
Seasonal Pattern in Serum Estradiol, Progesterone, and Prolactin Concentrations in Rescued Wild Female Taiwanese Pangolin (Manis pentadactyla pentadactyla)



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Animals
2.2. Blood Collection and Processing
2.3. Hormone Analysis
2.4. Data Analysis
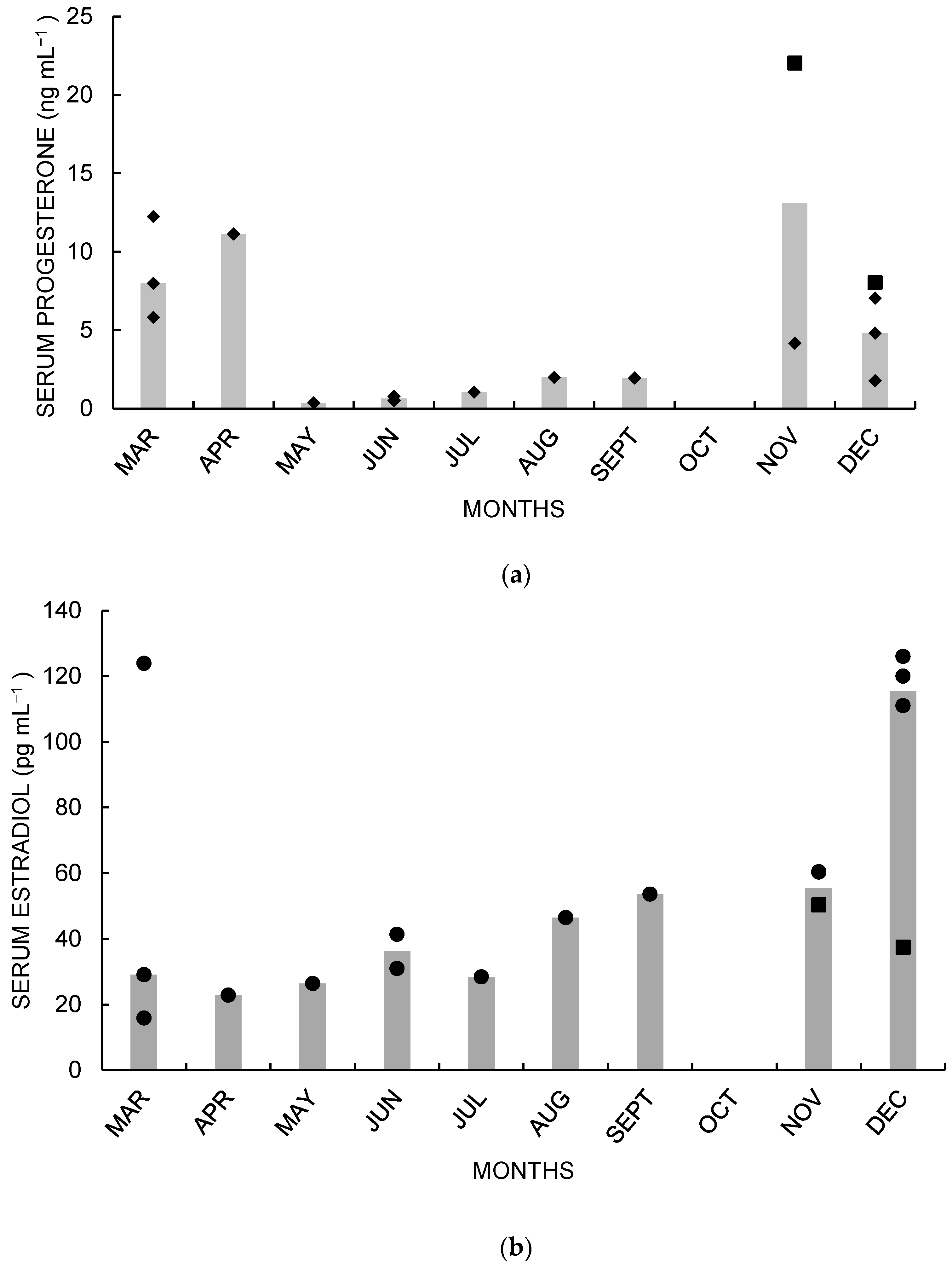
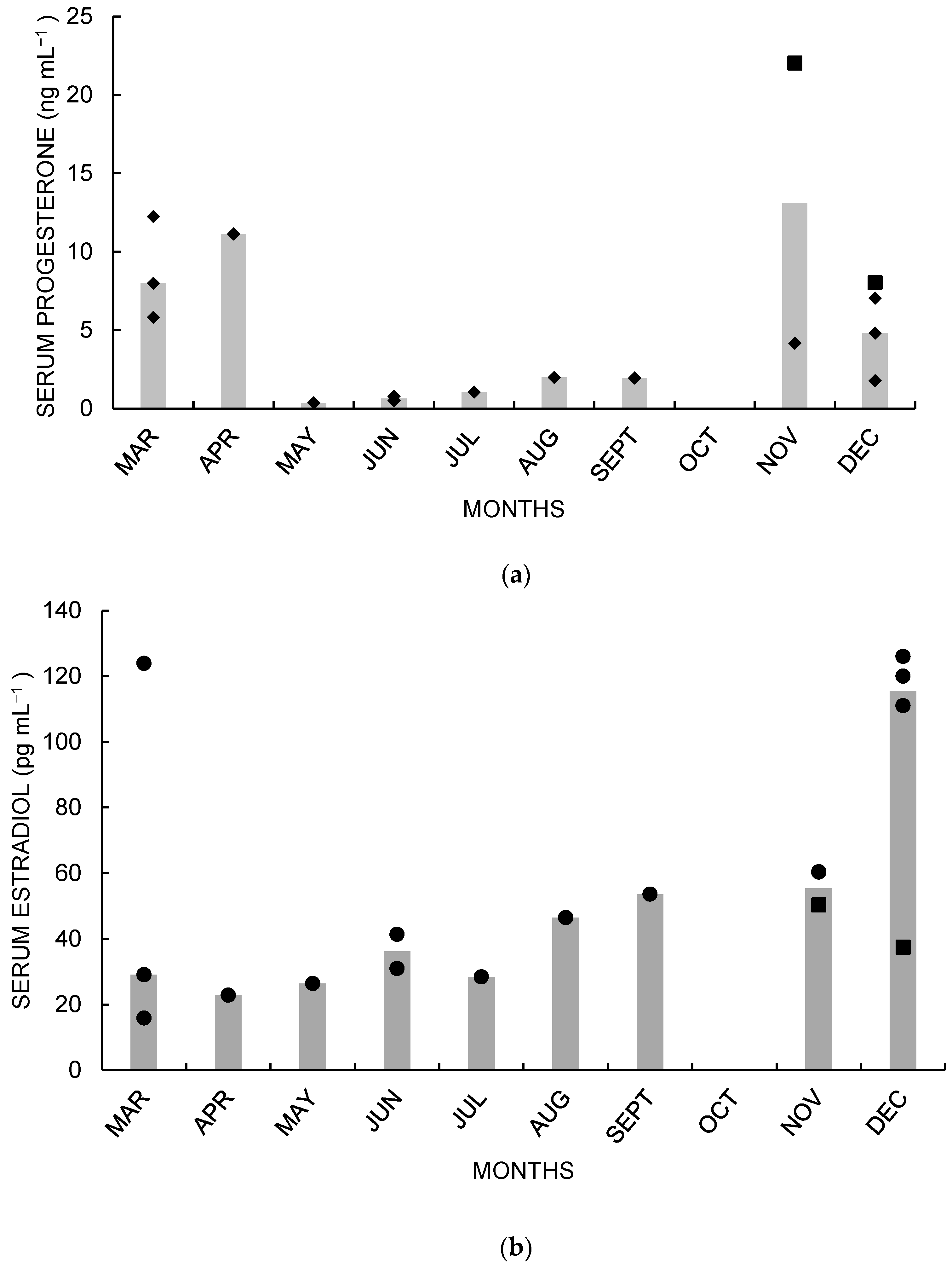
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wu, S.; Sun, N.C.-M.; Zhang, F.; Yu, Y.; Ades, G.; Suwal, T.L.; Jiang, Z. Chinese pangolin Manis pentadactyla (Linnaeus, 1758). In Pangolins; Academic Press; Elsevier: Cambridge, MA, USA, 2020; pp. 49–70. [Google Scholar] [CrossRef]
- Wilson, D.E.; Mittermeier, R.A. Carnivores; Lynx Edicions: Barcelona, Spain, 2009; Volume 1. [Google Scholar]
- Bekoff, M.; Daniels, T.J.; Gittleman, J.L. Life History Patterns and the Comparative Social Ecology of Carnivores. Annu. Rev. Ecol. Syst. 1984, 15, 191–232. [Google Scholar] [CrossRef]
- Gong, S.; Hua, L.; Wang, F.; Li, W.; Ge, Y.; Li, X.; Hou, F. Captive breeding of pangolins: Current status, problems and future prospects. ZooKeys 2015, 507, 99–114. [Google Scholar] [CrossRef] [PubMed]
- Wu, S. Notes on a Newborn Chinese Pangolin (Manis pentadactyla aurita). J. Qinghai Norm. Univ. 1998, 1, 40–42. [Google Scholar]
- Wu, S.; Zhang, F.; Zou, C.; Wang, Q.; Li, S.; Sun, R. A note on captive breeding and reproductive parameters of the Chinese pangolin, Manis pentadactyla Linnaeus, 1758. ZooKeys 2016, 618, 129–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chao, J.; Chen, Y.; Yeh, W.; Fang, K. Notes on a Newborn Formosan Pangolin, Manis pentadactyla pentadactyla. J. Taiwan Mus. 1993, 46, 43–46. [Google Scholar] [CrossRef]
- Yang, C.W.; Chen, S.; Chang, C.Y.; Lin, M.F.; Block, E.; Lorentsen, R.; Chin, J.S.; Dierenfeld, E.S. History and dietary husbandry of pangolins in captivity. Zoo Biol. 2007, 26, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Heath, M.E. Manis temminckii. Am. Soc. Mammal. 1992, 415, 1–5. [Google Scholar] [CrossRef]
- Heath, M.E. Manis pentadactyla. Am. Soc. Mammal. 1992, 414, 1–6. [Google Scholar] [CrossRef]
- Challender, D.; Hywood, L. African pangolins under increased pressure from poaching and intercontinental trade. TRAFFIC Bull. 2012, 24, 53–55. [Google Scholar]
- Challender, D.W.; Harrop, S.R.; MacMillan, D.C. Understanding markets to conserve trade-threatened species in CITES. Biol. Conserv. 2015, 187, 249–259. [Google Scholar] [CrossRef] [Green Version]
- Nash, H.C.; Wong, M.H.; Turvey, S.T. Using local ecological knowledge to determine status and threats of the Critically Endangered Chinese pangolin (Manis pentadactyla) in Hainan, China. Biol. Conserv. 2016, 196, 189–195. [Google Scholar] [CrossRef] [Green Version]
- Crandall, L.S. The Management of Wild Mammals in Captivity; University of Chicago Press: Chicago, IL, USA, 1964. [Google Scholar]
- Heath, M.E.; Vanderlip, S.L. Biology, husbandry, and veterinary care of captive Chinese pangolins (Manis pentadactyla). Zoo Biol. 1988, 7, 293–312. [Google Scholar] [CrossRef]
- Wilson, A.E. Husbandry of pangolins Manis spp. Int. Zoo Yearb. 1994, 33, 248–251. [Google Scholar] [CrossRef]
- Lim, N.T. Autecology of the Sunda Pangolin (Manis javanica) in Singapore; National University of Singapore: Singapore, 2008. [Google Scholar]
- Chin, S.-C.; Lien, C.-Y.; Chan, Y.-T.; Chen, C.-L.; Yang, Y.-C.; Yeh, L.-S. Monitoring the gestation period of rescued Formosan pangolin (Manis pentadactyla pentadactyla) with progesterone radioimmunoassay. Zoo Biol. 2011, 31, 479–489. [Google Scholar] [CrossRef]
- Arora, B.; Pei, K.J.-C.; Weng, C.F.; Sun, N.C.-M. Measuring fecal metabolites of endogenous steroids using ESI-MS/MS spectra in Taiwanese pangolin, (order Pholidota, family Manidae, Genus: Manis): A non-invasive method for endangered species. Gen. Comp. Endocrinol. 2020, 299, 113607. [Google Scholar] [CrossRef]
- Sun, N.C.-M.; Sompud, J.; Pei, K.J.-C. Nursing Period, Behavior Development, and Growth Pattern of a Newborn Formosan Pangolin (Manis pentadactyla pentadactyla) in the Wild. Trop. Conserv. Sci. 2018, 11, 1940082918788450. [Google Scholar] [CrossRef]
- Zhang, F.; Wu, S.; Yang, L.; Zhang, L.; Sun, R.; Li, S. Reproductive parameters of the Sunda pangolin, Manis javanica. Folia Zool. 2015, 64, 129–136. [Google Scholar] [CrossRef]
- Zhang, F.; Yu, J.; Wu, S.; Li, S.; Zou, C.; Wang, Q.; Sun, R. Keeping and breeding the rescued Sunda pangolins (Manis javanica) in captivity. Zoo Biol. 2017, 36, 387–396. [Google Scholar] [CrossRef]
- Payne, J.; Francis, C. A Field Guide to the Mammals of Borneo; The Sabah Society: Kota, Malaysia, 1998; p. 228. [Google Scholar]
- Wang, P.J. Application of Wildlife Rescue System in Conservation of the Formosan Pangolin (Manis pentadactyla pentadactyla). Master’s Thesis, National Taiwan University, Taipei, Taiwan, 2007. [Google Scholar]
- Sun, N.C.M.; Lo, F.H.Y.; Chen, B.Y.; Yu, H.Y.; Liang, C.C.; Lin, C.C.; Chin, S.C.; Li, H.F. Digesta retention time and recovery rates of ants and termites in Chinese pangolins (Manis pentadactyla). Zoo Biol. 2020, 39, 168–175. [Google Scholar] [CrossRef]
- Khatri-Chhetri, R.; Sun, C.-M.; Wu, H.-Y.; Pei, K.J.-C. Reference intervals for hematology, serum biochemistry, and basic clinical findings in free-ranging Chinese Pangolin (Manis pentadactyla) from Taiwan. Vet. Clin. Pathol. 2015, 44, 380–390. [Google Scholar] [CrossRef] [Green Version]
- Sun, N.C.-M.; Pei, K.J.-C.; Wu, L.-Y. Reproductive Behaviors of Wild Chinese Pangolin (Manis Pentadactyla): A Case Study Based on Long-Term Monitoring. Res. Sq. 2021. preprint. [Google Scholar] [CrossRef]
- Andersen, M.L.; Tufik, S. Does male sexual behavior require progesterone? Brain Res. Rev. 2006, 51, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Gentry, R.L. Northern fur Seal—Callorhinus ursinus; Academic Press: London, UK, 1981. [Google Scholar]
- Yan, D.; Zeng, X.; Jia, M.; Guo, X.; Deng, S.; Tao, L.; Huang, X.; Li, B.; Huang, C.; Que, T. Successful captive breeding of a Malayan pangolin population to the third filial generation. Commun. Biol. 2021, 4, 1212. [Google Scholar] [CrossRef] [PubMed]
- Arnason, U.; Adegoke, J.A.; Bodin, K.; Born, E.W.; Esa, Y.B.; Gullberg, A.; Nilsson, M.; Short, R.V.; Xu, X.; Janke, A. Mammalian mitogenomic relationships and the root of the eutherian tree. Proc. Natl. Acad. Sci. USA 2002, 99, 8151–8156. [Google Scholar] [CrossRef] [Green Version]
- Mead, R.A. Delayed implantation in mustelids, with special emphasis on the spotted skunk. J. Reprod. Fertil. Suppl. 1981, 29, 11–24. [Google Scholar]
- Mead, R.A. Role of the Corpus Luteum in Controlling Implantation in Mustelid Carnivores. Ann. N. Y. Acad. Sci. 1986, 476, 25–35. [Google Scholar] [CrossRef]
- Donato, J., Jr.; Frazão, R. Interactions between prolactin and kisspeptin to control reproduction. Arch. Endocrinol. Metab. 2016, 60, 587–595. [Google Scholar] [CrossRef] [Green Version]
- Boyd, I.L. Changes in plasma progesterone and prolactin concentrations during the annual cycle and the role of prolactin in the maintenance of lactation and luteal development in the Antarctic fur seal (Arctocephalus gazella). Reproduction 1991, 91, 637–647. [Google Scholar] [CrossRef] [Green Version]
- Masui, M. Birth of a Chinese pangolin Manis pentadactyle at Ueno Zoo, Tokyo. Int. Zoo Yearb. 1967, 7, 114–116. [Google Scholar] [CrossRef]
- Ee, C.A. A note on breeding the cape pangolin Manis temniincki at bloemfontein zoo. Int. Zoo Yearb. 1966, 6, 163–164. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Individual ID | Month/Year of Rescue | Body Weight (kg) | Number of Blood Samples Collected | Month/Year of Blood Collection |
|---|---|---|---|---|
| 1010313P01 | March, 2012 | 3.78 | 1 | March, 2012 |
| 1010327P01 | March, 2012 | 3.78 | 1 | March 2012 |
| 1050323P01 | March, 2016 | 3.43 | 1 | March, 2016 |
| 1060413P01 | April, 2017 | 3.48 | 2 | April 2017 |
| 1060327P01 | March, 2017 | 1.3 | 1 | May, 2017 |
| 1050527P01 | May, 2016 | 1.70 | 1 | June 2016 |
| 1030226P01 | February, 2014 | 2.06 | 1 | June, 2014 |
| 1060703P01 | July, 2017 | 1.36 | 2 | July, 2017 |
| 1030821P01 | August, 2014 | 1.33 | 1 | August, 2014 |
| 1070917P01 | September, 2018 | 2.58 | 1 | September, 2018 |
| 1041115P01 | November, 2015 | 4.40 | 1 | November, 2015 |
| 1051103P01 | November, 2016 | 2.1 | 1 | November, 2016 |
| 1041213P01 | December, 2015 | 2.08 | 2 | December, 2015 |
| 1051205P01 | December, 2016 | 4.6 | 2 | December, 2016 |
| 1051225P01 | December, 2016 | 2.59 | 2 | December, 2016 |
| 1061225P01 | December, 2017 | 3.94 | 3 | December, 2017 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arora, B.; Pei, K.J.-C.; Ganswindt, A. Seasonal Pattern in Serum Estradiol, Progesterone, and Prolactin Concentrations in Rescued Wild Female Taiwanese Pangolin (Manis pentadactyla pentadactyla). J. Zool. Bot. Gard. 2022, 3, 238-245. https://doi.org/10.3390/jzbg3020019
Arora B, Pei KJ-C, Ganswindt A. Seasonal Pattern in Serum Estradiol, Progesterone, and Prolactin Concentrations in Rescued Wild Female Taiwanese Pangolin (Manis pentadactyla pentadactyla). Journal of Zoological and Botanical Gardens. 2022; 3(2):238-245. https://doi.org/10.3390/jzbg3020019
Chicago/Turabian StyleArora, Bharti, Kurtis Jai-Chyi Pei, and Andre Ganswindt. 2022. "Seasonal Pattern in Serum Estradiol, Progesterone, and Prolactin Concentrations in Rescued Wild Female Taiwanese Pangolin (Manis pentadactyla pentadactyla)" Journal of Zoological and Botanical Gardens 3, no. 2: 238-245. https://doi.org/10.3390/jzbg3020019
APA StyleArora, B., Pei, K. J.-C., & Ganswindt, A. (2022). Seasonal Pattern in Serum Estradiol, Progesterone, and Prolactin Concentrations in Rescued Wild Female Taiwanese Pangolin (Manis pentadactyla pentadactyla). Journal of Zoological and Botanical Gardens, 3(2), 238-245. https://doi.org/10.3390/jzbg3020019