
Assessing the Impact of Agents with Antiviral Activities on Transmembrane Ionic Currents: Exploring Possible Unintended Actions

 , and
, and
Abstract
:1. Introduction
2. Summary
2.1. Artemisinin (ART)
2.2. Cannabidiol (CBD)
2.3. Memantine (MEM, Namenda®)
2.4. Mitoxantrone (MX, Novantrone®)
2.5. Molnupiravir (MOL, EIDD-2801, MK-4482, Lagevrio®)
2.6. Remdesivir (RDV, GS-5734)
2.7. SM-102
2.8. Sorafenib (SOR, Nexavar®)
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Ih | hyperpolarization-activated cation current |
| IK(erg) | erg-mediated K+ current |
| IK(IR) | inwardly rectifying K+ current |
| IK(M) | M-type K+ current |
| IK(S) | slowly activating delayed-rectifier K+ current |
| IMEP | MEP-induced inward current |
| INa | voltage-gated Na+ current |
| Kir channel | inwardly rectifying K+ channel |
| MEP | membrane electroporation |
| NaV channel | voltage-gated Na+ channel |
| QTc interval | corrected QT interval |
References
- Hong, C.M.; Liu, C.H.; Su, T.H.; Yang, H.C.; Chen, P.J.; Chen, Y.W.; Kao, J.H.; Liu, C.J. Real-world effectiveness of direct-acting antiviral agents for chronic hepatitis C in Taiwan: Real-world data. J. Microbiol. Immunol. Infect. 2020, 53, 569–577. [Google Scholar] [CrossRef] [PubMed]
- Panahi, Y.; Gorabi, A.M.; Talaei, S.; Beiraghdar, F.; Akbarzadeh, A.; Tarhriz, V.; Mellatyar, H. An overview on the treatments and prevention against COVID-19. Virol. J. 2023, 20, 23. [Google Scholar] [CrossRef] [PubMed]
- Ackerman, M.J.; Clapham, D.E. Ion channels—Basic science and clinical disease. N. Engl. J. Med. 1997, 336, 1575–1586. [Google Scholar] [CrossRef] [PubMed]
- Franciolini, F. Patch clamp technique and biophysical study of membrane channels. Experientia 1986, 42, 589–594. [Google Scholar] [CrossRef] [PubMed]
- Kausar, S.; Said Khan, F.; Ishaq Mujeeb Ur Rehman, M.; Akram, M.; Riaz, M.; Rasool, G.; Hamid Khan, A.; Saleem, I.; Shamim, S.; Malik, A. A review: Mechanism of action of antiviral drugs. Int. J. Immunopathol. Pharmacol. 2021, 35, 20587384211002621. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.S.; Lai, M.C.; Huang, H.I.; Wu, S.N.; Huang, C.W. Immunity, Ion Channels and Epilepsy. Int. J. Mol. Sci. 2022, 23, 6446. [Google Scholar] [CrossRef]
- Zhu, S.; Wang, J.; Wang, Y.; Chu, J.; Liu, Y.; Chen, X.; Chen, X. QTc prolongation during antiviral therapy in two COVID-19 patients. J. Clin. Pharm. Ther. 2020, 45, 1190–1193. [Google Scholar] [CrossRef] [PubMed]
- Santoro, F.; Monitillo, F.; Raimondo, P.; Lopizzo, A.; Brindicci, G.; Gilio, M.; Musaico, F.; Mazzola, M.; Vestito, D.; Benedetto, R.D.; et al. QTc Interval Prolongation and Life-Threatening Arrhythmias During Hospitalization in Patients with Coronavirus Disease 2019 (COVID-19): Results From a Multicenter Prospective Registry. Clin. Infect. Dis. 2021, 73, e4031–e4038. [Google Scholar] [CrossRef]
- Esmel-Vilomara, R.; Dolader, P.; Sabaté-Rotes, A.; Soriano-Arandes, A.; Gran, F.; Rosés-Noguer, F. QTc interval prolongation in patients infected with SARS-CoV-2 and treated with antiviral drugs. An. Pediatr. (Engl. Ed.) 2022, 96, 213–220. [Google Scholar] [CrossRef]
- Hu, C.H.; Wu, S.N.; So, E.C. Tyrosine kinase inhibitors, ionic currents, and cardiac arrhythmia. Front. Oncol. 2023, 13, 1218821. [Google Scholar] [CrossRef]
- Qiao, G.; Li, S.; Yang, B.; Li, B. Inhibitory effects of artemisinin on voltage-gated ion channels in intact nodose ganglion neurones of adult rats. Basic Clin. Pharmacol. Toxicol. 2007, 100, 217–224. [Google Scholar] [CrossRef] [PubMed]
- So, E.C.; Wu, S.N.; Wu, P.C.; Chen, H.Z.; Yang, C.J. Synergistic Inhibition of Delayed Rectifier K+ and Voltage-Gated Na+ Currents by Artemisinin in Pituitary Tumor (GH3) Cells. Cell. Physiol. Biochem. 2017, 41, 2053–2066. [Google Scholar] [CrossRef]
- Zhang, D.; Atochina-Vasserman, E.N.; Lu, J.; Maurya, D.S.; Xiao, Q.; Liu, M.; Adamson, J.; Ona, N.; Reagan, E.K.; Ni, H.; et al. The Unexpected Importance of the Primary Structure of the Hydrophobic Part of One-Component Ionizable Amphiphilic Janus Dendrimers in Targeted mRNA Delivery Activity. J. Am. Chem. Soc. 2022, 144, 4746–4753. [Google Scholar] [CrossRef]
- Liu, Y.-C.; So, E.C.; Wu, S.-N. Cannabidiol Modulates M-Type K+ and Hyperpolarization-Activated Cation Currents. Biomedicines 2023, 11, 2651. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.N.; Huang, H.C.; Yeh, C.C.; Yang, W.H.; Lo, Y.C. Inhibitory effect of memantine, an NMDA-receptor antagonist, on electroporation-induced inward currents in pituitary GH3 cells. Biochem. Biophys. Res. Commun. 2011, 405, 508–513. [Google Scholar] [CrossRef] [PubMed]
- Tsai, K.L.; Chang, H.F.; Wu, S.N. The inhibition of inwardly rectifying K+ channels by memantine in macrophages and microglial cells. Cell. Physiol. Biochem. 2013, 31, 938–951. [Google Scholar] [CrossRef]
- Wang, C.L.; Tsai, M.L.; Wu, S.N. Evidence for mitoxantrone-induced block of inwardly rectifying K+ channels expressed in the osteoclast precursor RAW 264.7 cells differentiated with lipopolysaccharide. Cell. Physiol. Biochem. 2012, 30, 687–701. [Google Scholar] [CrossRef]
- Shiau, A.L.; Lee, K.H.; Cho, H.Y.; Chuang, T.H.; Yu, M.C.; Wu, C.L.; Wu, S.N. Molnupiravir, a ribonucleoside antiviral prodrug against SARS-CoV-2, alters the voltage-gated sodium current and causes adverse events. Virology 2023, 587, 109865. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.T.; Liu, P.Y.; Lee, K.; Feng, Y.H.; Wu, S.N. Differential Inhibitory Actions of Multitargeted Tyrosine Kinase Inhibitors on Different Ionic Current Types in Cardiomyocytes. Int. J. Mol. Sci. 2020, 21, 1672. [Google Scholar] [CrossRef]
- Amarh, E.; Tisdale, J.E.; Overholser, B.R. Prolonged Exposure to Remdesivir Inhibits the Human Ether-A-Go-Go-Related Gene Potassium Current. J. Cardiovasc. Pharmacol. 2023, 82, 212–220. [Google Scholar] [CrossRef]
- Cho, H.Y.; Chuang, T.H.; Wu, S.N. Effective Perturbations on the Amplitude and Hysteresis of Erg-Mediated Potassium Current Caused by 1-Octylnonyl 8-[(2-hydroxyethyl)[6-oxo-6(undecyloxy)hexyl]amino]-octanoate (SM-102), a Cationic Lipid. Biomedicines 2021, 9, 1367. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.N.; Yeh, C.C.; Wu, P.Y.; Huang, H.C.; Tsai, M.L. Investigations into the correlation properties of membrane electroporation-induced inward currents: Prediction of pore formation. Cell Biochem. Biophys. 2012, 62, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.T.; Liu, P.Y.; Gao, Z.H.; Lee, S.W.; Lee, W.K.; Wu, S.N. Evidence for the Effectiveness of Remdesivir (GS-5734), a Nucleoside-Analog Antiviral Drug in the Inhibition of I (K(M)) or I (K(DR)) and in the Stimulation of I (MEP). Front. Pharmacol. 2020, 11, 1091. [Google Scholar] [CrossRef] [PubMed]
- Li, Y. Qinghaosu (artemisinin): Chemistry and pharmacology. Acta Pharmacol. Sin. 2012, 33, 1141–1146. [Google Scholar] [CrossRef]
- Ho, W.E.; Peh, H.Y.; Chan, T.K.; Wong, W.S. Artemisinins: Pharmacological actions beyond anti-malarial. Pharmacol. Ther. 2014, 142, 126–139. [Google Scholar] [CrossRef]
- Fuzimoto, A.D. An overview of the anti-SARS-CoV-2 properties of Artemisia annua, its antiviral action, protein-associated mechanisms, and repurposing for COVID-19 treatment. J. Integr. Med. 2021, 19, 375–388. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, J.R.; Antunes, B.S.; do Nascimento, G.O.; Kawall, J.C.S.; Oliveira, J.V.B.; Silva, K.; Costa, M.A.T.; Oliveira, C.R. Antiviral activity of medicinal plant-derived products against SARS-CoV-2. Exp. Biol. Med. 2022, 247, 1797–1809. [Google Scholar] [CrossRef] [PubMed]
- Urban, N.; Schaefer, M. Direct Activation of TRPC3 Channels by the Antimalarial Agent Artemisinin. Cells 2020, 9, 202. [Google Scholar] [CrossRef] [PubMed]
- Dillard, L.K.; Fullerton, A.M.; McMahon, C.M. Ototoxic hearing loss from antimalarials: A systematic narrative review. Travel Med. Infect. Dis. 2021, 43, 102117. [Google Scholar] [CrossRef]
- Peng, T.; Li, S.; Liu, L.; Yang, C.; Farhan, M.; Chen, L.; Su, Q.; Zheng, W. Artemisinin attenuated ischemic stroke induced cell apoptosis through activation of ERK1/2/CREB/BCL-2 signaling pathway in vitro and in vivo. Int. J. Biol. Sci. 2022, 18, 4578–4594. [Google Scholar] [CrossRef]
- Fraguas-Sánchez, A.I.; Torres-Suárez, A.I. Medical Use of Cannabinoids. Drugs 2018, 78, 1665–1703. [Google Scholar] [CrossRef] [PubMed]
- van Breemen, R.B.; Muchiri, R.N.; Bates, T.A.; Weinstein, J.B.; Leier, H.C.; Farley, S.; Tafesse, F.G. Cannabinoids Block Cellular Entry of SARS-CoV-2 and the Emerging Variants. J. Nat. Prod. 2022, 85, 176–184. [Google Scholar] [CrossRef] [PubMed]
- Marquez, A.B.; Vicente, J.; Castro, E.; Vota, D.; Rodríguez-Varela, M.S.; Lanza Castronuovo, P.A.; Fuentes, G.M.; Parise, A.R.; Romorini, L.; Alvarez, D.E.; et al. Broad-Spectrum Antiviral Effect of Cannabidiol Against Enveloped and Nonenveloped Viruses. Cannabis Cannabinoid Res. 2023; ahead of print. [Google Scholar]
- Viudez-Martínez, A.; García-Gutiérrez, M.S.; Manzanares, J. Cannabidiol regulates the expression of hypothalamus-pituitary-adrenal axis-related genes in response to acute restraint stress. J. Psychopharmacol. 2018, 32, 1379–1384. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.W.; Lin, P.C.; Chen, J.L.; Lee, M.J. Cannabidiol Selectively Binds to the Voltage-Gated Sodium Channel Na(v)1.4 in Its Slow-Inactivated State and Inhibits Sodium Current. Biomedicines 2021, 9, 1141. [Google Scholar] [CrossRef]
- Sankaranarayanan, S.; Simasko, S.M. Characterization of an M-like current modulated by thyrotropin-releasing hormone in normal rat lactotrophs. J. Neurosci. 1996, 16, 1668–1678. [Google Scholar] [CrossRef] [PubMed]
- Varghese, N.; Moscoso, B.; Chavez, A.; Springer, K.; Ortiz, E.; Soh, H.; Santaniello, S.; Maheshwari, A.; Tzingounis, A.V. KCNQ2/3 Gain-of-Function Variants and Cell Excitability: Differential Effects in CA1 versus L2/3 Pyramidal Neurons. J. Neurosci. 2023, 43, 6479–6494. [Google Scholar] [CrossRef] [PubMed]
- Simasko, S.M.; Sankaranarayanan, S. Characterization of a hyperpolarization-activated cation current in rat pituitary cells. Am. J. Physiol. 1997, 272, E405–E414. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.S.; So, E.C.; Wu, S.N. Modulating Hyperpolarization-Activated Cation Currents through Small Molecule Perturbations: Magnitude and Gating Control. Biomedicines 2023, 11, 2177. [Google Scholar] [CrossRef] [PubMed]
- Arvanitakis, Z.; Shah, R.C.; Bennett, D.A. Diagnosis and Management of Dementia: Review. JAMA 2019, 322, 1589–1599. [Google Scholar] [CrossRef]
- Pereira, A.; Santos, I.A.; da Silva, W.W.; Nogueira, F.A.R.; Bergamini, F.R.G.; Jardim, A.C.G.; Corbi, P.P. Memantine hydrochloride: A drug to be repurposed against Chikungunya virus? Pharmacol. Rep. 2021, 73, 954–961. [Google Scholar] [CrossRef]
- Butterworth, R.F. Adamantanes for the treatment of neurodegenerative diseases in the presence of SARS-CoV-2. Front. Neurosci. 2023, 17, 1128157. [Google Scholar] [CrossRef] [PubMed]
- Müller, T.; Riederer, P.; Kuhn, W. Aminoadamantanes: From treatment of Parkinson’s and Alzheimer’s disease to symptom amelioration of long COVID-19 syndrome? Expert Rev. Clin. Pharmacol. 2023, 16, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Cheek, E.R.; Fast, V.G. Nonlinear changes of transmembrane potential during electrical shocks: Role of membrane electroporation. Circ. Res. 2004, 94, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Dyachok, O.; Zhabyeyev, P.; McDonald, T.F. Electroporation-induced inward current in voltage-clamped guinea pig ventricular myocytes. J. Membr. Biol. 2010, 238, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Hibino, H.; Inanobe, A.; Furutani, K.; Murakami, S.; Findlay, I.; Kurachi, Y. Inwardly rectifying potassium channels: Their structure, function, and physiological roles. Physiol. Rev. 2010, 90, 291–366. [Google Scholar] [CrossRef] [PubMed]
- Eklund, J.; Kozloff, M.; Vlamakis, J.; Starr, A.; Mariott, M.; Gallot, L.; Jovanovic, B.; Schilder, L.; Robin, E.; Pins, M.; et al. Phase II study of mitoxantrone and ketoconazole for hormone-refractory prostate cancer. Cancer 2006, 106, 2459–2465. [Google Scholar] [CrossRef] [PubMed]
- Preet, G.; Oluwabusola, E.T.; Milne, B.F.; Ebel, R.; Jaspars, M. Computational Repurposing of Mitoxantrone-Related Structures against Monkeypox Virus: A Molecular Docking and 3D Pharmacophore Study. Int. J. Mol. Sci. 2022, 23, 14287. [Google Scholar] [CrossRef]
- Kitamura, H.; Yokoyama, M.; Akita, H.; Matsushita, K.; Kurachi, Y.; Yamada, M. Tertiapin potently and selectively blocks muscarinic K+ channels in rabbit cardiac myocytes. J. Pharmacol. Exp. Ther. 2000, 293, 196–205. [Google Scholar] [PubMed]
- Vignani, F.; Bertaglia, V.; Buttigliero, C.; Tucci, M.; Scagliotti, G.V.; Di Maio, M. Skeletal metastases and impact of anticancer and bone-targeted agents in patients with castration-resistant prostate cancer. Cancer Treat. Rev. 2016, 44, 61–73. [Google Scholar] [CrossRef]
- Stojilkovic, S.S.; Tabak, J.; Bertram, R. Ion channels and signaling in the pituitary gland. Endocr. Rev. 2010, 31, 845–915. [Google Scholar] [CrossRef]
- Sanderson, T.; Hisner, R.; Donovan-Banfield, I.A.; Hartman, H.; Løchen, A.; Peacock, T.P.; Ruis, C. A molnupiravir-associated mutational signature in global SARS-CoV-2 genomes. Nature 2023, 623, 594–600. [Google Scholar] [CrossRef] [PubMed]
- Zarenezhad, E.; Marzi, M. Review on molnupiravir as a promising oral drug for the treatment of COVID-19. Med. Chem. Res. 2022, 31, 232–243. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.N.; So, E.C.; Liao, Y.K.; Huang, Y.M. Reversal by ranolazine of doxorubicin-induced prolongation in the inactivation of late sodium current in rat dorsal root ganglion neurons. Pain Med. 2015, 16, 1032–1034. [Google Scholar] [CrossRef] [PubMed]
- Salazar, C.A.; Basilio Flores, J.E.; Veramendi Espinoza, L.E.; Mejia Dolores, J.W.; Rey Rodriguez, D.E.; Loza Munárriz, C. Ranolazine for stable angina pectoris. Cochrane Database Syst. Rev. 2017, 2, Cd011747. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, F.C.; Lewis, R.J. Sodium channels and pain: From toxins to therapies. Br. J. Pharmacol. 2018, 175, 2138–2157. [Google Scholar] [CrossRef]
- Gordon, C.J.; Tchesnokov, E.P.; Feng, J.Y.; Porter, D.P.; Götte, M. The antiviral compound remdesivir potently inhibits RNA-dependent RNA polymerase from Middle East respiratory syndrome coronavirus. J. Biol. Chem. 2020, 295, 4773–4779. [Google Scholar] [CrossRef] [PubMed]
- Morse, J.S.; Lalonde, T.; Xu, S.; Liu, W.R. Learning from the Past: Possible Urgent Prevention and Treatment Options for Severe Acute Respiratory Infections Caused by 2019-nCoV. Chembiochem 2020, 21, 730–738. [Google Scholar] [CrossRef]
- Sheahan, T.P.; Sims, A.C.; Leist, S.R.; Schäfer, A.; Won, J.; Brown, A.J.; Montgomery, S.A.; Hogg, A.; Babusis, D.; Clarke, M.O.; et al. Comparative therapeutic efficacy of remdesivir and combination lopinavir, ritonavir, and interferon beta against MERS-CoV. Nat. Commun. 2020, 11, 222. [Google Scholar] [CrossRef] [PubMed]
- Spinner, C.D.; Gottlieb, R.L.; Criner, G.J.; Arribas López, J.R.; Cattelan, A.M.; Soriano Viladomiu, A.; Ogbuagu, O.; Malhotra, P.; Mullane, K.M.; Castagna, A.; et al. Effect of Remdesivir vs Standard Care on Clinical Status at 11 Days in Patients with Moderate COVID-19: A Randomized Clinical Trial. JAMA 2020, 324, 1048–1057. [Google Scholar] [CrossRef]
- Wang, M.; Cao, R.; Zhang, L.; Yang, X.; Liu, J.; Xu, M.; Shi, Z.; Hu, Z.; Zhong, W.; Xiao, G. Remdesivir and chloroquine effectively inhibit the recently emerged novel coronavirus (2019-nCoV) in vitro. Cell Res. 2020, 30, 269–271. [Google Scholar] [CrossRef]
- Grundeis, F.; Ansems, K.; Dahms, K.; Thieme, V.; Metzendorf, M.I.; Skoetz, N.; Benstoem, C.; Mikolajewska, A.; Griesel, M.; Fichtner, F.; et al. Remdesivir for the treatment of COVID-19. Cochrane Database Syst. Rev. 2023, 1, Cd014962. [Google Scholar] [PubMed]
- Beigel, J.H.; Tomashek, K.M.; Dodd, L.E.; Mehta, A.K.; Zingman, B.S.; Kalil, A.C.; Hohmann, E.; Chu, H.Y.; Luetkemeyer, A.; Kline, S.; et al. Remdesivir for the Treatment of COVID-19—Final Report. N. Engl. J. Med. 2020, 383, 1813–1826. [Google Scholar] [CrossRef] [PubMed]
- Pilote, S.; Simard, C.; Drolet, B. Remdesivir (VEKLURY) for Treating COVID-19: Guinea Pig Ex Vivo and In Vivo Cardiac Electrophysiological Effects. J. Cardiovasc. Pharmacol. 2022, 80, 616–622. [Google Scholar] [CrossRef] [PubMed]
- Fung, J.S.; Levitan, M.; Landry, S.; McIsaac, S. Torsades de pointes associated with remdesivir treatment for COVID-19 pneumonia. J. Assoc. Med. Microbiol. Infect. Dis. Can. 2023, 8, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Tchesnokov, E.P.; Feng, J.Y.; Porter, D.P.; Götte, M. Mechanism of Inhibition of Ebola Virus RNA-Dependent RNA Polymerase by Remdesivir. Viruses 2019, 11, 326. [Google Scholar] [CrossRef] [PubMed]
- Nabati, M.; Parsaee, H. Potential Cardiotoxic Effects of Remdesivir on Cardiovascular System: A Literature Review. Cardiovasc. Toxicol. 2022, 22, 268–272. [Google Scholar] [CrossRef]
- Brown, A.J.; Won, J.J.; Graham, R.L.; Dinnon, K.H., 3rd; Sims, A.C.; Feng, J.Y.; Cihlar, T.; Denison, M.R.; Baric, R.S.; Sheahan, T.P. Broad spectrum antiviral remdesivir inhibits human endemic and zoonotic deltacoronaviruses with a highly divergent RNA dependent RNA polymerase. Antiviral Res. 2019, 169, 104541. [Google Scholar] [CrossRef] [PubMed]
- Hassett, K.J.; Benenato, K.E.; Jacquinet, E.; Lee, A.; Woods, A.; Yuzhakov, O.; Himansu, S.; Deterling, J.; Geilich, B.M.; Ketova, T.; et al. Optimization of Lipid Nanoparticles for Intramuscular Administration of mRNA Vaccines. Mol. Ther. Nucleic Acids 2019, 15, 1–11. [Google Scholar] [CrossRef]
- Lam, K.; Leung, A.; Martin, A.; Wood, M.; Schreiner, P.; Palmer, L.; Daly, O.; Zhao, W.; McClintock, K.; Heyes, J. Unsaturated, Trialkyl Ionizable Lipids are Versatile Lipid-Nanoparticle Components for Therapeutic and Vaccine Applications. Adv. Mater. 2023, 35, e2209624. [Google Scholar] [CrossRef]
- Naidu, G.S.; Yong, S.B.; Ramishetti, S.; Rampado, R.; Sharma, P.; Ezra, A.; Goldsmith, M.; Hazan-Halevy, I.; Chatterjee, S.; Aitha, A.; et al. A Combinatorial Library of Lipid Nanoparticles for Cell Type-Specific mRNA Delivery. Adv. Sci. 2023, 10, e2301929. [Google Scholar] [CrossRef]
- Behr, J.P.; Demeneix, B.; Loeffler, J.P.; Perez-Mutul, J. Efficient gene transfer into mammalian primary endocrine cells with lipopolyamine-coated DNA. Proc. Natl. Acad. Sci. USA 1989, 86, 6982–6986. [Google Scholar] [CrossRef] [PubMed]
- Albert, E.; Aurigemma, G.; Saucedo, J.; Gerson, D.S. Myocarditis following COVID-19 vaccination. Radiol. Case Rep. 2021, 16, 2142–2145. [Google Scholar] [CrossRef] [PubMed]
- Bozkurt, B.; Kamat, I.; Hotez, P.J. Myocarditis with COVID-19 mRNA Vaccines. Circulation 2021, 144, 471–484. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.W.; Jenista, E.R.; Wendell, D.C.; Azevedo, C.F.; Campbell, M.J.; Darty, S.N.; Parker, M.A.; Kim, R.J. Patients with Acute Myocarditis following mRNA COVID-19 Vaccination. JAMA Cardiol. 2021, 6, 1196–1201. [Google Scholar] [CrossRef] [PubMed]
- Vidula, M.K.; Ambrose, M.; Glassberg, H.; Chokshi, N.; Chen, T.; Ferrari, V.A.; Han, Y. Myocarditis and Other Cardiovascular Complications of the mRNA-Based COVID-19 Vaccines. Cureus 2021, 13, e15576. [Google Scholar] [CrossRef] [PubMed]
- Williams, C.B.; Choi, J.I.; Hosseini, F.; Roberts, J.; Ramanathan, K.; Ong, K. Acute Myocarditis Following mRNA-1273 SARS-CoV-2 Vaccination. CJC Open 2021, 3, 1410–1412. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.Y.; Chuang, T.H.; Wu, S.N. The Effectiveness in Activating M-Type K+ Current Produced by Solifenacin ([(3R)-1-azabicyclo[2.2.2]octan-3-yl] (1S)-1-phenyl-3,4-dihydro-1H-isoquinoline-2-carboxylate): Independent of Its Antimuscarinic Action. Int. J. Mol. Sci. 2021, 22, 12399. [Google Scholar] [CrossRef] [PubMed]
- Martín-Montañez, E.; López-Téllez, J.F.; Acevedo, M.J.; Pavía, J.; Khan, Z.U. Efficiency of gene transfection reagents in NG108-15, SH-SY5Y and CHO-K1 cell lines. Methods Find. Exp. Clin. Pharmacol. 2010, 32, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Azdaki, N.; Farzad, M. Long QT interval and syncope after a single dose of COVID-19 vaccination: A case report. Pan Afr. Med. J. 2021, 40, 67. [Google Scholar] [CrossRef]
- Abdelgalil, A.A.; Alkahtani, H.M.; Al-Jenoobi, F.I. Sorafenib. Profiles Drug Subst. Excip. Relat. Methodol. 2019, 44, 239–266. [Google Scholar]
- Cabrera, R.; Limaye, A.R.; Horne, P.; Mills, R.; Soldevila-Pico, C.; Clark, V.; Morelli, G.; Firpi, R.; Nelson, D.R. The anti-viral effect of sorafenib in hepatitis C-related hepatocellular carcinoma. Aliment. Pharmacol. Ther. 2013, 37, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.S.; Huang, S.H.; Yan, B.Y.; Lai, H.C.; Lin, C.W. Effective Antiviral Activity of the Tyrosine Kinase Inhibitor Sunitinib Malate against Zika Virus. Infect. Chemother. 2021, 53, 730–740. [Google Scholar] [CrossRef] [PubMed]
- Theerawatanasirikul, S.; Lueangaramkul, V.; Semkum, P.; Lekcharoensuk, P. Antiviral mechanisms of sorafenib against foot-and-mouth disease virus via c-RAF and AKT/PI3K pathways. Vet. Res. Commun. 2023, 48, 329–343. [Google Scholar] [CrossRef]
- Kloth, J.S.; Pagani, A.; Verboom, M.C.; Malovini, A.; Napolitano, C.; Kruit, W.H.; Sleijfer, S.; Steeghs, N.; Zambelli, A.; Mathijssen, R.H. Incidence and relevance of QTc-interval prolongation caused by tyrosine kinase inhibitors. Br. J. Cancer 2015, 112, 1011–1016. [Google Scholar] [CrossRef] [PubMed]
- Sung, R.J.; Kuo, C.-T.; Wu, S.-N.; Lai, W.-T.; Luqman, N.; Chan, N.-Y. Sudden cardiac death syndrome: Age, gender, ethnicity, and genetics. Acta Cardiol. Sin. 2008, 24, 65–74. [Google Scholar]
- Ansone, L.; Briviba, M.; Silamikelis, I.; Terentjeva, A.; Perkons, I.; Birzniece, L.; Rovite, V.; Rozentale, B.; Viksna, L.; Kolesova, O.; et al. Amino Acid Metabolism is Significantly Altered at the Time of Admission in Hospital for Severe COVID-19 Patients: Findings from Longitudinal Targeted Metabolomics Analysis. Microbiol. Spectr. 2021, 9, e0033821. [Google Scholar] [CrossRef]
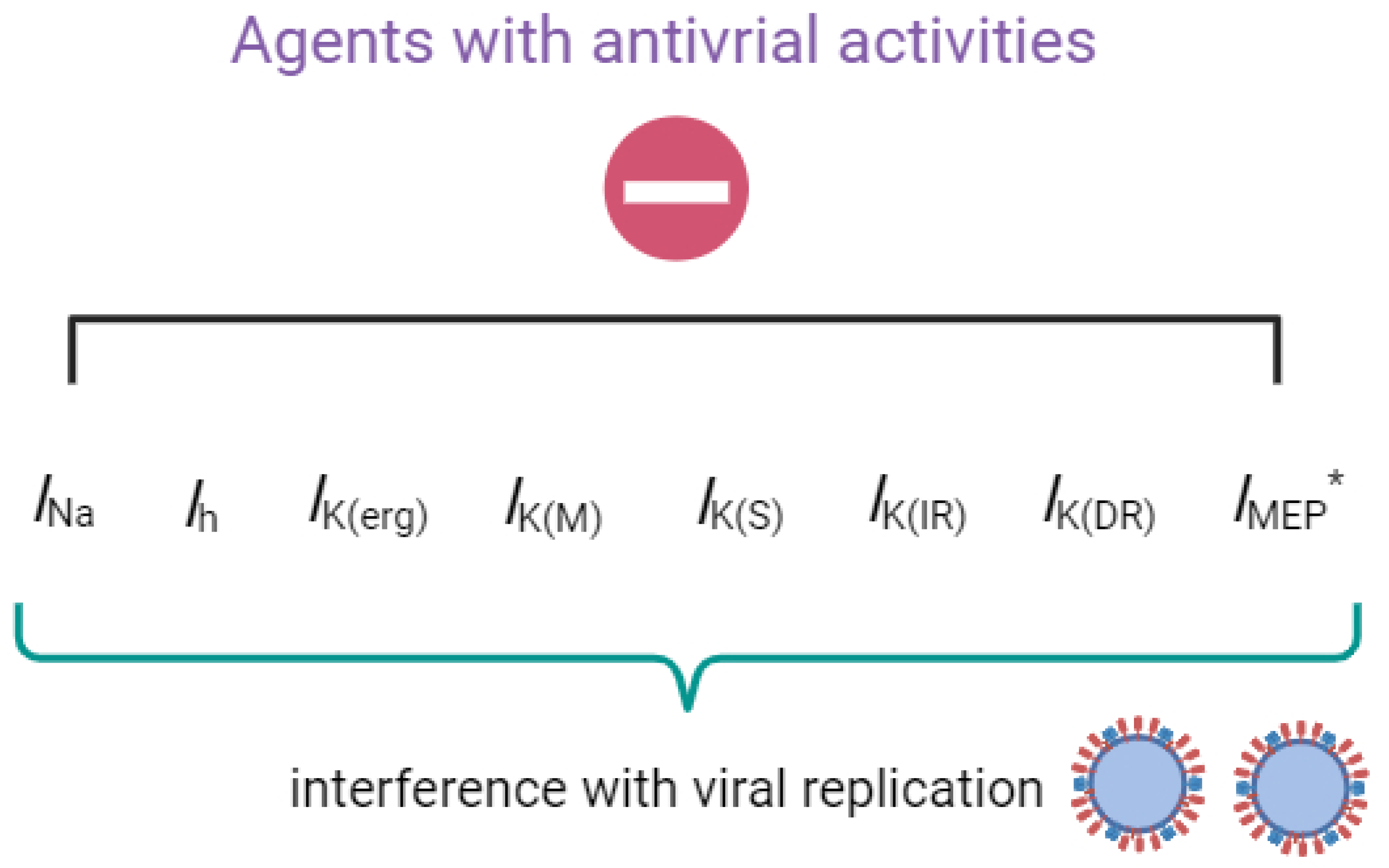

{kind=link}
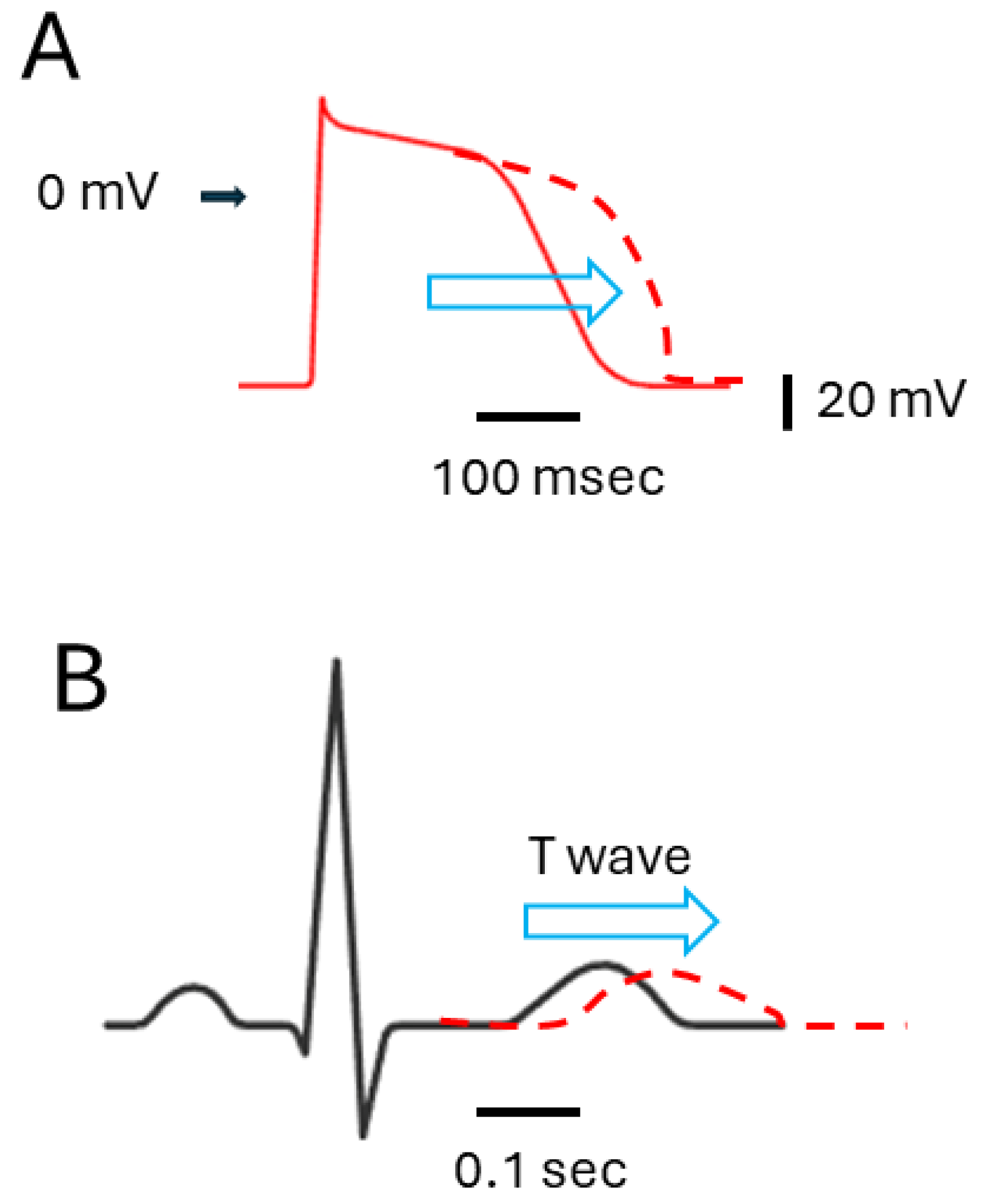
{kind=link}
| Compound or Drug | Chemical Structure | Cell Type Studied | Concentration Range Used |
|---|---|---|---|
| Artemisinin | 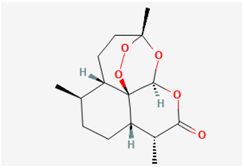 | GH3 cells, TRPC3-expressing HEK293 cells | 1–100 μM |
| Cannabidiol |  | GH3 cells | 0.3–100 μM |
| Memantine | 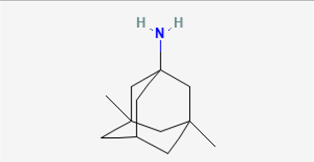 | GH3, RAW 264.7, and BV2 cells | 10–1 mM |
| Mitoxantrone | 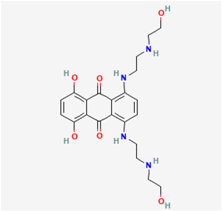 | RAW 264.7 cells | 1–100 μM |
| Molnupiravir |  | GH3 cells | 1–300 μM |
| Remdesivir |  | GH3 and Jurkat T cells | 0.3–100 μM |
| SM-102 | 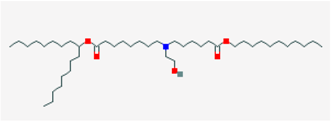 | GH3 cells and Leydig MA-10 cells | 3–1 mM |
| Sorafenib |  | H9c2, RUES2 cells *, and neonatal rat ventricular myocytes | 1–30 μM |
| Compound or Drug | Abbreviation | Actions on Ionic Currents *, ** | References |
|---|---|---|---|
| Artemisinin | ART | ↓ IK(DR) ↓ INa ↑ TRPC channel | Qiao et al. (2007) [11] So et al. (2017) [12] Zhang et al. (2022) [13] |
| Cannabidiol | CBD | ↓ IK(M) ↓ Ih | Liu et al. (2023) [14] |
| Memantine | MEM | ↓ IMEP ↓ IK(IR) | Wu et al. (2011) [15] Tsai et al. (2013) [16] |
| Mitoxantrone | MX | ↓ IK(IR) | Wang et al. (2012) [17] |
| Molnupiravir | MOL | ↓ INa | Shiau et al. (2023) [18] |
| Remdesivir | RDV | ↓ IK(DR) ↓ IK(M) ↑ IMEP ↓ IK(erg) | Chang et al. (2020) [19] Amarh et al. (2023) [20] |
| SM-102 | SM-102 | ↓ IK(erg) ↓ IK(IR) | Cho et al. (2021) [21] |
| Sorafenib | SOR | ↓ IK(erg) ↓ IK(S) ↓ IK(IR) | Wu et al. (2012) [22] Chang et al. (2020) [23] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, G.-B.; Shih, C.-L.; Liutkevičienė, R.; Rovite, V.; So, E.C.; Wu, C.-L.; Wu, S.-N. Assessing the Impact of Agents with Antiviral Activities on Transmembrane Ionic Currents: Exploring Possible Unintended Actions. Biophysica 2024, 4, 128-141. https://doi.org/10.3390/biophysica4020009
Lin G-B, Shih C-L, Liutkevičienė R, Rovite V, So EC, Wu C-L, Wu S-N. Assessing the Impact of Agents with Antiviral Activities on Transmembrane Ionic Currents: Exploring Possible Unintended Actions. Biophysica. 2024; 4(2):128-141. https://doi.org/10.3390/biophysica4020009
Chicago/Turabian StyleLin, Geng-Bai, Chia-Lung Shih, Rasa Liutkevičienė, Vita Rovite, Edmund Cheung So, Chao-Liang Wu, and Sheng-Nan Wu. 2024. "Assessing the Impact of Agents with Antiviral Activities on Transmembrane Ionic Currents: Exploring Possible Unintended Actions" Biophysica 4, no. 2: 128-141. https://doi.org/10.3390/biophysica4020009