

The Floristic Composition and Phytoecological Characterization of Plant Communities in the M’Goun Geopark, High Atlas, Morocco

 ,
,
Abstract
1. Introduction
2. Methodology
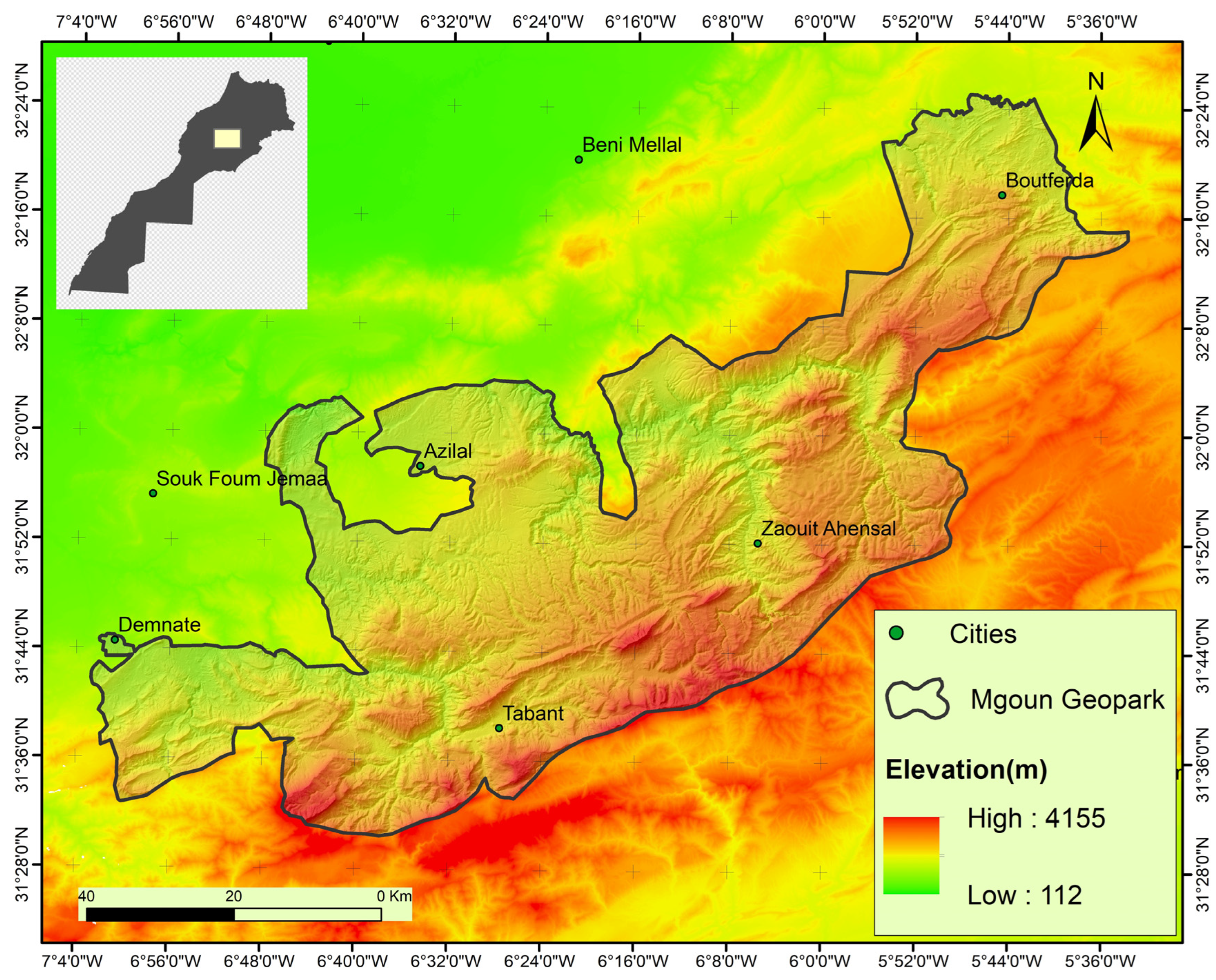
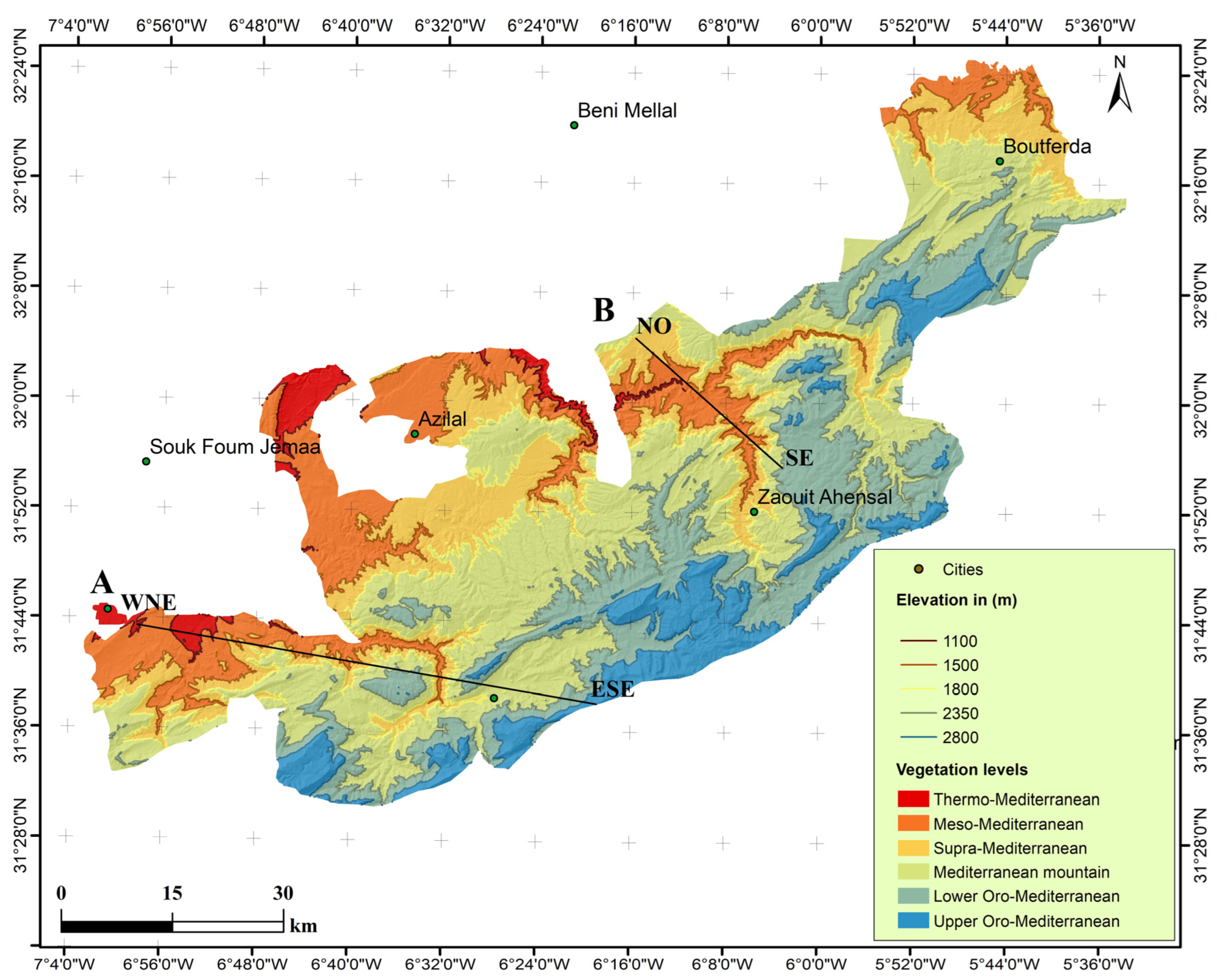
2.1. Study Area
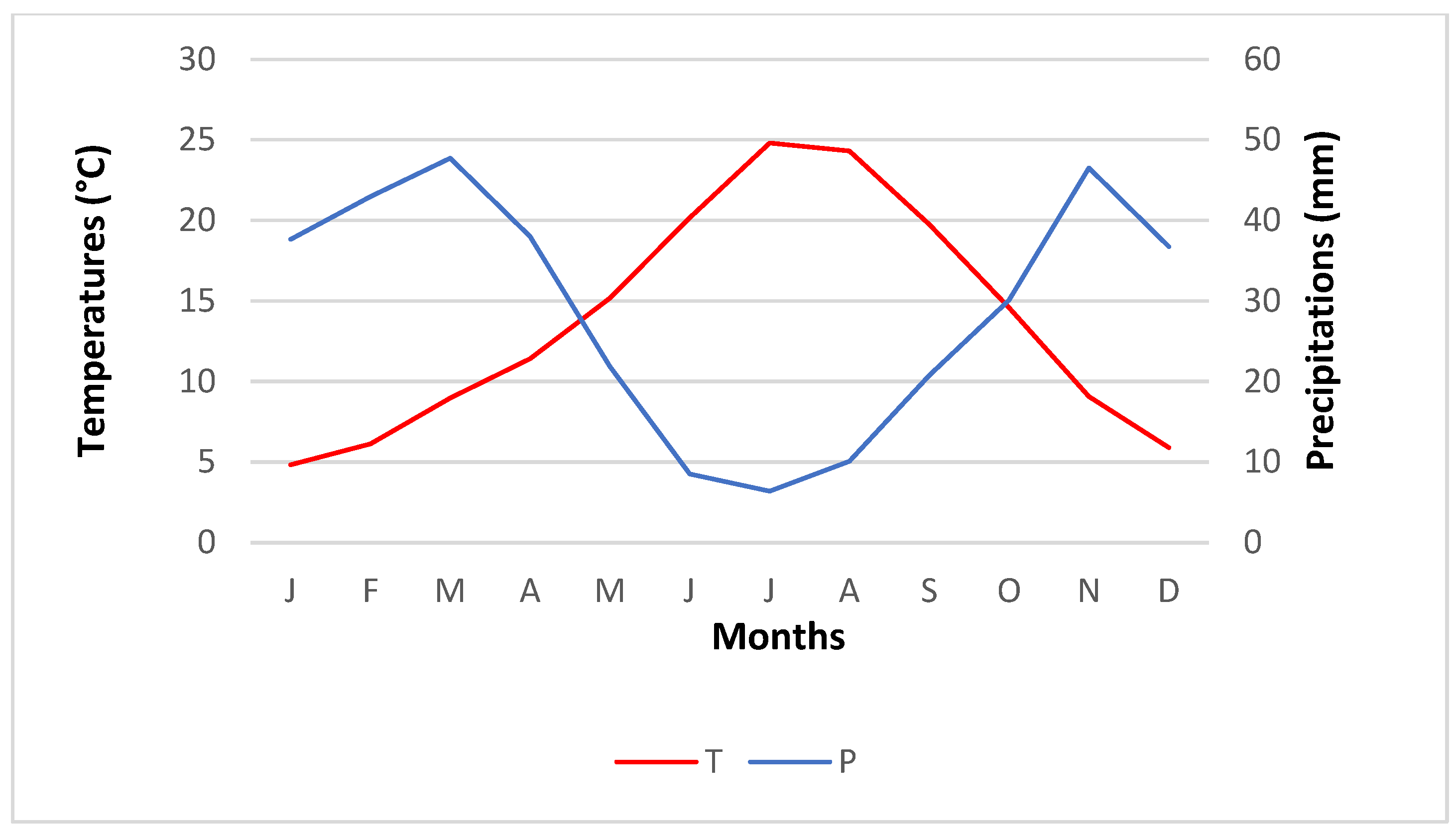
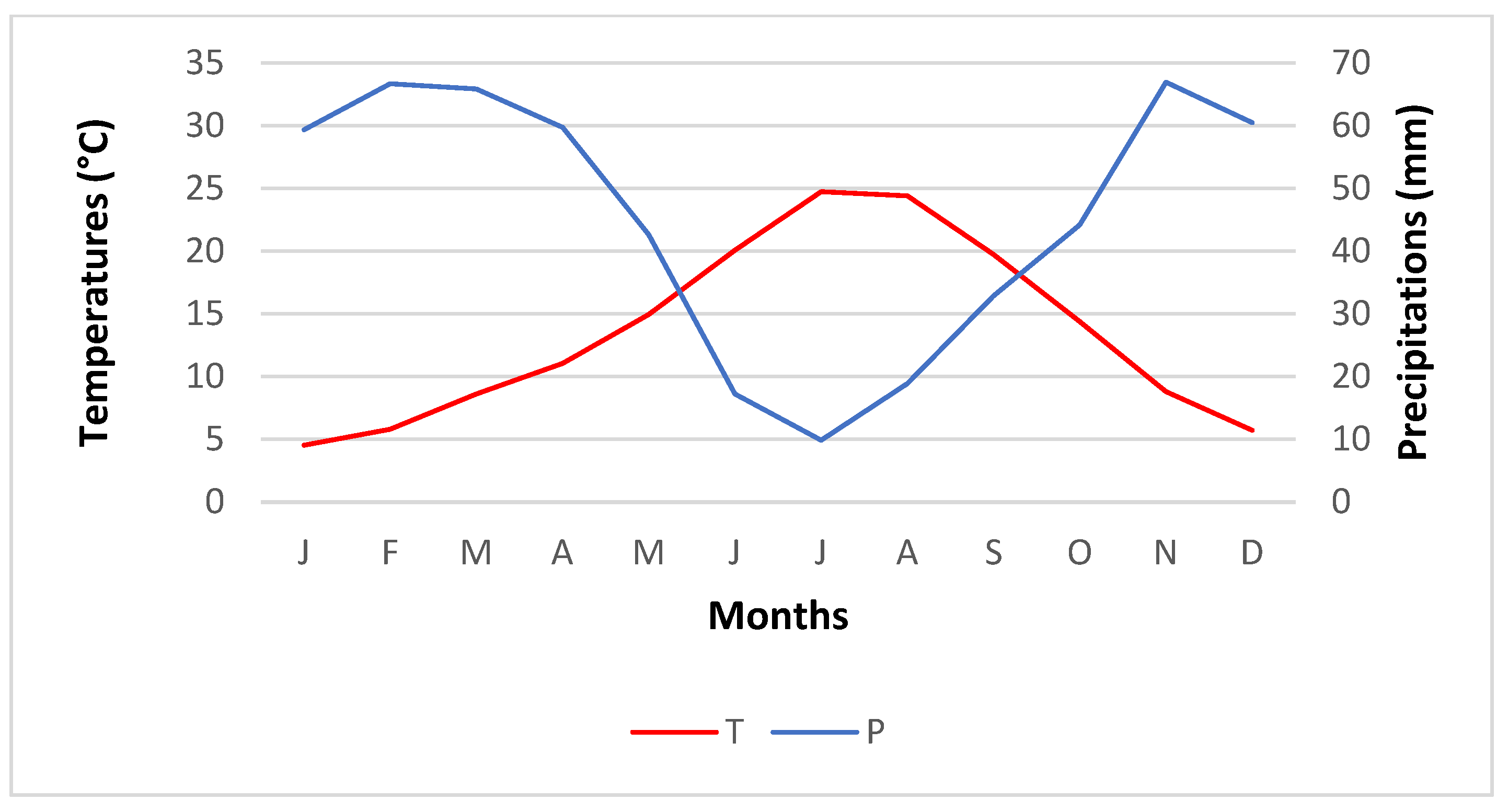
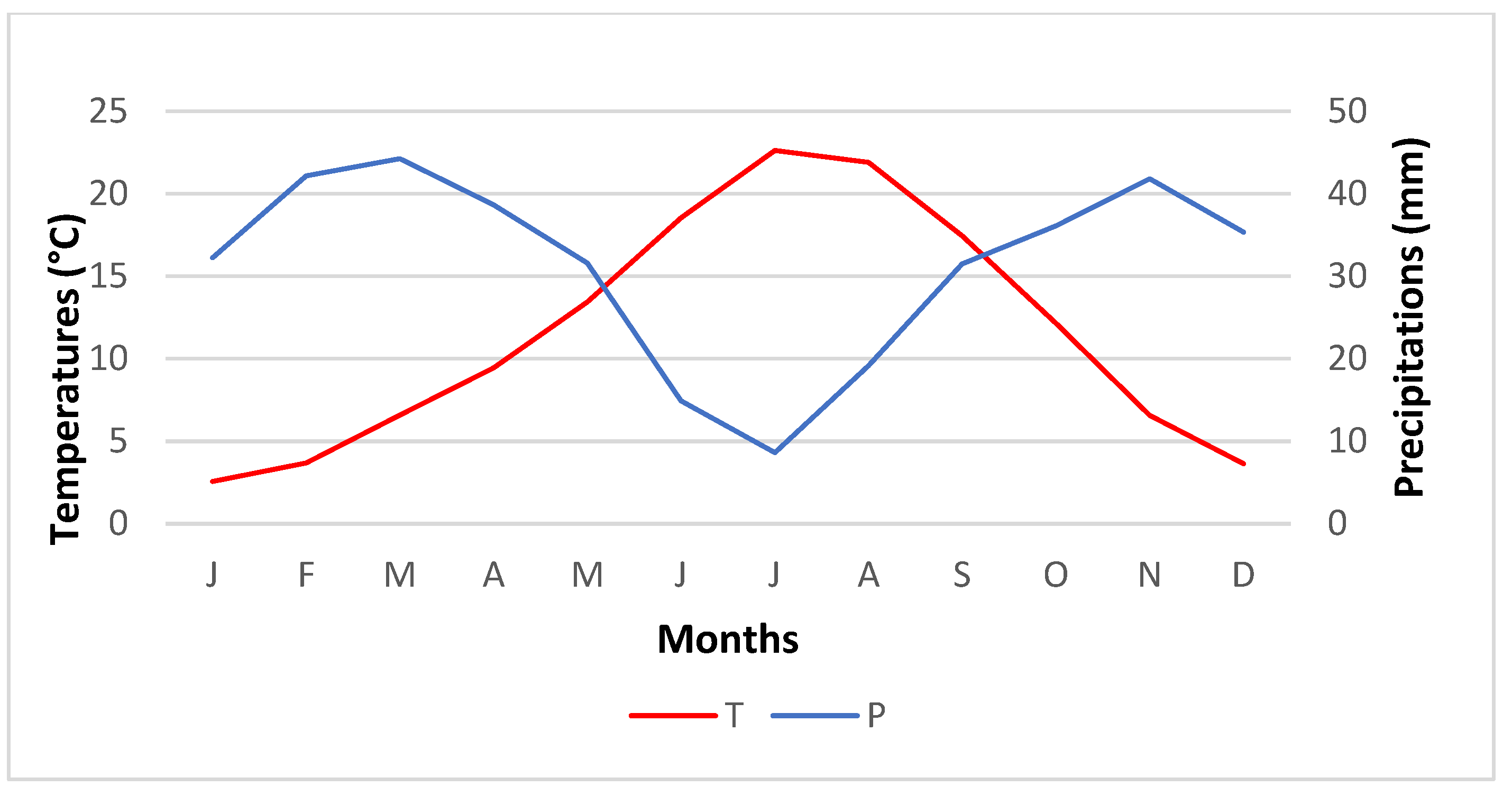
2.1.1. Temperature
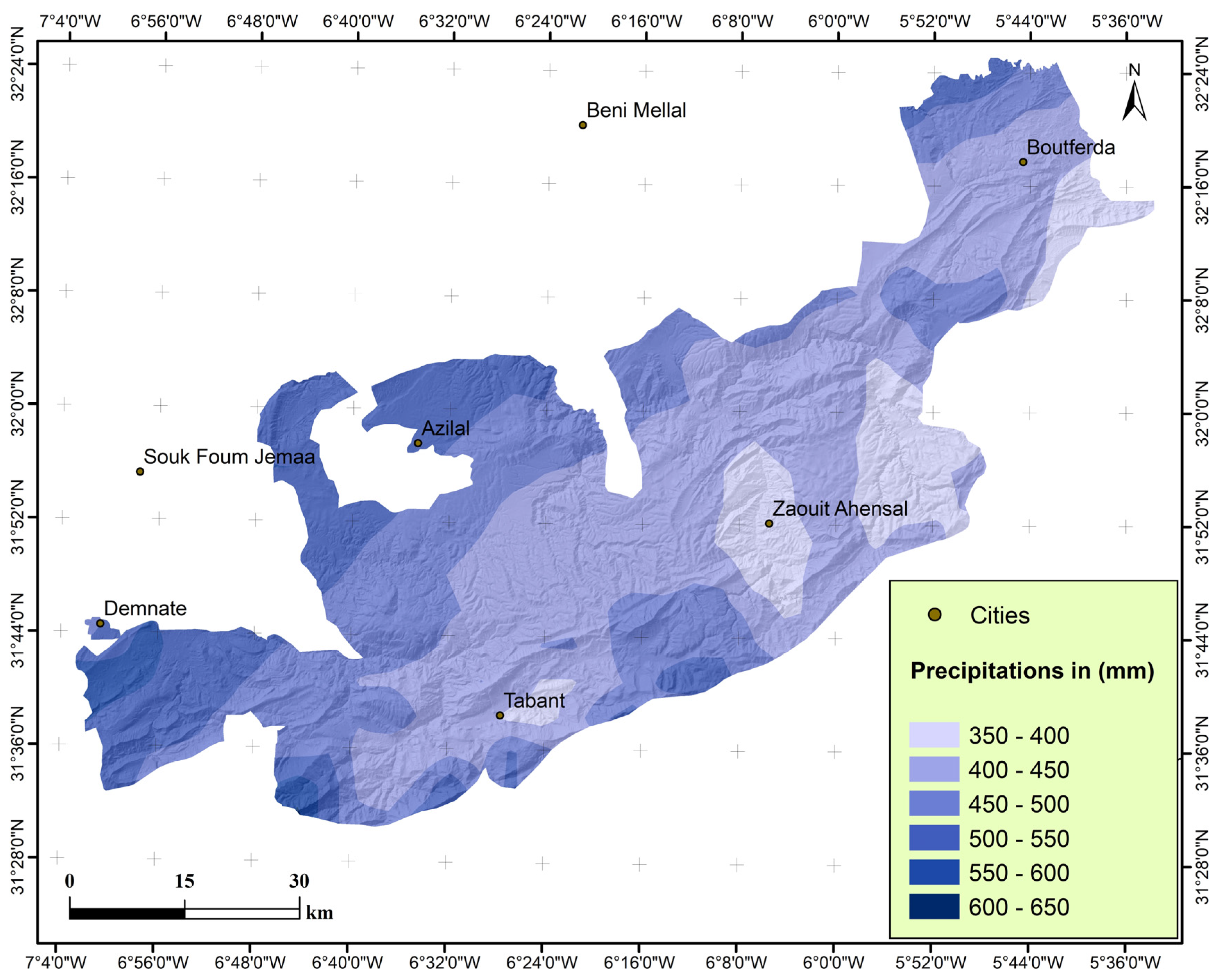
2.1.2. Rainfall Distribution
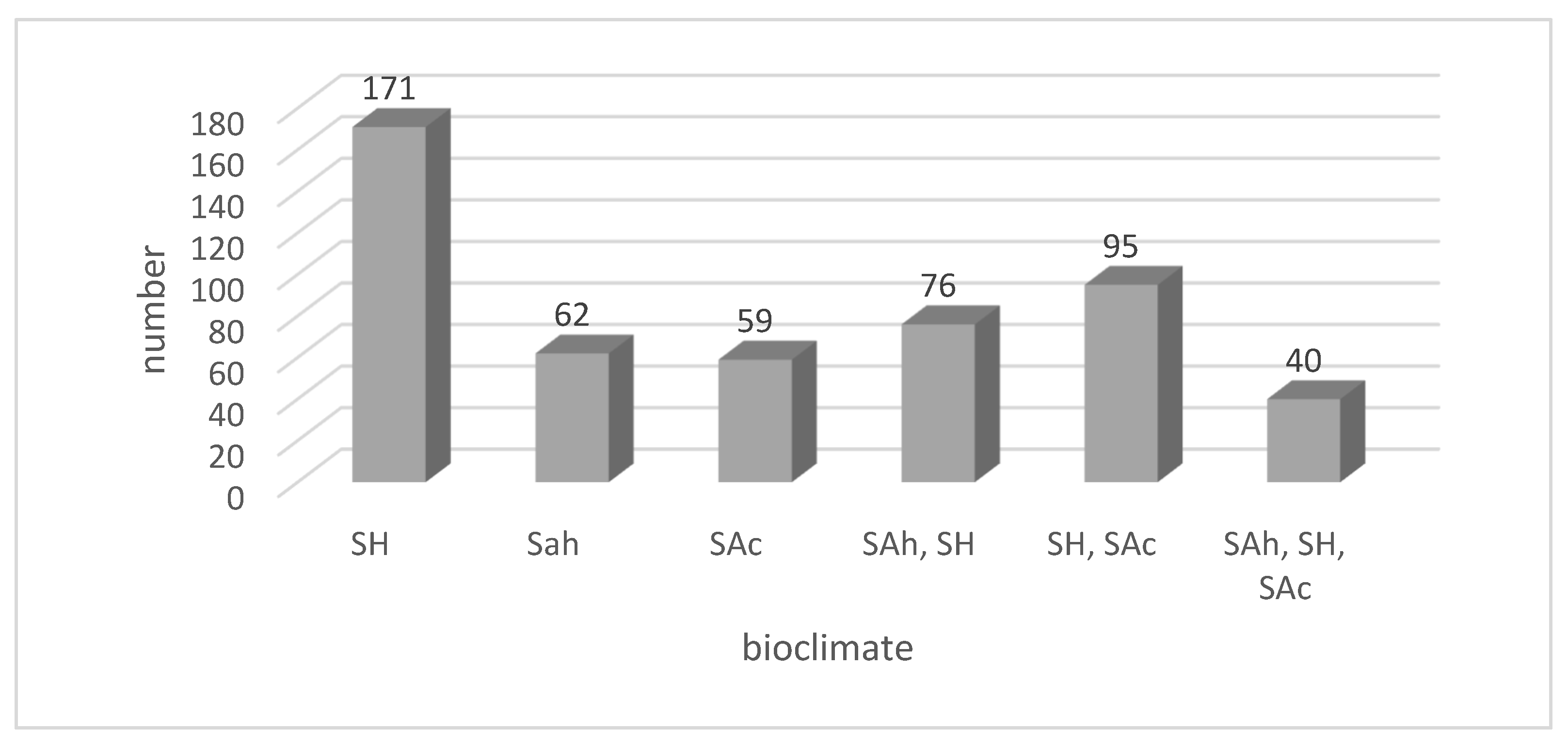
2.1.3. Bioclimatic Zones
2.2. Methods
2.2.1. Sampling, Species Identification, and Floristic Analysis
2.2.2. Phytoecology
3. Results
3.1. Plant Richness and Floristic Analysis
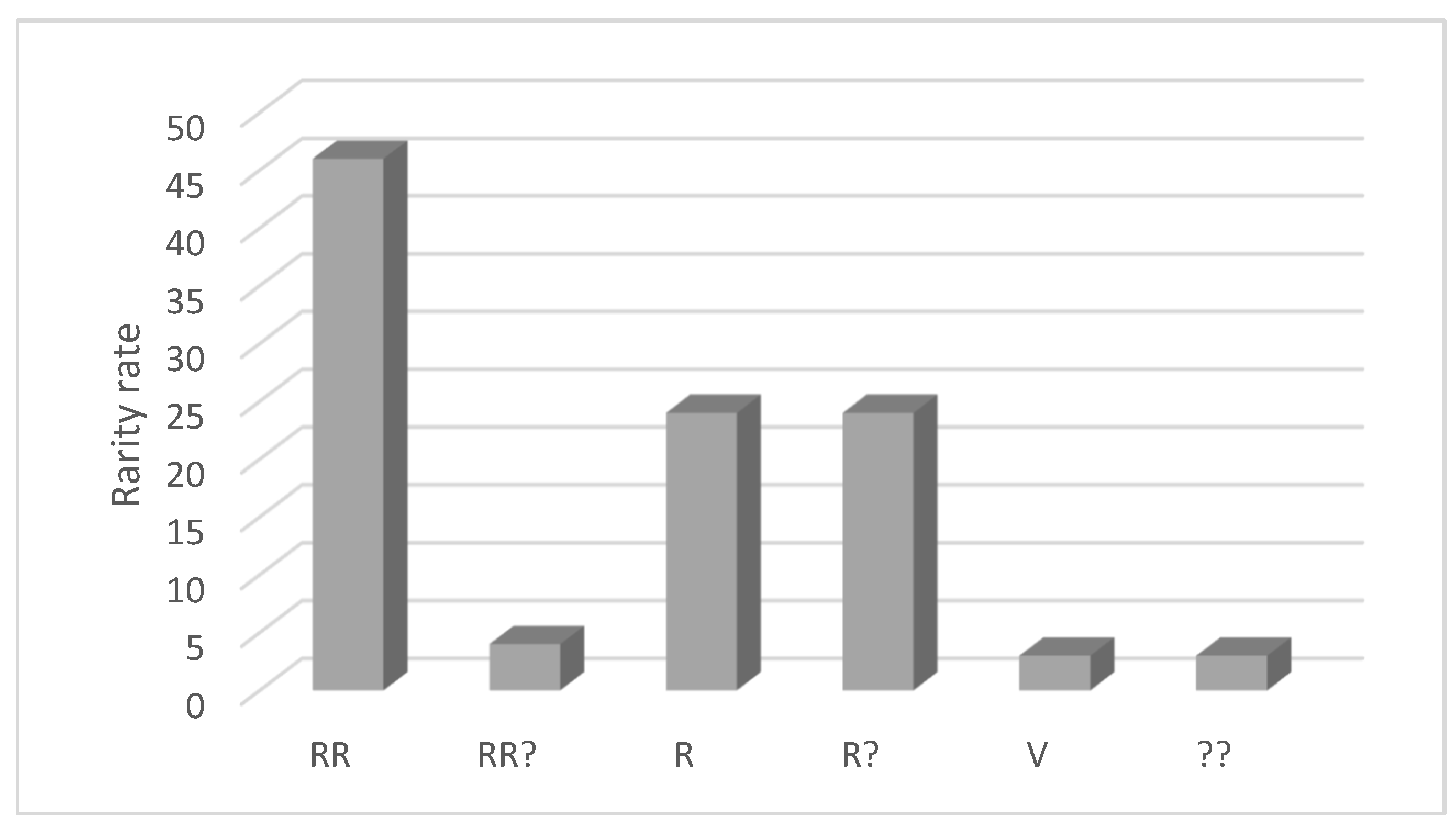
3.2. Endemism and Rarity
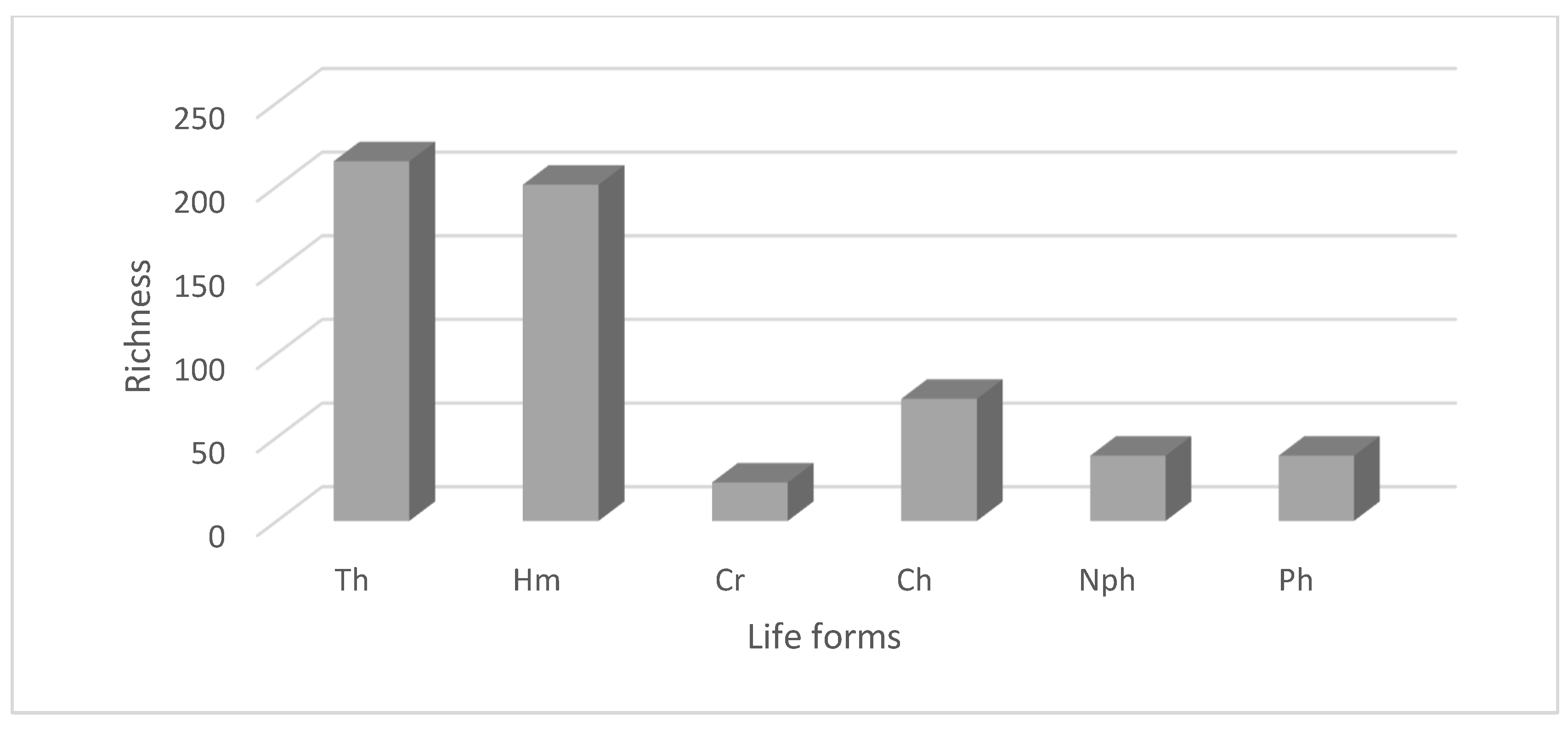
3.3. Life Forms
3.4. The Vegetation Cover
3.5. The Vegetation Levels and Associated Vegetation
4. Discussion
4.1. Floristic Richness
4.2. Phytoecology
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Séné, A.M. Perte et lutte pour la biodiversité: Perceptions et débats contradictoires. VertigO-la Rev. Électronique en Sci. L’environnement 2010, 10, 1–9. [Google Scholar]
- Mace, G.M.; Norris, K.; Fitter, A.H. Biodiversity and ecosystem services: A multilayered relationship. Trends Ecol. Evol. 2012, 27, 19–26. [Google Scholar]
- Bureau, D.; Bureau, J.-C.; Schubert, K. Biodiversité en danger: Quelle réponse économique? Notes du Cons. D’analyse Économique 2020, 59, 1–12. Available online: https://www.cairn.info/load_pdf.php?ID_ARTICLE=NCAE_059_0001 (accessed on 25 November 2024).
- Mauz, I.; Granjou, C. La construction de la biodiversité comme problème politique et scientifique, premiers résultats d’une enquête en cours. Sci. Eaux Territ. 2010, 3, 10–13. Available online: https://www.cairn.info/load_pdf.php?ID_ARTICLE=SET_003_0010 (accessed on 25 November 2024).
- Menioui, M. Biological diversity in Morocco. In Global Biodiversity; Apple Academic Press: New York, NY, USA, 2018; pp. 133–171+452. [Google Scholar]
- Pisani, D.; Pazienza, P.; Perrino, E.V.; Caporale, D.; De Lucia, C. The economic valuation of ecosystem services of biodiversity components in protected areas: A review for a framework of analysis for the Gargano National Park. Sustainability 2021, 13, 11726. [Google Scholar] [CrossRef]
- Gharnit, Y.; Outourakhte, A.; Moujane, A.; Ikhmerdi, H.; Hasib, A.; Boulli, A. Habitat diversity, ecology, and change assessment in the geoparc M’goun in High Atlas Mountains of Morocco. Geol. Ecol. Landsc. 2024; 1–22, in press. [Google Scholar] [CrossRef]
- Médail, F.; Diadema, K. Biodiversité végétale méditerranéenne. Ann. Georgr. 2006, 115, 618–640. [Google Scholar] [CrossRef]
- Mittermeier, R.A.; Myers, N.; Mittermeier, C.G.; Robles Gil, P. Hotspots: Earth’s Biologically Richest and Most Endangered Terrestrial Ecoregions; Cemex, S.A., Agrupación Sierra Madre, S.C., Eds.; Graphic Arts Center Publishing Company: Washington, DC, USA, 1999; 431p. [Google Scholar]
- Derneği, D. Ecosystem Profile: Mediterranean Basin Biodiversity Hotspot; BirdLife International: Cambridge, UK, 2010; p. 259. [Google Scholar]
- Underwood, E.C.; Viers, J.H.; Klausmeyer, K.R.; Cox, R.L.; Shaw, M.R. Threats and biodiversity in the mediterranean biome. In Diversity and Distributions; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2009; Volume 15, pp. 188–197. [Google Scholar]
- Cuttelod, A.; García, N.; Malak, D.A.; Temple, H.J.; Katariya, V. The Mediterranean: A biodiversity hotspot under threat. In Wildlife in a Changing World—An Analysis of the 2008 IUCN Red List of Threatened Species; IUCN: Gland, Switzerland, 2009; Volume 89, pp. 1–4. [Google Scholar]
- Ozenda, P. Sur les étages de végétation dans les montagnes du bassin méditerranéen. Doc. Cart. Ecol. 1975, 16, 1–32. [Google Scholar]
- Barbero, M.; Quézel, P. Les groupements forestiers de Grèce centro-méridionale. Ecol. Mediterr. 1976, 2, 3–86. [Google Scholar]
- Quézel, P.; Barbero, M. Definition and characterization of Mediterranean-type ecosystems. Ecol. Mediterr. 1982, 8, 15–29. [Google Scholar] [CrossRef]
- Tassin, C. Paysages Végétaux du Domaine Méditerranéen: Bassin Méditerranéen, Californie, Chili Central, Afrique du Sud, Australie Méridionale; IRD Éditions: Marseille, France, 2012; 424p. [Google Scholar]
- Benryane, M.A.; Belqadi, L.; Bounou, S.; Birouk, A. Analyse de la mise en œuvre du protocole de nagoya au maroc: Importance et limites de la gouvernance de la biodiversite. Rev. des Études Multidiscip. en Sci. Économiques Soc. 2022, 7, 167–196. [Google Scholar]
- Ilmen, R.; Benjelloun, H. Les écosystèmes forestiers marocains à l’épreuve des changements climatiques. Forêt Méditerranéenne 2013, XXXIV, 195–208. [Google Scholar]
- Gharnit, Y.; Outourakht, A.; Boulli, A.; Hassib, A. Biodiversity, Autecology and Status of Aromatic and Medicinal Plants in Geopark M’Goun (Morocco). Ann. di Bot. 2023, 13, 39–54. [Google Scholar] [CrossRef]
- Fennane, M.; Ibn Tattou, M. Statistiques et commentaires sur l’inventaire actuel de la flore vasculaire du Maroc. Bull. l’Institut Sci. Rabat, Sect. Sci. la Vie 2012, 34, 1–9. [Google Scholar]
- Mostakim, L.; Guennoun, F.Z.; Fetnassi, N.; Ghamizi, M. Analysis of floristic diversity of the forest ecosystems of the Zat valley-High Atlas of Morocco: Valorization and conservation perspectives. J. Adv. Biotechnol. Exp. Ther. 2021, 5, 126–135. [Google Scholar] [CrossRef]
- El Alami, A.; Fattah, A.; Bouzekraoui, H. Biodiversity, an essential component for the M’goun global geopark development (Morocco)-An overview. J. Anal. Sci. Appl. Biotechnol. 2021, 3, 103–106. [Google Scholar]
- Youssef, G.; Abdelaziz, M.; Aboubakre, O.; Abdelali, B.; Aziz, H. Impact of climate and demographic changes on the vegetation of the M’goun Geopark UNESCO of Morocco (1984–2021). Investig. Geográficas 2024, 81, 225–243. [Google Scholar]
- Gharnit, Y.; Moujane, A.; Outourakhte, A.; Hassan, I.; El Amraoui, K.; Hasib, A.; Boulli, A. Plant Richness, Species Assessment, and Ecology in the M’goun Geopark Rangelands, High Atlas Mountains, Morocco. Rangel. Ecol. Manag. 2025, 98, 357–376. Available online: https://www.sciencedirect.com/science/article/pii/S1550742424001490 (accessed on 4 January 2025). [CrossRef]
- Damiani, M.; Sinkko, T.; Caldeira, C.; Tosches, D.; Robuchon, M.; Sala, S. Critical review of methods and models for biodiversity impact assessment and their applicability in the LCA context. Environ. Impact Assess. Rev. 2023, 101, 107134. Available online: https://www.sciencedirect.com/science/article/pii/S0195925523001002 (accessed on 10 December 2024).
- Chiarucci, A.; Bacaro, G.; Scheiner, S.M. Old and new challenges in using species diversity for assessing biodiversity. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 2426–2437. [Google Scholar] [CrossRef]
- Lindenmayer, D.; Woinarski, J.; Legge, S.; Southwell, D.; Lavery, T.; Robinson, N.; Scheele, B.; Wintle, B. A checklist of attributes for effective monitoring of threatened species and threatened ecosystems. J. Environ. Manag. 2020, 262, 110312. Available online: https://www.sciencedirect.com/science/article/pii/S0301479720302474 (accessed on 20 November 2024).
- Green, M.J.B.; How, R.; Padmalal, U.; Dissanayake, S.R.B. The importance of monitoring biological diversity and its application in Sri Lanka. Trop. Ecol. 2009, 50, 41. [Google Scholar]
- Pauchard, A.; Meyerson, L.A.; Bacher, S.; Blackburn, T.M.; Brundu, G.; Cadotte, M.W.; Courchamp, F.; Essl, F.; Genovesi, P.; Haider, S. Biodiversity assessments: Origin matters. PLoS Biol. 2018, 16, e2006686. [Google Scholar]
- Vaughn, K.J.; Porensky, L.M.; Wilkerson, M.L.; Balachowski, J.; Peffer, E.; Riginos, C.; Young, T.P. Restoration ecology. Nat. Educ. Knowl. 2010, 3, 66. [Google Scholar]
- Van Der Maarel, E. The Braun-Blanquet approach in perspective. Vegetatio 1975, 30, 213–219. [Google Scholar] [CrossRef]
- Bussard, J.; Martin, S.; Monbaron, M.; Reynard, E.; El Khalki, Y. Geomorphological landscapes of the Central High Atlas (Morocco): Educative potential and resources for interpretation. Geomorphol. Process. Environ. 2022, 28, 173–185. [Google Scholar]
- Frizon de Lamotte, D.; Zizi, M.; Missenard, Y.; Hafid, M.; El Azzouzi, M.; Maury, R.C.; Charrière, A.; Taki, Z.; Benammi, M.; Michard, A. The Atlas System. In Continental Evolution: The Geology of Morocco: Structure, Stratigraphy, and Tectonics of the Africa-Atlantic-Mediterranean Triple Junction; Michard, A., Saddiqi, O., Chalouan, A., Lamotte, D.F.d., Eds.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 133–202. [Google Scholar]
- Bussard, J.; Martin, S.; Monbaron, M.; Reynard, E.; Khalki, Y. El Les paysages géomorphologiques du Haut Atlas central (Maroc): Potentiel éducatif et éléments pour la médiation scientifique. Géomorphologie Reli. Process. Environ. 2022, 28, 173–185. [Google Scholar]
- Taïbi, A.N.; Hannani, M.E.; Khalki, Y.E.; Ballouche, A. Les parcs agroforestiers d’Azilal (Maroc): Une construction paysagère pluri-séculaire et toujours vivante. Rev. Géographie Alp. 2019, 107, 1–17. [Google Scholar] [CrossRef]
- Moujane, A.; Boulli, A.; Gharnit, Y.; Outourakhte, A.; Ouigmane, A. Assessing of forest cover changes in Zaouit Ahansal (Central High Atlas, Morocco) using remote sensing and field data. J. Mater. Environ. Sci. 2024, 15, 1558. [Google Scholar]
- Ozenda, P. Les végétaux dans la biosphère. Rev. Géographie Alp. 1982, 6, 310–311. [Google Scholar]
- Fennane, M. Catalogue des plantes vascularies rares, menacées ou endémiques du Maroc. Bocconea 1998, 8, 5–243. [Google Scholar]
- APG Angiosperm Phylogeny Group III (APG III). An update of The Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III. Bot. J. Linn. Soc. 2009, 161, 105–121. [Google Scholar] [CrossRef]
- The Angiosperm Phylogeny Group; Chase, M.W.; Christenhusz, M.J.M.; Fay, M.F.; Byng, J.W.; Judd, W.S.; Soltis, D.E.; Mabberley, D.J.; Sennikov, A.N.; Soltis, P.S.; et al. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc. 2016, 181, 1–20. [Google Scholar] [CrossRef]
- Viscarra Rossel, R.A.; Behrens, T.; Ben-Dor, E.; Brown, D.J.; Demattê, J.A.M.; Shepherd, K.D.; Shi, Z.; Stenberg, B.; Stevens, A.; Adamchuk, V.; et al. A global spectral library to characterize the world’s soil. Earth-Sci. Rev. 2016, 155, 198–230. Available online: https://www.sciencedirect.com/science/article/pii/S0012825216300113 (accessed on 14 November 2024).
- ter Braak, C.J.F.; Verdonschot, P.F.M. Canonical correspondence analysis and related multivariate methods in aquatic ecology. Aquat. Sci. 1995, 57, 255–289. [Google Scholar] [CrossRef]
- Daget, P. Le bioclimat méditerranéen: Analyse des formes climatiques par le système d’Emberger. Vegetatio 1977, 34, 87–103. [Google Scholar]
- Calvet, C. Interprétation hydrique de la notion d’étage de végétation selon L. Emberger: Application au Maroc (Meteoric water and Emberger’s vegetation levels in Morocco). Bull. Assoc. Geogr. Fr. 1979, 56, 331–339. [Google Scholar]
- Achhal, A.; Akabli, A.; Barbero, M.; Benabid, A.; M’Hirit, O.; Peyre, C.; Quézel, P.; Rivas-Martinez, S. À propos de la valeur bioclimatique et dynamique de quelques essences forestières au Maroc. Ecol. Mediterr. 1979, 5, 211–249. [Google Scholar] [CrossRef]
- Benabid, A. Les écosystèmes forestiers, préforestiers et presteppiques du Maroc: Diversité, répartition biogéographique et problèmes posés par leur aménagement. Forêt Méditerranéenne 1985, 7, 53–64. [Google Scholar]
- Ibn Tattou, M.; Fennane, M. Aperçu historique et état actuel des connaissances sur la flore vasculaire du Maroc. Bull. Inst. Sci 1989, 13, 85–94. [Google Scholar]
- Körner, C. Mountain biodiversity, its causes and function: An overview. In Mountain Biodiversity; Routledge: London, UK, 2024; pp. 3–20. [Google Scholar]
- Spehn, E.M.; Rudmann-Maurer, K.; Körner, C. Mountain biodiversity. Plant Ecol. Divers. 2011, 4, 301–302. [Google Scholar] [CrossRef]
- Dundas, J. The Vegetation of Morocco and Western Algeria. J. Ecol. 1939, 27, 546. [Google Scholar] [CrossRef]
- El-Barougy, R.F.; Dakhil, M.A.; Halmy, M.W.A.; Cadotte, M.; Dias, S.; Farahat, E.A.; El-keblawy, A.; Bersier, L.-F. Potential extinction risk of Juniperus phoenicea under global climate change: Towards conservation planning. Glob. Ecol. Conserv. 2023, 46, e02541. Available online: https://www.sciencedirect.com/science/article/pii/S2351989423001762 (accessed on 20 October 2024). [CrossRef]
- Belghazi, B.; Ezzahiri, M.; Romane, F. Productivité de peuplements naturels de pin d’Alep (Pinus halepensis Miller) dans la forêt de Tamga (Haut Atlas, Maroc). Cah. Agric. 2000, 9, 39–46. [Google Scholar]
- Fougrach, H.; Badri, W.; Malki, M. Flore vasculaire rare et menacée du massif de Tazekka (région de Taza, Maroc). Bull. l’Institut Sci. Rabat Sect. Sci. la Vie 2007, 29, 1–10. [Google Scholar]
- Fennane, M.; De Montmollin, B. Réflexions sur les critères de l’UICN pour la Liste rouge: Cas de la flore marocaine. Bull. l’Institut Sci. Rabat 2015, 37, 1–11. [Google Scholar]
- Teixidor-Toneu, I.; M’Sou, S.; Salamat, H.; Baskad, H.A.; Illigh, F.A.; Atyah, T.; Mouhdach, H.; Rankou, H.; Babahmad, R.A.; Caruso, E.; et al. Which plants matter? A comparison of academic and community assessments of plant value and conservation status in the Moroccan High Atlas. Ambio 2022, 51, 799–810. [Google Scholar] [CrossRef]
- Medail, F.; Quezel, P. Hot-spots analysis for conservation of plant biodiversity in the Mediterranean Basin. Ann. Missouri Bot. Gard. 1997, 84, 112–127. [Google Scholar] [CrossRef]
- Maliha, N.S.; Chaloud, D.J.; Kepner, W.G.; Sarri, S. Regional Assessment of Landscape and Land Use Change in the Mediterranean Region BT—Environmental Change and Human Security: Recognizing and Acting on Hazard Impacts. In Environmental Change and Human Security: Recognizing and Acting on Hazard Impacts; Liotta, P.H., Mouat, D.A., Kepner, W.G., Lancaster, J.M., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 143–165. [Google Scholar]
- Defaut, B. Nouvelles considérations sur les phytoclimats du Maroc. Application au Maroc oriental. Matériaux Orthoptériques Et Entomocénotiques 2015, 20, 97–106. [Google Scholar]
- Benabid, A. Bref aperçu sur la zonation altitudinale de la végétation climacique du Maroc. Ecol. Mediterr. 1982, 8, 301–315. [Google Scholar] [CrossRef]
- Vargas, P. The Mediterranean Floristic Region: High Diversity of Plants and Vegetation Types; Goldstein, M.I., DellaSala DABTE of the WB, Eds.; Elsevier: Oxford, UK, 2020; pp. 602–616. Available online: https://www.sciencedirect.com/science/article/pii/B9780124095489120974 (accessed on 1 November 2024).
- Kim, A.-Y.; Lee, W.-S.; Son, Y. The Interaction between Climate Change and Biodiversity Can Be Assessed from a Material Cycle Perspective. Diversity 2024, 16, 506. [Google Scholar] [CrossRef]
- Kleespies, M.W.; Dierkes, P.W. Personal Assessment of Reasons for the Loss of Global Biodiversity—An Empirical Analysis. Sustainability 2020, 12, 4277. [Google Scholar] [CrossRef]
- Ebel, R.; Menalled, F.; Ahmed, S.; Gingrich, S.; Baldinelli, G.M.; Félix, G.F. How biodiversity loss affects society. In Handbook on the Human Impact of Agriculture; Edward Elgar Publishing: Cheltenham, UK, 2021; pp. 352–376. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Period | Tmin (Min–Max) in °C | Tmax (Min–Max) in °C | Precipitations (Min–Max) in mm | Q2 (Min–Max) |
|---|---|---|---|---|
| 1960–1969 | −9.97–2.18 | 20.9–30.83 | 369.67–658.68 | 42.63–77.62 |
| 1970–1979 | −9.37–3.12 | 23.5–34.42 | 350.32–605.81 | 37.92–66.91 |
| 1980–1989 | −10–2.34 | 23.5–34.63 | 295.56–506.28 | 31–54.39 |
| 1990–1999 | −9.74–2.89 | 23.87–35.17 | 307.88–541.58 | 32.34–58.25 |
| 2000–2009 | −8.86–2.39 | 24.42–35.65 | 309.58–533.49 | 32–57.67 |
| 2010–2019 | −9.39–3 | 24.60–36.12 | 329.25–571.19 | 33.96–60.76 |
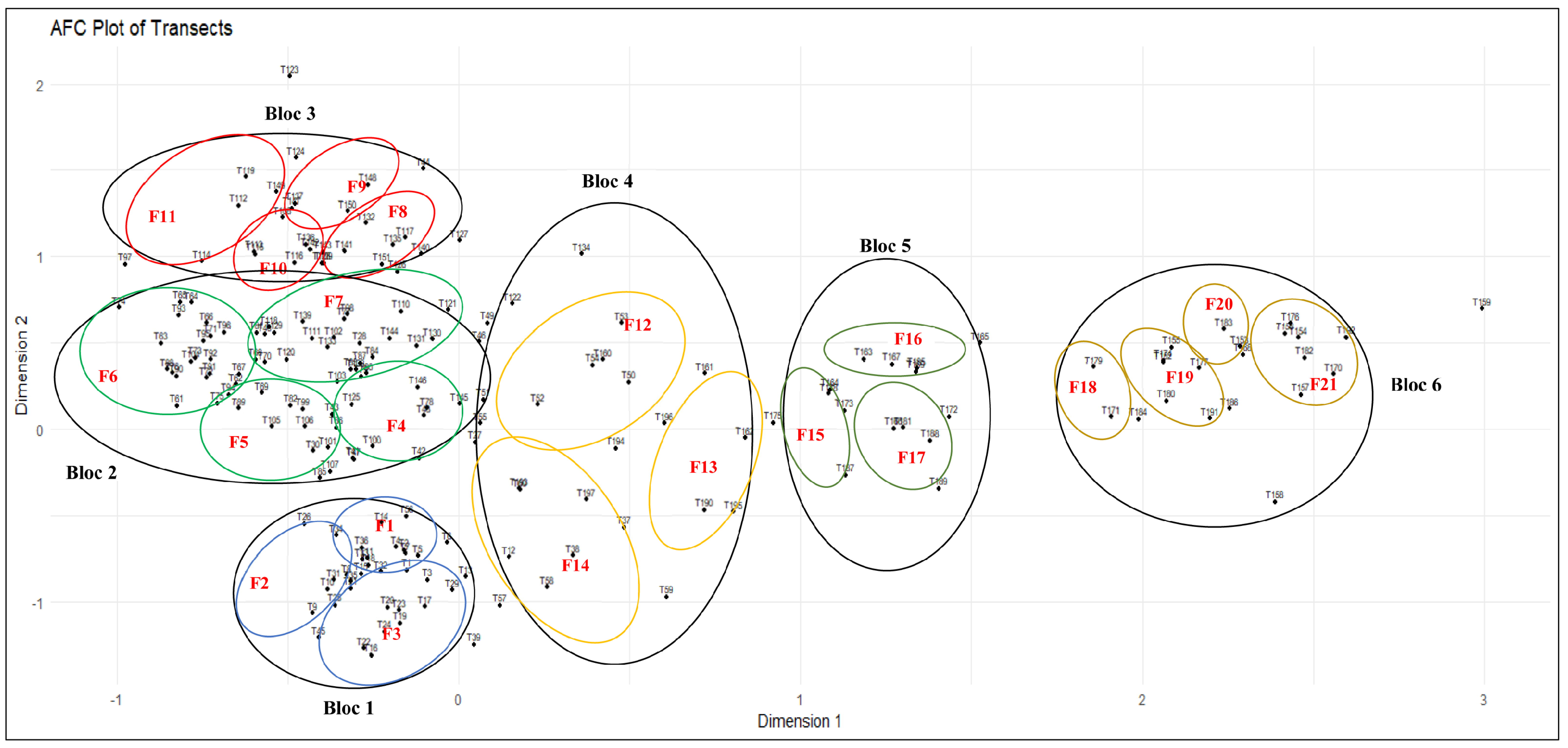
| Blocs | Transects | Abundant Species | |
|---|---|---|---|
| Bloc 1 | F1 | T6, T14, T4, T2, T5, T1, T56, T7 | Quercus ilex, Anchusa azurea, Astragalus incanus, Bromus madritensis, Campanula filicaulis, Catananche caerulea, Echinops spinosissimus, Lactuca tenerrima, Silene vulgaris, Asperula hirsuta, Picnomon acarna |
| F2 | T34, T36, T32, T15, T8, T35, T31, T10, T25, T9, T45, T18, T11 | Galium tricornicum, Papaver rhoeas, Bromus madritensis, Quercus ilex, Anchusa azurea, Medicago minima, Mantisalca salmantica, Plantago afra, Scolymus hispanicus, Beta macrocarpa, Medicago monspeliaca, Silene vulgaris, Sonchus asper | |
| F3 | T3, T13, T29, T17, T57, T39, T20, T23, T19, T24, T22, T16 | Quercus ilex, Hordeum murinum, Asphodelus macrocarpus, Galium tricornicum, Romulea bulbocodium, Anchusa azurea, Asperula hirsuta, Astragalus hamosus, Echinaria capitata, Echinops spinosissimus, Ononis cristata, Salvia verbenaca | |
| Bloc 2 | F4 | T146, T78, T145, T51, T55, T27, T42, T46 | Quercus ilex, Clematis cirrhosa, Echinops spinosissimus, Arthemisia herba alba, Bromus madritensis, Buxus balearica, Carlina hispanica, Chamaerops humilis, Convolvulus althaeoides, Hordeum murinum, Juniperus phoenicea |
| F5 | T100, T47, T41, T107, T85, T30, T101, T86, T125, T43, T106, T99, T82, T105 | Juniperus phoenicea, Anchusa azurea, Quercus ilex, Silene vulgaris, Alyssum alyssoides, Asperula arvensis, Atractylis cancellata, Avena fatua, Dactylis glomerata, Erodium cicutarium, Hedypnois rhagadioloides, Isatis tinctoria, Lactuca tenerrima, Medicago minima, Papaver rhoeas, Schismus barbatus, Torilis leptophylla | |
| F6 | T89, T69, T120, T75, T94, T62, T67, T68, T72, T118, T129, T81, T61, T92, T96, T66, T71, T95, T73, T104, T63, T81, T49, T88, T91, T93, T90, T101, T64, T93, T63 | Juniperus phoenicea, Stipellula capensis, Chamaerops humilis, Bromus madritensis, Pistacia lentiscus, Atractylis cancellata, A. sterilis, Torilis nodosa, Ceratonia siliqua, Cladanthus arabicus, Sonchus oleraceus, Tetraclinis articulata, Ziziphus lotus, Euphorbia resinifera… | |
| F7 | T139, T111, T133, T102 T77, T98, T28 T84, T87, T80, T103, T110, T144, T130, T131, T121, T49, T46 | Juniperus phoenicea, Picnomon acarna, Pinus halepensis, Buxus balearica, Quercus ilex, Thymus algeriensis, Lactuca tenerrima, Thymus saturejoides, Ajuga iva, Chondrella jencea, Crambe filiformis, Dactylis glomerata, Deverra scoparia, Genista scorpius, Globularia nainii, Lactuca viminea, Phagnalon saxatile, Phillyrea angustifolia, Salvia verbenaca, Teucrium polium, Thymus zygis | |
| Bloc 3 | F8 | T127, T140, T117, T135, T151, T126 | Pinus halepensis, Globularia nainii, Buxus balearica, C. corymbosa, Centaurea melitensis, Cistus creticus, Euphorbia niceensis, Fraxinus angustifolia, Fumana ericoides, Narsturia officinalis, Polygala balansae, Sedum sediforme, Stipellula capensis |
| F9 | T44, T148, T132, T150 | Thymus zygis, Anarrhinum fruticosum, Globularia nainii, Arbutus unedo, Astragalus incanus, Euphorbia taurinensis, Ononis pusilla, Fraxinus angustifolia, Schismus barbatus, Ajuga iva | |
| F10 | T141, T143, T116, T136, T142, 128, | Juniperus phoenicea, Xanthium spinosus, Ajuga iva, Andryala integrifolia, Capparis spinosa, Crambe filiformis, Dittrichia viscosa, Erodium cicutarium, Globularia alypum, Melilotus sulcatus, Mentha peligium, Mentha suaveolens, Nerium oleander, Pinus halepensis, Pistacia lentiscus, Polypogon minspeliensis, Schismus barbatus | |
| F11 | T124, T119, T112, T149, T137, T108, T114, T113, T115, T147 | Globularia nainii, Hirschfeldia incana, Lythrum junceum, Pinus halepensis, Pistacia lentiscus, Polygala balansae, Teucrium polium, Bombycilaena erecta, Centaurea calcitrapa, Chondrella jencea, Cistus creticus, Juniperus phoeniceae, Nerium oleander, Populus nigra, Schismus barbatus, Veronica polita, Anarrhinum fruticosum, Centaurea melitensis, Cistus albidus, Dianthus nudiflorus, Dittrichia viscosa | |
| Bloc 4 | F12 | T52, T54, T160, T53, T50, T194, T196, T161 | Euphorbia niceensis, Fraxinus angustifolia, Polycarpon tetraphyllum, Bellis annua, Eryngium campestre, Lactuca tenerrima, poclama brevifolia, Quercus, Astragalus incanus, Bromus madritensis, Crambe filiformis, Euphorbia terracina, Globularia nainii, Marrubium ayardii |
| F13 | T175, T162, T195, T190 | Teucrium chamaedrys, Alyssum spinosum, Polycarpon tetraphyllum, Arenaria pungens, asperula hirsuta, Bupleurum spinosum, Cerastium arvense, Helianthemum cenereum, Jurinea humilis, Minuartia funkii, Thymelaea virgata, Thymus zygis | |
| F14 | T193, T58, T38, T197, T37, T59 | Quercus ilex, Astragalus granatensis, Campanula filicaulis, Ononis spinosa, Polycarpon tetraphyllum, Scutellaria orientalis, Teucrium chamaedrys, Aegilops geniculate, Asperula hirsuta, Bromus madritensis, Cirsium acaule, Convolvulus lineatus, Coronilla minima, Crataegus lacinata | |
| Bloc 5 | F15 | T187, T173, T164, T178 | Cytisus balansae, Euphorbia niceensis, Alyssum spinosum, Hirschfeldia incana, Anthyllis vulneraria Arthemisia herba alba Buxus balearica, Cirsium dyris, Coronilla minima, Delphinium gracile, Helianthemum apenninum, Juniperus thurifera, Jurinea humilis, Ormenis scariosa, Quercus ilex, Scorzonera caespitosa |
| F16 | T163, T167, T185, T165 | Arthemisia herba alba, Ormenis scariosa, Euphorbia niceensis, Sanguisorba minor, Alyssum spinosum, Asperula cynanchica, Avena barbata, Bupleurum spinosum, Coronilla minima, Erinacea Anthyllis, Helianthemum cinereum, Hieracium pseudopilosella, Vella maierii | |
| F17 | T172, T188, T181, T189 | Euphorbia nicaeensis, Ribes uva-crispa, Coronilla minima, Erinacea Anthyllis, Alyssum serpyllifolium, Buxus balearica; Cytisus balansae, Juniperus thurifera, Raffenaldia platycarpa, Scorzonera angustifolia, Scorzonera caespitosa, Thymelaea virgata | |
| Bloc 6 | F18 | T179, T171, T184 | Alyssum spinosum, Scorzonera caespitosa, Thymus pallidus, Arenaria pungens, Carduncellus atractyloides, Centaurea takredensis, Convolvulus sabatius, Delphinium gracile, Erinacea Anthyllis, Euphorbia niceensis, Euphorbia sagitalis, Juniperus thurifera |
| F19 | T186, T191, T180, T177, T155, T152, T174 | Alyssum spinosum, Bupleurum spinosum, Carduncellus atractyloides, Centaurea takredensis, Cytisus balansae, Erinacea Anthyllis, Euphorbia nicaeensis, Vella maierii, Ribes uva-crispa, Arenaria serpyllifolia, Arenaria pungens, Berberis vulgaris, Bupleurum atlanticum, Cirsium dyris, Delphinium gracile, Juniperus thurifera, Minuartia funkii, Ormenis scariosa, Prunus prostrata, Rahmnus liscoides, Thymus pallidus | |
| F20 | T183, T153, T168 | Euphorbia nicaeensis, Alyssum spinosum Erinacea Anthyllis, Juniperus thurifera, Scorzonera caespitosa, Vella mairei, Arthemisia herba alba, Bupleurum, atlanticum, Bupleurum spinosum, Convolvulus lineatus, Ormenis scariosa | |
| F21 | T176, T154, T182, T170, T157, T192, T156 | Arenaria pungens, Valla mairei, Alyssum spinosum, Erinacea Anthyllis, Bupleurum spinosum, Clinopodium alpinum, Cytisus balansae, Euphorbia niceensis, Euphorbia megatlantica, Scorzonera caespitosa, Berberis vulgaris, Coronilla minima, Ormenis scariosa, Papaver atlanticum, Ruta montana, Stipa nitans | |
| Habitat | Elevation (m) | Tmin (°C) | Tmax (°C) | Precipitation (mm) | Bioclimate | Climatic Variation | Vegetation Zone |
|---|---|---|---|---|---|---|---|
| Euphorbia resinifera | Up to 1800 | 8.05–9.06 | 21.69–22.13 | 305.13–552.85 | Semi-arid and subhumid | Moderate and cold | Meso-Mediterranean |
| Juniperus phoenicea | Up to 2000 | 6.92–7.88 | 19.95–20.56 | 313.2–571.99 | Semi-arid and subhumid | Very cold and cold | Meso- and Thermo-Mediterranean |
| Chamaerops humilis | Up to 2000 | 6.62–7.69 | 20.98–21.43 | 302.59–545.73 | Subhumid and semi-arid | Cold | Supra-Mediterranean |
| Pinus halepensis | Up to 2200 | 6.29–7.36 | 19.13–19.72 | 278.23–501.08 | Subhumid and semi-arid | Cool and cold | Supra- and Meso-Mediterranean |
| Quercus ilex | Up to 2800 | 5.61–6.26 | 18.69–19.33 | 299.01–548.25 | Semi-arid and subhumid | All variants | Supra-, Meso-, Thermo-, and Montane Mediterranean |
| Juniperus thurifera | Up to 2700 | 3.41–4.42 | 16.26–16.98 | 288.48–499.47 | Subhumid and semi-arid | Cold and very cold | Montane Mediterranean |
| Buxus sempervirens | Up to 3000 | 1.99–2.93 | 14.9–15.51 | 281.61–478.13 | Subhumid and semi-arid | Cold and very cold | Montane Mediterranean |
| Xerophytes | Up to 3700 | 2.24–3.26 | 15.14–15.79 | 291.93–532.19 | Subhumid and semi-arid | Cold and very cold | Montane and Oro-Mediterranean |
| Bloc | SR | Formation | Vegetation Level | SR |
|---|---|---|---|---|
| B1 | 233 | F1 | Meso-Mediterranean | 91 |
| F2 | Meso-Mediterranean and Supra-Mediterranean | 176 | ||
| F3 | Meso-Mediterranean and Supra-Mediterranean | 160 | ||
| B2 | 277 | F4 | Supra-Mediterranean | 91 |
| F5 | Meso-Mediterranean | 114 | ||
| F6 | Thermo- Mediterranean | 172 | ||
| F7 | Meso-Mediterranean | 134 | ||
| B3 | 168 | F8 | Meso-Mediterranean | 64 |
| F9 | Meso-Mediterranean | 31 | ||
| F10 | Meso-Mediterranean | 91 | ||
| F11 | Meso-Mediterranean | 111 | ||
| B4 | 119 | F12 | Supra-Mediterranean and Montane Mediterranean | 53 |
| F13 | Montane Mediterranean | 41 | ||
| F14 | Supra-Mediterranean and Montane Mediterranean | 69 | ||
| B5 | 86 | F15 | Montane Mediterranean | 42 |
| F16 | Montane Mediterranean | 42 | ||
| F17 | Montane Mediterranean | 57 | ||
| B6 | 91 | F18 | Oro-Mediterranean | 27 |
| F19 | Oro-Mediterranean | 57 | ||
| F20 | Oro-Mediterranean | 18 | ||
| F21 | Oro-Mediterranean | 53 |
| Symbol | Dominant Species | Companion Species | Elevation | Bioclimatic Stage | Vegetation Zone | Tmin and Tmax | Precipitation | Substrate Type |
|---|---|---|---|---|---|---|---|---|
| F1 | Quercus ilex, Anchusa azurea, Astragalus incanus | Bromus madritensis, Campanula filicaulis, Catananche caerulea | 1100–1500 | Semi-arid fresh | Meso-Mediterranean | 0 °C, 32 °C | 400–600 mm | Limestones |
| F2 | Galium tricornicum, Quercus ilex, Anchusa azurea | Bromus madritensis, Medicago minima, Silene vulgaris | 1100–1800 | Semi-arid cold and sub-humid cold | Meso-Mediterranean Supra-Mediterranean | −3 °C, 28 °C | 550–650 mm | Limestones |
| F3 | Quercus ilex, Hordeum murinum, Asphodelus macrocarpus | Galium tricornicum, Romulea bulbocodium, Anchusa azurea | 1100–1800 | Semi-arid cold and sub-humid cold | Meso-Mediterranean Supra-Mediterranean | −3 °C, 30 °C | 550–650 mm | Limestones |
| F4 | Quercus ilex, Clematis cirrhosa, Echinops spinosissimus | Artemisia herba-alba, Juniperus phoenicea | 1500–1800 | Semi-arid, cold | Supra-Mediterranean | −3 °C, 30 °C | 550–600 mm | Limestones |
| F5 | Juniperus phoenicea, Anchusa azurea, Quercus ilex | Silene vulgaris, Avena fatua, Dactylis glomerata | 1100–1500 | Semi-arid fresh and Sub-humid fresh | Meso-Mediterranean | 0 °C, 32 °C | 600–650 mm | Limestones |
| F6 | Juniperus phoenicea, Stipellula capensis, Chamaerops humilis | Atractylis cancellata, Euphorbia resinifera | 544–1100 | Semi-arid fresh and Sub-humid fresh | Thermo- Mediterranean | 5 °C, 34 °C | 600–650 mm | Clays and sandstone |
| F7 | Juniperus phoenicea, Pinus halepensis, Buxus balearica | Thymus algeriensis, Lactuca tenerrima, Genista scorpius | 1400–1500 | Semi-arid fresh and Sub-humid fresh | Meso-Mediterranean and | 0 °C, 30 °C | 550–650 mm | Clays and limestone conglomerates |
| F8 | Pinus halepensis, Globularia nainii, Buxus balearica | Centaurea melitensis, Cistus creticus, Euphorbia niceensis | 1400–1500 | Semi-arid fresh and Sub-humid fresh | Meso-Mediterranean and | 0 °C, 30 °C | 550–650 mm | Clays and limestone |
| F9 | Thymus zygis, Anarrhinum fruticosum, Globularia nainii | Arbutus unedo, Astragalus incanus, Ononis pusilla | 1400–1500 | Semi-arid fresh and Sub-humid fresh | Meso-Mediterranean | 0 °C, 32 °C | 600–650 mm | Limestones and rocky limestones |
| F10 | Juniperus phoenicea, Xanthium spinosus, Ajuga iva | Capparis spinosa, Crambe filiformis, Nerium oleander | 1500–1800 | Semi-arid fresh and Sub-humid fresh | Meso-Mediterranean | 0 °C, 32 °C | 600–650 mm | Clays and sandstones |
| F11 | Globularia nainii, Hirschfeldia incana, Lythrum junceum | Pinus halepensis, Polygala balansae, Teucrium polium | 1500–1800 | Semi-arid fresh and Sub-humid fresh | Meso-Mediterranean | 0 °C, 32 °C | 600–650 mm | Clays |
| F12 | Euphorbia niceensis, Fraxinus angustifolia, Polycarpon tetraphyllum | Bellis annua, Lactuca tenerrima, Quercus ilex | 1700–2350 | Semi-arid cold and very cold | Supra-Mediterranean and Montane Mediterranean | −6 °C, 28 °C | 500–600 mm | Limestones |
| F13 | Teucrium chamaedrys, Alyssum spinosum, Polycarpon tetraphyllum | Arenaria pungens, Bupleurum spinosus, Cerastium arvense | 1800–2350 | Semi-arid cold and very cold | Montane Mediterranean | −6 °C, 28 °C | 400–450 mm | Limestones, marls, and dolomites |
| F14 | Quercus ilex, Astragalus granatensis, Campanula filicaulis | Ononis spinosa, Polycarpon tetraphyllum, Teucrium chamaedrys | 1700–2350 | Semi-arid cold, very cold, and Sub-humid cold | Supra-Mediterranean and Montane Mediterranean | −6 °C, 28 °C | 600–650 mm | Limestones |
| F15 | Cytisus balansae, Euphorbia nicaeensis, Alyssum spinosum | Hirschfeldia incana, Cirsium dyris, Coronilla minima | 2200–2350 | Semi-arid cold and very cold | Montane Mediterranean | −6 °C, 28 °C | 400–600 mm | Limestones |
| F16 | Artemisia herba-alba, Ormenis scariosa, Euphorbia niceensis | Sanguisorba minor, Alyssum spinosum, Bupleurum spinosus | 2200–2350 | Semi-arid cold and very cold | Montane Mediterranean | −6 °C, 28 °C | 400–600 mm | Limestones |
| F17 | Euphorbia niceensis, Ribes uva-crispa, Coronilla minima | Erinacea anthyllis, Alyssum serpyllifolium, Juniperus thurifera | 2000–2350 | Semi-arid cold and very cold | Montane Mediterranean | −6 °C, 28 °C | 400–600 mm | Limestones |
| F18 | Alyssum spinosum, Scorzonera caespitosa, Thymus pallidus | Arenaria pungens, Carduncellus atractyloides, Euphorbia niceensis | 2200–2350 | Semi-arid cold and very cold | Montane Mediterranean | −6 °C, 28 °C | 400–600 mm | Limestones |
| F19 | Alyssum spinosum, Bupleurum spinosum, Carduncellus atractyloides | Cytisus balansae, Erinacea anthyllis, Euphorbia niceensis | 2350–2800 | Sub-humid is extremely cold and Semi-arid extremely cold | Oro-Mediterranean | −9 °C, 24 °C | 550–650 mm | Limestones |
| F20 | Euphorbia niceensis, Alyssum spinosum, Erinacea anthyllis | Juniperus thurifera, Vella mairii, Ormenis scariosa | 2350–2800 | Sub-humid is extremely cold and Semi-arid is extremely cold | Oro-Mediterranean | −9 °C, 24 °C | 550–650 mm | Limestones |
| F21 | Arenaria pungens, Vella mairii, Alyssum spinosum | Erinacea anthyllis, Bupleurum spinosum, Cytisus balansae | 2350–2800 | Sub-humid is extremely cold and Semi-arid is extremely cold | Oro-Mediterranean | −9 °C, 24 °C | 550–650 mm | Limestones |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Outourakhte, A.; Gharnit, Y.; Moujane, A.; El Haddany, K.; Hasib, A.; Boulli, A. The Floristic Composition and Phytoecological Characterization of Plant Communities in the M’Goun Geopark, High Atlas, Morocco. Ecologies 2025, 6, 29. https://doi.org/10.3390/ecologies6020029
Outourakhte A, Gharnit Y, Moujane A, El Haddany K, Hasib A, Boulli A. The Floristic Composition and Phytoecological Characterization of Plant Communities in the M’Goun Geopark, High Atlas, Morocco. Ecologies. 2025; 6(2):29. https://doi.org/10.3390/ecologies6020029
Chicago/Turabian StyleOutourakhte, Aboubakre, Youssef Gharnit, Abdelaziz Moujane, Khalid El Haddany, Aziz Hasib, and Abdelali Boulli. 2025. "The Floristic Composition and Phytoecological Characterization of Plant Communities in the M’Goun Geopark, High Atlas, Morocco" Ecologies 6, no. 2: 29. https://doi.org/10.3390/ecologies6020029
APA StyleOutourakhte, A., Gharnit, Y., Moujane, A., El Haddany, K., Hasib, A., & Boulli, A. (2025). The Floristic Composition and Phytoecological Characterization of Plant Communities in the M’Goun Geopark, High Atlas, Morocco. Ecologies, 6(2), 29. https://doi.org/10.3390/ecologies6020029