
Tree Size Structure of Tectona grandis (Linn f.) Stand in Hilltop and Valley-Bottom of Omo Forest Reserve †


Abstract
:1. Introduction
2. Materials and Methods
2.1. The Study Area
2.2. Demarcation of Sample Plots and Method of Data Collection
2.3. Data Analysis
3. Results
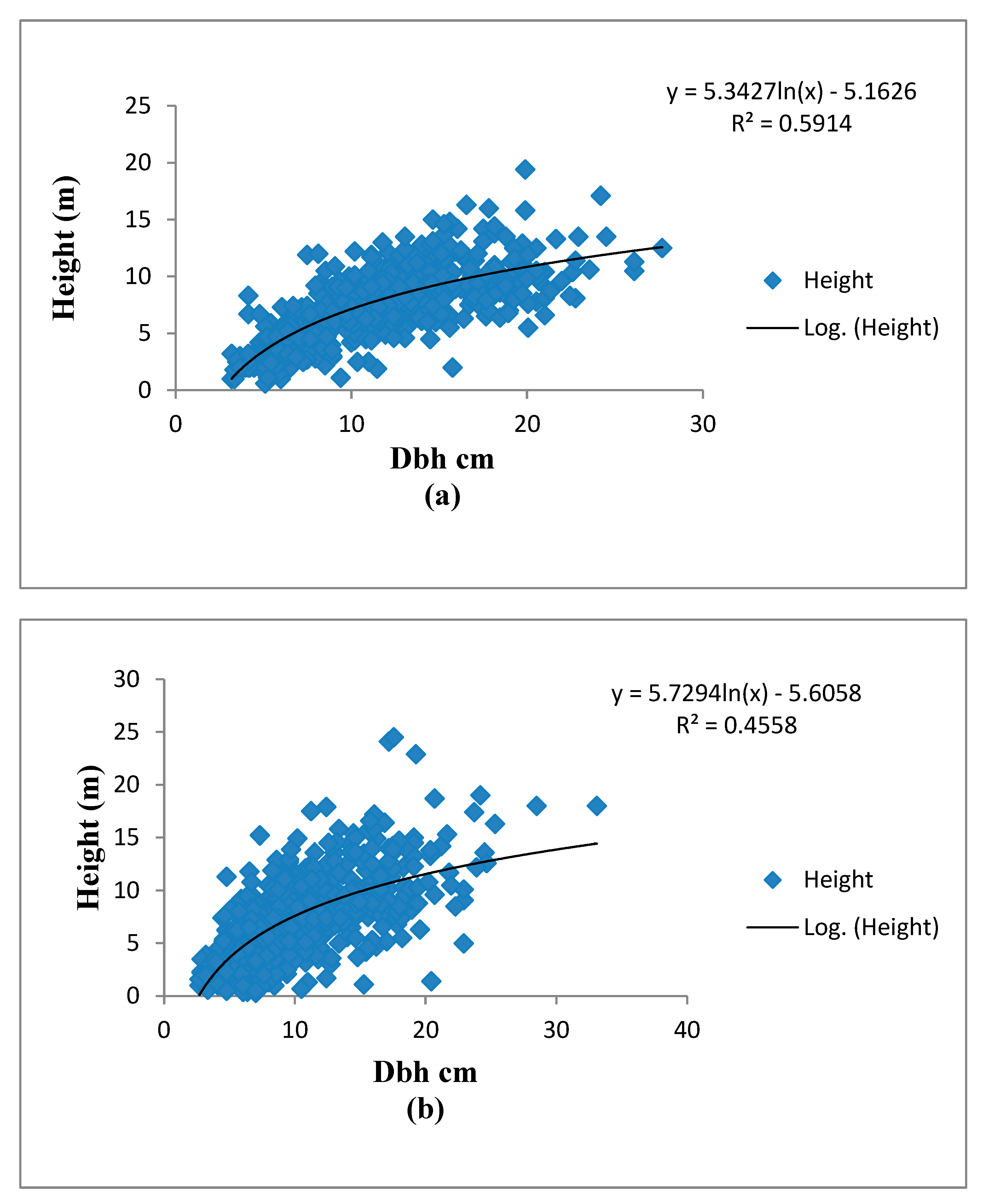
3.1. The H-D Allometry
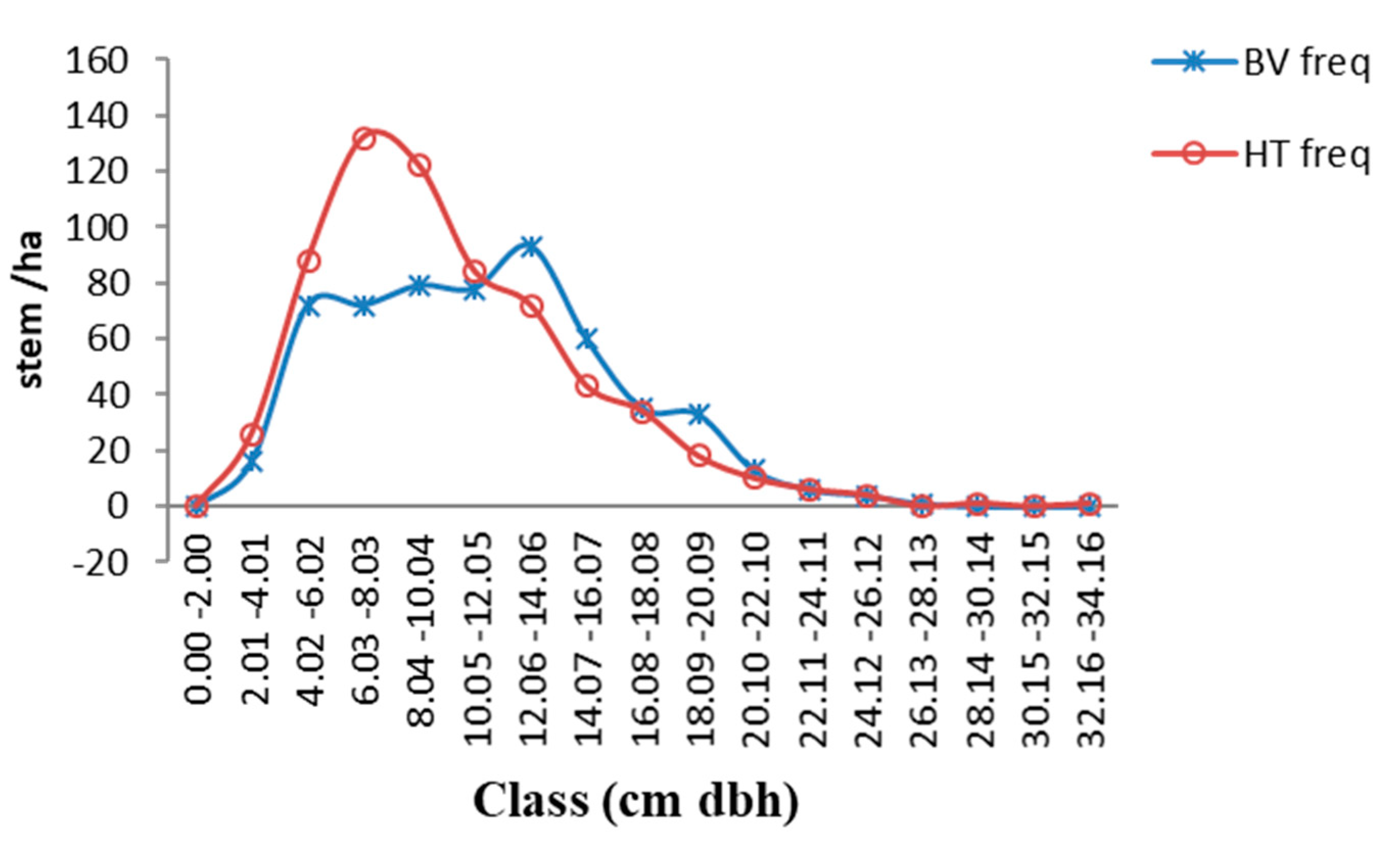
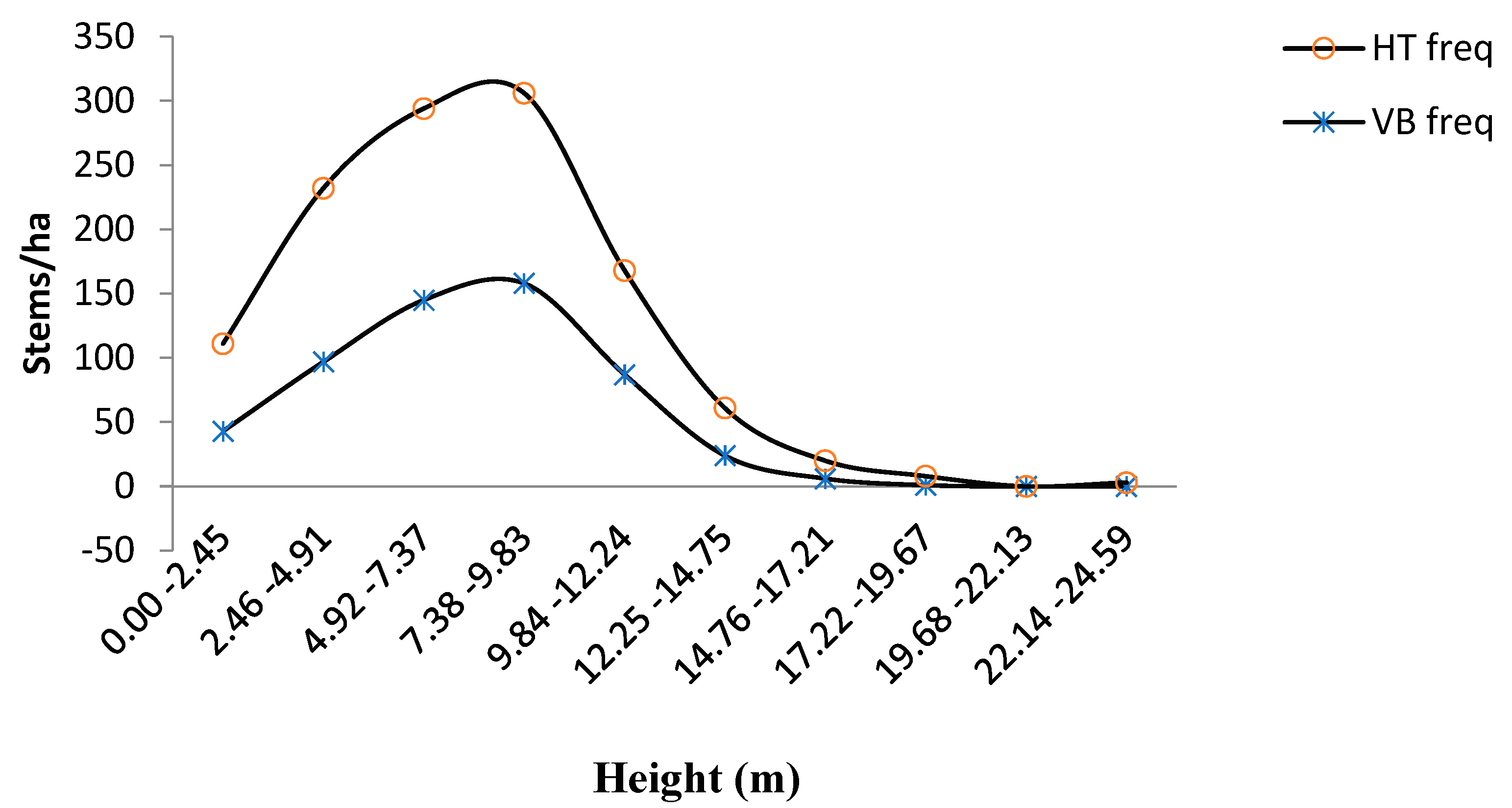
3.2. Diameter-Density and Height-Density Distributions of the Hilltop and Valley-Bottom Stands
3.3. Relationship between Inequality Measures and Diversity Indices of the Stem Diameter
3.4. Relationship between Inequality Measures and Diversity Indices of the Stem Height
4. Discussion
4.1. The H-D Allometry of the Hilltop and Valley-Bottom Stands
4.2. Diameter-Density and Height-Density Distribution of the Hilltop and Valley-Bottom Stands
4.3. Correlation between the Inequality and Diversity Measures
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Weiner, J. Asymmetric competition in plant populations. Trends Ecol. Evol. 1990, 5, 360–364. [Google Scholar] [CrossRef]
- Schwinning, S.; Weiner, J. Mechanisms determining the degree of size asymmetry in competition among plants. Oecologia 1998, 113, 447–455. [Google Scholar] [CrossRef] [PubMed]
- Sumata, B. Relationship between size hierarchy and density of trees in a tropical dry deciduous forest of Western India. J. Veg. Sci. 2010, 18, 389–394. [Google Scholar] [CrossRef]
- Wichmann, L. Annual variations in competition symmetry in Even-aged Sitka spruce. Ann. Bot. 2001, 88, 145–151. [Google Scholar] [CrossRef] [Green Version]
- Kroon, J.; Anderson, B.; Mullin, T.J. Genetic variation in the diameter-height relationship Scot pine (Pinus sylvestris). Can. J. For. Res. 2008, 38, 1493–1503. [Google Scholar] [CrossRef]
- Ojo, L.O. The fate of a tropical rainforest in Nigeria: Abeku sector of Omo Forest Reserve. Glob. Nest Int. J. 2004, 6, 116–130. [Google Scholar]
- Bi, H.; Turvey, N. Competition in mixed stands of Pinus radiata and Eucalyptus oblique. J. Appl. Ecol. 1996, 33, 87–99. [Google Scholar] [CrossRef]
- Fulton, M.R. Patterns in height-diameter relationship for selected tree species and site in Eastern Texas. Can. J. For. Res. 1999, 29, 1445–1448. [Google Scholar] [CrossRef]
- Coomes, D.A.; Allen, R.B. Effects of size, competition and altitude on tree growth. J. Ecol. 2007, 95, 1084–1097. [Google Scholar] [CrossRef]
- Hara, T. Dynamics of size structure in plant populations. Trends Ecol. Evol. 1988, 3, 129–133. [Google Scholar] [CrossRef]
- Ford, E.D. Competition and stand structure in some even-aged plant monocultures. J. Ecol. 1975, 63, 311–333. [Google Scholar] [CrossRef]
- Ford, E.D.; Newbould, P.J. Stand structure and dry weight production through the sweet chestnut (Castanea sativa Mill.) coppice cycle. J. Ecol. 1970, 58, 275–296. [Google Scholar] [CrossRef]
- West, P.W.; Borough, C.J. Tree suppression and the self-thinning rule in a monoculture of Pinus radiate D. Don. Ann. Bot. 1983, 52, 149–158. [Google Scholar] [CrossRef]
- Golf, F.G.; West, D.C. Canopy-understory interaction effects on forest population structure. For. Sci. 1975, 21, 98–108. [Google Scholar]
- Blend, M.; Lussier, J.-M.; Bergeron, Y.; Longpre, M.-H.; Beland, M. Structure, spatial distribution and competition in mixed jack pine (Pinus banksiana) stands on clay soils of eastern Canada. Ann. For. Sci. 2003, 60, 609–617. [Google Scholar] [CrossRef] [Green Version]
- Mendez-Alonzo, R.; Hernandez-Trejo, H.; Lope-Portillo, J. Salinity contrains size inequality and allometry in two contrasting mangrove habitats in the Mexico. J. Trop. Ecol. 2012, 28, 171–179. [Google Scholar] [CrossRef]
- Higgins, S.S.; Bendel, R.B.; Mack, R.N. Assessing competition among skewed distributions of plant biomass: An application of the Jackknife. Biometrics 1984, 40, 131–137. [Google Scholar] [CrossRef]
- Knox, R.G.; Peet, R.K.; Christensen, N.L. Population dynamics in Loblolly Pine stands: Changes in Skewness and size inequality. Ecology 1989, 70, 1153–1167. [Google Scholar] [CrossRef]
- Newton, P.F.; Jolliffe, P.A. Aboveground dry matter partitioning, size variation, and competition processes within second-growth black spruce stands. Can. J. For. Res. 1993, 23, 1917–1929. [Google Scholar] [CrossRef]
- Turner, M.D.; Rabinowitz, D. Factors affecting frequency distributions of plant mass: The absence of dominance and suppression in competing and monocultures of Festuca paradoxa. Ecology 1983, 64, 469–475. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Stand | Minimum (cm dbh) | Maximum (cm dbh) | Mean ± Std (cm dbh) | Gini | CV (%) | Skewness | Kurtosis | SD (Stems/ha) |
|---|---|---|---|---|---|---|---|---|
| Hilltop | 2.71 | 23.10 | 10.19 ± 4.62 | 0.24 | 45.37 | 0.97 | 1.18 | 1431.00 |
| Valley-Bottom | 3.18 | 24.68 | 11.30 ± 4.82 | 0.24 | 42.68 | 0.47 | 0.47 | 1251.00 |
| Stand | Minimum (m) | Maximum (m) | Mean ± Std (m) | Gini | CV (%) | Skewness | Kurtosis | SD (Stems/ha) |
|---|---|---|---|---|---|---|---|---|
| Hilltop | 0.30 | 24.50 | 7.12 ± 3.88 | 0.29 | 54.51 | 0.77 | 1.11 | 1431.00 |
| Valley-Bottom | 1.00 | 19.40 | 7.26 ± 3.21 | 0.25 | 44.30 | 0.22 | −0.21 | 1251.00 |
| Index 1 | Index 2 | Correlation Value | At 0.05 Level |
|---|---|---|---|
| Simpson index | Margalef index | 0.956 | 0.011 |
| Evenness | Equitability | 0.955 | 0.011 |
| Skewness coefficient | Margalef index | 0.936 | 0.019 |
| Skewness coefficient | Simpson index | 0.932 | 0.021 |
| Skewness coefficient | Shannon index | 0.905 | 0.034 |
| Evenness | Margalef index | −0.905 | 0.035 |
| Index 1 | Index 2 | Correlation Value | At 0.05 Level |
|---|---|---|---|
| Mean_D | Simpson | 0.915 | 0.029 |
| Mean_D | Stem_density | −0.913 | 0.030 |
| Stem_density | Shannon-Weiner | −0.917 | 0.029 |
| Index 1 | Index 2 | Correlation Value | At 0.05 Level |
|---|---|---|---|
| Simpson | Equitability | 0.953 | 0.012 |
| Simpson | Evenness | 0.918 | 0.028 |
| Index 1 | Index 2 | Correlation Values | At 0.05 Level |
|---|---|---|---|
| Gini-Coefficient | Shannon index | 0.945 | 0.015 |
| Coefficient of Variation | Margalef index | 0.945 | 0.016 |
| Skewness coefficient | Margalef index | 0.944 | 0.016 |
| Evenness | Equitability | 0.941 | 0.017 |
| Coefficient of Variation | Simpson-index | 0.931 | 0.022 |
| Gini-Coefficient | Skewness coefficient | 0.930 | 0.022 |
| Gini-Coefficient | Margalef index | 0.915 | 0.029 |
| Simpson index | Margalef index | 0.914 | 0.030 |
| Coefficient of Variation | Skewness coefficient | 0.901 | 0.037 |
| Skewness coefficient | Shannon-index | 0.883 | 0.047 |
| Gini-Coefficient | Simpson index | 0.878 | 0.050 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Falade, O.F.; Oguntona, S.B. Tree Size Structure of Tectona grandis (Linn f.) Stand in Hilltop and Valley-Bottom of Omo Forest Reserve. Environ. Sci. Proc. 2022, 13, 21. https://doi.org/10.3390/IECF2021-10823
Falade OF, Oguntona SB. Tree Size Structure of Tectona grandis (Linn f.) Stand in Hilltop and Valley-Bottom of Omo Forest Reserve. Environmental Sciences Proceedings. 2022; 13(1):21. https://doi.org/10.3390/IECF2021-10823
Chicago/Turabian StyleFalade, Oladele Fisayo, and Stephen Busola Oguntona. 2022. "Tree Size Structure of Tectona grandis (Linn f.) Stand in Hilltop and Valley-Bottom of Omo Forest Reserve" Environmental Sciences Proceedings 13, no. 1: 21. https://doi.org/10.3390/IECF2021-10823
APA StyleFalade, O. F., & Oguntona, S. B. (2022). Tree Size Structure of Tectona grandis (Linn f.) Stand in Hilltop and Valley-Bottom of Omo Forest Reserve. Environmental Sciences Proceedings, 13(1), 21. https://doi.org/10.3390/IECF2021-10823