Abstract
This study investigates the formation and impact of Endogenous Quasi-Pathogens (EQPs) within cellular environments, focusing on the role of Endogenous Smart Medicine (ESM) as a therapeutic intervention. This work elucidates how induced vibrations facilitate new molecular and atomic connections between adjacent cells, leading to endobiotic bond formation and significantly altered DNA behavior. These vibrations, which dominate cellular processes, induce both temporary and permanent changes in cellular dynamics. The resulting increase in extracellular impedance triggers the emergence of new EQP sources, potentially initiating divergent pathological cycles. Cells experiencing moderate impedance changes are classified as benign, while those with substantial alterations are considered malignant. This study highlights the medical diagnostic implications of EQPs and positions ESM as a precise method for modulating cellular impedance, reducing the effects of EQPs, and potentially treating diseases where disruptions in cellular dynamics and stiffness are critical. Additionally, the integration of ChronoBit Storage (CBS) within ESM introduces a novel energy management mechanism, enhancing therapeutic precision by synchronizing energy distribution with cellular needs. The ChronoVital Index (CVI), a temporal model for assessing time dynamics across biological systems from individual cells to whole organs further refines this approach. By advancing the CVI and CBS, this research paves the way for more sophisticated therapeutic strategies, offering promising applications in the fields of disease management and cellular restoration within the framework of Endogenous Smart Medicine.
Keywords:
Endogenous Quasi-Pathogens (EQPs); Endogenous Smart Medicine (ESM); ChronoCell Matrix (CCM); CellChrono Reference (C0), ChronoVital Index (CVI); ChronoBit Storage (CBS); Endobiotic Bond Quasi-Pathogens; Subtle Quasi-Pathogen; Gloomy Eyelet (GE); Warp Drive Hydro (WDH) Model; Swinging Spring Model; Co-Moving Reference Frame (CMRF); Rankine Model; Pseudo-Spherical Geometry; Space–Time 1. Introduction
Induced vibrations within cellular systems have garnered significant interest in biomedical research due to their potential implications for understanding pathogen behaviors and cellular disturbances. This paper investigates the interplay between induced vibrations and cellular dynamics, focusing on the emergence of Endogenous Quasi-Pathogens (EQPs). This paper investigates the complex interplay between induced vibrations, Gloomy Eyelets (GEs) within cell nuclei, and their impacts on cellular dynamics. By drawing insights from biophysics, nanotechnology, and cellular biology, our analysis illuminates the emergence of Gloomy Eyelets (GEs), nano distortions of space–time, and their interactions with the extracellular environment. Using Hamiltonian equations, fluid dynamics principles, and previous research, we elucidate the dynamics of coupled GEs and their implications for cellular disturbances [1,2].
2. Materials and Methods
We explore pressure wave generation and frequencies within DNA, revealing the role of Nano Latent Buoyancy in cellular processes. This study provides novel insights into pathogen behaviors, cellular disturbances, and their relevance to human health, with potential applications in medical diagnostics and therapeutic interventions.
For a circular plate, the shapes of modes with nodal lines can be rotated by using two excitation signals at different locations on the outer edge with certain phase and space differences. The superposition of the two standing waves results in a traveling wave that can be observed in a rotating mode shape. The circular cell is chosen for studying and exerting the excitation forces. The swinging spring model is made on the circular plate (as the forced vortex region of the Rankine model), which is faced with resonance frequency.
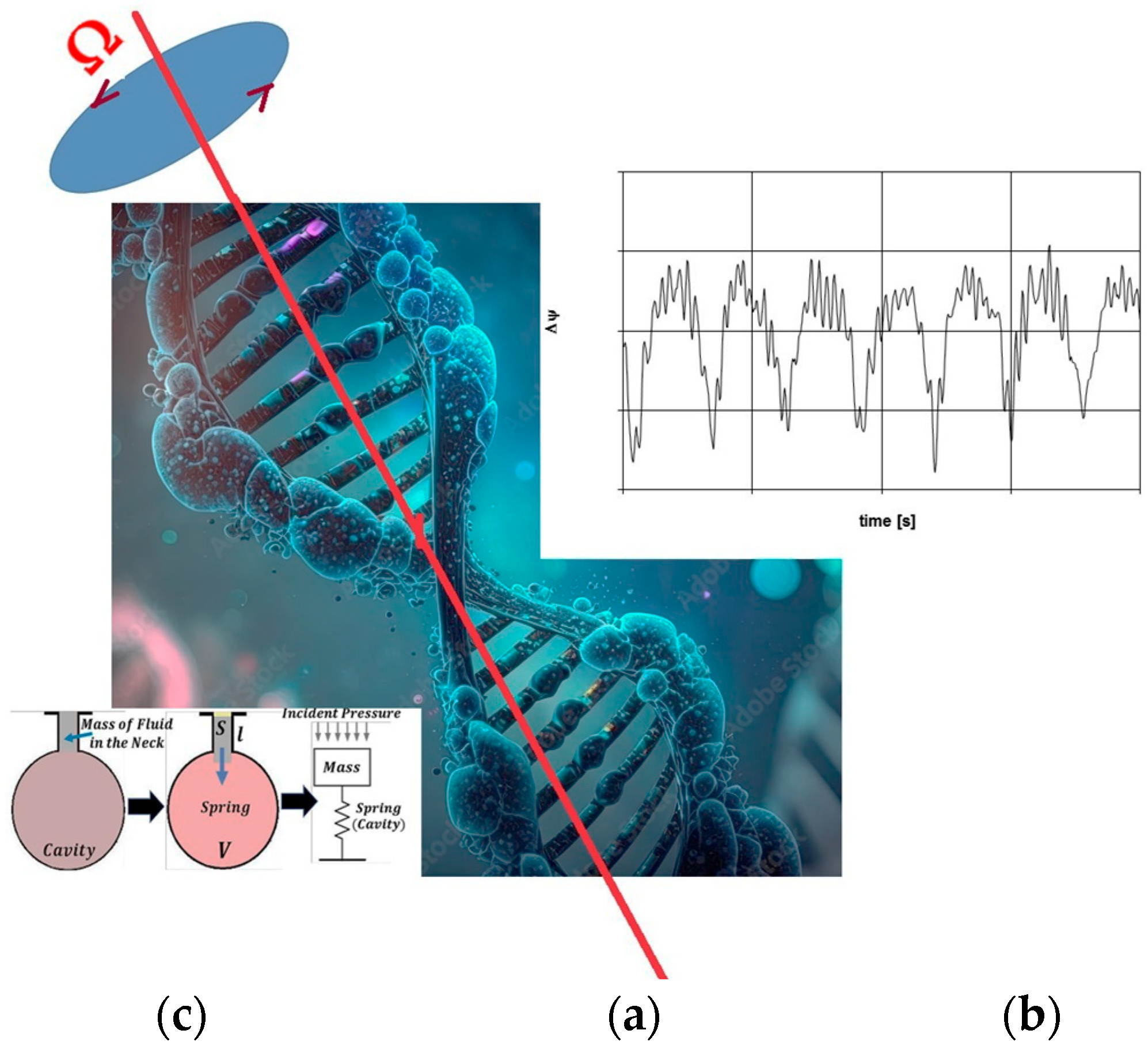
The synchronization between the beat frequency (Ωb) of blood pressure within vessels and the rotation speed (Ω′) of the left ventricle, as described by the Rankine model, leads to a sudden pressure drop and rapid cavity formation within the Warp Drive Hydro (WDH) medium. This phenomenon is anticipated when the DNA’s swinging spring frequency (Ω) converges with the blood pressure beat frequency (Ωb), potentially inducing a pressure drop similar to that of an accelerated cavity, as postulated by the Rankine model. This phenomenon results in DNA’s longitudinal expansion, creating nanoscale cavities and reducing internal pressure, leading to the emergence of Gloomy Eyelets (GEs) within the WDH model of DNA, as illustrated in Figure 1.
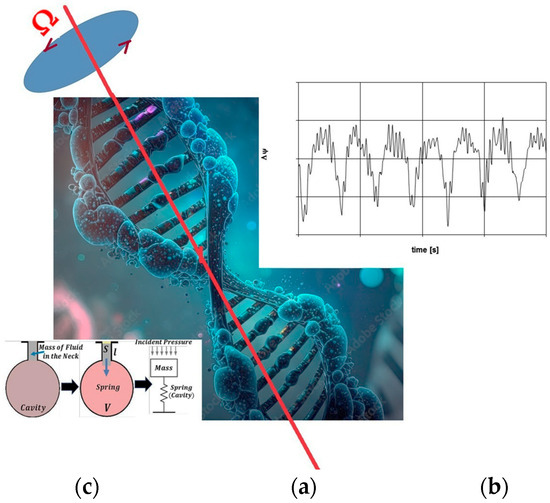
Figure 1.
The rotating standing wave within the cell nucleus represented by its rotational speed (Ω), as shown in (a) within the WDH Model, interacts with the pulsating pressure wave in the blood vessels, which is characterized by its envelope frequency (Ωb), as depicted in (b), alongside the Helmholtz resonator model, as indicated in (c).
Within nanoscale domains, photons generated by Quantum Electrodynamics (QED) processes may affect the dynamics of co-moving reference frames (CMRF), indicating subtle space–time distortions within cellular structures, particularly in the nucleus. These distortions can be likened to oscillatory perturbations that follow a pseudo-spherical trajectory within certain physical models.
To draw a comparison with a more tangible system, consider the Helmholtz resonator, which is a device comprising a resonant cavity and a neck through which air oscillates. The air’s motion within the neck can be modeled as a mass attached to a spring, obeying Newton’s Second Law. When subjected to an external sinusoidal driving force, the motion is governed by Equation (1):
where:
- m is the mass of the air in the neck,
- R is the resistance coefficient (damping),
- K is the spring constant,
- Rv is the resistance force,
- x is the displacement,
- f(t)e(iωt) is the external driving force.
In a driven, damped spring–mass system, three forces come into play: the driving force f(t)e(iωt), the resistance force Rv (where R represents resistance and v represents velocity), and the spring’s restorative force kx (where k is the spring constant and x is the displacement) [3,4,5,6].
Induced vibrations, arising from what we term “Gloomy Eyelets,” represent the effects of minute distortions in nanoscale space and time. These distortions have the potential to generate new oscillatory modes within biological systems by exciting a secondary frequency, denoted as ω′, in addition to the primary driving frequency ω. When such distortions are present, they can introduce these new frequencies, causing the system to oscillate not only at its original frequency but also at the newly induced frequency ω′. In a biological context, these additional oscillations may correspond to abnormal or altered behaviors in pathogens, potentially leading to the emergence of new pathogen dynamics. This phenomenon can play a significant role in modulating the behaviors of pathogens within the biological environment, as described by Equation (2), leading to unpredictable and potentially novel interactions.
The amplitude of oscillation in the spring–mass system is influenced by mechanical resistance. The quality factor q delineates the amplitude of oscillation concerning resonance frequency and resistance. High-quality factor systems exhibit minimal resistance and amplified oscillation at resonance, while low-quality factor systems exhibit dampened response at resonance, as characterized by its broader peak in the resonance curve [1,2,7,8].
2.1. Pathogen Behaviors and Cellular Disturbances
Our research explores how induced vibrations impact pathogen behaviors associated with Gloomy Eyelets. Gloomy Eyelets are hypothesized to be areas within the biological medium exhibiting unique vibrational characteristics. This type of vibration, with a frequency excitation denoted as ω, propagates pathogens with specific behaviors into the biological medium along a Gaussian curve; this is a common probability distribution that describes values distributed symmetrically around a mean.
We propose a theoretical framework where quasi-particle A harbors Quasi-Pathogens, which are not fully developed pathogens but instead have the potential to evolve under certain conditions. Induced vibrations from Gloomy Eyelets, including tiny distortions in nano space and time, can generate new oscillations through the excitation of frequency ω′ in pathogens. This phenomenon is expected to induce new pathogen behaviors in the biological medium, which are modeled on the Complementary Gaussian Distribution curve. Slight time dilation in the environment gives rise to quasi-particle B, representing Subtle Quasi-Pathogens.
2.2. Endogenous Quasi-Pathogens
Understanding the interplay between the cellular microenvironment and breast cancer metastasis is crucial for developing effective treatments. Although progress has been made in elucidating the molecular mechanisms of breast cancer progression, the quantitative characterization of cellular mechanical properties remains incomplete. Breast tumors are stiffer than normal breast tissue, and the stiffness of the extracellular matrix influences cancer cell motility, which is a key aspect of metastasis.
To study and create endogenous Quasi-Pathogens, it is crucial to consider certain parameters, such as interaction frequencies, viscosity, stiffness, and the impedance related to the cell’s medium.
2.2.1. Viscoelastic Behavior
Viscoelastic properties are important in understanding the time-dependent deformation characteristics of materials under stress. In one dimension, the stress history σ can be considered as a step function starting at time zero. The strain ε(t) in a viscoelastic material consequently increases over time. Creep is the slow, progressive deformation of a material under constant stress, and it can be described by the creep compliance in Equation (3):
- J(t): The function J(t) represents how the material’s deformation (strain) progresses over time under constant stress. It is a key property in viscoelasticity, reflecting the material’s ability to “creep”.
- ϵ(t): The strain ϵ(t) changes over time, indicating that the material is progressively deforming.
- σ0: The constant stress σ0 that is applied to the material.
For linear viscoelastic materials, the creep compliance J(t) is independent of the magnitude of the applied stress σ0. This independence means that doubling the applied stress doubles the strain, but the ratio ϵ(t)/σ0 (creep compliance) remains the same, as long as the material behaves linearly.
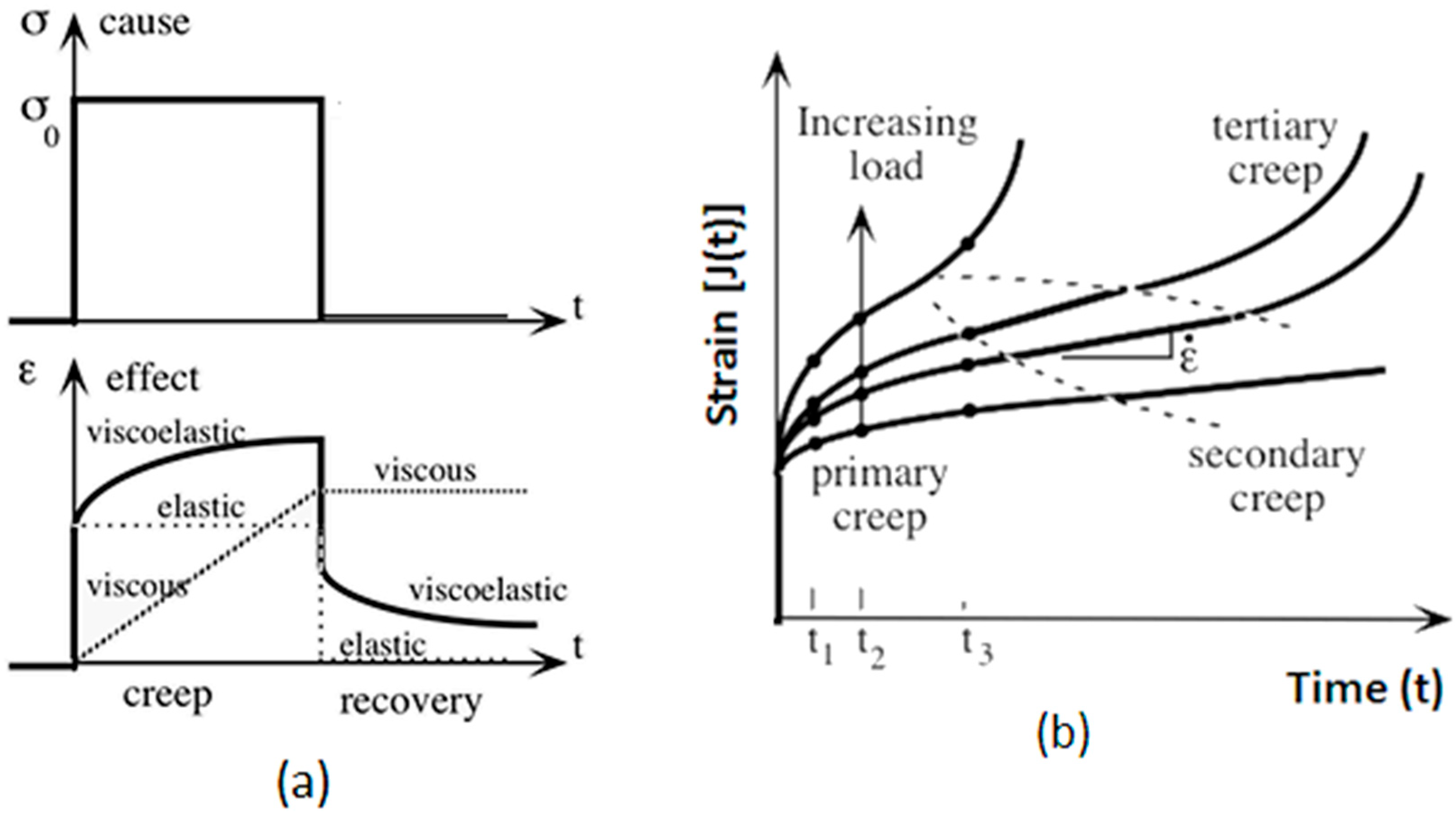
Upon unloading, strain recovery ensues based on the material’s inherent properties, as shown in Figure 2.
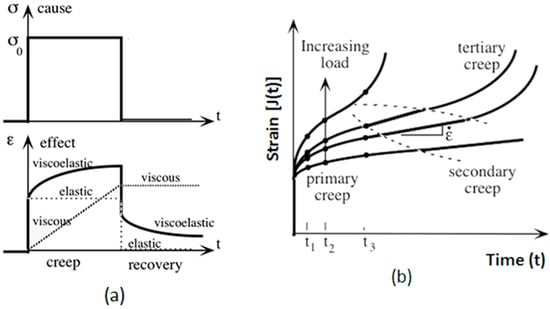
Figure 2.
The creep behavior regions are (a) the stress and strain plotted against time (b) and an illustration of the strain, J(t), as a function of time under varying load levels.
Viscoelastic behavior is often modeled using mechanical frameworks consisting of elastic springs and viscous dashpots. The time-dependent responses of viscoelastic materials mirror those of reactive electrical circuits, which are both governed by analogous ordinary differential equations over time.
Figure 2 depicts three distinct phases of creep: primary creep, characterized by a decreasing strain rate; secondary creep, where the strain rate stabilizes; and tertiary creep, marked by an accelerating strain rate leading to failure. The progression through these phases is shown for increasing load levels, with ϵ representing the total strain experienced at each stage.
A common approach to conceptualize these relationships involves employing ‘spring–dashpot’ models. Here, the spring represents the instantaneous deformation of bonds within the material, with its magnitude corresponding to the fraction of mechanical energy stored reversibly as strain energy. The process of entropic uncoiling behaves akin to a fluid modeled by a ‘Newtonian dashpot’.
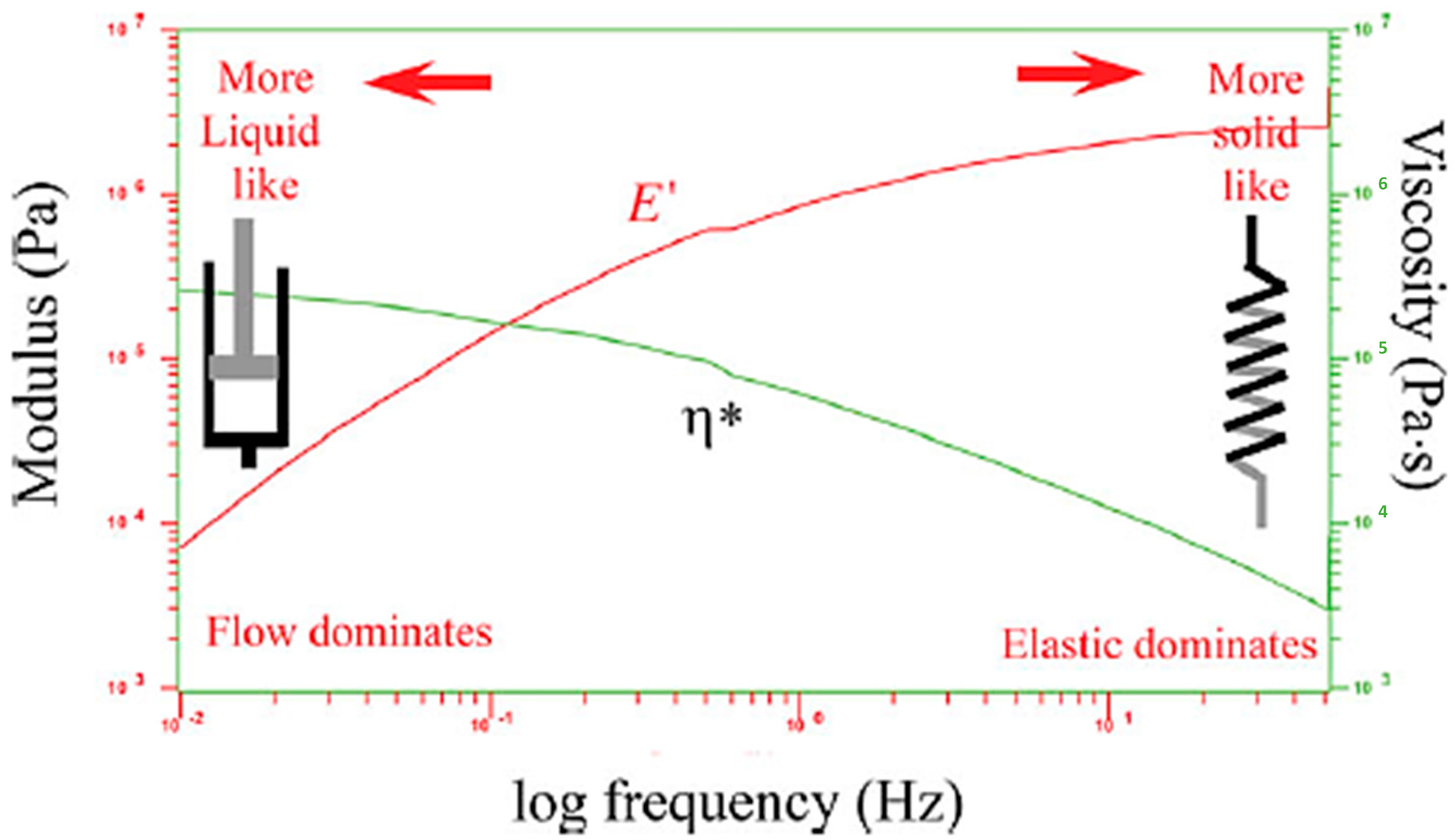
This version clarifies the analogy between mechanical and electrical models while maintaining clarity on how spring–dashpot models are used to understand viscoelastic behaviors (refer to Figure 3). As the frequency increases and shear thinning occurs, the viscosity (η*) decreases. At the same time, increasing the frequency increases the elasticity (E′) [9,10,11].
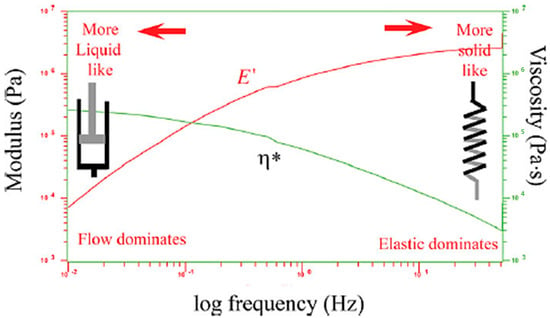
Figure 3.
The spring represents the material’s instant deformation and reversible strain energy, while the dashpot models represent entropic uncoiling as a fluid. This analogy clarifies how the mechanical model explains viscoelastic behavior, showing that as the frequency increases, the viscosity (η*) decreases because of shear thinning, and the elasticity (E′) increases.
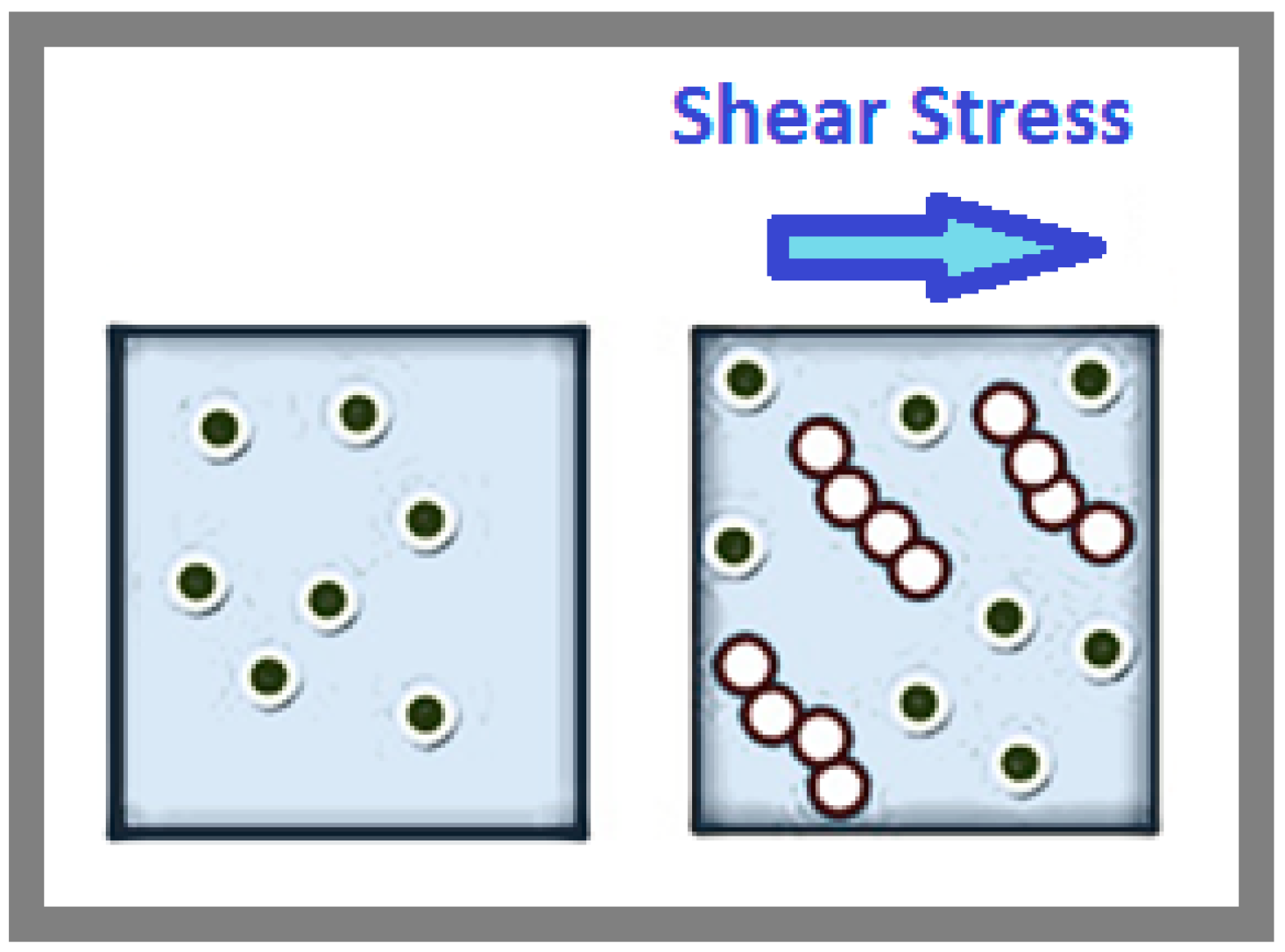
In fluid systems, shear stress can induce particles to reorganize into more efficient arrangements, reducing resistance and viscosity. Similarly, in biological systems, cellular vibrations can theoretically promote the alignment of molecules within cells. This alignment can optimize molecular interactions, improve cellular function, and minimize mechanical resistance, paralleling how shear stress aligns particles and enhances flow in fluid systems, as shown in Figure 4. The left image in Figure 4 represents the state of a particle suspension without the application of shear stress. Here, the particles are randomly distributed, maintaining a disordered structure. In this configuration, the particle interactions are primarily governed by repulsive forces, preventing them from coming too close to each other. The disorder in the system contributes to higher viscosity as the random arrangement increases the resistance to flow. In contrast, the right image in Figure 4 demonstrates the effect of shear stress on the same particle suspension. As shear stress is applied (indicated by the arrow), the particles overcome their repulsive interactions and align themselves in an ordered structure. This alignment reduces the overall resistance to flow, leading to a decrease in viscosity. The ordered rows of particles are a direct consequence of the directional forces exerted by the shear stress, organizing the particles in a more efficient arrangement for movement.
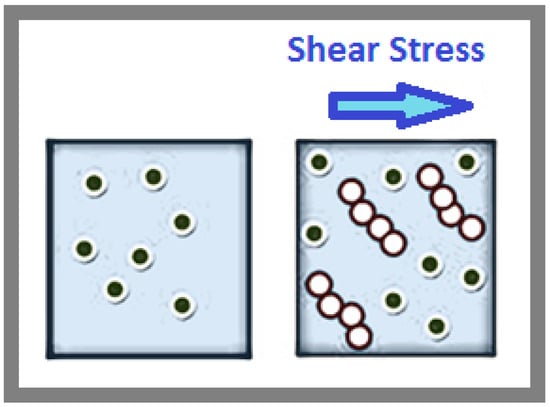
Figure 4.
The shear-induced ordering of particles under shear stress. The left image shows particles in a disordered state without shear stress, where repulsive forces dominate, increasing viscosity. The right image illustrates the effect of shear stress, where particles align into ordered rows, reducing resistance to flow and decreasing viscosity. This demonstrates the transformation from a disordered to an ordered structure under the influence of shear stress.
2.2.2. Impedance
Impedance is a valuable concept for understanding the flow of energy. The rate at which energy is used is referred to as power. When an object is pushed with a force and moves at a certain velocity, the power is the product of the force and the velocity. The movement of the object is dependent on the applied force, and the two are interrelated. For a harmonic force, the ratio of force to velocity indicates how difficult it is to move the object, which depends on the inertia of the object and the strength of the restoring force.
It is important to note that when discussing waves, velocity refers to the ‘particle’ velocity, which is the speed of displacement, not the propagation velocity of the wave itself. Moving an object through a viscous medium, such as molasses, requires a constant force to maintain a constant velocity. This relationship is expressed by the medium’s viscosity. In such viscous media, the impedance is equal to the viscosity.
Impedance is a complex quantity that represents the opposition a system offers to a sinusoidal driving force. In a driven oscillator, the system’s overall impedance is the sum of the individual impedances of all components/molecules of that system.
At high frequencies, the mass term dominates, resulting in mass-dominated impedance, where a high-frequency force has minimal effect on velocity due to the mass’s inability to react quickly.
Conversely, at low frequencies, the stiffness term prevails, leading to stiffness-dominated impedance. In this case, the velocity for a given force is significantly influenced by the spring’s stiffness. The impedance of the whole system is the sum of the impedances (Ztotal), as shown in Equation (4) [12]:
- Zm = iωm is the impedance due to the mass.
- Zk = −ik/ω is the impedance due to the stiffness.
- i is the imaginary unit, representing a 90-degree phase shift between the force and velocity in purely reactive components.
The velocities of Quasi-Pathogens vary across different media, each with a specific stiffness.
3. Main Models
3.1. Formation of Endogenous Quasi-Pathogens (EQPs)
Induced vibrations can facilitate the formation of new molecular and atomic connections within a cell and its adjacent cells, resulting in the creation of endobiotic bonds due to the presence of shear and dynamic forces. Certain vibration frequencies may become dominant, significantly altering the DNA behaviors of nearby cells. Consequently, human cellular DNA may experience a range of temporary and permanent behavioral changes due to the formation of these new endobiotic bonds, which in turn form EQPs.
As these changes occur, the stiffness of the surrounding medium increases, leading to a gradual rise in impedance. The vibrations of the newly formed EQPs generate waves that are partially transmitted through the altered medium, while others are reflected. If the frequencies of the reflected waves match or are close to the frequencies of other EQPs, there is a higher likelihood of resonance, which can amplify the wave intensity. This amplification leads to a significant increase in the medium’s impedance.
The interaction and accumulation characteristics of these amplified EQP sources further increase the medium’s stiffness, potentially resulting in continuous and divergent cycles of EQPs generation. Cells experiencing moderate changes in the stiffness of the surrounding medium may be classified as benign, whereas those undergoing significant changes may be classified as malignant. This continuous process of EQP generation, reflection, and resonance can lead to the occurrence of endless cycles, significantly impacting cellular dynamics and potentially contributing to disease progression.
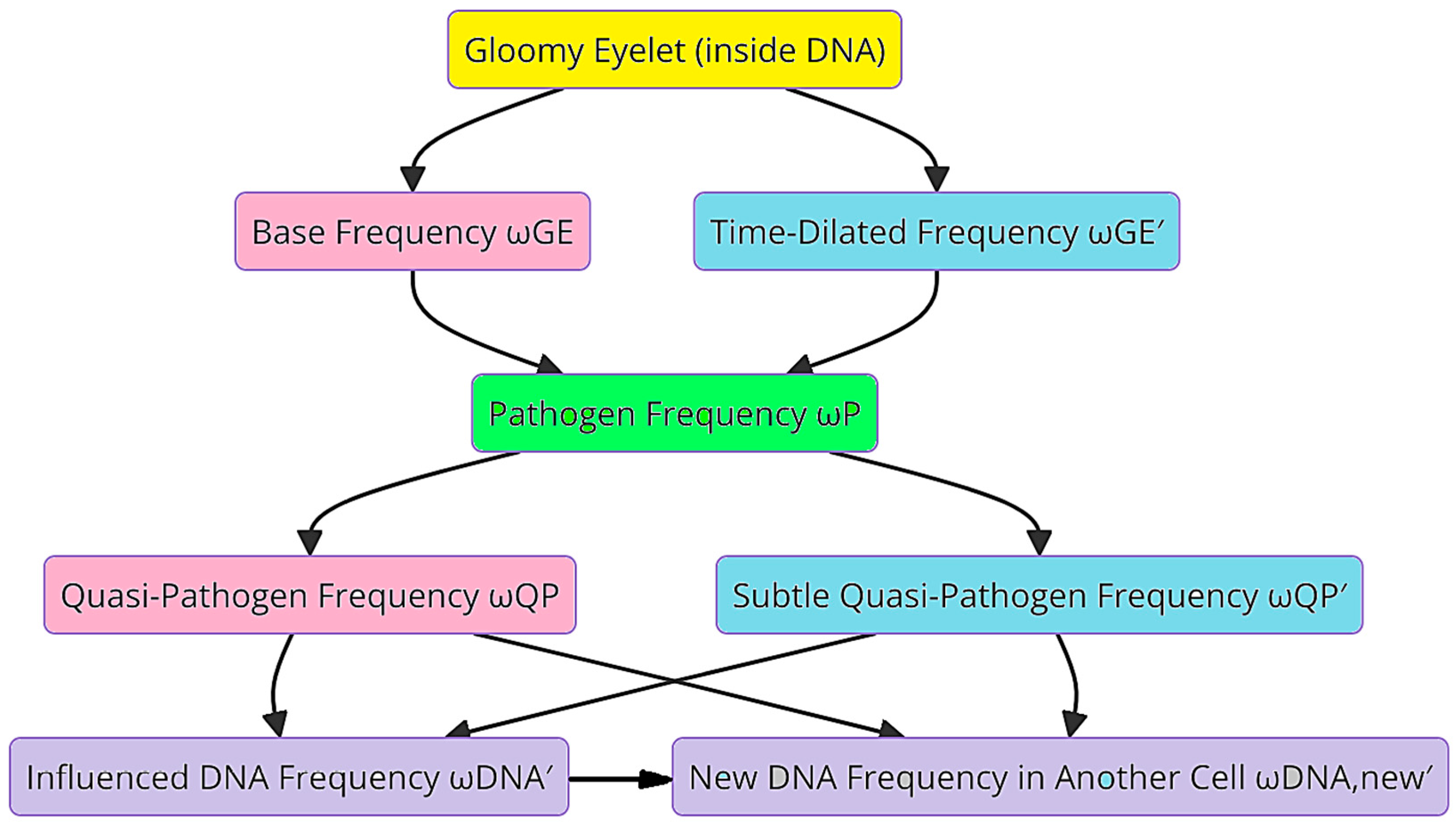
The Gloomy Eyelet, acting as an initial frequency source, generates both a base frequency and a time-dilated frequency. These frequencies propagate through a biological medium. Table 1 details various frequency components and their descriptions, including the base frequency, time-dilated frequency, pathogen frequency, and their effects on DNA. These findings are shown in Figure 5, which illustrates the flow diagram of how these frequencies interact with pathogens, Quasi-Pathogens, and DNA.

Table 1.
Frequency Modulation of Cellular and Pathogenic Interaction.
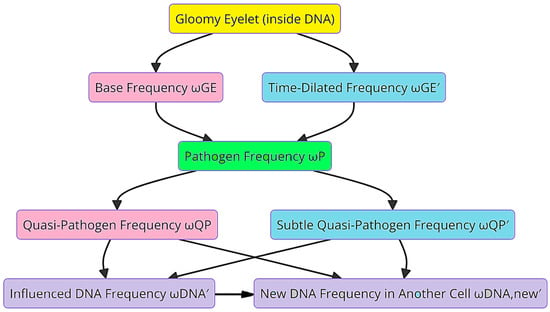
Figure 5.
A flow diagram of the propagation of base and time-dilated frequencies through pathogen modulation in biological applications.
The generation of endogenous Quasi-Pathogens involves complex frequency interactions between cells. These interactions significantly affect the impedance and stiffness of the surrounding medium. When the frequency of reflected waves matches or closely aligns with the frequencies of Quasi-Pathogens or Subtle Quasi-Pathogens, resonance occurs, amplifying the wave intensity and increasing the medium’s impedance. Understanding these interactions is crucial for comprehending the continuous and divergent cycles of Quasi-Pathogen generation and their impacts on cellular dynamics.
3.2. Endogenous Smart Medicine (ESM)
The synchronization of specific frequencies and induced vibrations triggers the formation of Endogenous Quasi-Pathogens (EQPs), creating new molecular bonds between adjacent cells. ESM offers a promising solution to counter these effects by regulating amplitude and stiffness, preventing the occurrence of harmful cycles within cells. By precisely managing the stiffness and impedance of the surrounding medium, ESM can modulate vibrations to influence Quasi-Pathogen behavior, reducing the formation of reflected waves that generate new Quasi-Pathogen sources. This targeted intervention helps maintain the integrity and stability of the medium, preventing the escalation of cyclic disturbances.
Practically, ESM enables the classification of cells experiencing moderate changes in surrounding medium stiffness and impedance as benign. This proactive approach mitigates the risk of malignant transformations, which is particularly crucial in diseases like cancer. By carefully modulating extracellular matrix stiffness, ESM holds promise in impeding tumor progression and metastasis.
3.3. Controlled Cellular Modulation with Endogenous Smart Medicine (ESM)
ESM aims to develop internal cellular mechanisms akin to Quasi-Pathogens for controlled modulation within cellular environments. ESM involves creating internal sources that mimic Quasi-Pathogens, facilitating the formation of beneficial endobiotic bonds through induced vibrations and specific frequency interactions.
A critical aspect of ESM is its ability to precisely control stiffness, impedance, frequency, and amplitude within cells. By understanding and manipulating these parameters, ESM seeks to optimize cellular dynamics and mitigate pathological conditions. Techniques may include using targeted acoustic or electromagnetic waves to modulate cellular behaviors without adverse effects.
Monitoring and feedback mechanisms are integral to ESM, enabling real-time adjustments to modulation strategies for optimal outcomes. Clinical validation is essential to ensure the safety and efficacy of ESM interventions in practical medical applications. This iterative approach aims to refine and enhance therapeutic strategies by leveraging insights from Quasi-Pathogen behaviors and cellular modulation processes.
3.4. ChronoCell Matrix (CCM)
The ChronoCell Matrix (CCM) is a complex temporal framework designed to understand and quantify the time dynamics across various biological levels from individual cells to entire organs and the whole human body itself. The goal is to create a cohesive model that can be used to analyze how time dilation and other temporal factors impact cellular functions, organ efficiency, and overall human health. The model begins with the concept of the CellChrono Reference, which serves as a standardized time frame for various cellular processes.
Subtle Quasi-Pathogens can significantly impact the ChronoCell Matrix (CCM) by disrupting the normal temporal rhythms of cells. This disruption can lead to altered cellular functions, which may manifest in various pathological conditions. Endogenous Smart Medicine aims to detect and counteract these disruptions by restoring the natural chrono-rhythms within cells, thereby preventing disease progression.
3.5. Conceptual Framework
The CellChrono Reference is a fundamental concept in the ChronoCell Matrix (CCM), representing a standardized time frame for cellular processes under ideal conditions. This concept acts as a baseline to measure how time behaves under various influences like aging, disease, and environmental stressors. The key idea is that time can dilate or stretch under different conditions, and this dilation is captured using a Dilation Factor (γ), which is the ratio of the observed time (T1) to the reference time (T0).
CellChrono Reference (C0): A standardized temporal unit that measures the time dynamics of cellular processes under normal conditions that is adjusted for time dilation effects.
ChronoCell Matrix (CCM): An aggregated temporal profile that extends the CellChrono concept to organs and the entire human body.
3.6. Improved Formula for CellChrono Reference for a Single Cell
To enhance the precision of the CellChrono Reference (C0) in Equation (5), we can incorporate additional factors that influence cellular time dynamics:
where:
- T0: Reference time under standard conditions.
- T1: Observed time in a specific environment.
- T′: Co-Moving Reference Time accounting for time dilation.
- Es and E0: Intracellular energy states (e.g., ATP levels) under specific and reference conditions, respectively.
- Fs and F0: Frequency of cellular processes (e.g., mitosis, protein synthesis) under specific and reference conditions, respectively.
- α and β: Empirical constants that adjust the influence of energy and frequency on the CellChrono Reference (C0).
This refined formula allows for a more comprehensive evaluation of the temporal dynamics within a cell, accounting for both the energy state and frequency of cellular processes [6,13].
3.7. Application to ChronoCell Matrix (CCM) at the Organ Level
To extend the model from a single cell to an organ like the liver, we aggregate the CellChrono References (C0) for all relevant cell types within that organ. Since different cells contribute differently to the organ’s overall function, a weighted average is used based on the proportion of each cell type.
The ChronoCell Matrix (CCM) at the organ level aggregates the CellChrono References from various cell types, as described in Equation (6), with each reference weighted according to its contribution to the organ’s overall function:
where:
- Csi: CellChrono Reference for each cell type within the organ.
- wi: Weight of each cell type’s contribution to organ function.
- Is and Io: Integrity of the cellular matrix or tissue under specific and reference conditions, respectively.
- γ: Empirical constant for the tissue integrity’s influence on the organ-level CCM.
3.8. Total ChronoCell Matrix (CCM) for the Human Body
The ChronoCell Matrix (CCM) for the entire human body is an aggregate of organ-based CellChrono References, yielding a temporal profile for the entire system. Each organ’s contribution is weighted based on its functional importance to achieving bodily homeostasis. The total ChronoCell Matrix (CCM) for the human body, as detailed in Equation (7), integrates the organ-level ChronoCell Matrix (CCMs), aggregating them to reflect the overall system-wide dynamics:
where:
- wj: Weight of each organ’s contribution to the human body’s overall function.
- Hs and Ho: Stable balance under specific and reference conditions, respectively.
- δ: Empirical constant for homeostatic balance’s influence on the body-level CCM.
This final formula accounts for a stable balance across the body, offering a holistic temporal profile.
3.9. ChronoVital Index (CVI): A Transformative Health Metric
The ChronoVital Index (CVI) is a groundbreaking health metric designed to evaluate the efficiency and time-based performance of organs at the cellular level. By examining the temporal dynamics within cells and tissues, the CVI can detect early signs of dysfunction or disease. For instance, a higher CVI score in a particular organ may signal slower cellular processes, which can indicate the onset of health issues.
Beyond individual organs, the CVI provides a holistic view of overall human health by aggregating data from various organs. This comprehensive assessment is invaluable for monitoring the aging process, gauging the impact of disease, and evaluating the effectiveness of medical treatment. The CVI has the potential to revolutionize personalized medicine by offering a dynamic, individualized approach to health assessment.
The ChronoVital Index (CVI) is a health metric that aggregates the temporal dynamics at both cellular and organ levels. Based on the provided formulas, CVI can be represented as an integrated function of time and the total CCM:
where:
- CCCM-Total (t): Total ChronoCell Matrix dynamics at the organ level across time.
- T: Total observation time.
3.10. ChronoBit Storage (CBS): A Novel System for Energy Modulation and Cellular Synchronization
The capability of CBS to store and release energy at nanoscale intervals allows for the fine-tuning of the energy states involved in the CellChrono Reference (C0), enhancing the precision of time dilation measurements and their influences on cellular functions. By synchronizing CBS with the natural rhythms of cellular activities, precise treatment delivery can be triggered in response to real-time physiological signals. This process enables a highly advanced form of smart medicine, where therapies are administered with pinpoint accuracy—exactly when and where they are most effective within the body. Embedding CBS into tissue engineering scaffolds can manipulate bioelectric fields to guide tissue regeneration, enhance wound healing, repair nerve damage, and support recovery from fractures by modulating bioelectric signals in real time. CBS can power bioelectronic devices and nanorobots at the cellular level, such as neural interfaces or pacemakers. In brain –computer interfaces (BCIs), CBS facilitates accelerated data transfer and interaction processes with neural systems. The rapid energy storage and release processes can improve signal processing and interactions between artificial systems and biological neurons.
Integrating CBS into the ChronoCell Matrix (CCM) and ChronoVital Index (CVI) for Advanced Temporal Regulation
The observed time T1 in a specific environment is related to the reference time T0 by the following equation,
where γ is the Lorentz factor given by
The biological stiffness constant CS is related to C0 by
- T0: Initial time parameter.
- E0/Es: Ratio of energy stored in a cell under different conditions.
- F0/Fs: Ratio of frequencies acting on the cell.
- α, β: Exponents for energy and force impact.
- ζ: Empirical constant.
The energy stored in ChronoBit Storage (CBS) is given by:
where:
- A: Amplitude of the system’s oscillation.
- ω*: Frequency of oscillation.
- ϕ: Phase constant.
- t: Time
4. Discussion
The influences of Quasi-Pathogens and Subtle Quasi-Pathogens on cellular dynamics highlight their critical roles in disrupting molecular interactions and altering DNA behaviors, which can lead to both transient and lasting cellular changes. This study emphasizes the potential of Endogenous Smart Medicine (ESM) as a therapeutic approach to mitigate these effects. ESM seeks to precisely modulate key cellular parameters like amplitude and stiffness, ensuring that pathological cycles induced by Quasi-Pathogens are prevented or minimized. By regulating induced vibrations within cells, ESM optimizes cellular environments, reducing the formation of new Quasi-Pathogen sources and promoting cellular stability. This targeted approach not only addresses the immediate effects of Quasi-Pathogens but also holds promise for managing diseases where cellular stiffness plays a critical role, such as cancer.
ChronoBit Storage (CBS) is a pivotal enhancement to ESM, as it allows for precise energy management at the nanoscale. CBS provides the ability to store and release energy in sync with cellular needs, enabling ESM to finely control cellular dynamics, especially in environments affected by Quasi-Pathogens. By integrating CBS, ESM can optimize energy distribution during therapeutic interventions, maintaining cellular homeostasis by providing timely energy bursts or modulating energy flows to restore or maintain balance.
The integration of the ChronoVital Index (CVI) within Endogenous Smart Medicine (ESM) provides a powerful tool for therapeutic intervention. The CVI enables differentiation between benign and malignant cellular dynamics by analyzing deviations in energy states, frequencies, and tissue integrity as reflected in the CellChrono References. By adjusting the variables within the CVI formulas, therapeutic interventions can be tailored to restore or optimize cellular and organ-level time dynamics. This approach holds the potential to reverse pathological conditions or enhance overall health by re-establishing cellular balance.
Further research into the mechanisms through which CBS and CVI influence Quasi-Pathogen dynamics will be essential to advancing therapeutic strategies. This research can deepen our understanding of cellular interactions and enable more precise interventions to restore cellular equilibrium and promote long-term health.
5. Conclusions
This study provides valuable insights into how induced vibrations contribute to the formation of Endogenous Quasi-Pathogens (EQPs) and their significant impacts on cellular dynamics. Understanding these processes is essential for developing innovative diagnostic and therapeutic strategies for diseases driven by alterations in cellular behavior and tissue stiffness.
Endogenous Smart Medicine (ESM), enhanced by ChronoBit Storage (CBS), emerges as a promising intervention, offering precise control over cellular impedance and mitigating the disruptive effects of EQPs. CBS allows for the real-time modulation of cellular energy states, ensuring that therapeutic interventions are optimally timed and effective. This dynamic control of energy provides new possibilities for managing complex diseases such as cancer, where cellular stiffness and time dynamics are disrupted.
The ChronoVital Index (CVI) integrated with CBS serves as a powerful model for capturing the temporal dynamics within biological systems. By refining CVI formulas and incorporating CBS into ESM, the capacity to manipulate these dynamics for therapeutic purposes is greatly enhanced. Future research should focus on further exploring the interactions between CBS systems, induced vibrations, EQPs, and cellular environments. Such investigations are crucial for advancing our understanding of these phenomena and their broader implications for human health.
Author Contributions
Conceptualization, F.A.; methodology, F.A.; software; validation, J.-M.S. and F.A.; formal analysis, F.A.; investigation, F.A.; resources, F.A.; data curation, F.A.; writing—original draft preparation, F.A.; writing—review and editing, F.A. and J.-M.S.; visualization, F.A.; supervision, J.-M.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sabatier, M.J.; Amini, F. Emergence of Gloomy Eyelet inside DNA. Biophysica 2023, 3, 35–45. [Google Scholar] [CrossRef]
- Sabatier, M.J.; Amini, F. Exploring the Impact of Induced Vibrations and Gloomy Eyelets within DNA on Pathogen Behavior: Insights into Quasi-Pathogens. Austin J. Microbiol. 2024, 1–3. [Google Scholar]
- Richoux, O.; Lombard, B.; Mercier, J. Generation of acoustic solitary waves in a lattice of Helmholtz resonators. Wave Motion 2015, 56, 85–99. [Google Scholar] [CrossRef]
- Smith, H. A Thermoacoustic Approach to Exploring Resonance; University of Central Arkansas: Conway, AR, USA, 2008. [Google Scholar]
- Petrov, G.A. Rotation of the Apparent Vibration Plane of a Swinging Spring at the 1:1:2 Resonance. Mech. Solids 2017, 52, 243–253. [Google Scholar] [CrossRef]
- Ridgely, T.C. On the Origin of Inertia. Galilean Electrodyn. 2001, 12, 17–19. [Google Scholar]
- Sabatier, M.J.; Amini, F. Gloomy Eyelet Along with the Latent Buoyancy Inside Nucleus Cell. Lupine Online J. Med. Sci. 2023, 640–642. [Google Scholar] [CrossRef]
- Sabatier, M.J.; Amini, F. Coupling Gloomy Eyelets in the Nucleus of Cells and Forming Unknown Distinct Structure (UDS). Austin J. Anal. Pharm. Chem. 2023, 10, 1155. [Google Scholar] [CrossRef]
- Lakes, R. Viscoelasticity Notes Adapted from Viscoelastic Materials; University of Wisconsin, Rod Lakes Home: Madison, WI, USA, 2009. [Google Scholar]
- Li, M.; Li, D.; Wang, L.J.; Adhikari, B. Creep behavior of starch-based nanocomposite films with cellulose nanofibrils. Carbohydr. Polym. 2014, 117, 957–963. [Google Scholar] [CrossRef] [PubMed]
- Dealy, M.J. Dynamic Mechanical Analysis in the Analysis of Polymers and Rubbers. In Encyclopedia of Polymer Science and Technology; John Wiley & Sons: Hoboken, NJ, USA, 2015. [Google Scholar] [CrossRef]
- Smith, A.B.; Jones, C.D. Fundamentals of Oscillator Impedance: Theory and Applications; Cambridge University Press: Cambridge, UK, 2015. [Google Scholar]
- Felicetti, M.A.; Piantino, F.; Coury, J.R.; Aguiar, M.L. Influences of Removal Time and Particle Size on the Particle Substrate Adhesion Force. Braz. J. Chem. Eng. 2008, 25, 71–82. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).