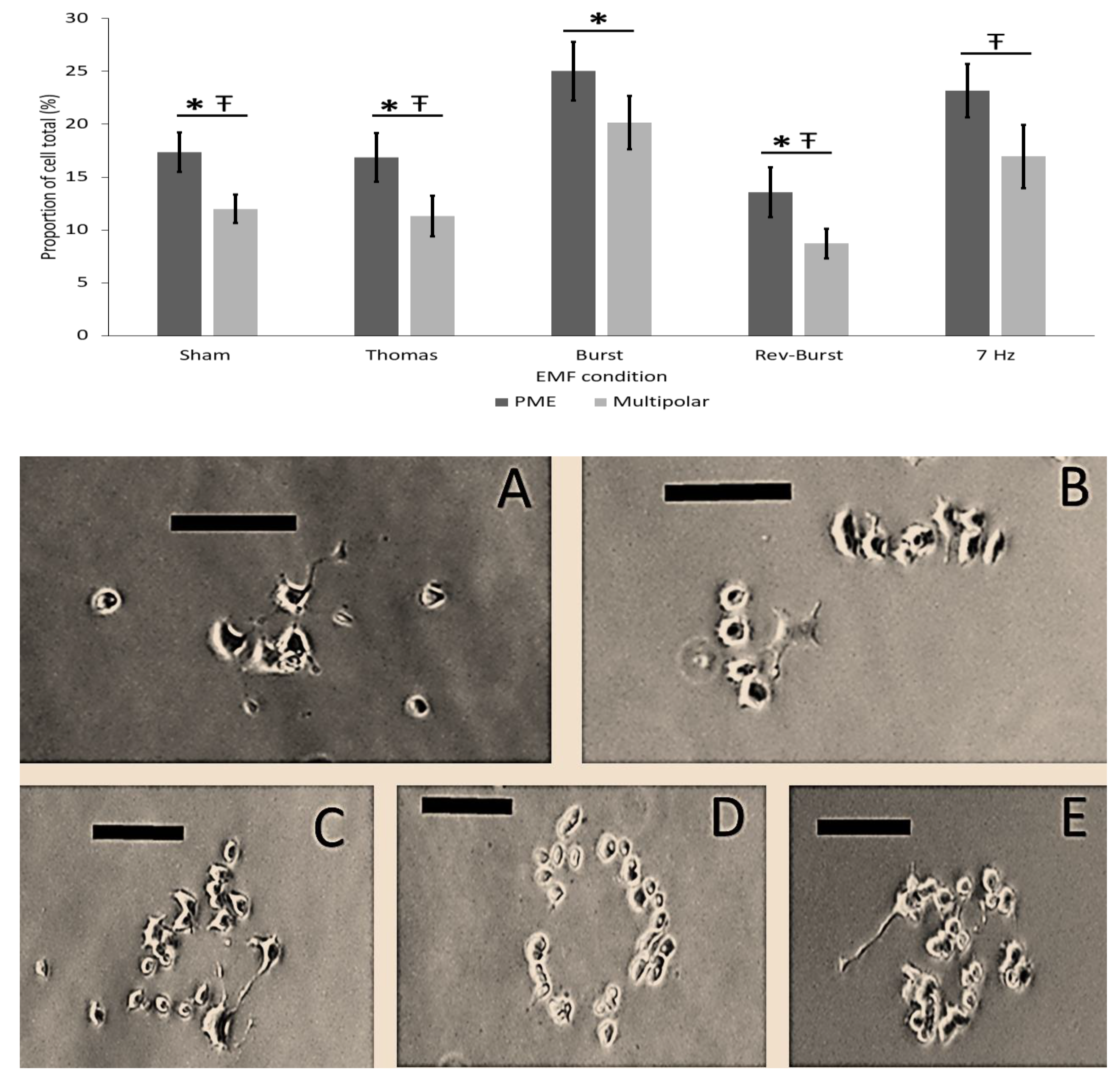
3.5. Results of Experiment # 2–Pattern Specificity of EMF Application on Forskolin Induced PC12 Plasma Membrane Extensions
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A




References
- Greene, L.A.; Tischler, A.S. PC12 Pheochromocytoma Cultures in Neurobiological Research. In Advances in Cellular Neurobiology; Elsevier: Amsterdam, The Netherlands, 1982; Volume 3, pp. 373–414. [Google Scholar]
- Greene, L.A.; Tischler, A. Establishment of a noradrenergic clonal line of rat adrenal pheochromocytoma cells which respond to nerve growth factor. Proc. Natl. Acad. Sci. USA 1976, 73, 2424–2428. [Google Scholar] [CrossRef] [PubMed]
- Tischler, A.S.; A Greene, L. Morphologic and cytochemical properties of a clonal line of rat adrenal pheochromocytoma cells which respond to nerve growth factor. Lab. Investig. 1978, 39, 77–89. [Google Scholar]
- Tischler, A.; Greene, L.; Kwan, P.; Slayton, V. Ultrastructural effects of nerve growth factor on PC 12 pheochromocytoma cells in spinner culture. Cell Tissue Res. 1983, 228, 641–648. [Google Scholar] [CrossRef] [PubMed]
- Satpute, R.; Hariharakrishnan, J.; Bhattacharya, R. Effect of alpha-ketoglutarate andN-acetyl cysteine on cyanide-induced oxidative stress mediated cell death in PC12 cells. Toxicol. Ind. Heal. 2010, 26, 297–308. [Google Scholar] [CrossRef]
- Shafer, T.; Atchison, W.D. Transmitter, ion channel and receptor properties of pheochromocytoma (PC12) cells: A model for neurotoxicological studies. NeuroToxicology 1991, 12, 473–492. [Google Scholar] [PubMed]
- Caillaud, T.; Opstal, W.Y.X.-V.; Scarcériaux, V.; Billardon, C.; Rostène, W. Treatment of PC12 cells by nerve growth factor, dexamethasone, and forskolin. Mol. Neurobiol. 1995, 10, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Gunning, P.W.; E Landreth, G.; A Bothwell, M.; Shooter, E.M. Differential and synergistic actions of nerve growth factor and cyclic AMP in PC12 cells. J. Cell Biol. 1981, 89, 240–245. [Google Scholar] [CrossRef]
- Loeb, D.M.; Maragos, J.; Martin-Zanca, D.; Chao, M.; Parada, L.F.; Greene, L.A. The trk proto-oncogene rescues NGF responsiveness in mutant NGF-nonresponsive PC12 cell lines. Cell 1991, 66, 961–966. [Google Scholar] [CrossRef]
- Vaudry, D.; Stork, P.J.S.; Lazarovici, P.; Eiden, L.E. Signaling Pathways for PC12 Cell Differentiation: Making the Right Connections. Science 2002, 296, 1648–1649. [Google Scholar] [CrossRef] [PubMed]
- Bos, J.; De Bruyn, K.; Enserink, J.; Kuiperij, B.; Rangarajan, S.; Rehmann, H.; Riedl, J.; De Rooij, J.; Van Mansfeld, F.; Zwartkruis, F. The role of Rap1 in integrin-mediated cell adhesion. Biochem. Soc. Trans. 2003, 31, 83–86. [Google Scholar] [CrossRef]
- Greene, L.A.; McGuire, J.C. Induction of ornithine decarboxylase by nerve growth factor dissociated from effects on survival and neurite outgrowth. Nat. Cell Biol. 1978, 276, 191–194. [Google Scholar] [CrossRef] [PubMed]
- Park, K.H.; Park, H.J.; Shin, K.S.; Choi, H.S.; Kai, M.; Lee, M.K. Modulation of PC12 Cell Viability by Forskolin-Induced Cyclic AMP Levels Through ERK and JNK Pathways: An Implication for L-DOPA-Induced Cytotoxicity in Nigrostriatal Dopamine Neurons. Toxicol. Sci. 2012, 128, 247–257. [Google Scholar] [CrossRef]
- York, R.D.; Yao, H.; Dillon, T.; Ellig, C.L.; Eckert, S.P.; McCleskey, E.W.; Stork, P.J.S. Rap1 mediates sustained MAP kinase activation induced by nerve growth factor. Nat. Cell Biol. 1998, 392, 622–626. [Google Scholar] [CrossRef]
- Wang, Z.; Dillon, T.J.; Pokala, V.; Mishra, S.; Labudda, K.; Hunter, B.; Stork, P.J.S. Rap1-Mediated Activation of Extracellular Signal-Regulated Kinases by Cyclic AMP Is Dependent on the Mode of Rap1 Activation. Mol. Cell. Biol. 2006, 26, 2130–2145. [Google Scholar] [CrossRef]
- Park, T.; Kim, K.-T. Cyclic AMP-Independent Inhibition of Voltage-Sensitive Calcium Channels by Forskolin in PC12 Cells. J. Neurochem. 2002, 66, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Richter-Landsberg, C.; Jastorff, B. The role of cAMP in nerve growth factor-promoted neurite outgrowth in PC12 cells. J. Cell Biol. 1986, 102, 821–829. [Google Scholar] [CrossRef]
- Seamon, K.B.; Daly, J.W. Forskolin: A unique diterpene activator of cyclic AMP-generating systems. J. Cycl. Nucleotide Res. 1981, 7, 201–224. [Google Scholar]
- Seamon, K.B.; Daly, J.W. Forskolin, cyclic AMP and cellular physiology. Trends Pharmacol. Sci. 1983, 4, 120–123. [Google Scholar] [CrossRef]
- Seamon, K.B.; Padgett, W.; Daly, J.W. Forskolin: Unique diterpene activator of adenylate cyclase in membranes and in intact cells. Proc. Natl. Acad. Sci. USA 1981, 78, 3363–3367. [Google Scholar] [CrossRef]
- Zhang, K.; Duan, L.; Ong, Q.; Lin, Z.; Varman, P.M.; Sung, K.; Cui, B. Light-Mediated Kinetic Control Reveals the Temporal Effect of the Raf/MEK/ERK Pathway in PC12 Cell Neurite Outgrowth. PLoS ONE 2014, 9, e92917. [Google Scholar] [CrossRef]
- Cook, C.M.; Persinger, M.A. Experimental Induction of the “Sensed Presence” in Normal Subjects and an Exceptional Subject. Percept. Mot. Ski. 1997, 85, 683–693. [Google Scholar] [CrossRef]
- Karbowski, L.M.; Harribance, S.L.; Buckner, C.A.; Mulligan, B.P.; Koren, S.A.; Lafrenie, R.M.; Persinger, M.A. Digitized quantitative electroencephalographic patterns applied as magnetic fields inhibit melanoma cell proliferation in culture. Neurosci. Lett. 2012, 523, 131–134. [Google Scholar] [CrossRef]
- Mach, Q.H.; Persinger, M.A. Behavioral changes with brief exposures to weak magnetic fields patterned to stimulate long-term potentiation. Brain Res. 2009, 1261, 45–53. [Google Scholar] [CrossRef]
- Meli, S.C.; Persinger, M.A. Red Light Facilitates the Sensed Presence Elicited by Application of Weak, Burst-Firing Magnetic Fields Over the Temporal Lobes. Int. J. Neurosci. 2009, 119, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Murugan, N.J.; Karbowski, L.M.; Lafrenie, R.; Persinger, M.A. Maintained Exposure to Spring Water but Not Double Distilled Water in Darkness and Thixotropic Conditions to Weak (~1 µT) Temporally Patterned Magnetic Fields Shift Photon Spectroscopic Wavelengths: Effects of Different Shielding Materials. J. Biophys. Chem. 2015, 6, 14–28. [Google Scholar] [CrossRef]
- Murugan, N.J.; Karbowski, L.M.; Persinger, M.A. Serial pH increments (~20 to 40 milliseconds) in water during exposures to weak, physiologically patterned magnetic fields: Implications for consciousness. Water 2014, 6, 45–60. [Google Scholar]
- Murugan, N.J.; Karbowski, L.M.; Persinger, M.A. Weak Burst-firing magnetic fields that produce analgesia equivalent to morphine do not initiate activation of proliferation pathways in human breast cells in culture. Integr. Cancer Sci. Ther. 2014, 1, 47–50. [Google Scholar]
- Ross, M.L.; Koren, S.A.; Persinger, M.A. Physiologically Patterned Weak Magnetic Fields Applied Over Left Frontal Lobe Increase Acceptance of False Statements as True. Electromagn. Biol. Med. 2008, 27, 365–371. [Google Scholar] [CrossRef] [PubMed]
- Whissell, P.D.; Persinger, M.A. Emerging synergisms between drugs and physiologically-patterned weak magnetic fields: Implications for neuropharmacology and the human population in the twenty-first century. Curr. Neuropharmacol. 2007, 5, 278–288. [Google Scholar] [CrossRef]
- Buckner, C.A.; Buckner, A.L.; Koren, S.A.; Persinger, M.A.; Lafrenie, R.M. Inhibition of Cancer Cell Growth by Exposure to a Specific Time-Varying Electromagnetic Field Involves T-Type Calcium Channels. PLoS ONE 2015, 10, e0124136. [Google Scholar] [CrossRef]
- Buckner, C.A.; Buckner, A.L.; Koren, S.A.; Persinger, M.A.; Lafrenie, R.M. The effects of electromagnetic fields on B16-BL6 cells are dependent on their spatial and temporal character. Bioelectromagnetics 2017, 38, 165–174. [Google Scholar] [CrossRef]
- Buckner, C.A.; Buckner, A.L.; Koren, S.A.; Persinger, M.A.; Lafrenie, R.M. Exposure to a specific time-varying electromagnetic field inhibits cell proliferation via cAMP and ERK signaling in cancer cells. Bioelectromagnetics 2018, 39, 217–230. [Google Scholar] [CrossRef]
- Dotta, B.T.; Buckner, C.A.; Lafrenie, R.M.; Persinger, M.A. Photon emissions from human brain and cell culture exposed to distally rotating magnetic fields shared by separate light-stimulated brains and cells. Brain Res. 2011, 1388, 77–88. [Google Scholar] [CrossRef]
- Dotta, B.T.; Lafrenie, R.M.; Karbowski, L.M.; Persinger, M.A. Photon emission from melanoma cells during brief stimulation by patterned magnetic fields: Is the source coupled to rotational diffusion within the membrane? Gen. Physiol. Biophys. 2014, 33, 63–73. [Google Scholar] [CrossRef][Green Version]
- Dotta, B.T.; Murugan, N.J.; Karbowski, L.M.; Koren, S.A. Rotational Frequency Matching of the Energy of the Changing Angular Velocity Magnetic Field Intensity and the Proton Magnetic Moment Produces a Ten Fold Increased Excess Correlation in pH Shifts in Spring Water. NeuroQuantology 2015, 14. [Google Scholar] [CrossRef]
- Dotta, B.T.; Persinger, M.A. “Doubling” of local photon emissions when two simultaneous, spatially-separated, chemiluminescent reactions share the same magnetic field configurations. J. Biophys. Chem. 2012, 03, 72–80. [Google Scholar] [CrossRef]
- Murugan, N.J.; Karbowski, L.M.; Lafrenie, R.M.; Persinger, M.A. Temporally-Patterned Magnetic Fields Induce Complete Fragmentation in Planaria. PLoS ONE 2013, 8, e61714. [Google Scholar] [CrossRef] [PubMed]
- Tessaro, L.W.; Persinger, M.A. Optimal durations of single exposures to a frequency-modulated magnetic field immediately after bisection in planarian predict final growth values. Bioelectromagnetics 2013, 34, 613–617. [Google Scholar] [CrossRef] [PubMed]
- Martin, L.; Koren, S.A.; Persinger, M.A. Thermal Analgesic Effects from Weak, Complex Magnetic Fields: Critical Parameters. Electromagn. Biol. Med. 2005, 24, 65–85. [Google Scholar] [CrossRef]
- Martin, L.; Persinger, M.A. Spatial Heterogeneity Not Homogeneity of the Magnetic Field during Exposures to Complex Frequency-Modulated Patterns Facilitates Analgesia. Percept. Mot. Ski. 2003, 96, 1005–1012. [Google Scholar] [CrossRef] [PubMed]
- Martin, L.; Persinger, M. Thermal analgesia induced by 30-min exposure to 1 μT burst-firing magnetic fields is strongly enhanced in a dose-dependent manner by the α2 agonist clonidine in rats. Neurosci. Lett. 2004, 366, 226–229. [Google Scholar] [CrossRef] [PubMed]
- Martin, L.; Persinger, M.A. The Influence of Various Pharmacological Agents on the Analgesia Induced by an Applied Complex Magnetic Field Treatment: A Receptor System Potpourri. Electromagn. Biol. Med. 2005, 24, 87–97. [Google Scholar] [CrossRef]
- Fleming, J.L.; Persinger, M.A.; Koren, S.A. Magnetic Pulses Elevate Nociceptive Thresholds: Comparisons with Opiate Receptor Compounds in Normal and Seizure-Induced Brain-Damaged Rats. Electro-Magnetobiology 1994, 13, 67–75. [Google Scholar] [CrossRef]
- Baker-Price, L.; Persinger, M.A. Intermittent Burst-firing weak (1 microTesla) magnetic fields reduce psychometric depression in patients who sustained closed head injuries: A replication and electroencephalographic validation. Percept. Mot. Ski. 2003, 96, 965–974. [Google Scholar] [CrossRef] [PubMed]
- Tsang, E.W.; Koren, S.A.; Persinger, M.A. Specific Patterns of Weak (1 microTesla) Transcerebral Complex Magnetic Fields Differentially Affect Depression, Fatigue, and Confusion in Normal Volunteers. Electromagn. Biol. Med. 2009, 28, 365–373. [Google Scholar] [CrossRef]
- Thompson, M.E.; Zimmer, W.E.; Wear, L.B.; MacMillan, L.A.; Thompson, W.; Huttner, W.B.; Hidaka, H.; Scammell, J.G. Differential regulation of chromogranin B/secretogranin I and secretogranin II by forskolin in PC12 cells. Mol. Brain Res. 1992, 12, 195–202. [Google Scholar] [CrossRef]
- Hussain, S.M.; Javorina, A.K.; Schrand, A.M.; Duhart, H.M.; Ali, S.F.; Schlager, J.J. The Interaction of Manganese Nanoparticles with PC-12 Cells Induces Dopamine Depletion. Toxicol. Sci. 2006, 92, 456–463. [Google Scholar] [CrossRef]
- Pothos, E.; Przedborski, S.; Davila, V.; Schmitz, Y.; Sulzer, D. D2-Like Dopamine Autoreceptor Activation Reduces Quantal Size in PC12 Cells. J. Neurosci. 1998, 18, 5575–5585. [Google Scholar] [CrossRef]
- Westerink, R.H.S.; Ewing, A.G. The PC12 cell as model for neurosecretion. Acta Physiol. 2008, 192, 273–285. [Google Scholar] [CrossRef]
- Calon, F.; Tahar, A.H.; Blanchet, P.J.; Morissette, M.; Grondin, R.; Goulet, M.; Doucet, J.-P.; Robertson, G.S.; Nestler, E.; Di Paolo, T.; et al. Dopamine-receptor stimulation: Biobehavioral and biochemical consequences. Trends Neurosci. 2000, 23, S92–S100. [Google Scholar] [CrossRef]
- Greengard, P.; Allen, P.B.; Nairn, A.C. Beyond the Dopamine Receptor: The DARPP-32/Protein Phosphatase-1 Cascade. Neuron 1999, 23, 435–447. [Google Scholar] [CrossRef]
- Thompson, A.C.; Zapata, A.; Justice, J.B.; Vaughan, R.A.; Sharpe, L.G.; Shippenberg, T.S. κ-Opioid Receptor Activation Modifies Dopamine Uptake in the Nucleus Accumbens and Opposes the Effects of Cocaine. J. Neurosci. 2000, 20, 9333–9340. [Google Scholar] [CrossRef] [PubMed]
- Fournier, N.M.; Mach, Q.H.; Whissell, P.D.; Persinger, M.A. Neurodevelopmental anomalies of the hippocampus in rats exposed to weak intensity complex magnetic fields throughout gestation. Int. J. Dev. Neurosci. 2012, 30, 427–433. [Google Scholar] [CrossRef] [PubMed]
- Friedman, H.; Becker, R.O.; Bachman, C.H. Psychiatric Ward Behaviour and Geophysical Parameters. Nat. Cell Biol. 1965, 205, 1050–1052. [Google Scholar] [CrossRef]
- Persinger, M.A. Wars and increased solar-geomagnetic activity: Aggression or change in intraspecies dominance? Percept. Mot. Ski. 1999, 88 (Suppl. S3), 1351–1355. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiment | Condition | Average Cell Total | Number of Replicates | Total Number of Plates Sampled | Average Cell Density (Cells/cm2) | Average PME | Average PME Density (Cells/cm2) |
|---|---|---|---|---|---|---|---|
| Experiment #1 Exposure Duration | Sham 1 h | 628.5 (175.64) | 3 | 6 | 4.76 × 104 | 105.5 (32.35) | 7.99 × 103 |
| Sham 3 h | 613.00 (180.92) | 3 | 6 | 4.64 × 104 | 101.33 (28.98) | 7.67 × 103 | |
| Burst 1 h | 421.5 (160.82) | 3 | 6 | 3.19 × 104 | 113.67 (47.92) | 8.61 × 103 | |
| Burst 3 h | 392.17 (166.22) | 3 | 6 | 2.97 × 104 | 103.77 (42.36) | 7.86 × 103 | |
| Experiment #2 Rotation vs. Constant | Rotation | 163.94 (57.96) | 4 | 8 | 1.24 × 104 | 71.75 (20.61) | 5.43 × 103 |
| Constant | 222.13 (60.31) | 4 | 8 | 1.68 × 104 | 68.00 (22.06) | 5.13 × 103 | |
| Experiment #3 Point Duration | Sham | 594.75 (78.30) | 12 | 24 | 4.50 × 104 | 107.00 (14.79) | 8.10 × 103 |
| Burst (3,3) | 348.5 (132.73) | 3 | 6 | 2.64 × 104 | 107.17 (33.26) | 8.11 × 103 | |
| Burst (1,1) | 298.33 (166.64) | 3 | 6 | 2.26 × 104 | 91.83 (49.61) | 6.96 × 103 | |
| Burst (33,000) | 159.00 (23.39) | 3 | 6 | 1.20 × 104 | 53.83 (7.61) | 4.08 × 103 | |
| Experiment #4 Intensity | Sham | 690.75 (112.35) | 6 | 12 | 5.23 × 104 | 119.54 (21.98) | 9.06 × 103 |
| Low | 465.17 (27.16) | 3 | 6 | 3.52 × 104 | 120.50 (10.83) | 9.13 × 103 | |
| Med | 314.5 (89.31) | 3 | 6 | 2.38 × 104 | 85.17 (26.87) | 6.45 × 103 | |
| Hi | 310.67 (43.25) | 3 | 6 | 2.35 × 104 | 88.00 (4.49) | 6.67 × 103 | |
| Experiment #5 EMF Pattern | Sham | 594.75 (78.30) | 12 | 24 | 4.51 × 104 | 107.42 (14.79) | 8.14 × 103 |
| Thomas | 846.17 (166.88) | 3 | 6 | 6.41 × 104 | 52.03 (21.24) | 3.94 × 103 | |
| Burst | 212.67 (54.73) | 3 | 6 | 1.61 × 104 | 47.00 (10.89) | 3.26 × 103 | |
| Rev-Burst | 251.17 (57.28) | 3 | 6 | 1.90 × 104 | 33.33 (7.07) | 2.53 × 103 | |
| 7-Hz | 304.84 (81.01) | 3 | 6 | 2.31 × 104 | 63.29 (25.84) | 4.79 × 103 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carniello, T.N.; Lafrenie, R.M.; Dotta, B.T. The Influence of Burst-Firing EMF on Forskolin-Induced Pheochromocytoma (PC12) Plasma Membrane Extensions. NeuroSci 2021, 2, 383-399. https://doi.org/10.3390/neurosci2040028
Carniello TN, Lafrenie RM, Dotta BT. The Influence of Burst-Firing EMF on Forskolin-Induced Pheochromocytoma (PC12) Plasma Membrane Extensions. NeuroSci. 2021; 2(4):383-399. https://doi.org/10.3390/neurosci2040028
Chicago/Turabian StyleCarniello, Trevor N., Robert M. Lafrenie, and Blake T. Dotta. 2021. "The Influence of Burst-Firing EMF on Forskolin-Induced Pheochromocytoma (PC12) Plasma Membrane Extensions" NeuroSci 2, no. 4: 383-399. https://doi.org/10.3390/neurosci2040028
APA StyleCarniello, T. N., Lafrenie, R. M., & Dotta, B. T. (2021). The Influence of Burst-Firing EMF on Forskolin-Induced Pheochromocytoma (PC12) Plasma Membrane Extensions. NeuroSci, 2(4), 383-399. https://doi.org/10.3390/neurosci2040028