

Trends in Biological Ammonia Production
 ,
,
Abstract
1. Introduction
2. Economic Importance of Ammonia
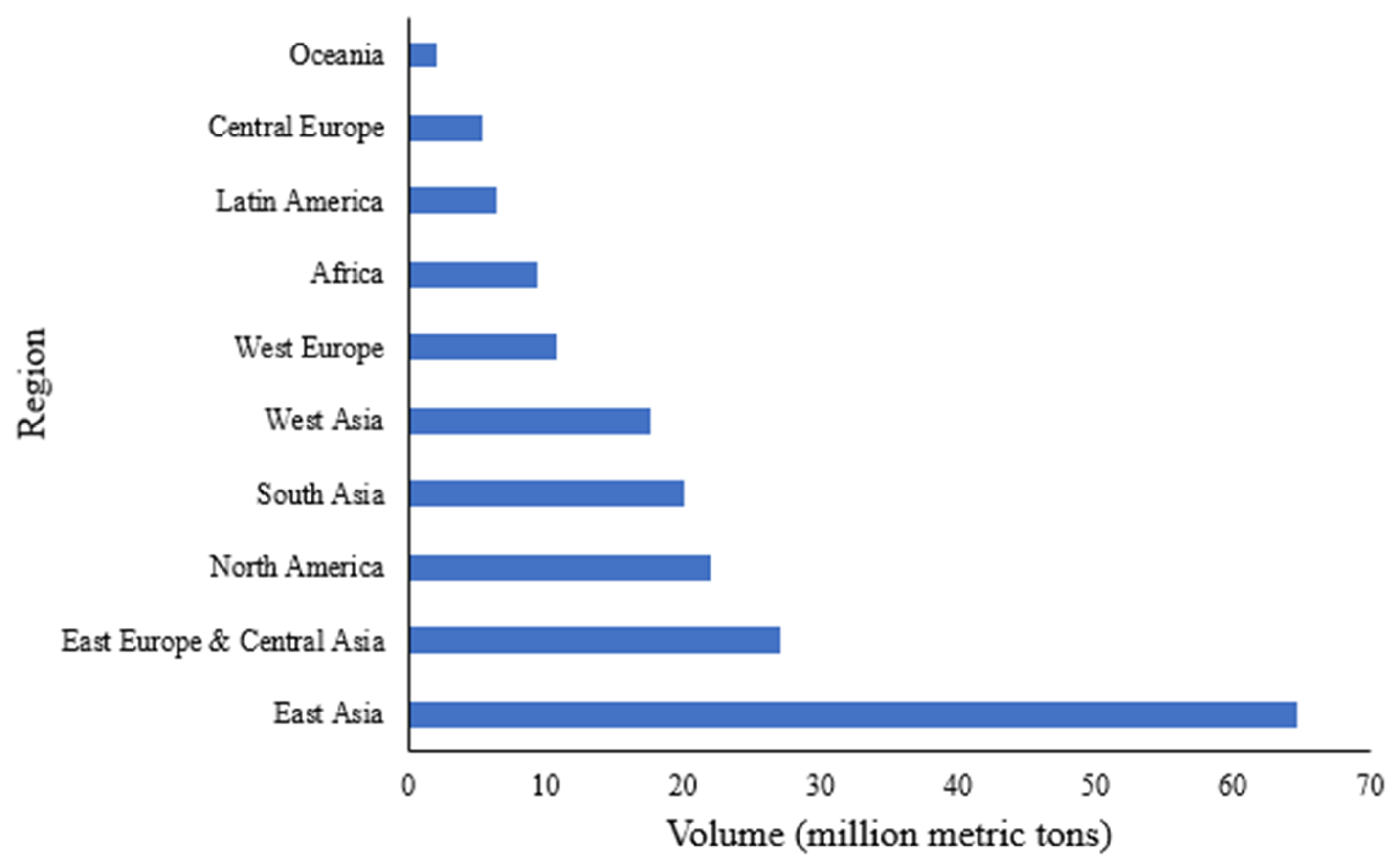
2.1. Scale of Production
2.2. Application as Fertilizer
2.3. Fuel Potential
3. Ammonia Classification
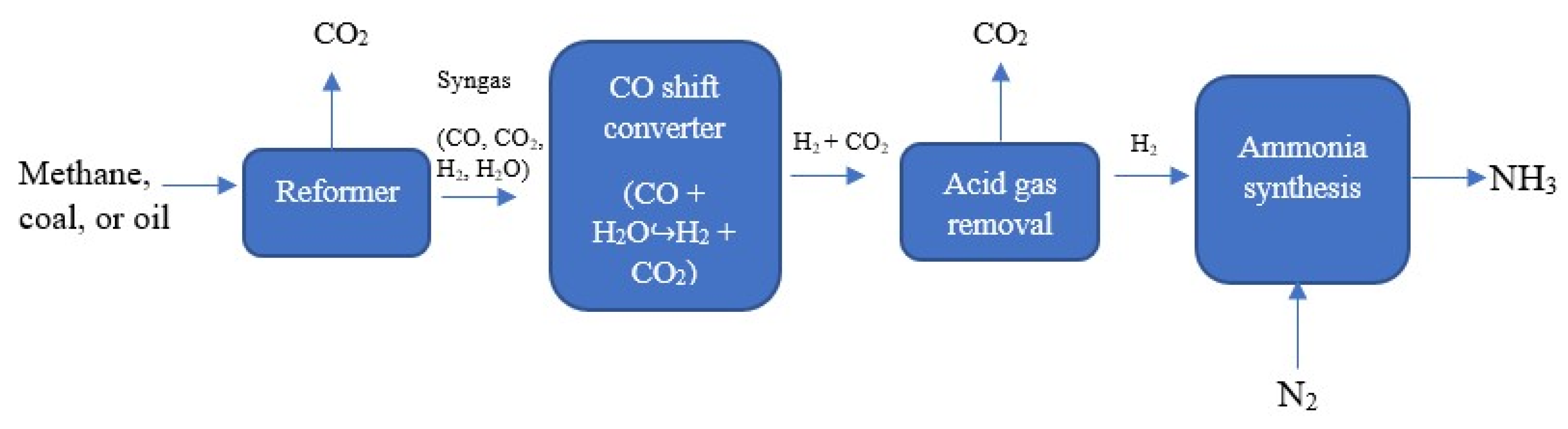
3.1. Brown (or Grey) Ammonia
3.2. Blue Ammonia
3.3. Green Ammonia
4. Biological Ammonia Production
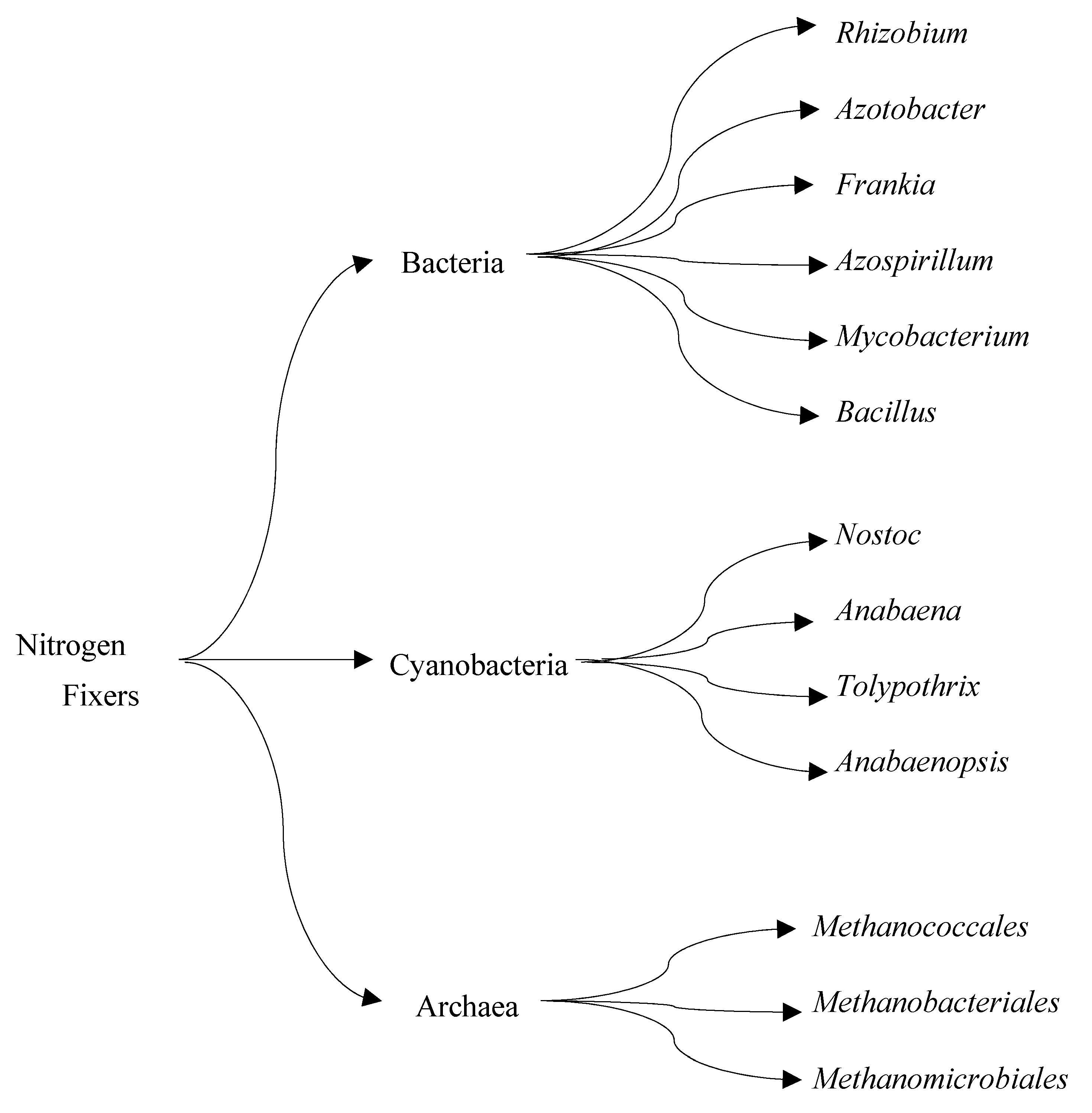
4.1. Biological Nitrogen Fixation by Nitrogenase
4.2. Cell and Metabolic Engineering for Ammonia Production
4.3. Ammonia from Wastewater Treatment Plants
4.4. Hyper Ammonia-Producing Bacteria Route
5. Biomanufacturing
5.1. Conceptual Bioprocess Flow
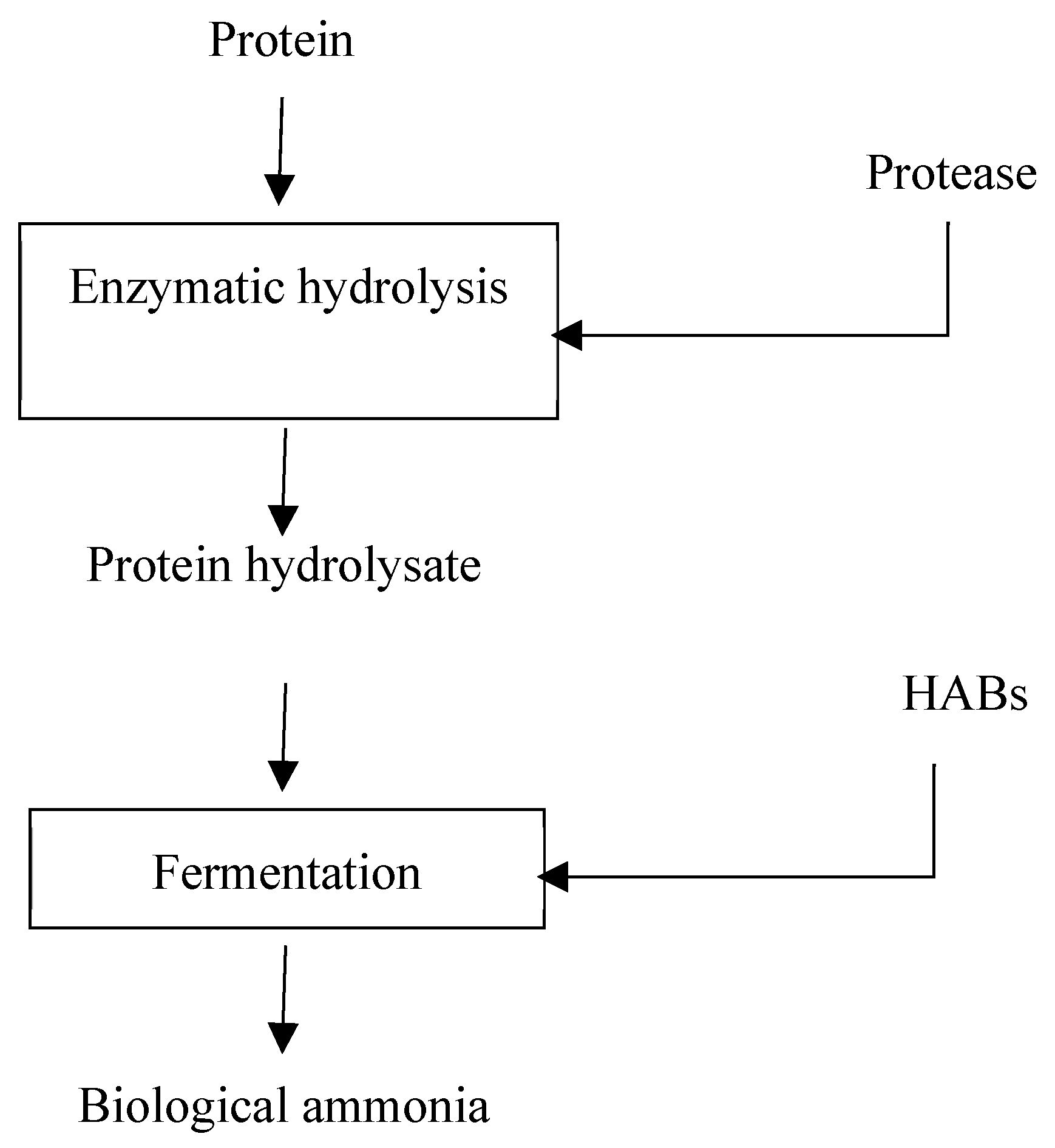
5.2. Protein Hydrolysis
5.2.1. Biological/Enzymatic Protein Hydrolysis
5.2.2. Multi-Enzymatic Hydrolysis
5.3. Leading HABs for Biological Ammonia Production
5.4. Factors Affecting Biological Ammonia Production
5.4.1. Effect of Diet, Substrate, and Substrate Combination
5.4.2. Effect of pH
5.4.3. Effect of Temperature
5.4.4. Effect of Time
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pattabathula, V.; Richardson, J. Introduction to Ammonia Production. Chem. Eng. Prog. 2016, 112, 69–75. [Google Scholar]
- Norskov, J.; Chen, J.; Bullock, M.; Chirik, P.; Chorkendorff, I. Sustainable Ammonia Synthesis; DOE Roundtable Report. In Proceedings of the DOE Roundtable Discussion, Dulles, VA, USA, 18 February 2016. [Google Scholar]
- Ghavam, S.; Vahdati, M.; Wilson, I.G.; Styring, P. Sustainable Ammonia Production Processes. Front. Energy Res. 2021, 9, 580808. [Google Scholar] [CrossRef]
- Kojima, Y. Physical and Chemical Properties of Ammonia as Energy and Hydrogen Carriers. In CO2 Free Ammonia as an Energy Carrier: Japan’s Insights; Springer: Berlin/Heidelberg, Germany, 2022; pp. 17–28. [Google Scholar]
- Stoecker, W.F. Industrial Refrigeration Handbook; McGraw-Hill Education: New York, NY, USA, 1998; ISBN 9780070616233. [Google Scholar]
- Smith, C.; Hill, A.K.; Torrente-Murciano, L. Current and Future Role of Haber-Bosch Ammonia in a Carbon-Free Energy Landscape. Energy Environ. Sci. 2020, 13, 331–344. [Google Scholar] [CrossRef]
- Smil, V. Enriching the Earth: Fritz Haber, Carl Bosch, and the Transformation of World Food Production; MIT Press: Cambridge, MA, USA, 2004. [Google Scholar]
- Liu, B.; Manavi, N.; Deng, H.; Huang, C.; Shan, N.; Chikan, V.; Pfromm, P. Activation of N2 on Manganese Nitride-Supported Ni3 and Fe3 Clusters and Relevance to Ammonia Formation. J. Phys. Chem. Lett. 2021, 12, 6535–6542. [Google Scholar] [CrossRef] [PubMed]
- Ibendahl, G. The Russia-Ukraine Conflict and the Effect on Fertilizer; Department of Agricultural Economics, Kansas State University: Manhattan, KS, USA, 2022. [Google Scholar]
- Ben Hassen, T.; El Bilali, H. Impacts of the Russia-Ukraine War on Global Food Security: Towards More Sustainable and Resilient Food Systems? Foods 2022, 11, 2301. [Google Scholar] [CrossRef]
- Jagtap, S.; Trollman, H.; Trollman, F.; Garcia-Garcia, G.; Parra-López, C.; Duong, L.; Martindale, W.; Munekata, P.E.; Lorenzo, J.M.; Hdaifeh, A. The Russia-Ukraine Conflict: Its Implications for the Global Food Supply Chains. Foods 2022, 11, 2098. [Google Scholar] [CrossRef]
- Voigt, C.; Lamprecht, R.E.; Marushchak, M.E.; Lind, S.E.; Novakovskiy, A.; Aurela, M.; Martikainen, P.J.; Biasi, C. Warming of Subarctic Tundra Increases Emissions of All Three Important Greenhouse Gases–Carbon Dioxide, Methane, and Nitrous Oxide. Glob. Chang. Biol. 2017, 23, 3121–3138. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, L.; Van, J.; Maréchal, F.; Desideri, U. Techno-Economic Comparison of Green Ammonia Production Processes. Appl. Energy 2019, 259, 114135. [Google Scholar] [CrossRef]
- Bello, I.; Adeniyi, A.; Mukaila, T.; Hammed, A. Optimization of Soybean Protein Extraction with Ammonium Hydroxide (NH4OH) Using Response Surface Methodology. Foods 2023, 12, 1515. [Google Scholar] [CrossRef]
- Bello, I.; Adeniyi, A.; Mukaila, T.; Hammed, A. Developing and Modelling of Sustainable Protein Extraction Using Ammonium Hydroxide–a Recoverable and Reusable Solvent. Food Bioprod. Process. 2023, 140, 16–28. [Google Scholar] [CrossRef]
- Tawalbeh, M.; Murtaza, S.Z.; Al-Othman, A.; Alami, A.H.; Singh, K.; Olabi, A.G. Ammonia: A Versatile Candidate for the Use in Energy Storage Systems. Renew. Energy 2022, 194, 955–977. [Google Scholar] [CrossRef]
- Barton, P.; McCormick, R.H.; Fenske, M.R. Ammonia, Versatile Liquid Extraction Solvent. Ind. Eng. Chem. Process Des. Dev. 1968, 7, 366–371. [Google Scholar] [CrossRef]
- Sánchez, G.; Gaikwad, V.; Holdsworth, C.; Dlugogorski, B.; Kennedy, E.; Stockenhuber, M. Catalytic Conversion of Glycerol to Polymers in the Presence of Ammonia. Chem. Eng. J. 2016, 291, 279–286. [Google Scholar] [CrossRef]
- Fernandez, L. Ammonia Production Worldwide from 2010 to 2021. Available online: https://www.statista.com/statistics/1266378/global-ammonia-production/#:~:text=In2021%2Ctheglobalproduction,approximately%2064.6%20million%20metric%20tons (accessed on 17 May 2023).
- Pfromm, P.H. Towards Sustainable Agriculture: Fossil-Free Ammonia Towards Sustainable Agriculture: Fossil-Free Ammonia. J. Renew. Sustain. Energy 2017, 9, 034702. [Google Scholar] [CrossRef]
- FAO. World Fertilizer Trends and Outlook to 2020; Food and Agriculture Organization of the United Nations: Rome, Italy, 2017. [Google Scholar]
- Bird, F.; Clarke, A.; Davies, P.; Surkovic, E. Ammonia: Zero-Carbon Fertiliser, Fuel and Energy Store; KBR Inc.: London, UK, 2020; ISBN 9781782524489. [Google Scholar]
- Papavisasam, S. Oil and Gas Industry Network. In Corrosion Control in the Oil and Gas Industry; Gulf Professional Publishing: Houston, TX, USA, 2014; pp. 41–131. [Google Scholar]
- Paschkewitz, T.M. Ammonia Production at Ambient Temperature and Pressure. Ph.D. Thesis, University of Iowa, Iowa City, IA, USA, 2012. [Google Scholar]
- Boerner, L.K. Industrial Ammonia Production Emits More CO2 than Any Other Chemical-Making Reaction. Chemists Want to Change That. Chem. Eng. News 2019, 97, 1–9. [Google Scholar]
- Yüzbaşıoğlu, A.E.; Tatarhan, A.H.; Gezerman, A.O. Decarbonization in Ammonia Production, New Technological Methods in Industrial Scale Ammonia Production and Critical Evaluations. Heliyon 2021, 7, e08257. [Google Scholar] [CrossRef]
- Hussin, F.; Aroua, M.K. Recent Trends in the Development of Adsorption Technologies for Carbon Dioxide Capture: A Brief Literature and Patent Reviews (2014–2018). J. Clean. Prod. 2020, 253, 119707. [Google Scholar] [CrossRef]
- Abdelkareem, M.A.; Lootah, M.A.; Sayed, E.T.; Wilberforce, T.; Alawadhi, H.; Yousef, B.A.A.; Olabi, A.G. Fuel Cells for Carbon Capture Applications. Sci. Total Environ. 2021, 769, 144243. [Google Scholar] [CrossRef]
- Kamble, A.R.; Patel, C.M.; Murthy, Z.V.P. A Review on the Recent Advances in Mixed Matrix Membranes for Gas Separation Processes. Renew. Sustain. Energy Rev. 2021, 145, 111062. [Google Scholar] [CrossRef]
- Mutch, G.A.; Qu, L.; Triantafyllou, G.; Xing, W.; Fontaine, M.L.; Metcalfe, I.S. Supported Molten-Salt Membranes for Carbon Dioxide Permeation. J. Mater. Chem. A 2019, 7, 12951–12973. [Google Scholar] [CrossRef]
- Siqueira, R.M.; Freitas, G.R.; Peixoto, H.R.; Nascimento, J.F.D.; Musse, A.P.S.; Torres, A.E.B.; Azevedo, D.C.S.; Bastos-Neto, M. Carbon Dioxide Capture by Pressure Swing Adsorption. Energy Procedia 2017, 114, 2182–2192. [Google Scholar] [CrossRef]
- Bello, I.; Rasaq, N.; Adeniyi, A.; Hammed, A. Enzyme Aided Processing of Oil. Int. J. Halal Res. 2021, 3, 60–72. [Google Scholar] [CrossRef]
- Miller, H.A.; Bouzek, K.; Hnat, J.; Loos, S.; Bernäcker, C.I.; Weißgärber, T.; Röntzsch, L.; Meier-Haack, J. Green Hydrogen from Anion Exchange Membrane Water Electrolysis: A Review of Recent Developments in Critical Materials and Operating Conditions. Sustain. Energy Fuels 2020, 4, 2114–2133. [Google Scholar] [CrossRef]
- Yang, J.; Weng, W.; Xiao, W. Electrochemical Synthesis of Ammonia in Molten Salts. J. Energy Chem. 2020, 43, 195–207. [Google Scholar] [CrossRef]
- Zhang, L.; Ji, X.; Ren, X.; Ma, Y.; Shi, X.; Tian, Z.; Asiri, A.M.; Chen, L.; Tang, B.; Sun, X. Electrochemical Ammonia Synthesis via Nitrogen Reduction Reaction on a MoS2 Catalyst: Theoretical and Experimental Studies. Adv. Mater. 2018, 30, 1800191. [Google Scholar] [CrossRef]
- Casallas, C.; Dincer, I. Assessment of an Integrated Solar Hydrogen System for Electrochemical Synthesis of Ammonia. Int. J. Hydrogen Energy 2017, 42, 21495–21500. [Google Scholar] [CrossRef]
- Giddey, S.; Badwal, S.P.S.; Kulkarni, A. Review of Electrochemical Ammonia Production Technologies and Materials. Int. J. Hydrogen Energy 2013, 38, 14576–14594. [Google Scholar] [CrossRef]
- Frattini, D.; Cinti, G.; Bidini, G.; Desideri, U.; Cioffi, R.; Jannelli, E. A System Approach in Energy Evaluation of Different Renewable Energies Sources Integration in Ammonia Production Plants. Renew. Energy 2016, 99, 472–482. [Google Scholar] [CrossRef]
- Boyd, E.S.; Peters, J.W. New Insights into the Evolutionary History of Biological Nitrogen Fixation. Front. Microbiol. 2013, 4, 201. [Google Scholar] [CrossRef]
- Swarnalakshmi, K.; Yadav, V.; Murugeasn, S.; Dhar, D. Biofertilizers for Higher Pulse Production in India: Scope, Accessibility and Challenges. Indian J. Agron. 2016, 61, 173–181. [Google Scholar]
- Khosro, M.; Yousef, S. Bacterial Biofertilizers for Sustainable Crop Production: A Review. J. Agric. Bological Sci. 2012, 7, 307–316. [Google Scholar]
- Rapson, T.D.; Wood, C.C. Analysis of the Ammonia Production Rates by Nitrogenase. Catalysts 2022, 12, 844. [Google Scholar] [CrossRef]
- Lindström, K.; Mousavi, S.A. Effectiveness of Nitrogen Fixation in Rhizobia. Microb. Biotechnol. 2020, 13, 1314–1335. [Google Scholar] [CrossRef] [PubMed]
- Philippot, L.; Raaijmakers, J.M.; Lemanceau, P.; Van Der Putten, W.H. Going Back to the Roots: The Microbial Ecology of the Rhizosphere. Nat. Rev. Microbiol. 2013, 11, 789–799. [Google Scholar] [CrossRef]
- Temme, K.; Zhao, D.; Voigt, C.A. Refactoring the Nitrogen Fixation Gene Cluster from Klebsiella Oxytoca. Proc. Natl. Acad. Sci. USA 2012, 109, 7085–7090. [Google Scholar] [CrossRef]
- López-Torrejón, G.; Burén, S.; Veldhuizen, M.; Rubio, L.M. Biosynthesis of Cofactor-Activatable Iron-Only Nitrogenase in Saccharomyces Cerevisiae. Microb. Biotechnol. 2021, 14, 1073–1083. [Google Scholar] [CrossRef] [PubMed]
- Takimoto, R.; Tatemichi, Y.; Aoki, W.; Kosaka, Y.; Minakuchi, H.; Ueda, M.; Kuroda, K. A Critical Role of an Oxygen-Responsive Gene for Aerobic Nitrogenase Activity in Azotobacter Vinelandii and Its Application to Escherichia coli. Sci. Rep. 2022, 12, 4182. [Google Scholar] [CrossRef]
- Wang, D.; Xu, A.; Elmerich, C.; Ma, L.Z. Biofilm Formation Enables Free-Living Nitrogen-Fixing Rhizobacteria to Fix Nitrogen under Aerobic Conditions. ISME J. 2017, 11, 1602–1613. [Google Scholar] [CrossRef]
- Bhatti, M.; Feng, P.C.C.; Pitkin, J. Methods and Compositions for Improving Plant Health. U.S. Patent 8,754,011, 17 June 2014. [Google Scholar]
- Yenigün, O.; Demirel, B. Ammonia Inhibition in Anaerobic Digestion: A Review. Process Biochem. 2013, 48, 901–911. [Google Scholar] [CrossRef]
- Whelan, M.J.; Everitt, T.; Villa, R. A Mass Transfer Model of Ammonia Volatilisation from Anaerobic Digestate. Waste Manag. 2010, 30, 1808–1812. [Google Scholar] [CrossRef]
- Walker, M.; Iyer, K.; Heaven, S.; Banks, C.J. Ammonia Removal in Anaerobic Digestion by Biogas Stripping: An Evaluation of Process Alternatives Using a First Order Rate Model Based on Experimental Findings. Chem. Eng. J. 2011, 178, 138–145. [Google Scholar] [CrossRef]
- Choi, K.; Wernick, D.G.; Tat, C.A.; Liao, J.C. Consolidated Conversion of Protein Waste into Biofuels and Ammonia Using Bacillus Subtilis. Metab. Eng. 2014, 23, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Mikami, Y.; Yoneda, H.; Tatsukami, Y.; Aoki, W.; Ueda, M. Ammonia Production from Amino Acid—Based Biomass—like Sources by Engineered Escherichia coli. AMB Express 2017, 7, 83. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, Y.; Aoki, W.; Ueda, M. Improved Ammonia Production from Soybean Residues by Cell Surface-Displayed l-Amino Acid Oxidase on Yeast. Biosci. Biotechnol. Biochem. 2021, 85, 972–980. [Google Scholar] [CrossRef] [PubMed]
- Tatemichi, Y.; Kuroda, K.; Nakahara, T.; Ueda, M. Efficient Ammonia Production from Food by—Products by Engineered Escherichia coli. AMB Express 2020, 10, 150. [Google Scholar] [CrossRef] [PubMed]
- Ueda, M. Establishment of Cell Surface Engineering and Its Development. Biosci. Biotechnol. Biochem. 2016, 80, 1243–1253. [Google Scholar] [CrossRef]
- Kuroda, K.; Ueda, M. Cell Surface Engineering of Yeast for Applications in White Biotechnology. Biotechnol. Lett. 2011, 33, 1–9. [Google Scholar] [CrossRef]
- Watanabe, Y.; Kuroda, K.; Tatemichi, Y.; Nakahara, T.; Aoki, W.; Ueda, M. Construction of Engineered Yeast Producing Ammonia from Glutamine and Soybean Residues (Okara). AMB Express 2020, 10, 70. [Google Scholar] [CrossRef]
- Lu, P.; Ma, D.; Chen, Y.; Guo, Y.; Chen, G.Q.; Deng, H.; Shi, Y. L-Glutamine Provides Acid Resistance for Escherichia coli through Enzymatic Release of Ammonia. Cell Res. 2013, 23, 635–644. [Google Scholar] [CrossRef]
- Bloess, S.; Beuel, T.; Krüger, T.; Sewald, N.; Dierks, T.; Fischer von Mollard, G. Expression, Characterization, and Site-Specific Covalent Immobilization of an L-Amino Acid Oxidase from the Fungus Hebeloma Cylindrosporum. Appl. Microbiol. Biotechnol. 2019, 103, 2229–2241. [Google Scholar] [CrossRef]
- Fenner, K.; Men, Y. Comment on “Role of Ammonia Oxidation in Organic Micropollutant Transformation during Wastewater Treatment”: Overlooked Evidence to the Contrary. Environ. Sci. Technol. 2021, 55, 12128–12129. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Peng, Y.; Ma, B.; Wang, S.; Zhu, G. Anaerobic Ammonium Oxidation in Traditional Municipal Wastewater Treatment Plants with Low-Strength Ammonium Loading: Widespread but Overlooked. Water Res. 2015, 84, 66–75. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Singh, L.; Zularisam, A.W. Exoelectrogens: Recent Advances in Molecular Drivers Involved in Extracellular Electron Transfer and Strategies Used to Improve It for Microbial Fuel Cell Applications. Renew. Sustain. Energy Rev. 2016, 56, 1322–1336. [Google Scholar] [CrossRef]
- Ye, Y.; Ngo, H.H.; Guo, W.; Liu, Y.; Chang, S.W.; Nguyen, D.D.; Liang, H.; Wang, J. A Critical Review on Ammonium Recovery from Wastewater for Sustainable Wastewater Management. Bioresour. Technol. 2018, 268, 749–758. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Monedero, M.A.; Roig, A.; Paredes, C.; Bernal, M.P. Nitrogen Transformation during Organic Waste Composting by the Rutgers System and Its Effects on PH, EC and Maturity of the Composting Mixtures. Bioresour. Technol. 2001, 78, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.; Yau, Y.; Sze, E.T.-P. Application of Anaerobic Bacterial Ammonification Pretreatment to Microalgal Food Waste Leachate Cultivation and Biofuel Production. Mar. Pollut. Bull. 2020, 153, 111007. [Google Scholar] [CrossRef] [PubMed]
- Bello, I.A. Challenges in Textile Wastewater and Current Palliative Methods: An Overview. IIUM Eng. J. 2017, 18, 71–78. [Google Scholar] [CrossRef]
- Palatsi, J.; Ripoll, F.; Benzal, A.; Pijuan, M.; Romero-Güiza, M.S. Enhancement of Biological Nutrient Removal Process with Advanced Process Control Tools in Full-Scale Wastewater Treatment Plant. Water Res. 2021, 200, 117212. [Google Scholar] [CrossRef]
- Cyprowski, M.; Stobnicka-Kupiec, A.; Lawniczek-Walczyk, A.; Bakal-Kijek, A.; Golofit-Szymczak, M.; Górny, R.L. Anaerobic Bacteria in Wastewater Treatment Plant. Int. Arch. Occup. Environ. Health 2018, 91, 571–579. [Google Scholar] [CrossRef]
- Haynes, R.J. Nitrification. In Mineral Nitrogen in the Plant-Soil System; Academic Press: Cambridge, MA, USA, 1986; pp. 127–165. [Google Scholar]
- Van Kessel, M.A.; Speth, D.R.; Albertsen, M.; Nielsen, P.H.; Op den Camp, H.J.; Kartal, B.; Jetten, M.S.; Lücker, S. Complete Nitrification by a Single Microorganism. Nature 2015, 528, 555–559. [Google Scholar] [CrossRef]
- Foley, J.; De Haas, D.; Hartley, K.; Lant, P. Comprehensive Life Cycle Inventories of Alternative Wastewater Treatment Systems. Water Res. 2010, 44, 1654–1666. [Google Scholar] [CrossRef]
- Iskander, S.M.; Brazil, B.; Novak, J.T.; He, Z. Resource Recovery from Landfill Leachate Using Bioelectrochemical Systems: Opportunities, Challenges, and Perspectives. Bioresour. Technol. 2016, 201, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Kankanamge, N.R.; Chow, C.; Welsh, D.T.; Li, T.; Teasdale, P.R. Removing Ammonium from Water and Wastewater Using Cost-Effective Adsorbents: A Review. J. Environ. Sci. 2018, 63, 174–197. [Google Scholar] [CrossRef] [PubMed]
- Barbera, E.; Bertucco, A.; Kumar, S. Nutrients Recovery and Recycling in Algae Processing for Biofuels Production. Renew. Sustain. Energy Rev. 2018, 90, 28–42. [Google Scholar] [CrossRef]
- He, C.; Wang, K.; Yang, Y.; Amaniampong, P.N.; Wang, J.-Y. Effective Nitrogen Removal and Recovery from Dewatered Sewage Sludge Using a Novel Integrated System of Accelerated Hydrothermal Deamination and Air Stripping. Environ. Sci. Technol. 2015, 49, 6872–6880. [Google Scholar] [CrossRef]
- Huang, H.; Yang, J.; Li, D. Recovery and Removal of Ammonia–Nitrogen and Phosphate from Swine Wastewater by Internal Recycling of Struvite Chlorination Product. Bioresour. Technol. 2014, 172, 253–259. [Google Scholar] [CrossRef]
- Husnain, T.; Mi, B.; Riffat, R. A Combined Forward Osmosis and Membrane Distillation System for Sidestream Treatment. J. Water Resour. Prot. 2015, 7, 1111. [Google Scholar] [CrossRef]
- Katehis, D.; Diyamandoglu, V.; Fillos, J. Stripping and Recovery of Ammonia from Centrate of Anaerobically Digested Biosolids at Elevated Temperatures. Water Environ. Res. 1998, 70, 231–240. [Google Scholar] [CrossRef]
- Kim, T.; An, J.; Jang, J.K.; Chang, I.S. Coupling of Anaerobic Digester and Microbial Fuel Cell for COD Removal and Ammonia Recovery. Bioresour. Technol. 2015, 195, 217–222. [Google Scholar] [CrossRef]
- Kuntke, P.; Śmiech, K.; Bruning, H.; Zeeman, G.; Saakes, M.; Sleutels, T.; Hamelers, H.V.M.; Buisman, C.J.N. Ammonium Recovery and Energy Production from Urine by a Microbial Fuel Cell. Water Res. 2012, 46, 2627–2636. [Google Scholar] [CrossRef]
- Puniya, A.K.; Singh, R.; Kamra, D.N. Rumen Microbiology: From Evolution to Revolution. In Rumen Microbiology: From Evolution to Revolution; Springer: New Delhi, India, 2015; pp. 1–379. [Google Scholar] [CrossRef]
- Rychlik, J.L.; Russell, J.B. Mathematical Estimations of Hyper-Ammonia Producing Ruminal Bacteria and Evidence for Bacterial Antagonism That Decreases Ruminal Ammonia Production. FEMS Microbiol. Ecol. 2000, 32, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Pengpeng, W.; Tan, Z. Ammonia Assimilation in Rumen Bacteria: A Review. Anim. Biotechnol. 2013, 24, 107–128. [Google Scholar] [CrossRef]
- Rychlik, J.L.; Russell, J.B. Bacteriocin-like Activity of Butyrivibrio Fibrisolvens JL5 and Its Effect on Other Ruminal Bacteria and Ammonia Production. Appl. Environ. Microbiol. 2002, 68, 1040–1046. [Google Scholar] [CrossRef] [PubMed]
- Bach, A.; Calsamiglia, S.; Stern, M.D. Nitrogen Metabolism in the Rumen. J. Dairy Sci. 2005, 88, E9–E21. [Google Scholar] [CrossRef]
- Van Soest, P.J. Nitrogen Metabolism. In Nutritional Ecology of the Ruminant; Cornell University Press: Ithaca, NY, USA, 1994. [Google Scholar]
- Rychlik, J.L.; Lavera, R.; Russell, J.B. Amino Acid Deamination by Ruminal Megasphaera elsdenii Strains. Curr. Microbiol. 2002, 45, 340–345. [Google Scholar] [CrossRef] [PubMed]
- Doelle, H.W. Nitrogen Metabolism as an Energy Source for Anaerobic Microorganisms (Clostridium). In Bacterial Metabolism; Elsevier: Amsterdam, The Netherlands, 1969; pp. 402–422. [Google Scholar]
- Strock, J.S. Ammonification. In Encyclopedia of Ecology, Five-Volume Set; Elsevier Inc.: Amsterdam, The Netherlands, 2008; pp. 162–165. [Google Scholar]
- Nolan, J.V. Quantitative Models of Nitrogen Metabolism in Sheep. In Digenstion and Metabolism in the Ruminant; Oxford University Press: Oxford, UK, 1975. [Google Scholar]
- Valente, T.N.P.; da Silva Lima, E.; dos Santos, W.B.R.; Cesário, A.S.; Tavares, C.J.; de Freitas, M.A.M. Ruminal Microorganism Consideration and Protein Used in the Metabolism of the Ruminants: A Review. Afr. J. Microbiol. Res. 2016, 10, 456–464. [Google Scholar]
- Adeniyi, A.; Bello, I.; Mukaila, T.; Monono, E.; Hammed, A. Developing Rumen Mimicry Process for Biological Ammonia Synthesis. Bioprocess Biosyst. Eng. 2023, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Nagaraja, T.G. Microbiology of the Rumen. In Rumenology; Springer: Berlin/Heidelberg, Germany, 2016; pp. 39–61. [Google Scholar]
- Whitehead, T.R.; Cotta, M.A. Isolation and Identification of Hyper-Ammonia Producing Bacteria from Swine Manure Storage Pits. Curr. Microbiol. 2004, 48, 20–26. [Google Scholar] [CrossRef]
- Adeniyi, A.; Bello, I.; Mukaila, T.; Hammed, A. A Review of Microbial Molecular Profiling during Biomass Valorization. Biotechnol. Bioprocess Eng. 2022, 27, 515–532. [Google Scholar] [CrossRef]
- Slyter, L.L. Influence of Acidosis on Rumen Function. J. Anim. Sci. 1976, 43, 910–929. [Google Scholar] [CrossRef]
- Chen, G.; Russell, J.B. More Monensin-Sensitive, Ammonia-Producing Bacteria from the Rumen. Appl. Environ. Microbiol. 1989, 55, 1052–1057. [Google Scholar] [CrossRef] [PubMed]
- McDonald, I.W. The Absorption of Ammonia from the Rumen of the Sheep. Biochem. J. 1948, 42, 584. [Google Scholar] [CrossRef] [PubMed]
- Bryant, M.P. The Characteristics of Strains of Selenomonas Isolated from Bovine Rumen Contents. J. Bacteriol. 1956, 72, 162–167. [Google Scholar] [CrossRef] [PubMed]
- Bryant, M.P.; Small, N. The Anaerobic Monotrichous Butyric Acid-Producing Curved Rod-Shaped Bacteria of the Rumen. J. Bacteriol. 1956, 72, 16–21. [Google Scholar] [CrossRef]
- Ward, B.K.; Dufault, R.J.; Hassell, R.; Cutulle, M.A. Affinity of Hyperammonia-Producing Bacteria to Produce Bioammonium/Ammonia Utilizing Five Organic Nitrogen Substrates for Potential Use as an Organic Liquid Fertilizer. ACS Omega 2018, 3, 11817–11822. [Google Scholar] [CrossRef]
- Fischer, M. Limiting Factors for the Enzymatic Accessibility of Soybean Protein; Wageningen University and Research: Wageningen, The Netherlands, 2006. [Google Scholar]
- Sun, X.S. Bio-Based Polymers and Composites. In Thermal and Mechanical Properties of Soy Proteins; Elsevier Inc.: Amsterdam, The Netherlands, 2005; pp. 292–326. [Google Scholar]
- Wang, L.; Liu, K.; Wang, Z.; Bai, X.; Peng, Q.; Jin, L. Bacterial Community Diversity Associated with Different Utilization Efficiencies of Nitrogen in the Gastrointestinal Tract of Goats. Front. Microbiol. 2019, 10, 239. [Google Scholar] [CrossRef]
- Thorpe, J.; Beal, J.D. Vegetable Protein Meals and the Effects of Enzymes. In Enzymes in Farm Animal Nutrition; Bedford, M.R., Partridge, G.G., Eds.; CABI Publishing: Wallingford, UK, 2000; pp. 125–143. [Google Scholar]
- Bedford, M.R. Exogenous Enzymes in Monogastric Nutrition—Their Current Value and Future Benefits. Anim. Feed. Sci. Technol. 2000, 86, 1–13. [Google Scholar] [CrossRef]
- Hur, S.J.; Lim, B.O.; Decker, E.A.; McClements, D.J. In Vitro Human Digestion Models for Food Applications. Food Chem. 2011, 125, 1–12. [Google Scholar] [CrossRef]
- Wang, R.; Mohammadi, M.; Mahboubi, A.; Taherzadeh, M.J. In-Vitro Digestion Models: A Critical Review for Human and Fish and a Protocol for in-Vitro Digestion in Fish. Bioengineered 2021, 12, 3040–3064. [Google Scholar] [CrossRef]
- Banskota, A.H.; Tibbetts, S.M.; Jones, A.; Stefanova, R.; Behnke, J. Biochemical Characterization and In Vitro Digestibility of Protein Isolates from Hemp (Cannabis sativa L.) By-Products for Salmonid Feed Applications. Molecules 2022, 27, 4794. [Google Scholar] [CrossRef]
- Bohn, T.; Carriere, F.; Day, L.; Deglaire, A.; Egger, L.; Freitas, D.; Golding, M.; Le Feunteun, S.; Macierzanka, A.; Ménard, O. Correlation between in Vitro and in Vivo Data on Food Digestion. What Can We Predict with Static in Vitro Digestion Models? Crit. Rev. Food Sci. Nutr. 2018, 58, 2239–2261. [Google Scholar] [CrossRef] [PubMed]
- Moyano, F.J.; Saenz de Rodriganez, M.A.; Díaz, M.; Tacon, A.G. Application of in Vitro Digestibility Methods in Aquaculture: Constraints and Perspectives. Rev. Aquac. 2015, 7, 223–242. [Google Scholar] [CrossRef]
- Lajterer, C.; Levi, C.S.; Lesmes, U. An in Vitro Digestion Model Accounting for Sex Differences in Gastro-Intestinal Functions and Its Application to Study Differential Protein Digestibility. Food Hydrocoll. 2022, 132, 107850. [Google Scholar] [CrossRef]
- Liu, J.; Chen, J.; Wang, S.; Xie, J.; Wang, Y.; Chai, T.-T.; Ong, M.K.; Wu, J.; Tian, L.; Bai, W. Effects of Monascus Application on in Vitro Digestion and Fermentation Characteristics of Fish Protein. Food Chem. 2022, 377, 132000. [Google Scholar] [CrossRef]
- Lucas-González, R.; Viuda-Martos, M.; Pérez-Alvarez, J.A.; Fernández-López, J. In Vitro Digestion Models Suitable for Foods: Opportunities for New Fields of Application and Challenges. Food Res. Int. 2018, 107, 423–436. [Google Scholar] [CrossRef]
- Xue, P.; Sun, N.; Li, Y.; Cheng, S.; Lin, S. Targeted Regulation of Hygroscopicity of Soybean Antioxidant Pentapeptide Powder by Zinc Ions Binding to the Moisture Absorption Sites. Food Chem. 2018, 242, 83–90. [Google Scholar] [CrossRef]
- Islam, M.; Huang, Y.; Islam, S.; Fan, B.; Tong, L.; Wang, F. Influence of the Degree of Hydrolysis on Functional Properties and Antioxidant Activity of Enzymatic Soybean Protein Hydrolysates. Molecules 2022, 27, 6110. [Google Scholar] [CrossRef]
- Marsman, G.J.P.; Gruppen, H.; Mul, A.J.; Voragen, A.G.J. In Vitro Accessibility of Untreated, Toasted, and Extruded Soybean Meals for Proteases and Carbohydrases. J. Agric. Food Chem. 1997, 45, 4088–4095. [Google Scholar] [CrossRef]
- Lallés, J.P.; Huet, A.; Quillien, L.; Plumb, G.W.; Mills, E.N.C.; Morgen, M.R.A.; Toullec, R. Duodenal Passage of Immunoreactive Glycinin and B−conglycinin from Soya Bean in Preruminant Calves. In Recent Advances of Research in Antinutritional Factors in Legume Seeds and Rapeseed; Jansman, A.J.M., Hill, G.D., Huisman, J., van der Poel, A.F.B., Eds.; Wageningen Press: Wageningen, The Netherlands, 1998; pp. 255–258. [Google Scholar]
- Merz, M.; Eisele, T.; Berends, P.; Appel, D.; Rabe, S.; Blank, I.; Stressler, T.; Fischer, L. Flavourzyme, an Enzyme Preparation with Industrial Relevance: Automated Nine-Step Purification and Partial Characterization of Eight Enzymes. J. Agric. Food Chem. 2015, 63, 5682–5693. [Google Scholar] [CrossRef]
- Nchienzia, H.A.; Morawicki, R.O.; Gadang, V.P. Enzymatic Hydrolysis of Poultry Meal with Endo-and Exopeptidases. Poult. Sci. 2010, 89, 2273–2280. [Google Scholar] [CrossRef]
- Chae, H.J.; In, M.-J.; Kim, M.-H. Process Development for the Enzymatic Hydrolysis of Food Protein: Effects of Pre-Treatment and Post-Treatments on Degree of Hydrolysis and Other Product Characteristics. Biotechnol. Bioprocess Eng. 1998, 3, 35–39. [Google Scholar] [CrossRef]
- Kamnerdpetch, C.; Weiss, M.; Kasper, C.; Scheper, T. An Improvement of Potato Pulp Protein Hydrolyzation Process by the Combination of Protease Enzyme Systems. Enzym. Microb. Technol. 2007, 40, 508–514. [Google Scholar] [CrossRef]
- Hunsakul, K.; Laokuldilok, T.; Sakdatorn, V.; Klangpetch, W.; Brennan, C.S.; Utama-ang, N. Optimization of Enzymatic Hydrolysis by Alcalase and Flavourzyme to Enhance the Antioxidant Properties of Jasmine Rice Bran Protein Hydrolysate. Sci. Rep. 2022, 12, 12582. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Bu, G.; Chen, F. The Influence of Composite Enzymatic Hydrolysis on the Antigenicity of β-Conglycinin in Soy Protein Hydrolysates. J. Food Biochem. 2018, 42, e12544. [Google Scholar] [CrossRef]
- Paster, B.J.; Russell, J.B.; Yang, C.M.J.; Chow, J.M.; Woese, C.R.; Tanner, R. Phylogeny of the Ammonia-Producing Ruminal Bacteria Peptostreptococcus anaerobius, Clostridium sticklandii, and Clostridium aminophilum sp. nov. Int. J. Syst. Evol. Microbiol. 1993, 43, 107–110. [Google Scholar] [CrossRef] [PubMed]
- Balows, A.; Trüper, H.G.; Dworkin, M.; Harder, W.; Schleifer, K.-H. The Prokaryotes: A Handbook on the Biology of Bacteria: Ecophysiology, Isolation, Identification, Applications; Springer: Berlin/Heidelberg, Germany, 1992; Volume 3. [Google Scholar]
- Gano, J.M. Amino Acid-Fermenting Bacteria from the Rumen of Dairy Cattle-Enrichment, Isolation, Characterization, and Interaction with Entodinium Caudatum. Ph.D. Thesis, The Ohio State University, Columbus, OH, USA, 2013. [Google Scholar]
- Chen, G.J.; Russell, J.B. Fermentation of Peptides and Amino Acids by a Monensin-Sensitive Ruminal Peptostreptococcus. Appl. Environ. Microbiol. 1988, 54, 2742–2749. [Google Scholar] [CrossRef] [PubMed]
- Attwood, G.T.; Klieve, A.V.; Ouwerkerk, D.; Patel, B.K. Ammonia-Hyperproducing Bacteria from New Zealand Ruminants. Appl. Environ. Microbiol. 1998, 64, 1796–1804. [Google Scholar] [CrossRef]
- Eschenlauer, S.C.P.; McKain, N.; Walker, N.D.; McEwan, N.R.; Newbold, C.J.; Wallace, R.J. Ammonia Production by Ruminal Microorganisms and Enumeration, Isolation, and Characterization of Bacteria Capable of Growth on Peptides and Amino Acids from the Sheep Rumen. Appl. Environ. Microbiol. 2002, 68, 4925–4931. [Google Scholar] [CrossRef]
- Kuiken, K.A.; Lyman, C.M.; Bradford, M.; Trant, M.; Dieterich, S. Essential Amino Acid Composition of Soy Bean Meals Prepared from Twenty Strains of Soy Beans. J. Biol. Chem. 1949, 177, 29–36. [Google Scholar] [CrossRef]
- Bladen, H.A.; Bryant, M.P.; Doetsch, R.N. A Study of Bacterial Species from the Rumen Which Produce Ammonia from Protein Hydrolyzate. Appl. Microbiol. 1961, 9, 175–180. [Google Scholar] [CrossRef]
- Chen, G.J.; Russell, J.B. Effect of Monensin and a Protonophore on Protein Degradation, Peptide Accumulation, and Deamination by Mixed Ruminal Microorganisms in Vitro. J. Anim. Sci. 1991, 69, 2196–2203. [Google Scholar] [CrossRef] [PubMed]
- Russell, J.B.; Sniffen, C.J.; Van Soest, P.J. Effect of Carbohydrate Limitation on Degradation and Utilization of Casein by Mixed Rumen Bacteria. J. Dairy Sci. 1983, 66, 763–775. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Approach | Description | Host | Substrates | Ref |
|---|---|---|---|---|
| Gene knockout | Deletion of CodY gene which regulates genes:
| Bacillus subtilis | Amino acid | [53] |
| Gene knockout | Deletion of gene BkdB which helps in the biosynthesis of branched chain fatty acids | Bacillus subtilis | Amino acid | [53] |
| Gene overexpression | Over expression of proteins leuDH, and two-keto-acid decarboxylase which respectively converts amino acids to important metabolic intermediates and increases the availability of metabolic precursors for ammonia production | Bacillus subtilis | Amino acid | [53] |
| Gene knockout | Deletion of genes glnA and gdhA which aids ammonia assimilation | Eschericia coli | Amino acid | [54] |
| Gene knockout | Deletion of ptsG (glucose transporter gene) and deletion of phosphoenol pyruvate (glucose transporter) | Eschericia coli | Soybean residue and food waste | [56] |
| Cell surface engineering | HcLAAO (L-amino acid oxidase) display on yeast cell surface by gene insertion | Yeast cells | Amino acids from soybean residue | [55] |
| Cell surface engineering | Glutaminase gene (Ybas) display on yeast cell surface by gene insertion. | Yeast cells | Soybean residue and glutamine | [59]. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adeniyi, A.; Bello, I.; Mukaila, T.; Sarker, N.C.; Hammed, A. Trends in Biological Ammonia Production. BioTech 2023, 12, 41. https://doi.org/10.3390/biotech12020041
Adeniyi A, Bello I, Mukaila T, Sarker NC, Hammed A. Trends in Biological Ammonia Production. BioTech. 2023; 12(2):41. https://doi.org/10.3390/biotech12020041
Chicago/Turabian StyleAdeniyi, Adewale, Ibrahim Bello, Taofeek Mukaila, Niloy Chandra Sarker, and Ademola Hammed. 2023. "Trends in Biological Ammonia Production" BioTech 12, no. 2: 41. https://doi.org/10.3390/biotech12020041
APA StyleAdeniyi, A., Bello, I., Mukaila, T., Sarker, N. C., & Hammed, A. (2023). Trends in Biological Ammonia Production. BioTech, 12(2), 41. https://doi.org/10.3390/biotech12020041