
Response of Herbaceous and Woody Plant Species in Southern Portugal to Cope Oak Decline Associated to Phytophthora cinnamomi

 ,
,  and
and
Abstract
1. Introduction
2. Material and Methods
2.1. Inoculum and Preparation of Soil and Plants
2.2. Plant Response to P. cinnamomi
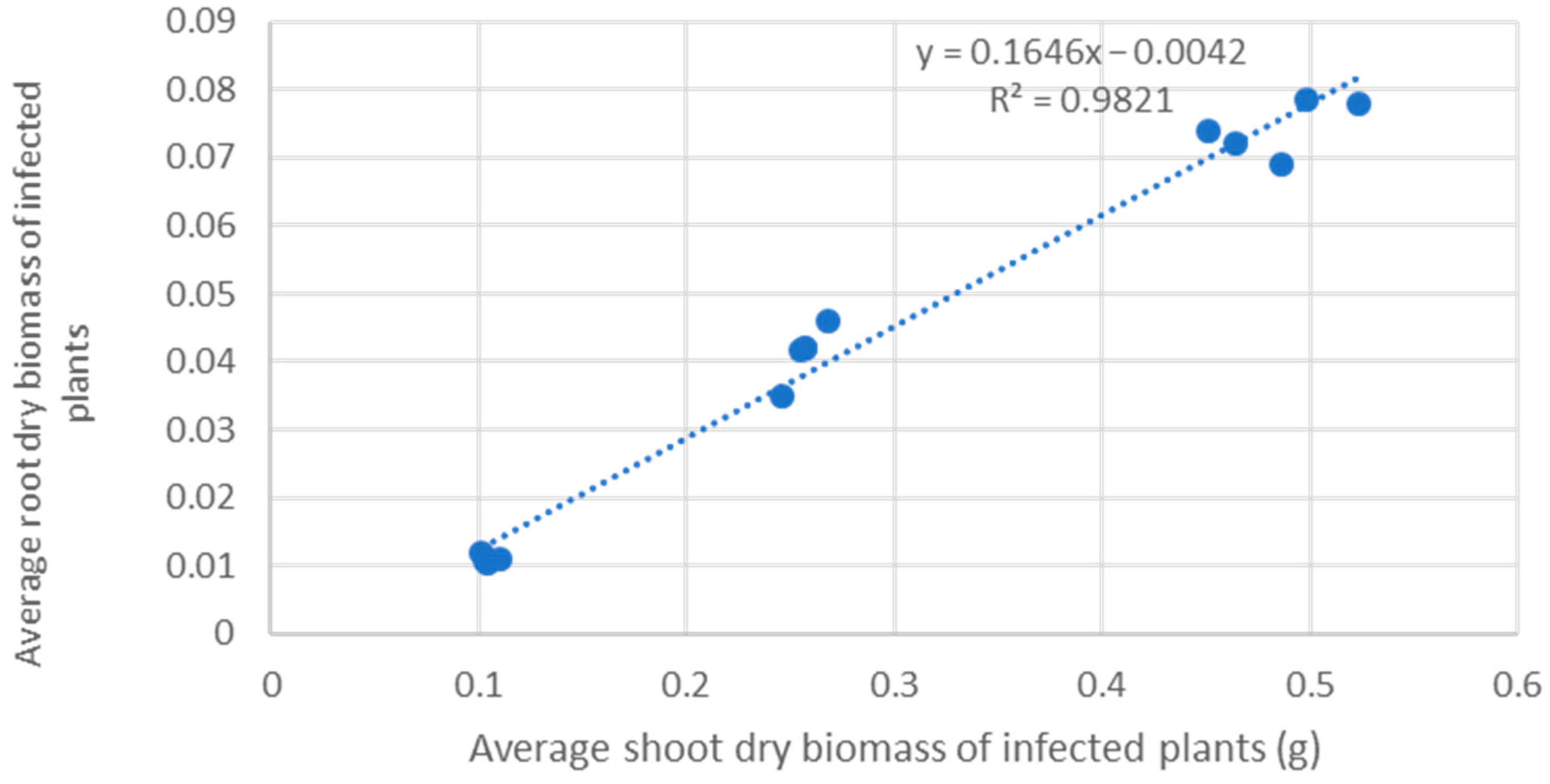
3. Results
3.1. Susceptibility of Woody Species
3.1.1. Trees
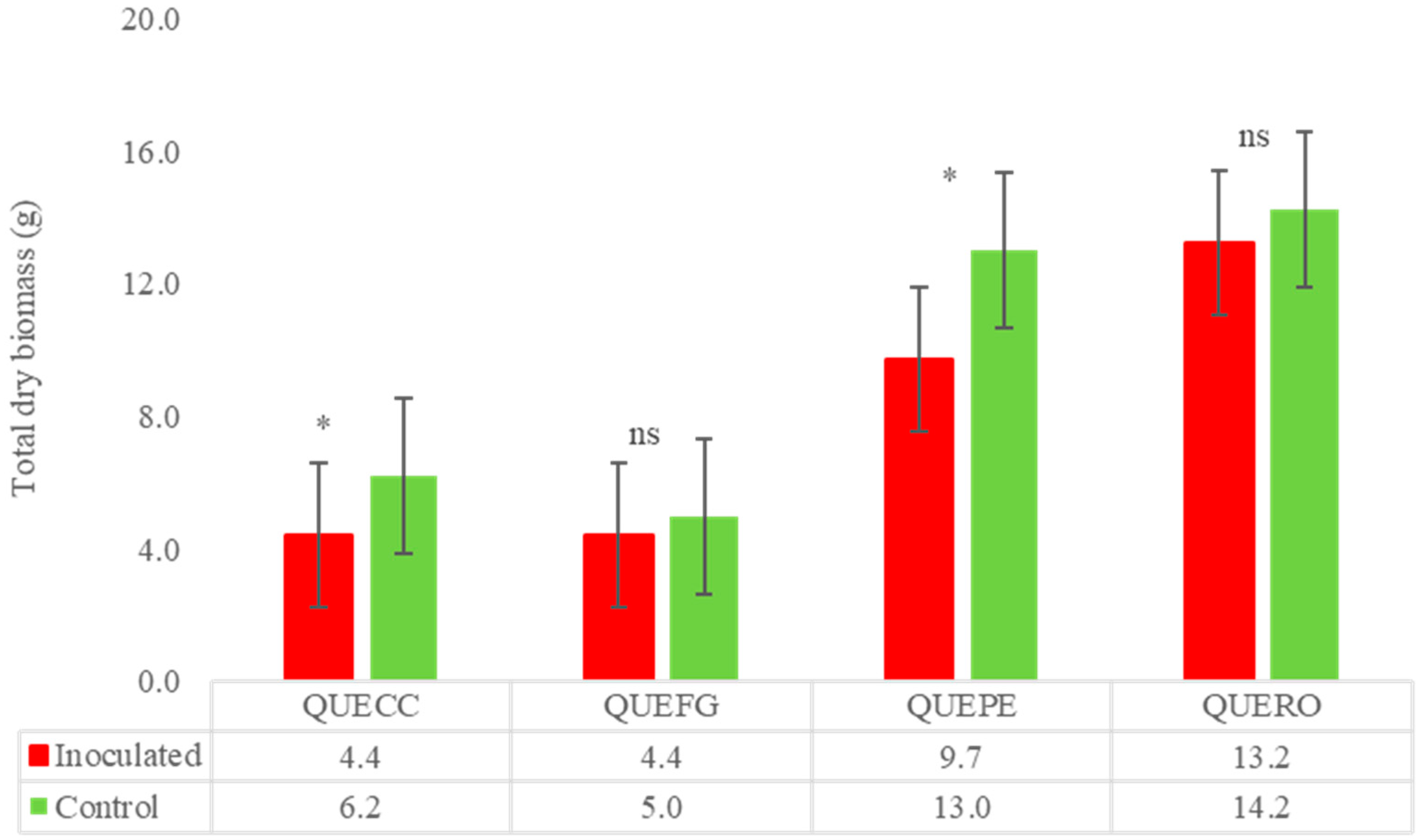
Quercus spp.
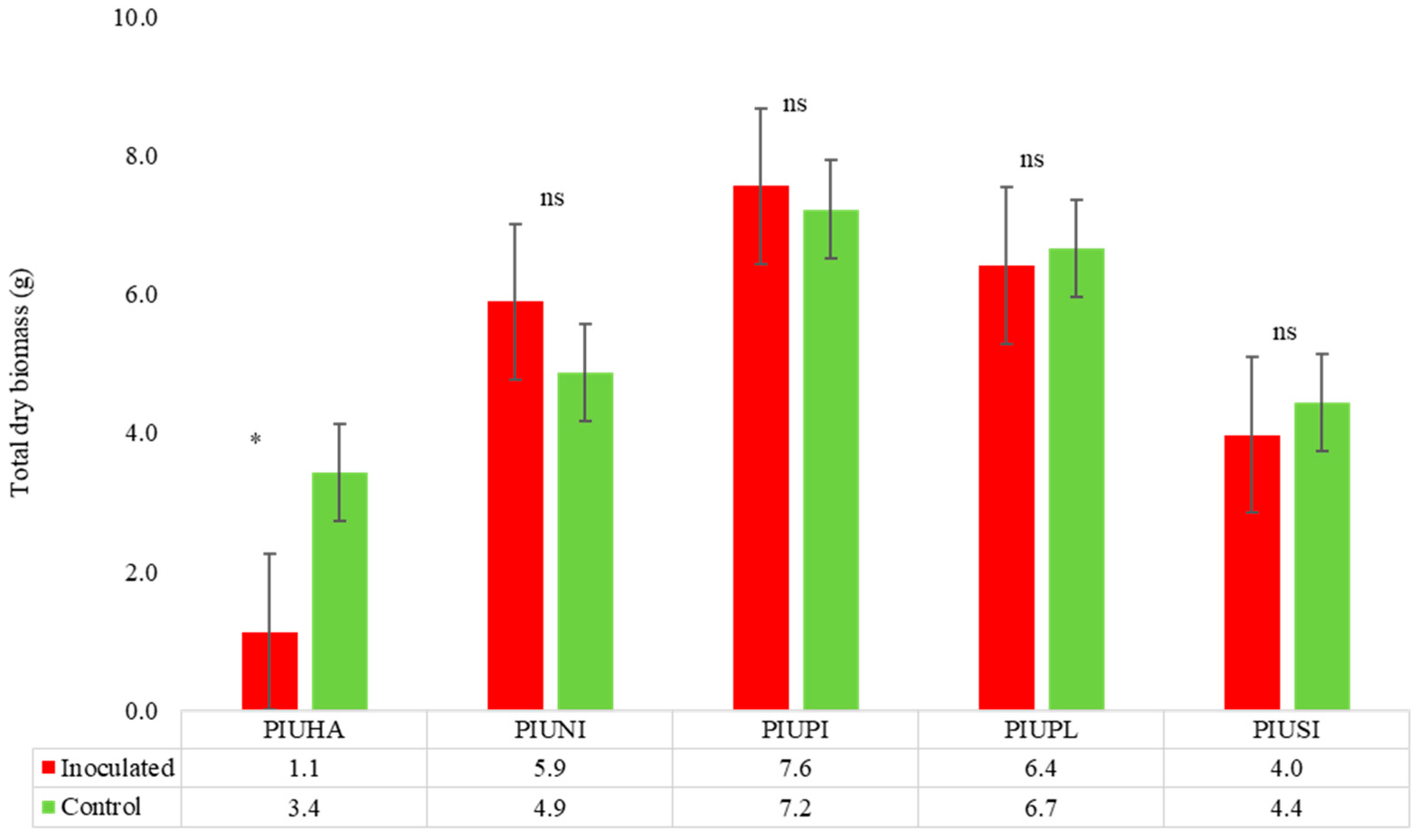
Pinus spp.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | a Mortality (%) | b Inoculated Plants Nº | Dead Nº | c Infection Reaction | Control Nº | d Total Biomass Reduction (%) | e Category of Susceptibility |
|---|---|---|---|---|---|---|---|
| Quercus coccifera | 80.0 | 15 | 12 | Symp-host | 15 | 29.0 | HS |
| Quercus faginea | 0 | 15 | 0 | Symp-host/Asymp host field | 15 | 11.2 | SS |
| Quercus petrae | 0 | 15 | 0 | Symp-host | 15 | 25.2 | MS |
| Quercus robur | 0 | 15 | 0 | Symp-host | 15 | 6.8 | SS |
| Pinus halepensis | 67.0 | 15 | 10 | Symp-host | 15 | 67.1 | HS |
| Pinus nigra | 0 | 15 | 0 | Asymp host | 15 | No-reduction | HT |
| Pinus pinea | 6.7 | 15 | 1 | Symp-host | 15 | No-reduction | MS |
| Pinus pinaster | 20 | 15 | 3 | Symp-host/Asymp host field | 15 | 3.8 | MS |
| Pinus sylvestris | 0 | 15 | 0 | Symp-host | 15 | 10.6 | SS |
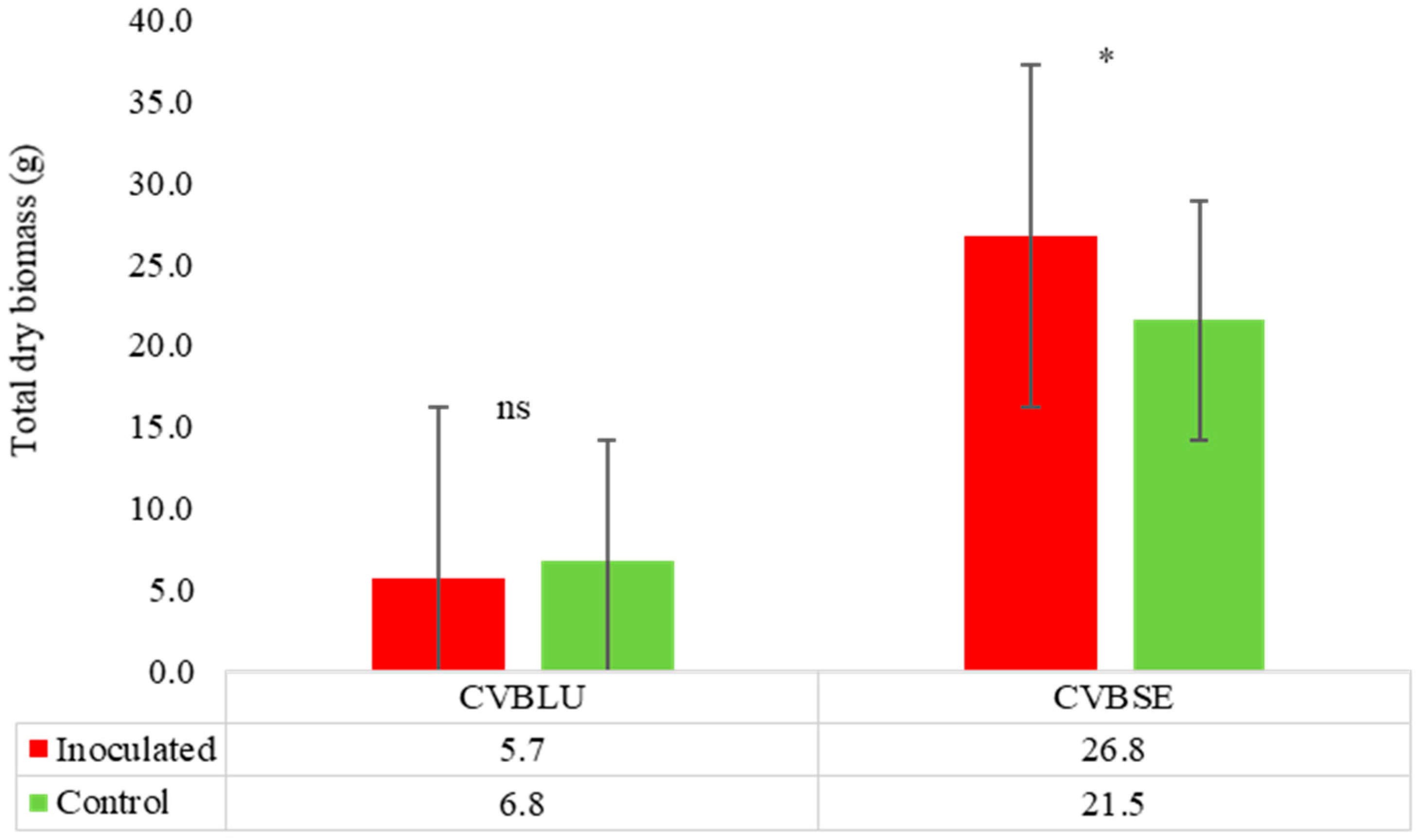
| Cupressus lusitanica | 0 | 15 | 0 | Asymp-host | 15 | 15.8 | T |
| Cumpressus sempervirens | 0 | 15 | 0 | Asymp-host | 15 | No-reduction | HT |
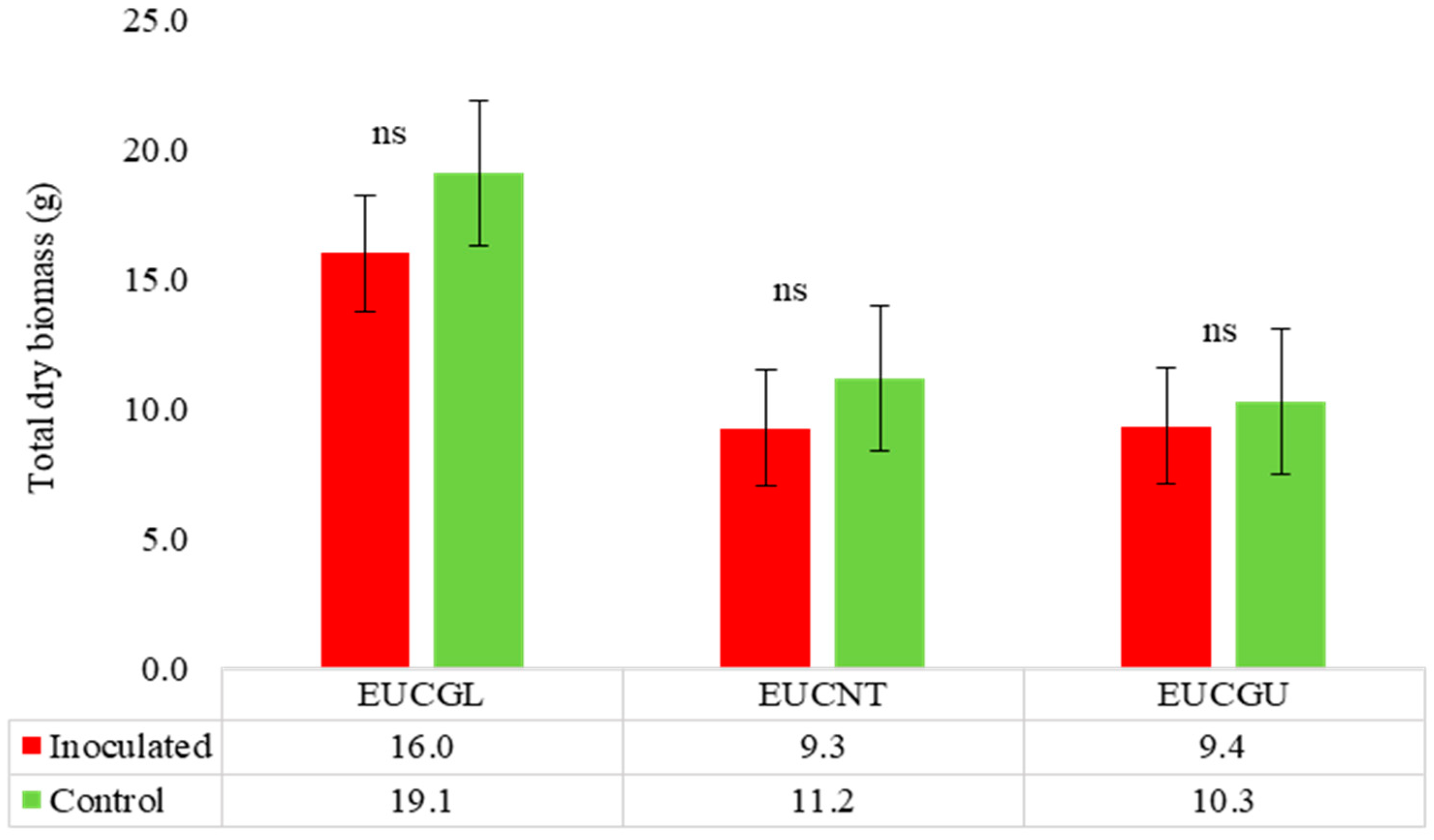
| Eucalyptus globulus | 0 | 15 | 0 | Symp-host | 15 | 16.1 | T |
| Eucalyptus guinnii | 0 | 15 | 0 | Symp-host | 15 | 8.7 | SS |
| Eucalyptus nitens | 0 | 15 | 0 | Symp-host | 15 | 17.0 | SS |
| Cistus albidus | 75.0 | 8 | 6 | Symp-host | 8 | 57.2 | HS |
| Cistus ladanifer | 87.5 | 8 | 7 | Symp-host | 8 | 36.4 | HS |
| Cistus monspeliensis | 12.5 | 8 | 1 | Symp-host | 8 | 12.9 | MS |
| Lavandula dentata | 0 | 8 | 0 | Asymp-host | 8 | 5.8 | T |
| Myrtus communis | 12.5 | 8 | 1 | Symp-host | 8 | 49.9 | MS |
Cupressus spp.
Eucalyptus spp.
3.1.2. Shrub Species
3.2. Susceptibility of Herbaceous Species
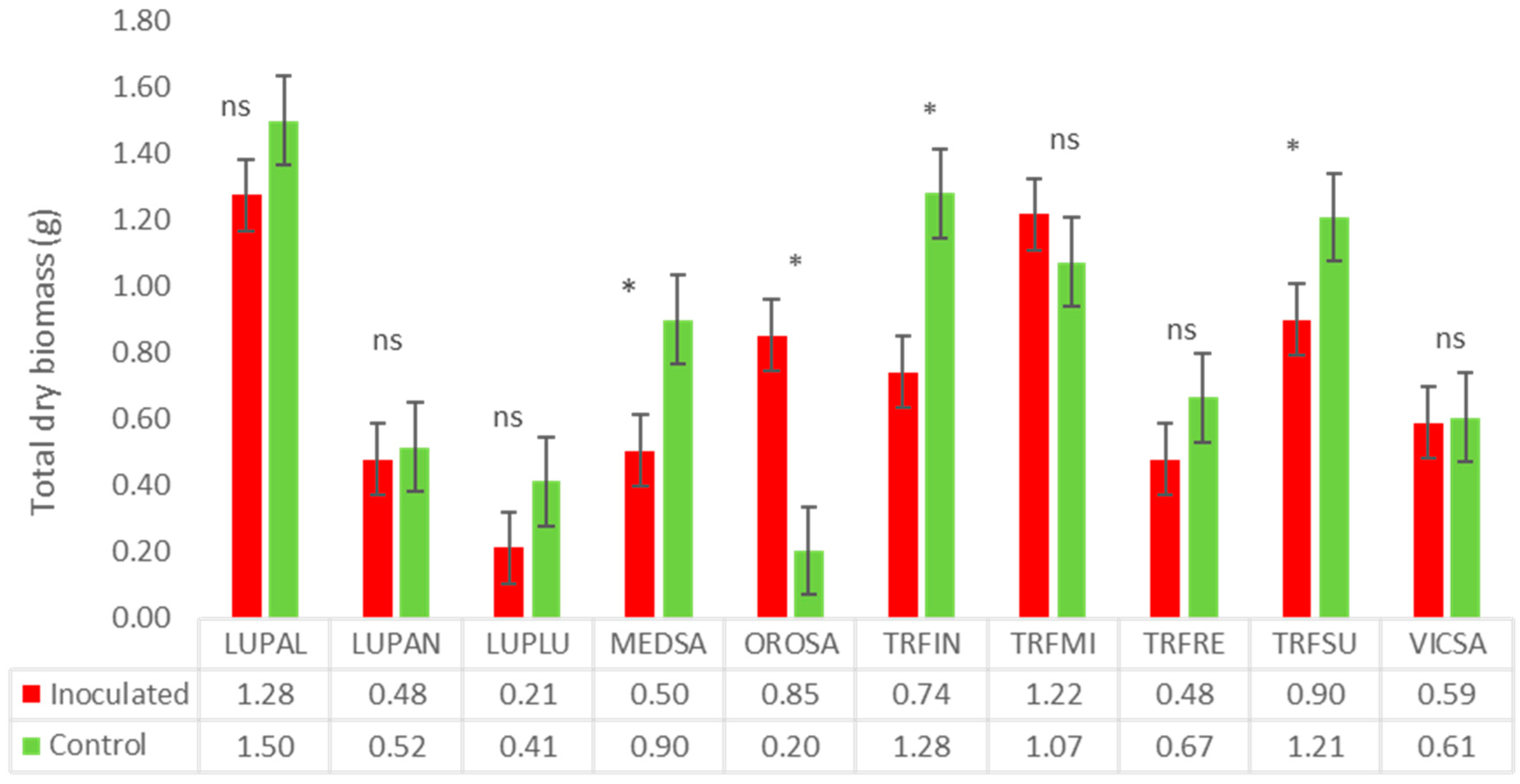
3.2.1. Fabaceae Family
3.2.2. Poaceae Family
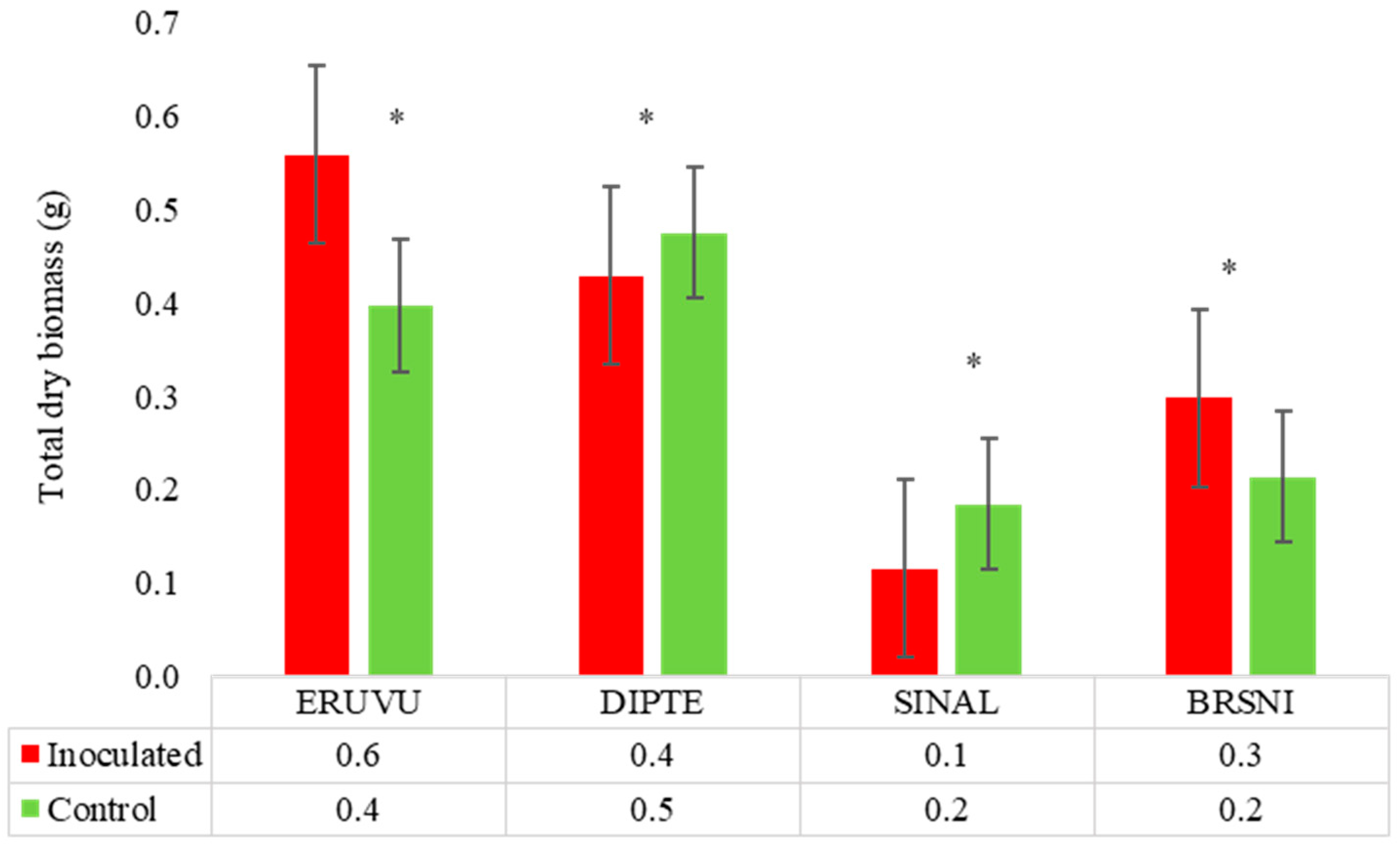
3.2.3. Brassicaceae Family
| Species | a Mortality (%) | b Inoculated Plants Nº | c Infection Reaction | Control Plants Nº | d Root Biomass Reduction (%) | e Shoot Biomass Reduction (%) | f Colonies Number cfu/g Soil | g Category of Susceptibility |
|---|---|---|---|---|---|---|---|---|
| Trifolium incarnatum | 0 | 98 | Symp-no-host | 98 | 76.2 | 13.5 | 5 | SS |
| T. repens | 0 | 80 | Asymp-no-host | 99 | 2.8 | 32.4 | 17 | SS |
| T. subterraneum | 0 | 112 | Asymp-no-host | 112 | 43.1 | 8.9 | 12 | SS |
| T. michelianum | 0 | 90 | Asymp-no-host | 90 | 11.7 | No reduction | 20 | SS |
| Lupinus angustifolius | 0 | 36 | Asymp-host | 27 | 14.4 | 5.6 | 162 | SS |
| L. luteus | 56.0 | 18 | Symp-host | 22 | 40.6 | 49.5 | 49 | HS |
| L. albus | 0 | 18 | Symp-host | 24 | 2.7 | 9.52 | 2 | SS |
| Ornithopus sativus | 18.0 | 56 | Symp-host | 109 | No reduction | No reduction | 61 | MS |
| Vicia sativa | 0 | 72 | Asymp-no-host | 81 | No reduction | 2.9 | 21 | T |
| Medicago sativa | 0 | 72 | Symp-no-host | 72 | 61.1 | 19.6 | 23 | SS |
| Lolium multiflorum | 0 | 128 | Asymp-no-host | 92 | 31.0 | 31.0 | 13 | SS |
| L. rigidum | 0 | 93 | Asymp-no-host | 93 | 21.1 | 24.3 | 28 | SS |
| L. perenne | 0 | 86 | Asymp-no-host | 86 | 29.9 | 29.9 | 8 | SS |
| Dactylis glomerata | 0 | 73 | Symp-no-host | 73 | 59.3 | 59.3 | 22 | SS |
| Festuca arundinacea | 0 | 80 | Asymp-no-host | 80 | 24.1 | 24.1 | 5 | SS |
| Brachypodium distachyon | 0 | 67 | Asymp-host | 126 | 5.6 | 5.6 | 22 | SS |
| Hordeum murinum | 0 | 53 | Asymp-host | 102 | 12.4 | No reduction | 2 | SS |
| Secale cereale * | 0 | 17 | Asymp-no-host | 19 | No reduction | No reduction | 125 | T |
| Eruca vesicaria | 0 | 44 | Asymp-no-host | 47 | 8.2 | No reduction | 23 | T |
| Diplotaxis tenuifolia | 0 | 38 | Asymp-no-host | 38 | 64.0 | No reduction | 23 | T |
| Sinapis alba | 0 | 145 | Symp-host | 100 | 35.0 | 35.0 | 3 | SS |
| Brassica nigra * | 0 | 43 | Asymp-no-host | 53 | No reduction | No reduction | 85 | T |
4. Discussion
4.1. Response of Woody Species
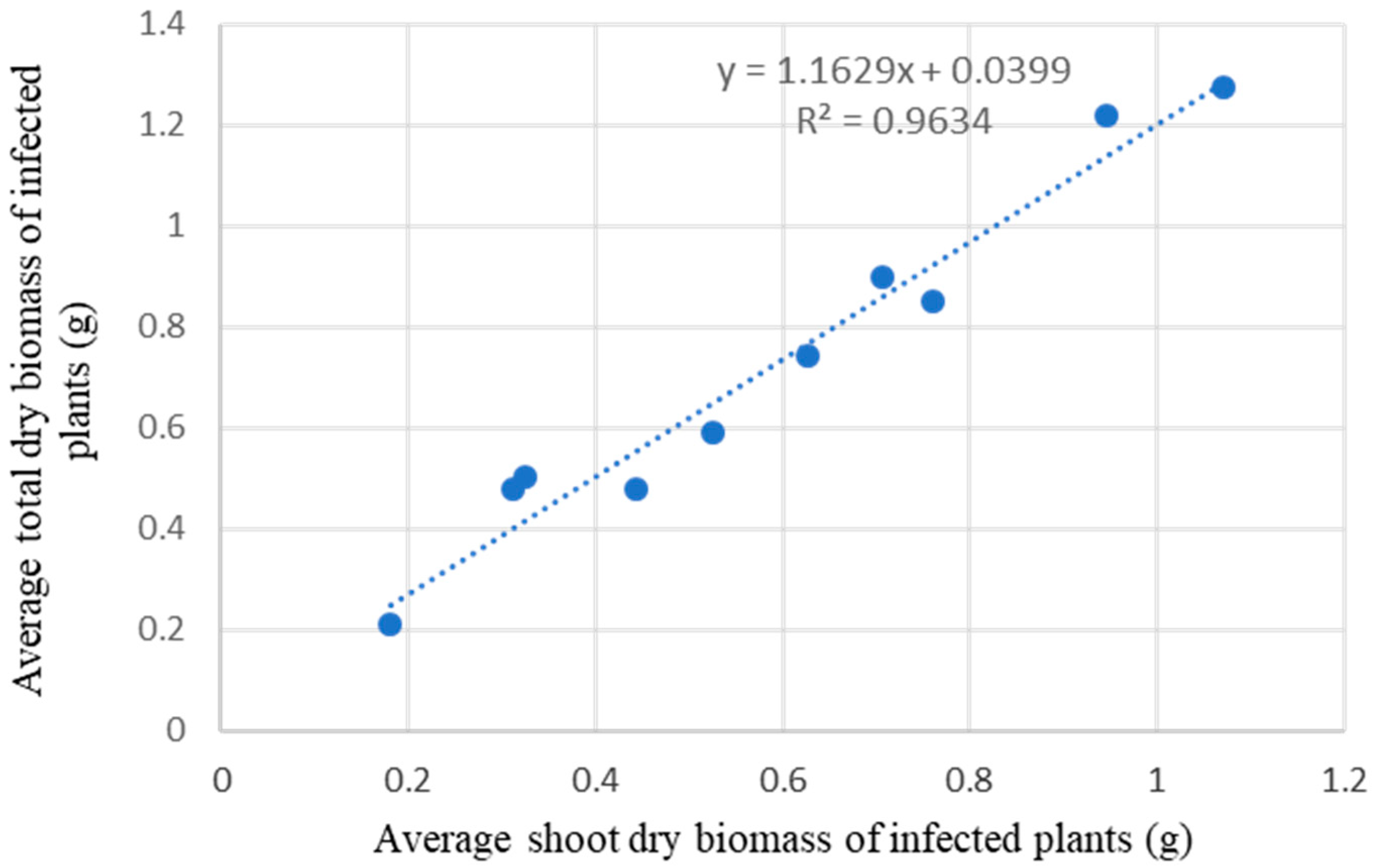
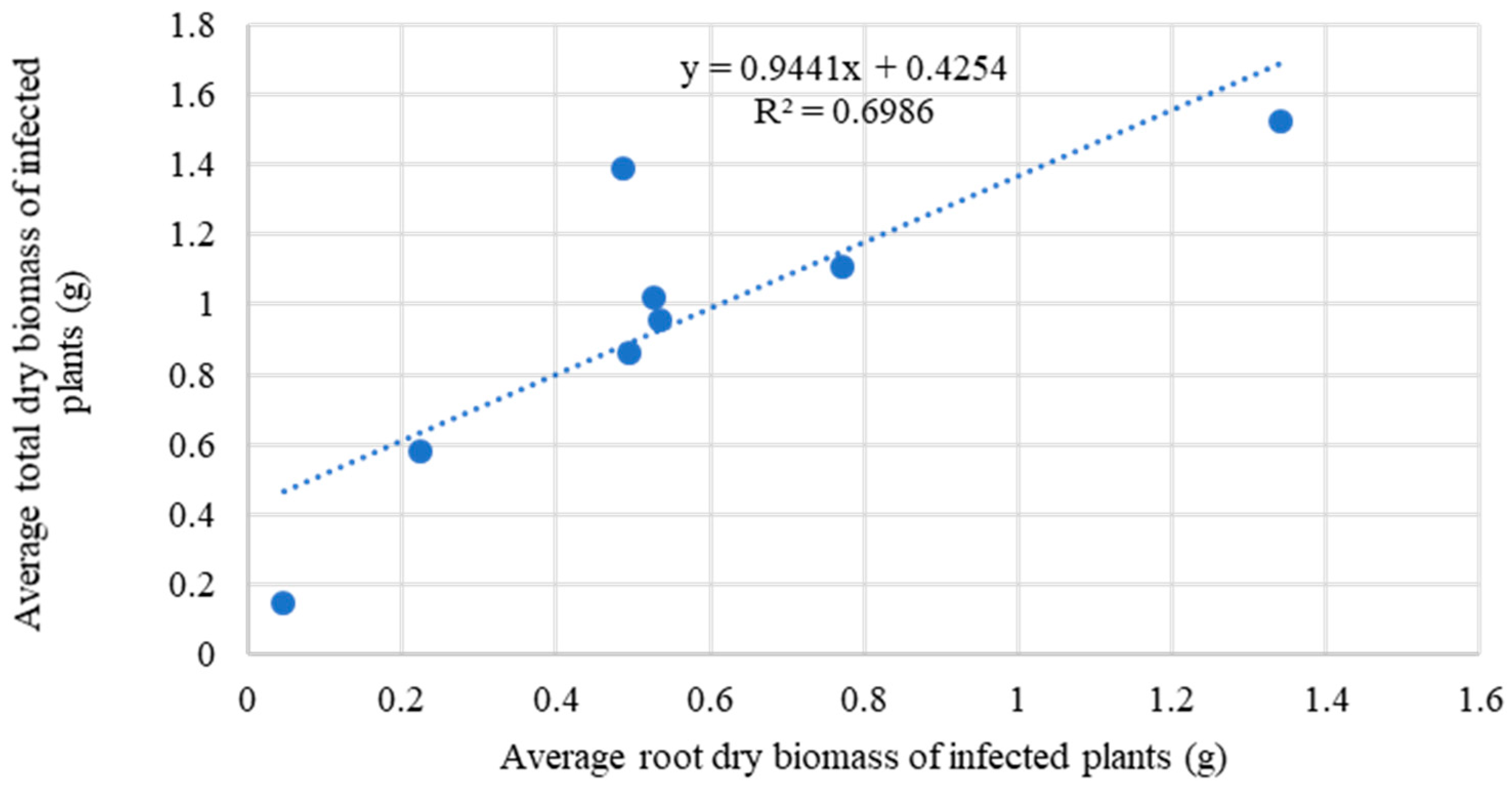
4.2. Response of Herbaceous Species
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Simonson, W.D.; Allen, H.D.; Parham, E.; Basto e Santos, E.; Hotham, P. Modelling biodiversity trends in the montado (wood pasture) landscapes of the Alentejo, Portugal. Landsc. Ecol. 2018, 33, 811–827. [Google Scholar] [CrossRef]
- Simões, M.P.; Belo, A.F.; Fernandes, M.; Madeira, M. Regeneration patterns of Quercus suber according to Montado management systems. Agrofor. Syst. 2015, 90, 107–115. [Google Scholar] [CrossRef]
- Pereira, J.S.P.; Correia, A.; Correia, A.; Borges, J.G. Uma Avaliação dos ecossistemas de Portugal Floresta. In Ecossistemas e Bem–Estar Humano; Pereira, H.M., Domingos, T., Vicente, L., Eds.; Escolar Editora: Lisboa, Portugal, 2009; Capítulo 6; pp. 183–211. ISBN 978-972-592-274-3. [Google Scholar]
- Capelo, J.; Aguiar, C. A Vegetação de Portugal, 1st ed.; Imprensa Nacional-Casa da Moeda (A.INCM): Lisboa, Portugal, 2021; 335p, ISBN 978-972-27-2879-9. [Google Scholar]
- Brasier, C.M. Oak tree mortality in Iberia. Nature 1992, 360, 539. [Google Scholar] [CrossRef]
- Brasier, C.M.; Robredo, F.; Ferraz, J.P.F. Evidence for Phytophthora cinnamomi involvement in Iberian oak decline. Plant Pathol. 1993, 42, 140–145. [Google Scholar] [CrossRef]
- Moreira, A.C.; Martins, J.M. Influence of site factors on the impact of Phytophthora cinnamomi in cork oak stands in Portugal. For. Pathol. 2005, 35, 154–162. [Google Scholar] [CrossRef]
- Hardham, A.R.; Blackman, L.M. Phytophthora cinnamomi. Mol. Plant Pathol. 2018, 19, 260–285. [Google Scholar] [CrossRef]
- Moreira, A.C.; Calha, I.; Rodriguez-Romero, M.; Costa, A.; Neno, J.; Passarinho, J.A. A Vegetação dos Montados em Areas Afetadas por Declínio—Situação de Ourique; Moreira, A.C., Ed.; INIAV, I.P.: Oeiras, Portugal, 2023; 75p, ISBN 978-972-579-074-8. Available online: https://www.iniav.pt/divulgacao/publicacoes-bd/a-vegetacao-dos-montados-em-areas-afetadas-por-declinio-situacao-de-ourique (accessed on 22 August 2024).
- Erwin, D.C.; Ribeiro, O.K. Phytophthora Diseases Worldwide; APS Press: St. Paull, MN, USA, 1996; 562p. [Google Scholar]
- Dunstan, W.A.; Rudman, T.; Nr, B.L.; Moore, N.A.; Paap, T.; Calver, M.C.; Hardy, G.E.S.J. Containment and spot eradication of a highly destructive, invasive plant pathogen (Phytophthora cinnamomi) in natural ecosystems. Biol. Invasions 2010, 12, 913–925. [Google Scholar] [CrossRef]
- Zentmyer, G.A. Phytophthora cinnamomi and the Diseases It Causes; American Phytopathological Society: St. Paul, MN, USA, 1980. [Google Scholar]
- Little, T.J.; Shuker, D.M.; Colegrave, N.; Day, T.; Graham, A.L. The Coevolution of Virulence: Tolerance in Perspective. PLoS Pathog. 2010, 6, e1001006. [Google Scholar] [CrossRef]
- Clarke, D.D. Tolerance of parasites and disease in plants and its significance in host-parasite interactions. Adv. Plant Pathol. 1986, 5, 161–198. [Google Scholar]
- Pagán, I.; García-Arenal, F. Tolerance to Plant Pathogens: Theory and Experimental Evidence. Int. J. Mol. Sci. 2018, 19, 810. [Google Scholar] [CrossRef]
- Stowe, K.A.; Marquis, R.J.; Hochwender, C.G.; Simms, E.L. The Evolutionary Ecology of Tolerance to Consumer Damage. Annu. Rev. Ecol. Syst. 2000, 31, 565–595. [Google Scholar] [CrossRef]
- Moreira-Marcelino, A.C. Aspectos da Interacção Entre Phytophthora cinnamomi e a Doença do Declínio em Q. suber e Q. rotundifolia. Ph.D. Thesis, Faculdade de Engenharia de Recursos Naturais, Universidade do Algarve, Faro, Portugal, 2001; 279p. [Google Scholar]
- Sampaio, A.R.B. Selecção de Plantas com Efeito Alelopático para Controlar Phytophthora cinnamomi. Master’s Thesis, Universidade Técnica de Lisboa, ISA, Lisboa, Portugal, 2017; 73p. [Google Scholar]
- Kueh, K.H.; McKay, S.F.; Facelli, E.; Facelli, J.M.; Velzeboer, R.M.A.; Able, A.J.; Scott, E.S. Response of selected South Australian native plants species to Phytophthora cinnamomi. For. Pathol. 2012, 61, 1165–1178. [Google Scholar] [CrossRef]
- Moreira, A.; Calha, I.; Passarinho, J.; Sampaio, A. Anti-Phytophthora activity of root extracts from herbaceous species. Rev. Ciências Agrárias 2018, 41, 39–47. [Google Scholar]
- Rios, P.; Obregón, S.; Gonzaléz, M.; Haro, A.; Sánchez, M.E. Screening brassicaceous plants as biofumigants for management of Phytophthora cinnamomi oak disease. For. Pathol. 2015, 46, 618. [Google Scholar] [CrossRef]
- Shearer, B.L.; Dillon, M. Susceptibility of plant species in Eucalyptus marginata forest to infection by Phytophthora cinnamomi. Aust. J. Bot. 1995, 43, 113–134. [Google Scholar] [CrossRef]
- McDougall, K.L.; Hobbs, R.J.; Hardy, G.E.S.J. Distribution of understorey species in forest affected by Phytophthora cinnamomi in south-western Western Australia. Aust. J. Bot. 2005, 53, 813–819. [Google Scholar] [CrossRef]
- Robertson, G.I. Susceptibility of exotic and indigenous trees and shrubs to Phytophthora cinnamomi Rands. N. Z. J. Agric. Res. 1970, 13, 297–307. [Google Scholar] [CrossRef]
- Balci, Y.; Halmschlager, E. Phytophthora species in oak ecosystems in Turkey and their association with declining oak trees. Plant Pathol. 2003, 52, 694–702. [Google Scholar] [CrossRef]
- Gea−Izquierdo, G.; Férriz, M.; García-Garrido, S.; Aguín, O.; Elvira-Recuenco, M.; Escribano, L.H.; Benito, D.M.; Raposo, R. Synergistic abiotic and biotic stressors explain widespread decline of Pinus pinaster in a mixed forest. Sci. Total Environ. 2019, 685, 963–975. [Google Scholar] [CrossRef]
- Pérez-Sierra, A.; Lopez-Garcia, C.; Leon, M.; Garcia-Jimenez Abad-Campos, P.; Jung, T. Previously unrecorded Low-temperature Phytophthora species associated with Quercus decline in a Mediterranean forest in eastern Spain. For. Pathol. 2013, 43, 331–339. [Google Scholar] [CrossRef]
- Moreira, A.C.; Tapias, R.; Fernandes, L.; Rodrigues, A. Field susceptibility of cork oak trees with different provenances to Phytophthora cinnamomi. For. Pathol. 2018, 48, e12461. [Google Scholar] [CrossRef]
- Wardlaw, T.J.; Palzer, C. Regeneration of Eucalyptus Species in an Eastern Tasmanian Coastal Forest in the Presence of Phytophthora cinnamomi. Aust. J. Bot. 1988, 36, 205–215. [Google Scholar] [CrossRef]
- Bregant, C.; Batista, E.; Hilário, S.; Linaldeddu, B.T.; Alves, A. First report of Phytophthora hibernalis, P. multivora and P. niederhauserii causing root rot and bleeding cankers on Eucalyptus globulus in Portugal. New Dis. Rep. 2023, 47, e12171. [Google Scholar] [CrossRef]
- Eschen, R. Introduction. In Field Guide for the Identification of Damage on Woody Sentinel Plants; Roque, A., Claeary, M., Matsiakh, I., Eschen, R., Eds.; CABI: Oxfordshire, UK, 2017; 292p, ISBN-13 978 1 78639 440 8. [Google Scholar]
- Moreira, A.C.; Rodrigues, A. Effect of Soil Water Content and Soil Texture on Phytophthora cinnamomi Infection on Cork and Holm Oak. Silva Lusit. 2021, 29, 133. [Google Scholar] [CrossRef]
- Phillips, D.; Weste, G. Field resistance in three native monocotyledon species that colonize indigenous sclerophyll forest after invasion by Phytophthora cinnamomi. Aust. J. Bot. 1984, 32, 339–352. [Google Scholar] [CrossRef]
- D’Souza, N.K.; Colquhoun, I.J.; Shearer, B.L.; Hardy, G.E.S.J. Assessing the potential for biological control of Phytophthora cinnamomi by fifteen native Western Australian jarrah-forest legume species. Aust. Plant Pathol. 2005, 34, 533–540. [Google Scholar] [CrossRef]
- Masini, L.; Grenville-Briggs, L.J.; Andreasson, E.; Råberg, L. Tolerance and overcompensation to infection by Phytophthora infestans in the wild perennial climber Solanum dulcamara. Ecol. Evol. 2019, 9, 4557–4567. [Google Scholar] [CrossRef]
- Homet, P.; González, M.; Matías, L.; Godoy, O.; Pérez-Ramos, I.M.; García, L.V.; Gómez-Aparicio, L. Exploring interactive effects of climate change and exotic pathogens on Quercus suber performance: Damage caused by Phytophthora cinnamomi varies across contrasting scenarios of soil moisture. Agric. For. Meteorol. 2019, 276–277, 107605. [Google Scholar] [CrossRef]
- Nilsson, M.C.; Wardle, D. Understory Vegetation as a Forest Ecosystem Driver. Front. Ecol. Environ. 2005, 3, 421–428. [Google Scholar] [CrossRef]
- Wu, J.P.; Liu, Z.F.; Wang, X.L.; Sun, Y.X.; Zhou, L.X.; Lin, Y.B.; Fu, S.L. Effects of understory removal and tree girdling on soil microbial community composition and litter decomposition in two Eucalyptus plantations in South China. Funct. Ecol. 2011, 25, 921–931. [Google Scholar] [CrossRef]
- Crone, M.; McComb, J.A.; O’Brien, P.A.; Hardy, G.E.S.J. Assessment of Australian native annual/herbaceous perennial plant species as asymptomatic or symptomatic hosts of Phytophthora cinnamomi under controlled conditions. For. Pathol. 2013, 43, 245–251. [Google Scholar] [CrossRef]
- Serrano, M.S.; Fernández-Rebollo, P.; De Vita, P.; Sánchez, M.E. Susceptibility of common herbaceous crops to Phytophthora cinnamomi and its influence on Quercus root rot in rangelands. Eur. J. Plant Pathol. 2012, 134, 409–414. [Google Scholar] [CrossRef]
- Fernández, P.; Carbonero, M.D.; Sánchez, M.E.; Trapero, A. Cómo manejar las explotaciones afectadas por decaimento florestal? Eur. Cork 2008, 36, 40–46. [Google Scholar]
- Serrano, M.S.; Fernández-Rebollo, P.; De Vita, P.; Carbonero, M.D.; Trapero, A.; Sánchez, M.E. The role of yellow lupin (Lupinus luteus) in the decline affecting oak agroforestry ecosystems. For. Pathol. 2011, 41, 382–386. [Google Scholar] [CrossRef]
- Rodríguez-Romero, M.; Godoy-Cancho, B.; Calha, I.; Passarinho, J.A.; Moreira, A.C. Allelopathic Effects of Three Herb Species on Phytophthora cinnamomi, a Pathogen Causing Severe Oak Decline in Mediterranean Wood Pastures. Forests 2021, 12, 285. [Google Scholar] [CrossRef]
- Agrawal, A.A. Host-range Evolution: Adaptation and trade-offs in fitness of mites on alternative hosts. Ecology 2000, 81, 500–508. [Google Scholar] [CrossRef]
- Jayasekera, A.U.; McComb, J.A.; Shearer, B.; Hardy, G. Suppression of Phytophthora cinnamomi by the root exudates of Acacia pulchella. In Progress in Research on Phytophthora Diseases of Forest Trees; Brasier, C., Jung, T., Oßwald, W., Eds.; Forest Research: Farnham, UK, 2006; pp. 92–94. [Google Scholar]
- Bais, H.P.; Weir, T.L.; Perry, L.G.; Gilroy, S.; Vivanco, J.M. The role of root exudates in rhizosphere interactions with plants and others organisms. Ann. Rev. Plant Biol. 2006, 57, 233–266. [Google Scholar] [CrossRef]
- Ríos, P.; Obregón, S.; Haro, A.; Fernández-Rebollo, P.; Serrano, M.; Sánchez, M.E. Effect of Brassica biofumigant amendments on different stages of the life cycle of Phytophthora cinnamomi. J. Phytopathol. 2016, 164, 582–594. [Google Scholar] [CrossRef]
- Serrano, M.S.; Romero, M.A.; Homet, P.; Gómez-Aparicio, L. Climate change impact on the population dynamics of exotic pathogens: The case of the worldwide pathogen Phytophthora cinnamomi. Agric. For. Meteorol. 2022, 322, 109002. [Google Scholar] [CrossRef]
- Serrano, M.S.; Villa-Sanabria, H.; Homet, P.; Gutiérrez Gómez-Aparicio, L. Impact of a drier climate on the exotic pathogen Phytophthora cinnamomi in Mediterranean forests differing in soil properties and species composition. For. Ecol. Manag. 2024, 556, 121721. [Google Scholar] [CrossRef]
- Brasier, C.M.; Scott, J.K. European oak declines and global warming: A theoretical assessment with special reference to the activity of Phytophthora cinnamomi. EPPO Bull. 1994, 24, 221–232. [Google Scholar] [CrossRef]


| Species | Common Name | Family | EPPO Code |
|---|---|---|---|
| Batch 1-Inoculated in Spring Woody plants: shrubs | |||
| Cistus ladanifer L. | Rockrose | Cistaceae | CSTLA |
| Cistus albidus L. | Big rosete | Cistaceae | CSTAL |
| Cistus monspeliensis L. | Montpellier rockrose | Cistaceae | CSTMO |
| Myrtus communis L. | Myrtle | Myrtaceae | MYVCO |
| Lavandula dentata L. | Lavender | Lamiaceae | LAUDE |
| Woody plants: trees | |||
| Quercus coccifera L. | Kermes oak | Fagaceae | QUECC |
| Quercus petrae (Mattuschka) Liebl. | Sessile oak | Fagaceae | QUEPE |
| Quercus robur L. | Oak | Fagaceae | QUERO |
| Quercus faginea Lam. | Oak fence | Fagaceae | QUEFG |
| Cupressus sempervirens L. | Mediterranean cypress | Cupressaceae | CVBSE |
| Cupressus lusitânica Miller | Cedar of Goa | Cupressaceae | CVBLU |
| Eucalyptus globulus Labill. | Blue gum | Myrtaceae | EUCGL |
| Eucalyptus gunnii Hook. F. | Cider gum | Myrtaceae | EUCGU |
| Eucalyptus nitens Maiden | Shining gum | Myrtaceae | EUCNT |
| Pinus pinea L. | Stone pine | Pinaceae | PIUPI |
| Pinus pinaster Aiton | Maritime pine | Pinaceae | PIUPL |
| Pinus nigra Arnold | Black pine | Pinaceae | PIUNI |
| Pinus sylvestris L. | Scots pine | Pinaceae | PIUSI |
| Pinus halepensis Miller | Allepo pine | Pinaceae | PIUHA |
| Batch 2-Inoculated in Spring Herbaceous plants | |||
| Lupinus luteus L. | Yellow lupin | Fabaceae | LUPLU |
| Lupinus angustifolius L. | Blue lupin | Fabaceae | LUPAN |
| Lupinus albus L. | White lupin | Fabaceae | LUPAL |
| Hordeum murinum L. | Hare barley | Poaceae | HORDMU |
| Brachypodium distachyon (L.) P.Beauv | Purple false brome | Poaceae | BRCDI |
| Secale cereal L. | Rye | Poaceae | SECCE |
| Brassica nigra L. | Black mustard | Brassicaceae | BRSNI |
| Sinapis alba L. | White mustard | Brassicaceae | SINAL |
| Diplotaxis tenuifolia (L.) D.C. | Wallrocket | Brassicaceae | DIPTE |
| Eruca vesicaria (L.) Cav. | Rocket | Brassicaceae | ERUVU |
| Ornithopus sativus Brot. | Serradella | Fabaceae | OROSA |
| Vicia sativa L. | Common vetch | Fabaceae | VICSA |
| Batch 3-Inoculated in Autumn | |||
| Lolium multiflorum Lam. | Annual ryegrass | Poaceae | LOLMU |
| Lolium rigidum subsp. rigidum Gaudin. | Annual ryegrass | Poaceae | LOLRI |
| Lolium perenne L. | Perennial ryegrass | Poaceae | LOLPE |
| Dactylis glomerata L. | Cocksfoot | Poaceae | DACGL |
| Festuca arundinacea Schreb. | Tall fescue | Poaceae | FESAR |
| Trifolium incarnatum L. | Italian clover | Fabaceae | TRFIN |
| Trifolium repens L. | White clover | Fabaceae | TRFRE |
| Trifolium subterraneum L. | Subclover | Fabaceae | TRFSU |
| Trifolium michelianum Savi | Balansa | Fabaceae | TRFMI |
| Medicago sativa L. | Alfafa | Fabaceae | MEDSA |
| Texture (1) | Csand (%) | Fsand (%) | Lime (%) | Clay (%) | pH | N (gKg−1) | P2O5 (mgKg−1) | K2O (mgKg−1) | OM (%) |
|---|---|---|---|---|---|---|---|---|---|
| Loamy-Sand (A) | 79.8 | 8.2 | 3.4 | 8.6 | 6.4 | 1.12 | 113.6 | 294.0 | 5.3 |
| Loamy-Sand (B) | 68.2 | 16.6 | 5.7 | 5.5 | 5,9 | 0.3 | 25.0 | 41.0 | 27.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moreira, A.C.; Rodriguez-Romero, M.; Neno, J.; Rodrigues, A.; Calha, I. Response of Herbaceous and Woody Plant Species in Southern Portugal to Cope Oak Decline Associated to Phytophthora cinnamomi. Ecologies 2024, 5, 432-454. https://doi.org/10.3390/ecologies5030027
Moreira AC, Rodriguez-Romero M, Neno J, Rodrigues A, Calha I. Response of Herbaceous and Woody Plant Species in Southern Portugal to Cope Oak Decline Associated to Phytophthora cinnamomi. Ecologies. 2024; 5(3):432-454. https://doi.org/10.3390/ecologies5030027
Chicago/Turabian StyleMoreira, Ana Cristina, Manuela Rodriguez-Romero, Joana Neno, Abel Rodrigues, and Isabel Calha. 2024. "Response of Herbaceous and Woody Plant Species in Southern Portugal to Cope Oak Decline Associated to Phytophthora cinnamomi" Ecologies 5, no. 3: 432-454. https://doi.org/10.3390/ecologies5030027
APA StyleMoreira, A. C., Rodriguez-Romero, M., Neno, J., Rodrigues, A., & Calha, I. (2024). Response of Herbaceous and Woody Plant Species in Southern Portugal to Cope Oak Decline Associated to Phytophthora cinnamomi. Ecologies, 5(3), 432-454. https://doi.org/10.3390/ecologies5030027