
Biocatalysts Based on Immobilized Lipases for the Production of Fatty Acid Ethyl Esters: Enhancement of Activity through Ionic Additives and Ion Exchange Supports

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Esterase Activity and Protein Determination
2.3. Immobilization of Lipases: Obtaining Lipase Derivatives
2.4. One-Step Solvent-Free Fatty Acid Ethyl Ester Production (EE)
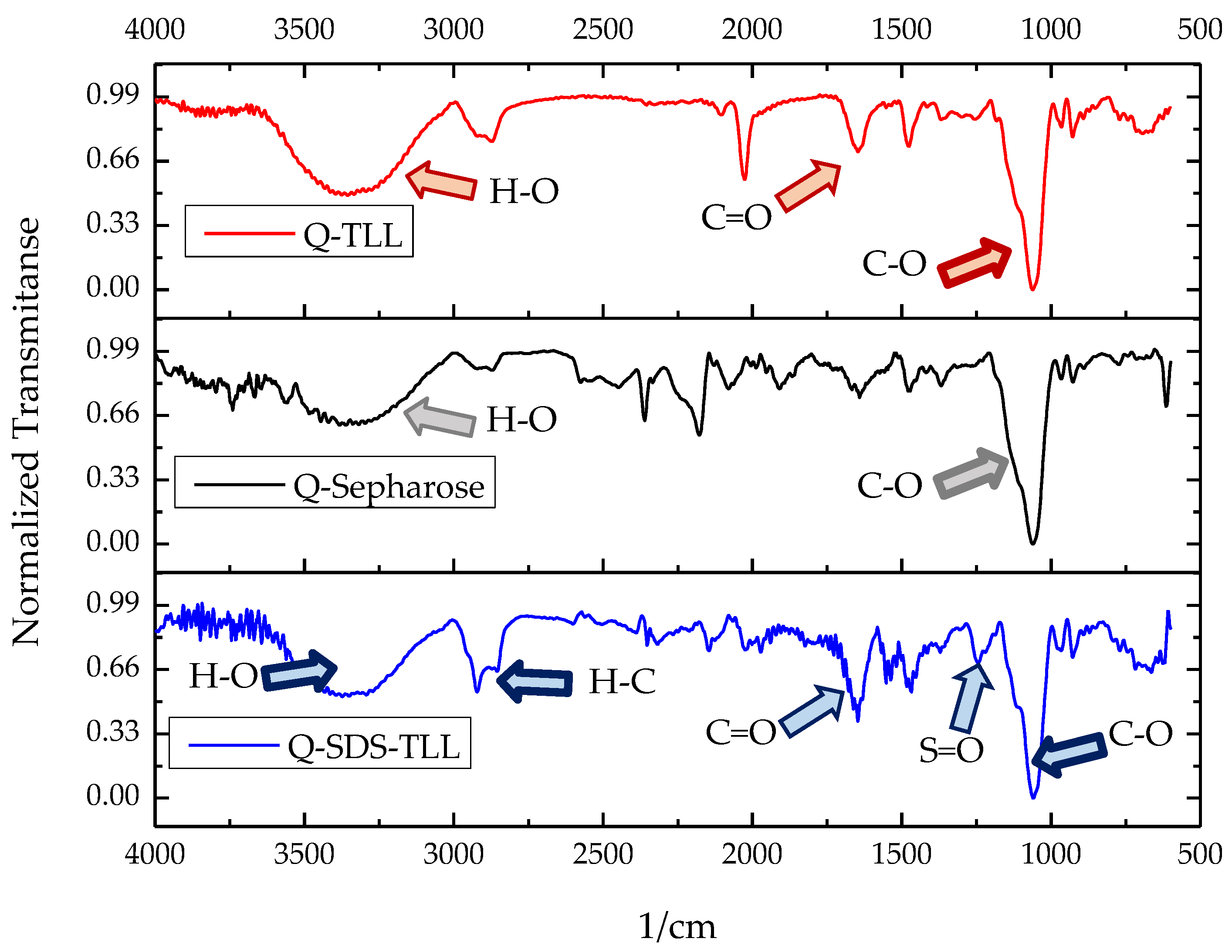
2.5. Spectroscopic Measurements of Derivatives and Supports
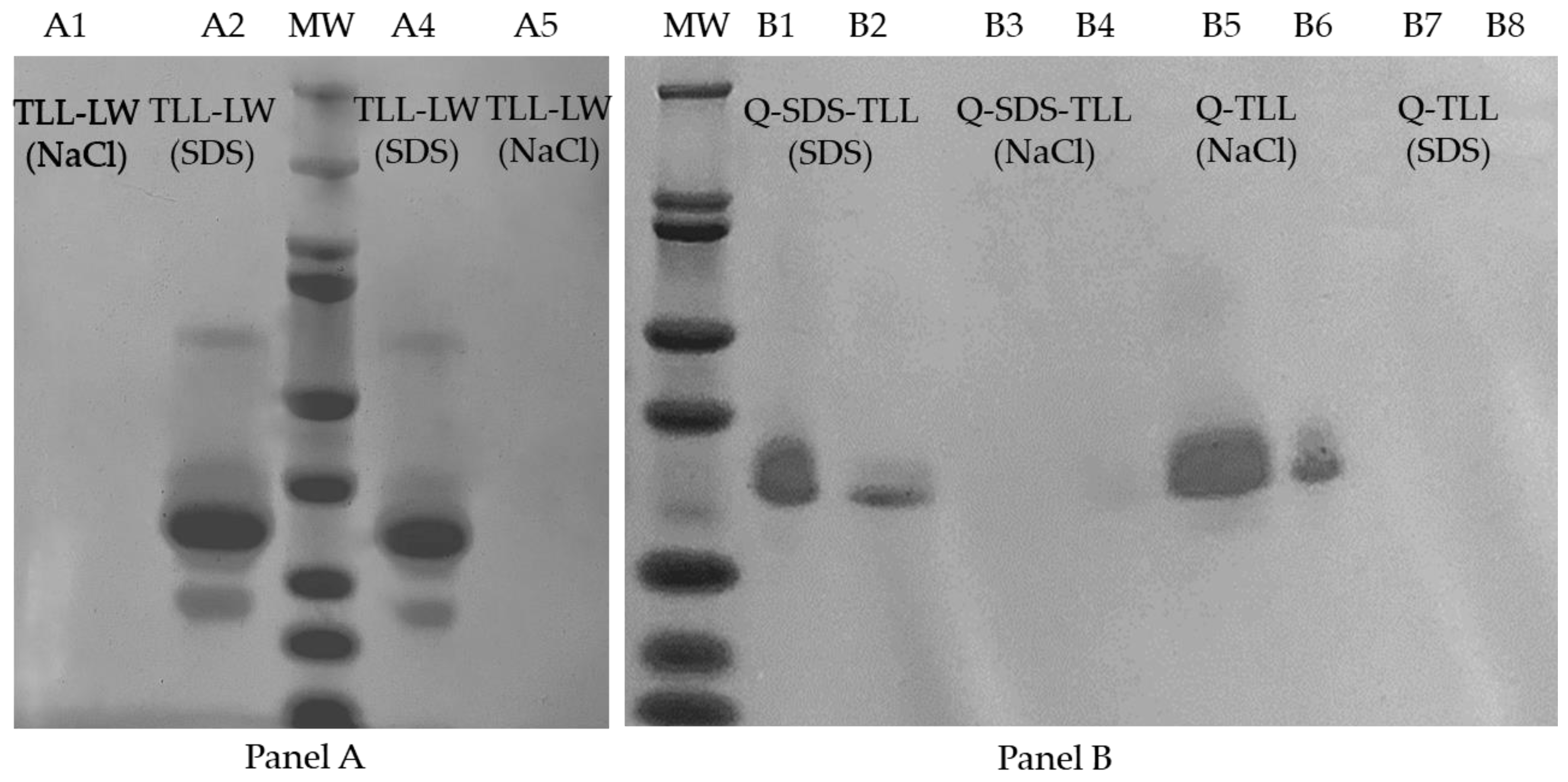
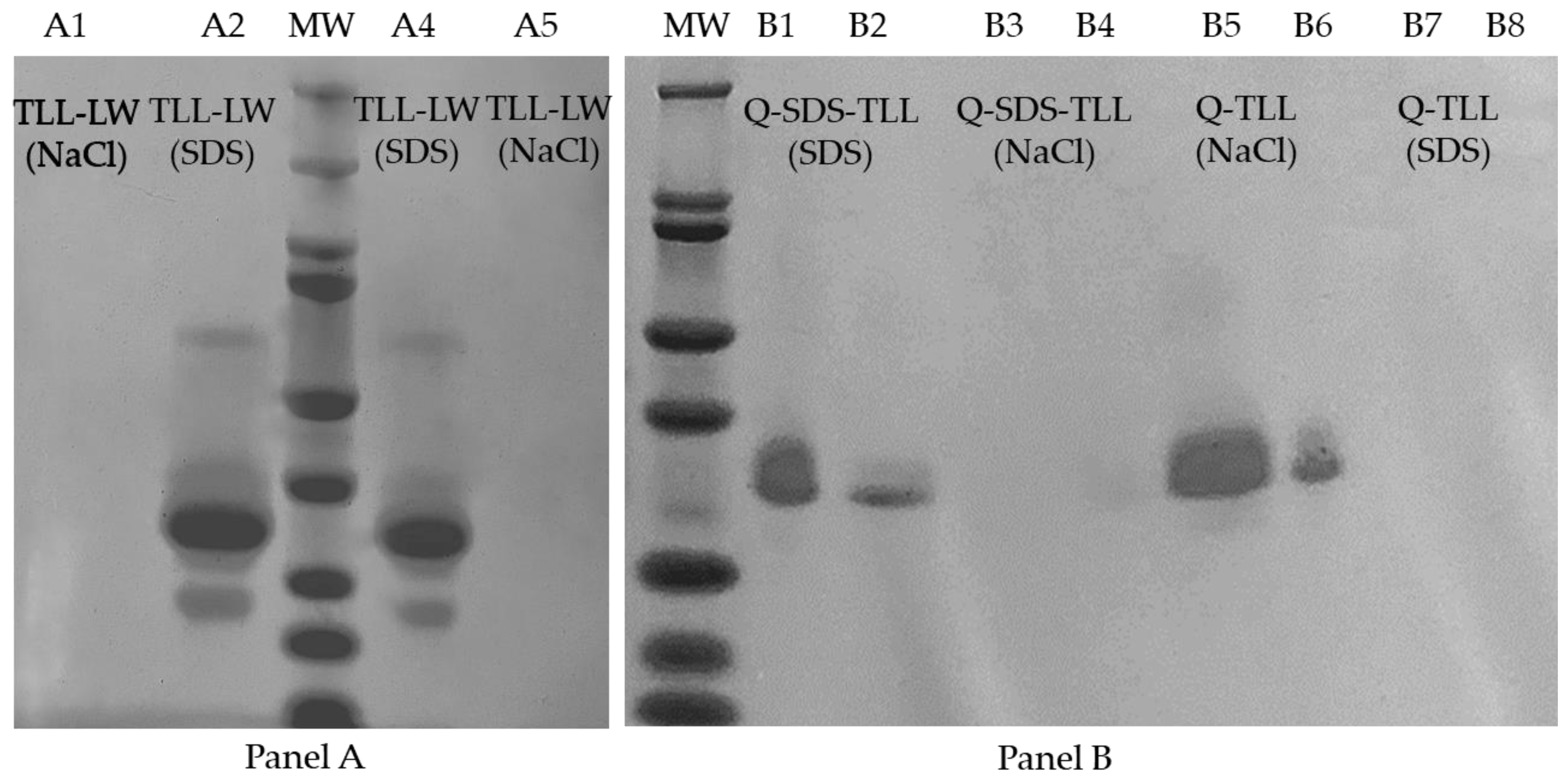
2.6. SDS-PAGE Characterization of the Biocatalysts
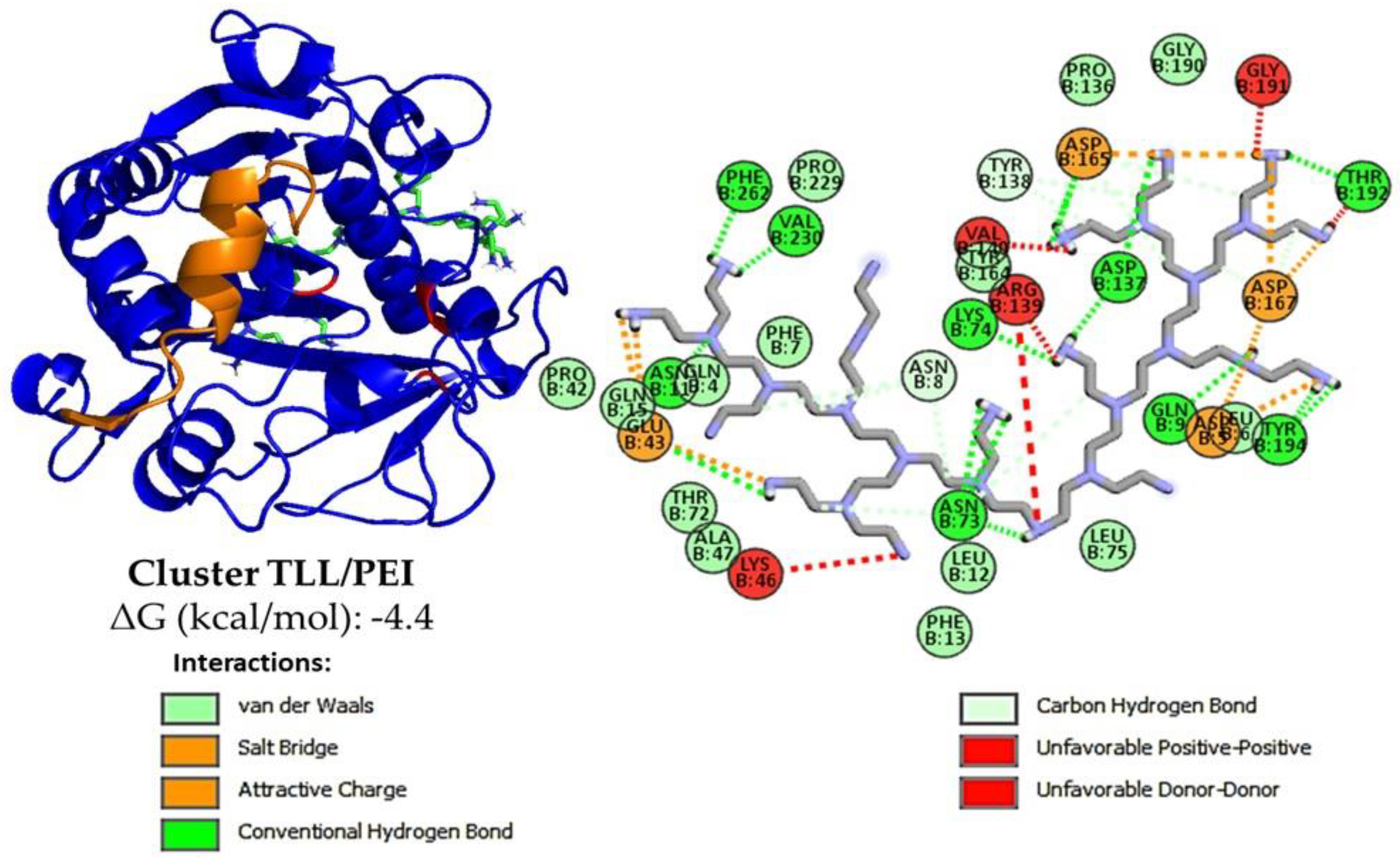
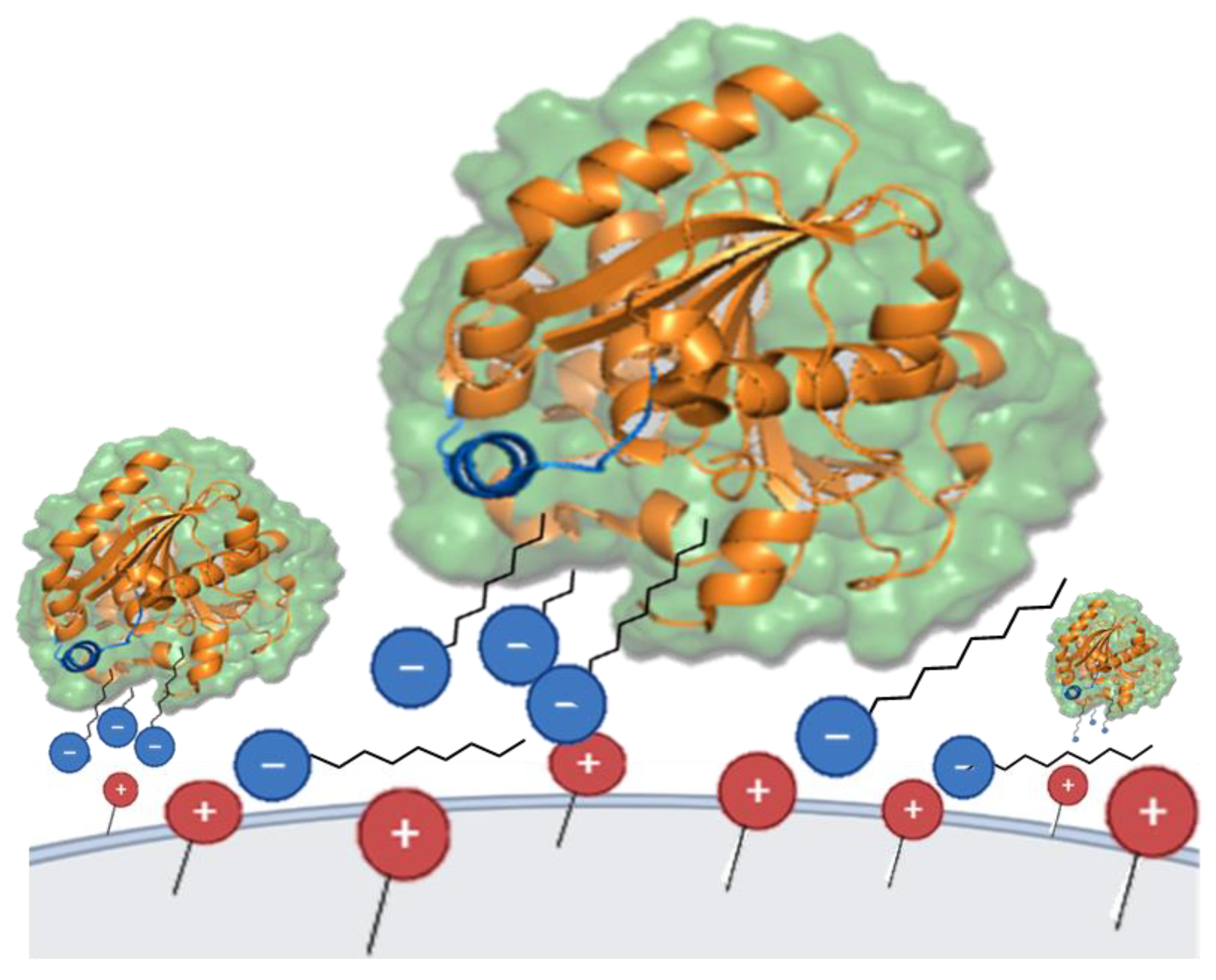
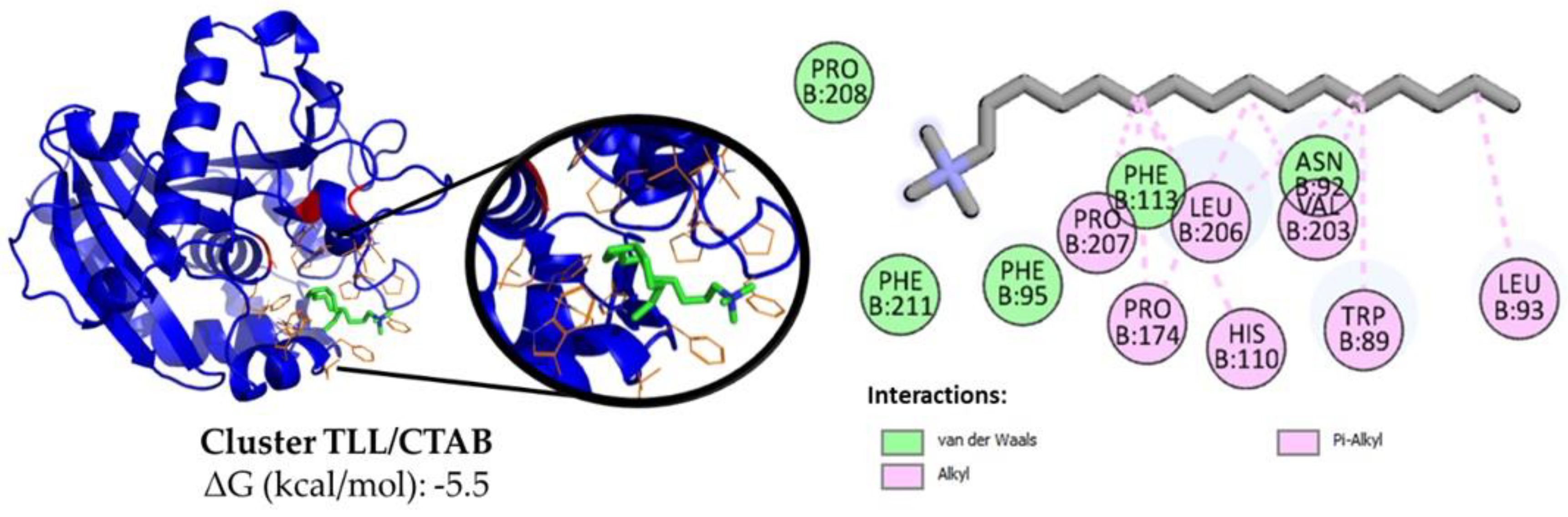
2.7. TLL-Additive In Silico Modeling
2.8. Statistical Analysis
3. Results and Discussion
3.1. Reversible Immobilization of Lipases in Different Types of Commercial Supports
3.1.1. Synthesis of Ethyl Esters of Fatty Acids Using Lipase Derivatives
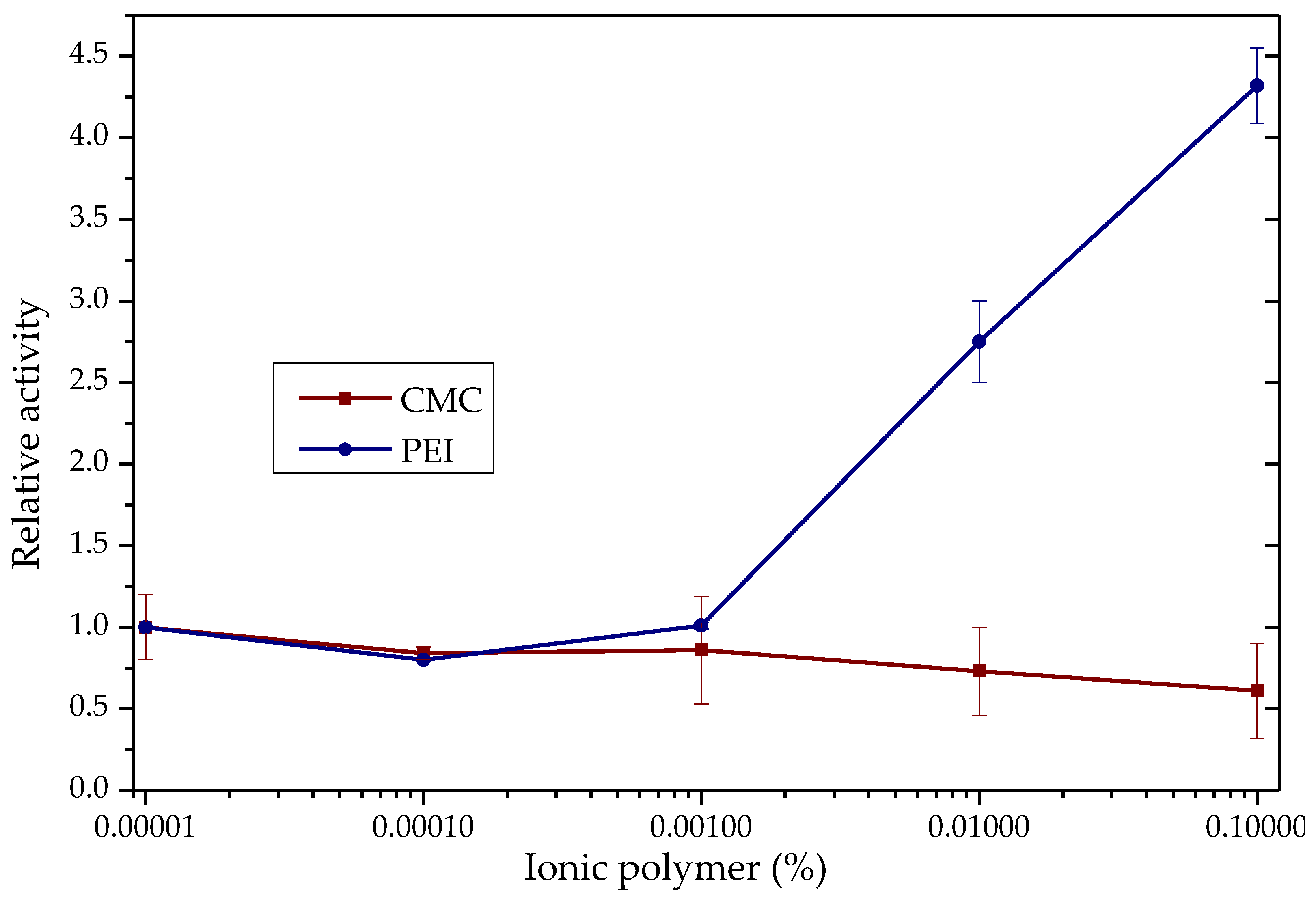
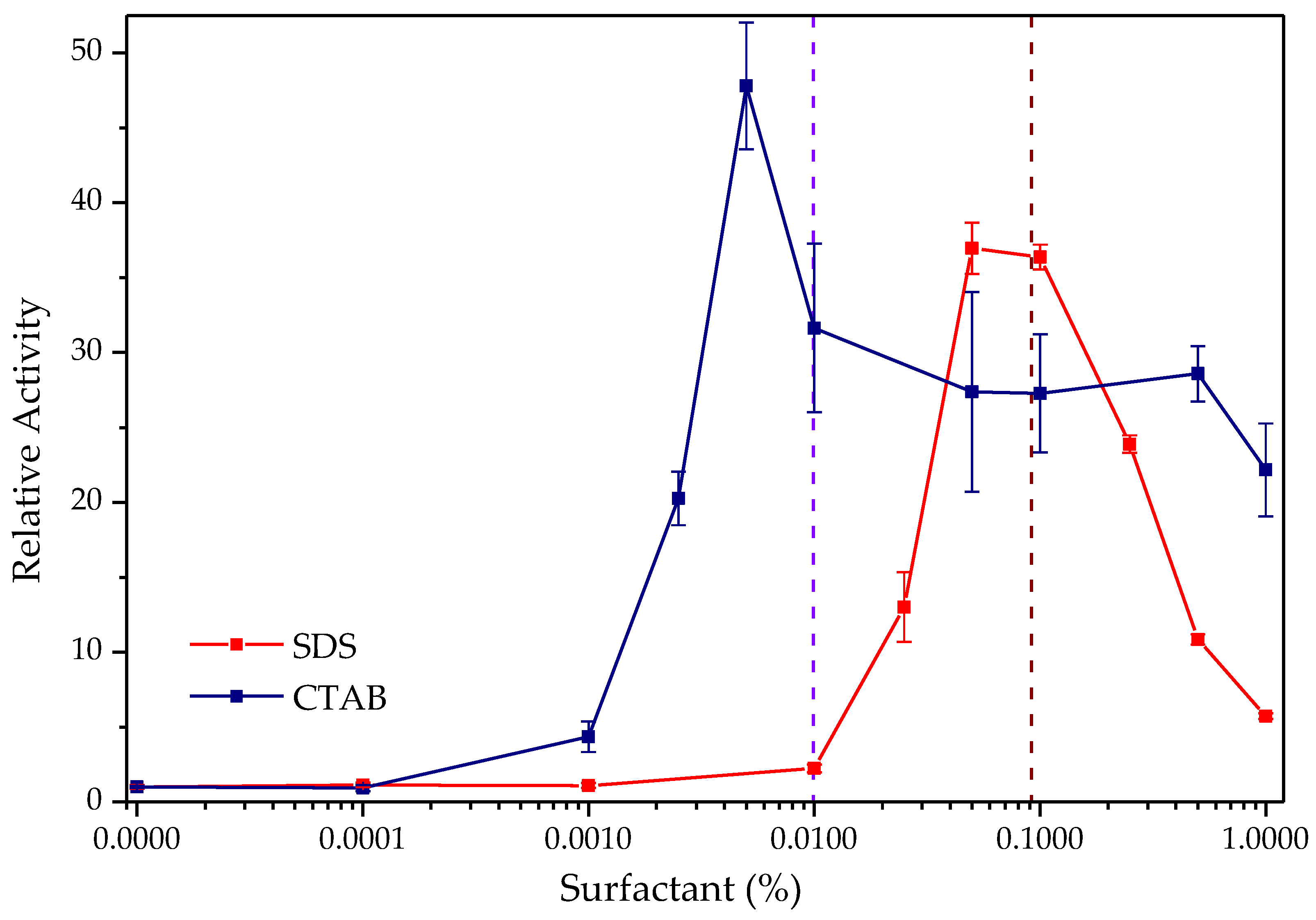
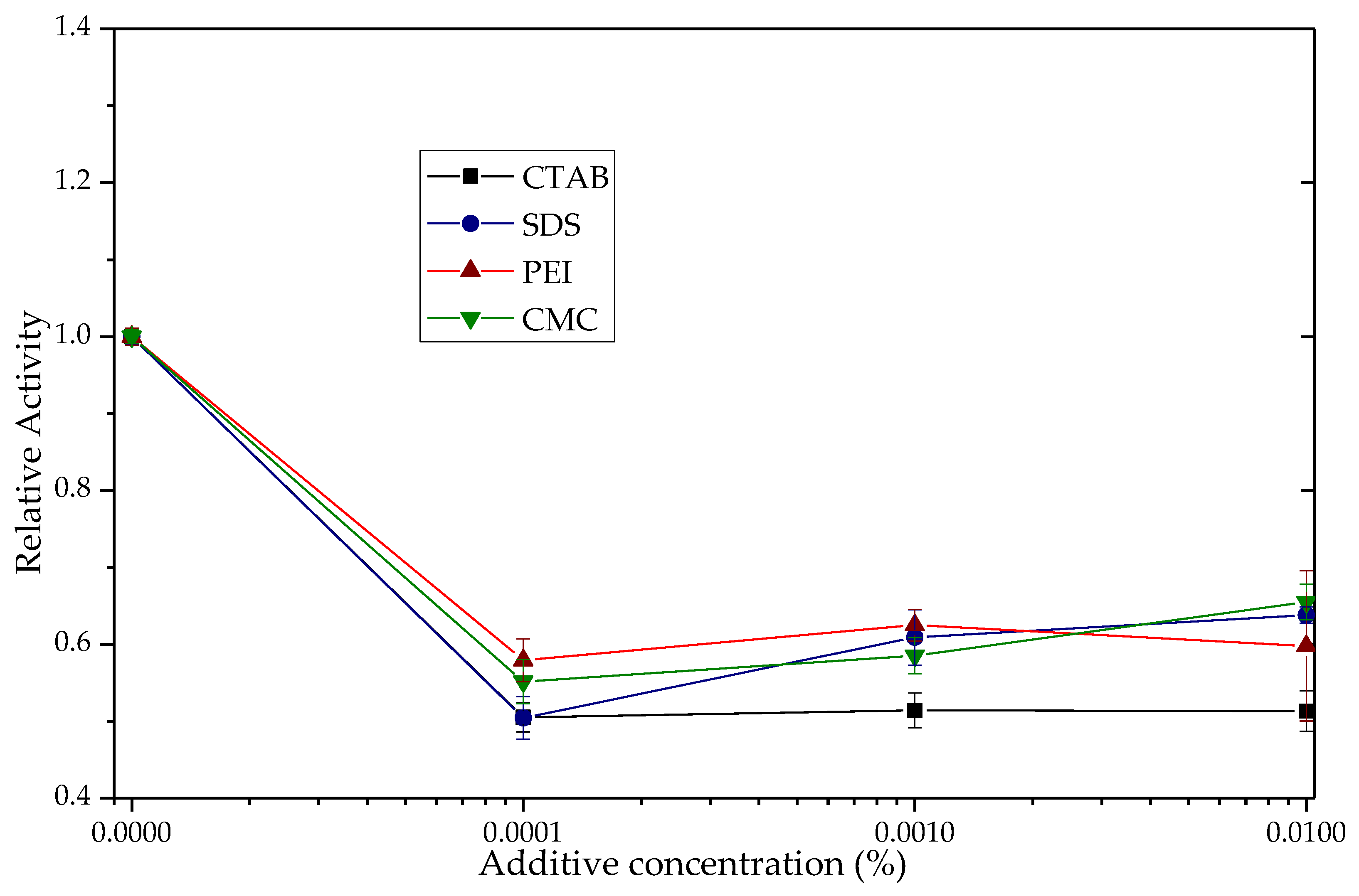
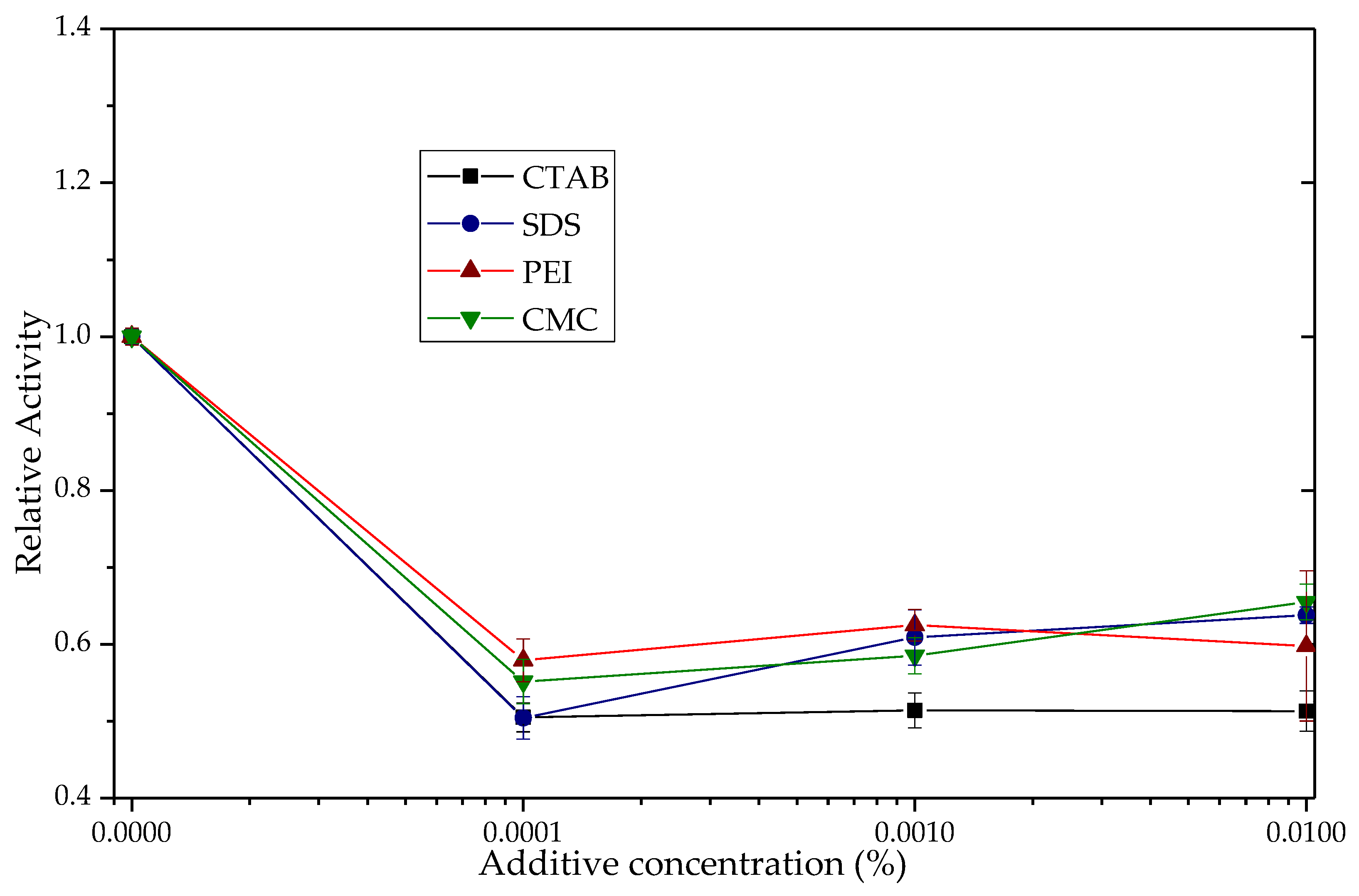
3.2. Effects of the Ionic Additives CTAB, SDS, PEI and CMC on the Hydrolytic Activity of TLL
3.3. TLL Immobilization on Q-Sepharose® Supports Using Ionic Surfactants
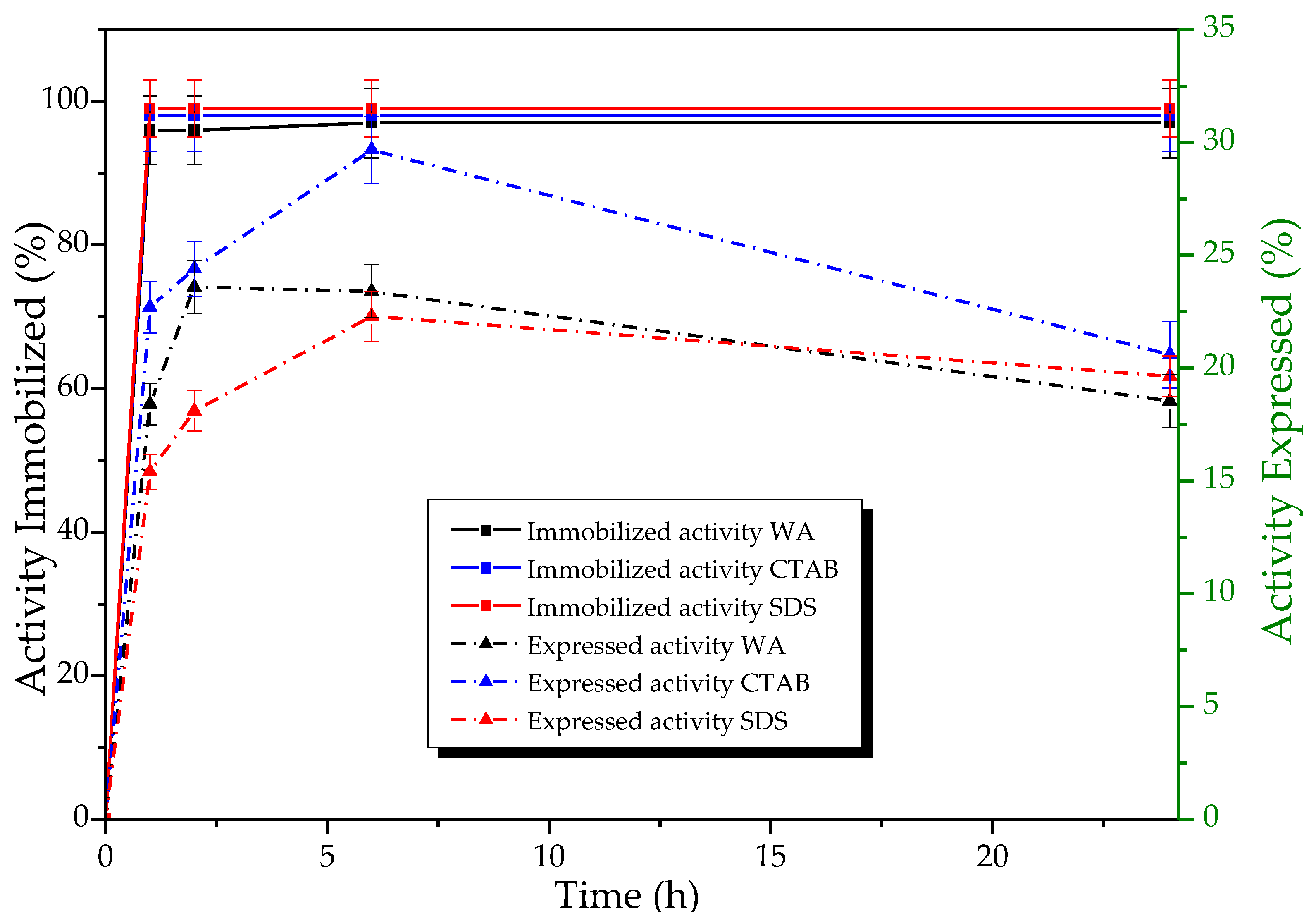
3.3.1. Effect of Ionic Surfactants on the Immobilization Process
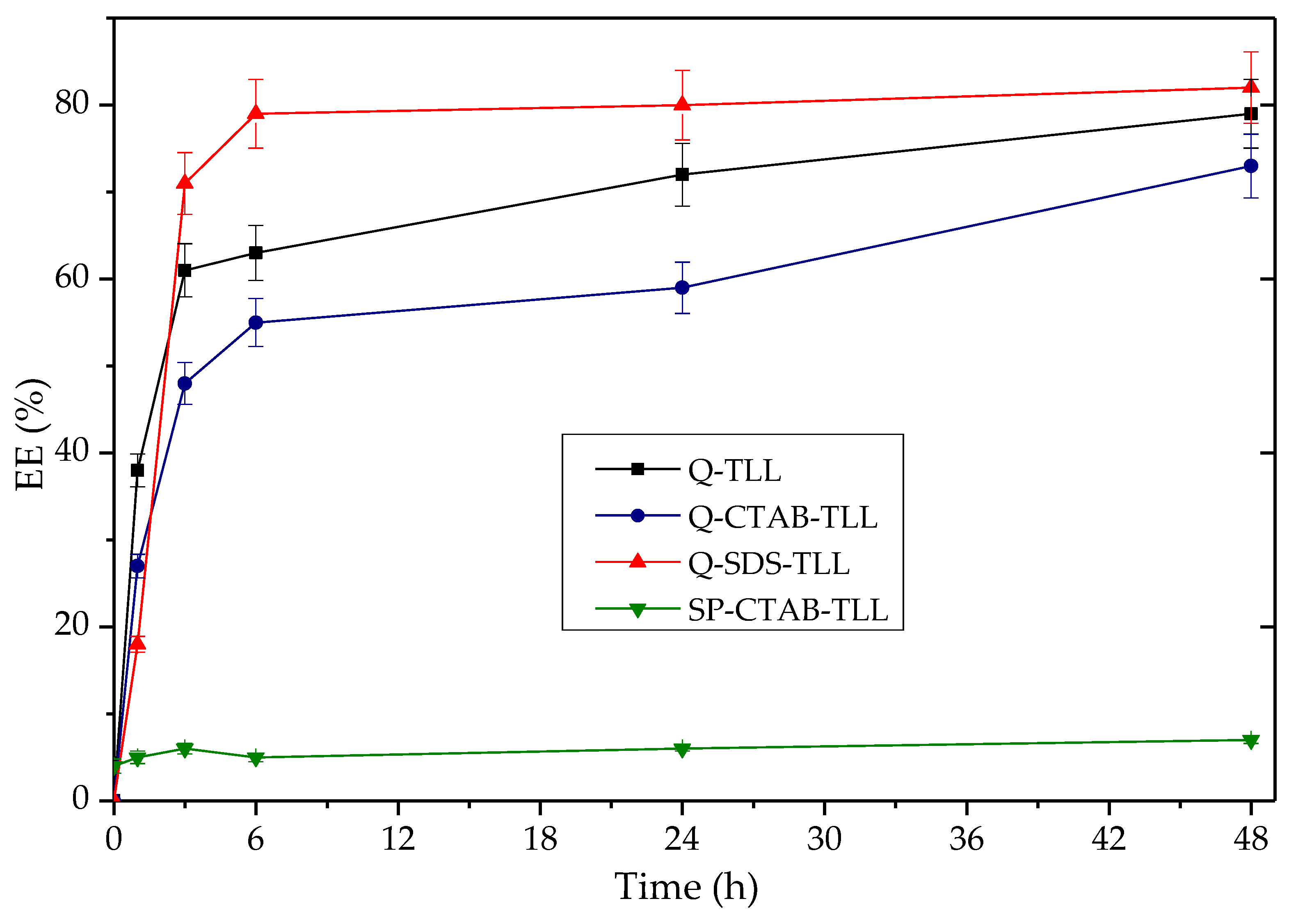
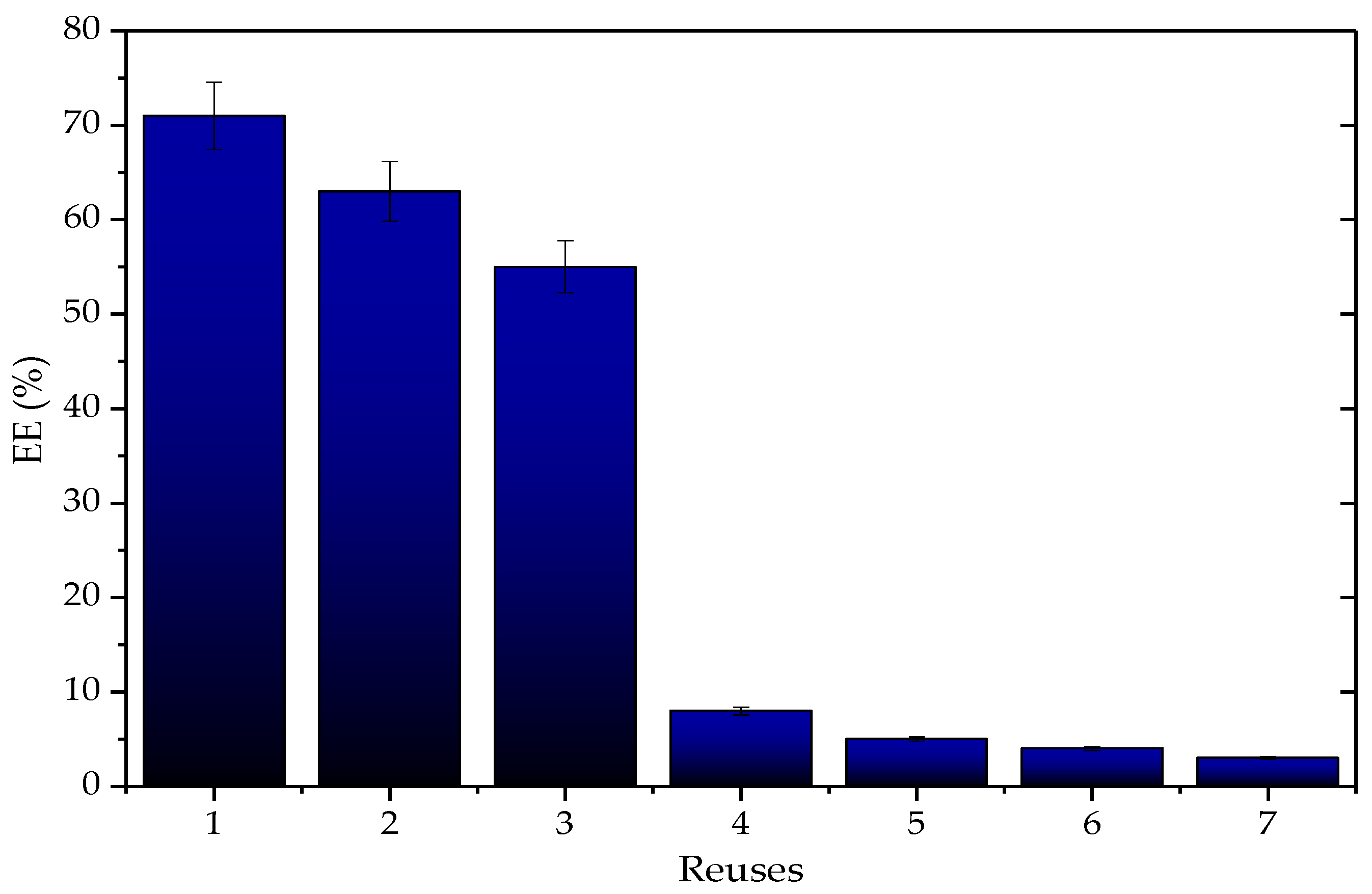
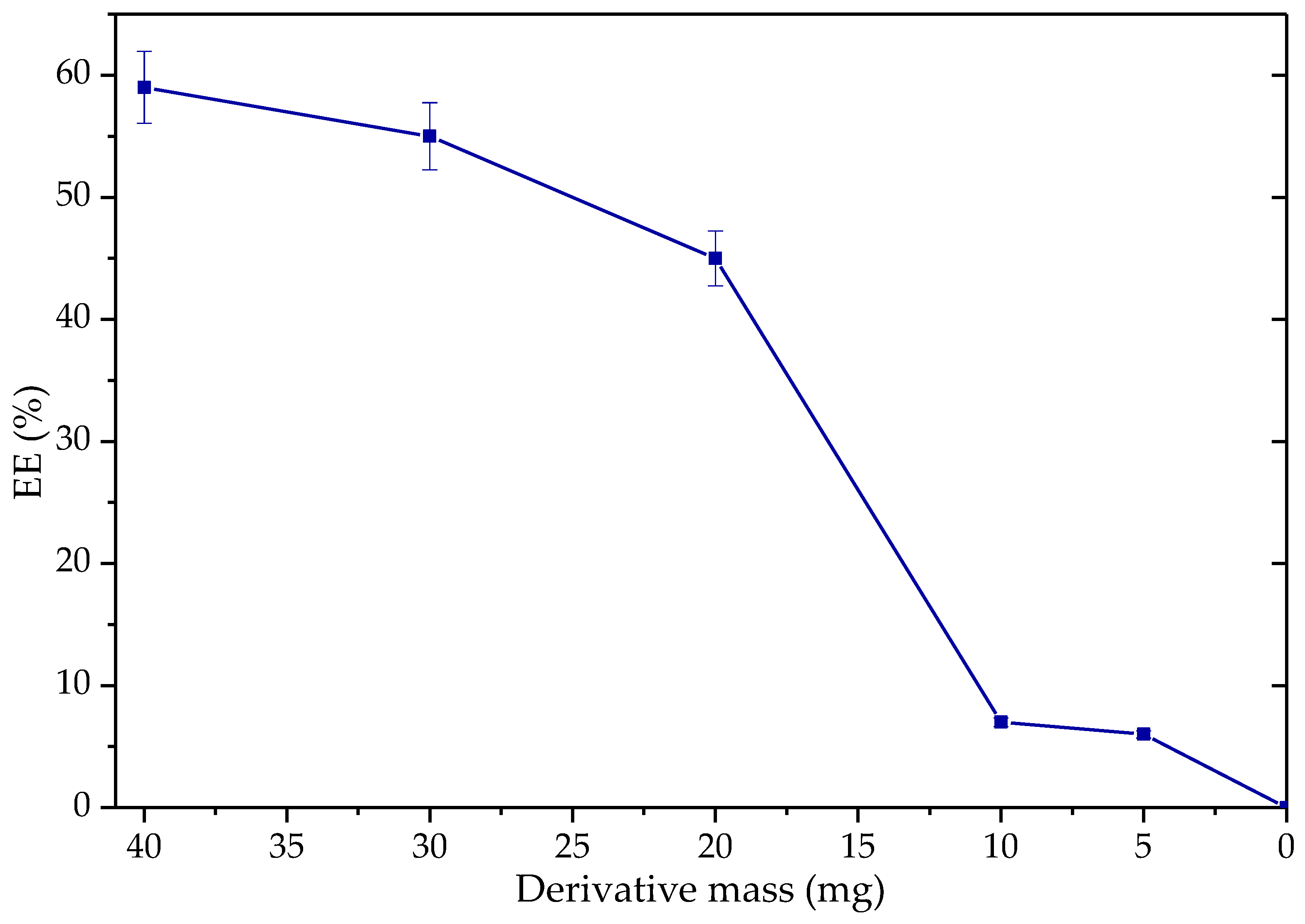
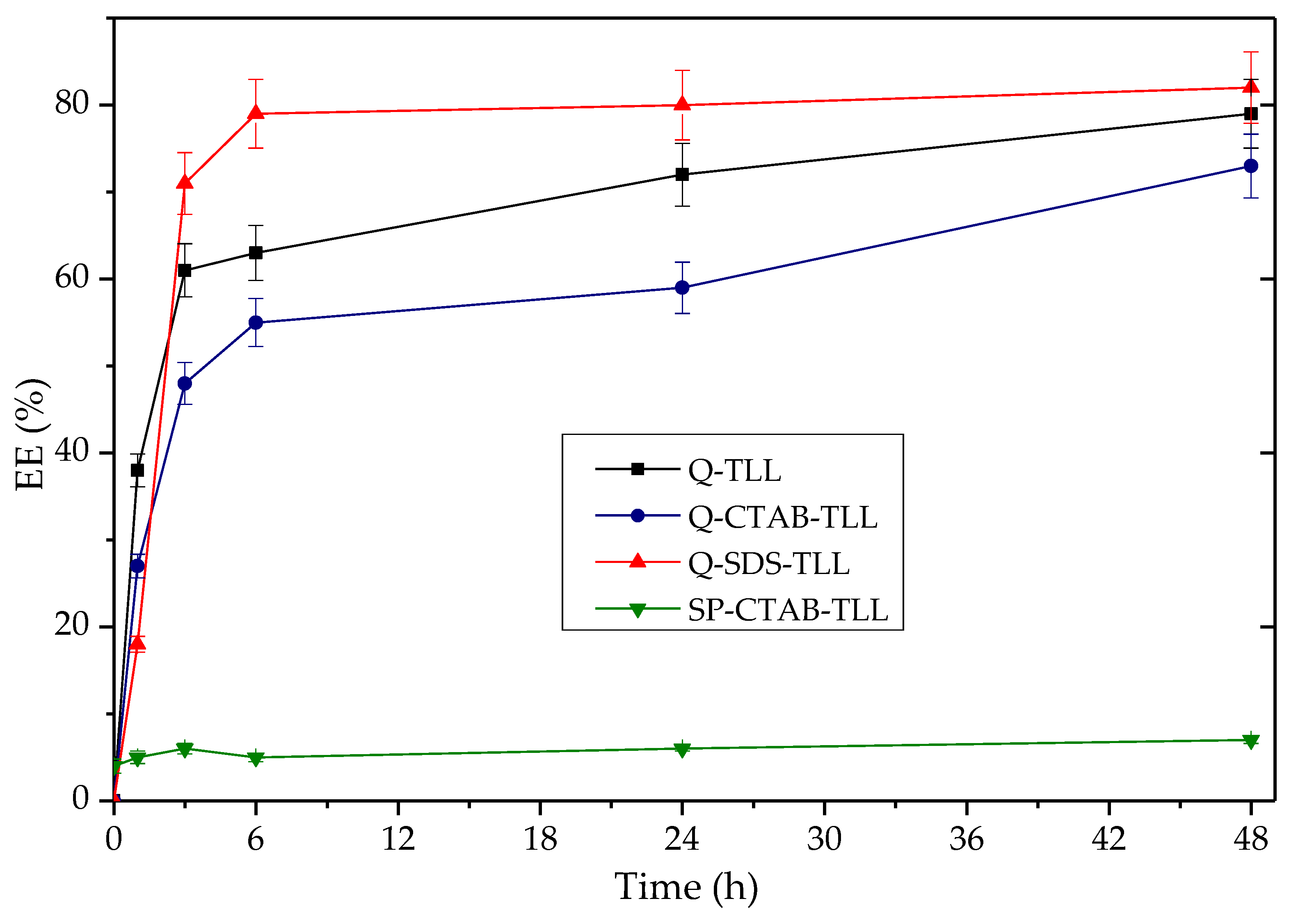
3.3.2. Application of Q-TLL Derivatives in Ethyl Ester Production
3.4. Comparison of EE Production of the Derivative Q-SDS-TLL Versus Lipozyme® TL IM
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BCA | Bicinchoninic acid |
| BSA | Bovine serum albumin |
| CALB | Candida antarctica B |
| CMC | Carboxy-methylcellulose |
| CTAB | Hexadecyltrimethylammonium bromide |
| DexSO4 | Dextran Sulfate agarose (−) |
| EE | Ethyl esters fatty acids |
| MP800 | Lewatit® MP800 |
| MPSP | Lewatit® MPSP112H |
| NK | Nekrolith®(+) |
| OC | Octyl-Sepharose® * |
| PEI | Polyethylenimine |
| PL® | Purolite® ECR1604 |
| p-NPB | p-nitrophenyl butyrate |
| Q | Q-Sepharose® (+) |
| RML | Rhizomucor miehei |
| SDS | Sodium dodecyl sulfate |
| SP | Sulfopropyl Sepharose® (−) |
| TLL | Thermomyces lanuginosus lipase |
| TX | Triton® X-100 |
| VPOC | Lewatit® VPOC1600 |
References
- Sheldon, R.A.; Woodley, J.M. Role of Biocatalysis in Sustainable Chemistry. Chem. Rev. 2018, 118, 801–838. [Google Scholar] [CrossRef] [PubMed]
- Bornscheuer, U.T.; Huisman, G.W.; Kazlauskas, R.J.; Lutz, S.; Moore, J.C.; Robins, K. Engineering the Third Wave of Biocatalysis. Nature 2012, 485, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Insight, T. Expanding Biocatalysis for a Sustainable Future. Nat. Catal. 2020, 3, 179–180. [Google Scholar] [CrossRef]
- Amoah, J.; Ho, S.H.; Hama, S.; Yoshida, A.; Nakanishi, A.; Hasunuma, T.; Ogino, C.; Kondo, A. Lipase Cocktail for Efficient Conversion of Oils Containing Phospholipids to Biodiesel. Bioresour. Technol. 2016, 211, 224–230. [Google Scholar] [CrossRef] [PubMed]
- Grunwald, P. Immobilized Biocatalysts. Catalysts 2018, 8, 386. [Google Scholar] [CrossRef]
- Chen, X.; Li, J.; Deng, L.; Pedersen, J.N.; Li, L.; Guo, Z.; Cong, F.; Xu, X. Chapter 10—Biodiesel Production Using Lipases. In Lipid Modification by Enzymes and Engineered Microbes; Bornscheuer, U.T., Ed.; AOCS Press: Urbana, IL, USA, 2018; pp. 203–238. ISBN 978-0-12-813167-1. [Google Scholar]
- Cavalcante, F.T.T.; Neto, F.S.; Rafael de Aguiar Falcão, I.; Erick da Silva Souza, J.; de Moura Junior, L.S.; da Silva Sousa, P.; Rocha, T.G.; de Sousa, I.G.; de Lima Gomes, P.H.; de Souza, M.C.M.; et al. Opportunities for Improving Biodiesel Production via Lipase Catalysis. Fuel 2021, 288, 119577. [Google Scholar] [CrossRef]
- Lv, L.; Dai, L.; Du, W.; Liu, D. Progress in Enzymatic Biodiesel Production and Commercialization. Processes 2021, 9, 355. [Google Scholar] [CrossRef]
- Toro, E.C.; Rodríguez, D.F.; Morales, N.; García, L.M.; Godoy, C.A. Novel Combi-Lipase Systems for Fatty Acid Ethyl Esters Production. Catalysts 2019, 9, 546. [Google Scholar] [CrossRef]
- Degfie, T.A.; Mamo, T.T.; Mekonnen, Y.S. Optimized Biodiesel Production from Waste Cooking Oil (WCO) Using Calcium Oxide (CaO) Nano-Catalyst. Sci. Rep. 2019, 9, 18982. [Google Scholar] [CrossRef]
- Vela, M.A.F.; Acevedo-Páez, J.C.; Urbina-Suárez, N.; Basto, Y.A.R.; González-Delgado, A.D. Enzymatic Transesterification Ofwaste Frying Oil from Local Restaurants in East Colombia Using a Combined Lipase System. Appl. Sci. 2020, 10, 3566. [Google Scholar] [CrossRef]
- Palacio-Ciro, S.; Vasco-Correa, C.A. Biofuels Policy in Colombia: A Reconfiguration to the Sugar and Palm Sectors? Renew. Sustain. Energy Rev. 2020, 134, 110316. [Google Scholar] [CrossRef]
- Tan, E.C.D.; Lamers, P. Circular Bioeconomy Concepts—A Perspective. Front. Sustain. 2021, 2, 1–8. [Google Scholar] [CrossRef]
- Chandra, P.; Enespa; Singh, R.; Arora, P.K. Microbial Lipases and Their Industrial Applications: A Comprehensive Review. Microb. Cell Fact. 2020, 19, 169. [Google Scholar] [CrossRef]
- Godoy, C.A.; Pardo-Tamayo, J.S.; Barbosa, O. Microbial Lipases and Their Potential in the Production of Pharmaceutical Building Blocks. Int. J. Mol. Sci. 2022, 23, 9933. [Google Scholar] [CrossRef]
- Verma, S.; Meghwanshi, G.K.; Kumar, R. Current Perspectives for Microbial Lipases from Extremophiles and Metagenomics. Biochimie 2021, 182, 23–36. [Google Scholar] [CrossRef] [PubMed]
- Bello, M.; Basilio-Antonio, L.; Fragoso-Vázquez, J.; Avalos-Soriano, A.; Correa-Basurto, J. Molecular Recognition between Pancreatic Lipase and Natural and Synthetic Inhibitors. Int. J. Biol. Macromol. 2017, 98, 855–868. [Google Scholar] [CrossRef] [PubMed]
- Ravindran, R.; Jaiswal, A.K. Microbial Enzyme Production Using Lignocellulosic Food Industry Wastes as Feedstock: A Review. Bioengineering 2016, 3, 30. [Google Scholar] [CrossRef]
- Mangiagalli, M.; Ami, D.; de Divitiis, M.; Brocca, S.; Catelani, T.; Natalello, A.; Lotti, M. Short-Chain Alcohols Inactivate an Immobilized Industrial Lipase through Two Different Mechanisms. Biotechnol. J. 2022, 17, 2100712. [Google Scholar] [CrossRef] [PubMed]
- José, C.; Briand, L.E. Deactivation of Novozym® 435 during the Esterification of Ibuprofen with Ethanol: Evidences of the Detrimental Effect of the Alcohol. React. Kinet. Mech. Catal. 2010, 99, 17–22. [Google Scholar] [CrossRef]
- Koutinas, M.; Yiangou, C.; Osório, N.M.; Ioannou, K.; Canet, A.; Valero, F.; Ferreira-Dias, S. Application of Commercial and Non-Commercial Immobilized Lipases for Biocatalytic Production of Ethyl Lactate in Organic Solvents. Bioresour. Technol. 2018, 247, 496–503. [Google Scholar] [CrossRef]
- Khan, F.I.; Lan, D.; Durrani, R.; Huan, W.; Zhao, Z.; Wang, Y. The Lid Domain in Lipases: Structural and Functional Determinant of Enzymatic Properties. Front. Bioeng. Biotechnol. 2017, 5, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Khan, I.M.; He, W.; Li, Y.; Jin, P.; Campanella, O.H.; Zhang, H.; Huo, Y.; Chen, Y.; Yang, H.; et al. Rebuilding the Lid Region from Conformational and Dynamic Features to Engineering Applications of Lipase in Foods: Current Status and Future Prospects. Compr. Rev. Food Sci. Food Saf. 2022, 21, 2688–2714. [Google Scholar] [CrossRef] [PubMed]
- Casas-Godoy, L.; Duquesne, S.; Bordes, F.; Sandoval, G.; Marty, A. Lipases: An Overview. In Lipases and Phospholipases: Methods and Protocols, Methods in Molecular Biology; Sandoval, G., Ed.; Springer Science & Business Media: New York, NY, USA, 2012; pp. 3–30. [Google Scholar]
- Moreno-Perez, S.; Ghattas, N.; Filice, M.; Guisan, J.M.; Fernandez-Lorente, G. Dramatic Hyperactivation of Lipase of Thermomyces Lanuginosa by a Cationic Surfactant: Fixation of the Hyperactivated Form by Adsorption on Sulfopropyl-Sepharose. J. Mol. Catal. B Enzym. 2015, 122, 199–203. [Google Scholar] [CrossRef]
- Gao, Z.; Chu, J.; Jiang, T.; Xu, T.; Wu, B.; He, B. Lipase Immobilization on Functionalized Mesoporous TiO2: Specific Adsorption, Hyperactivation and Application in Cinnamyl Acetate Synthesis. Process Biochem. 2018, 64, 152–159. [Google Scholar] [CrossRef]
- Fernandez-Lorente, G.; Palomo, J.; Cabrera, Z.; Fernandez-Lafuente, R.; Guisán, J. Improved Catalytic Properties of Inmobilized Lipases by the Presence of Very Low Concentrations of Detergents in the Reaction Medium. Biotechnol. Bioeng. 2007, 97, 242–250. [Google Scholar] [CrossRef]
- Adlercreutz, P. Immobilisation and Application of Lipases in Organic Media. Chem. Soc. Rev. 2013, 42, 6406–6436. [Google Scholar] [CrossRef]
- Mogensen, J.E.; Sehgal, P.; Otzen, D.E. Activation, Inhibition, and Destabilization of Thermomyces Lanuginosus Lipase by Detergents. Biochemistry 2005, 44, 1719–1730. [Google Scholar] [CrossRef]
- Jasińska, K.; Zieniuk, B.; Jankiewicz, U.; Fabiszewska, A. Bio-Based Materials versus Synthetic Polymers as a Support in Lipase Immobilization: Impact on Versatile Enzyme Activity. Catalysts 2023, 13, 395. [Google Scholar] [CrossRef]
- Torres, R.; Ortiz, C.; Pessela, B.C.C.; Palomo, J.M.; Mateo, C.; Guisán, J.M.; Fernández-Lafuente, R. Improvement of the Enantioselectivity of Lipase (Fraction B) from Candida Antarctica via Adsorpiton on Polyethylenimine-Agarose under Different Experimental Conditions. Enzym. Microb. Technol. 2006, 39, 167–171. [Google Scholar] [CrossRef]
- Fuentes, M.; Maquiese, J.V.; Pessela, B.C.C.; Torres, R.; Grazú, V.; Fernández-Lafuente, R.; Guisán, J.M.; Mateo, C. Use of Polyvalent Cations to Improve the Adsorption Strength between Adsorbed Enzymes and Supports Coated with Dextran Sulfate. Enzym. Microb. Technol. 2006, 39, 332–336. [Google Scholar] [CrossRef]
- Fernandez-Lorente, G.; Cabrera, Z.; Godoy, C.; Fernandez-Lafuente, R.; Palomo, J.M.; Guisan, J.M. Interfacially Activated Lipases against Hydrophobic Supports: Effect of the Support Nature on the Biocatalytic Properties. Process Biochem. 2008, 43, 1061–1067. [Google Scholar] [CrossRef]
- Godoy, C.A. New Strategy for the Immobilization of Lipases on Glyoxyl–Agarose Supports: Production of Robust Biocatalysts for Natural Oil Transformation. Int. J. Mol. Sci. 2017, 18, 2130. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, A.; Cruz, J.; Rueda, N.; Santos, J.C.S.d.; Torres, R.; Ortiz, C.; Villalonga, R.; Fernandez-Lafuente, R. Inactivation of Immobilized Trypsin under Dissimilar Conditions Produces Trypsin Molecules with Different Structures. RSC Adv. 2016, 6, 27329–27334. [Google Scholar] [CrossRef]
- Abellanas-Perez, P.; Carballares, D.; Fernandez-Lafuente, R.; Rocha-Martin, J. Glutaraldehyde Modification of Lipases Immobilized on Octyl Agarose Beads: Roles of the Support Enzyme Loading and Chemical Amination of the Enzyme on the Final Enzyme Features. Int. J. Biol. Macromol. 2023, 248, 125853. [Google Scholar] [CrossRef]
- Dallakyan, S.; Olson, A. Small-Molecule Library Screening by Docking with PyRX. In Chemical Biology: Mehtods and Protocols, Methods in Molecular Biology; Hempel, J., Ed.; Springer Science & Business Media: New York, NY, USA, 2015; pp. 243–250. ISBN 978-1-4939-2268-0. [Google Scholar]
- Zaak, H.; Fernandez-Lopez, L.; Otero, C.; Sassi, M.; Fernandez-Lafuente, R. Improved Stability of Immobilized Lipases via Modification with Polyethylenimine and Glutaraldehyde. Enzym. Microb. Technol. 2017, 106, 67–74. [Google Scholar] [CrossRef]
- Palomo, J.M.; Muñoz, G.; Fernández-Lorente, G.; Mateo, C.; Fernández-Lafuente, R.; Guisán, J.M. Interfacial Adsorption of Lipases on Very Hydrophobic Support (Octadecyl–Sepabeads): Immobilization, Hyperactivation and Stabilization of the Open Form of Lipases. J. Mol. Catal. B Enzym. 2002, 19–20, 279–286. [Google Scholar] [CrossRef]
- Scordino, M.; Mauro, A.D.; Passerini, A.; Maccarone, E. Highly Purified Sugar Concentrate from a Residue of Citrus Pigments Recovery Process. LWT—Food Sci. Technol. 2007, 40, 713–721. [Google Scholar] [CrossRef]
- Skjold-Jørgensen, J.; Vind, J.; Svendsen, A.; Bjerrum, M.J. Lipases That Activate at High Solvent Polarities. Biochemistry 2016, 55, 146–156. [Google Scholar] [CrossRef]
- Rodrigues, R.C.; Virgen-Ortíz, J.J.; dos Santos, J.C.S.; Berenguer-Murcia, Á.; Alcantara, A.R.; Barbosa, O.; Ortiz, C.; Fernandez-Lafuente, R. Immobilization of Lipases on Hydrophobic Supports: Immobilization Mechanism, Advantages, Problems, and Solutions. Biotechnol. Adv. 2019, 37, 746–770. [Google Scholar] [CrossRef]
- Zhao, H. Chapter 4—Lipase Activation and Stability Enhancement in Ionic Liquids. In Ionic Liquids in Lipid Processing and Analysis; Xu, X., Guo, Z., Cheong, L.-Z., Eds.; AOCS Press: Urbana, IL, USA, 2016; pp. 99–152. ISBN 978-1-63067-047-4. [Google Scholar]
- Manoel, E.A.; dos Santos, J.C.S.; Freire, D.M.G.; Rueda, N.; Fernandez-Lafuente, R. Immobilization of Lipases on Hydrophobic Supports Involves the Open Form of the Enzyme. Enzym. Microb. Technol. 2015, 71, 53–57. [Google Scholar] [CrossRef]
- Barbosa, O.; Ortiz, C.; Berenguer-Murcia, Á.; Torres, R.; Rodrigues, R.C.; Fernandez-Lafuente, R. Strategies for the One-Step Immobilization–Purification of Enzymes as Industrial Biocatalysts. Biotechnol. Adv. 2015, 33, 435–456. [Google Scholar] [CrossRef] [PubMed]
- Bi, Y.; Wang, Z.; Zhang, R.; Diao, Y.; Tian, Y.; Jin, Z. Improved Catalytic Properties of Thermomyces Lanuginosus Lipase Immobilized onto Newly Fabricated Polydopamine-Functionalized Magnetic Fe3O4 Nanoparticles. Processes 2020, 8, 629. [Google Scholar] [CrossRef]
- Yu, D.; Zhang, X.; Zou, D.; Wang, T.; Liu, T.; Wang, L.; Elfalleh, W.; Jiang, L. Immobilized CALB Catalyzed Transesterification of Soybean Oil and Phytosterol. Food Biophys. 2018, 13, 208–215. [Google Scholar] [CrossRef]
- Rauwerdink, A.; Kazlauskas, R.J. How the Same Core Catalytic Machinery Catalyzes 17 Different Reactions: The Serine-Histidine-Aspartate Catalytic Triad of α/β-Hydrolase Fold Enzymes. ACS Catal. 2015, 5, 6153–6176. [Google Scholar] [CrossRef]
- Ortiz, C.; Ferreira, M.L.; Barbosa, O.; dos Santos, J.C.; Rodrigues, R.C.; Berenguer-Murcia, Á.; Briand, L.E.; Fernandez-Lafuente, R. Novozym 435: The “Perfect” Lipase Immobilized Biocatalyst? Catal. Sci. Technol. 2019, 9, 2380–2420. [Google Scholar] [CrossRef]
- Coelho, A.L.S.; Orlandelli, R.C. Immobilized Microbial Lipases in the Food Industry: A Systematic Literature Review. Crit. Rev. Food Sci. Nutr. 2021, 61, 1689–1703. [Google Scholar] [CrossRef]
- Khor, G.K.; Sim, J.H.; Kamaruddin, A.H.; Uzir, M.H. Thermodynamics and Inhibition Studies of Lipozyme TL IM in Biodiesel Production via Enzymatic Transesterification. Bioresour. Technol. 2010, 101, 6558–6561. [Google Scholar] [CrossRef]
- Subhedar, P.B.; Botelho, C.; Ribeiro, A.; Castro, R.; Pereira, M.A.; Gogate, P.R.; Cavaco-Paulo, A. Ultrasound Intensification Suppresses the Need of Methanol Excess during the Biodiesel Production with Lipozyme TL-IM. Ultrason. Sonochemistry 2015, 27, 530–535. [Google Scholar] [CrossRef]
- Arana-Peña, S.; Carballares, D.; Cortés Corberan, V.; Fernandez-Lafuente, R. Multi-Combilipases: Co-Immobilizing Lipases with Very Different Stabilities Combining Immobilization via Interfacial Activation and Ion Exchange. The Reuse of the Most Stable Co-Immobilized Enzymes after Inactivation of the Least Stable Ones. Catalysts 2020, 10, 1207. [Google Scholar] [CrossRef]
- Elgharbawy, A.A.; Riyadi, F.A.; Alam, M.Z.; Moniruzzaman, M. Ionic Liquids as a Potential Solvent for Lipase-Catalysed Reactions: A Review. J. Mol. Liq. 2018, 251, 150–166. [Google Scholar] [CrossRef]
- Vesoloski, J.F.; Todero, A.S.; Macieski, R.J.; de Oliveira Pereira, F.; Dallago, R.M.; Mignoni, M.L. Immobilization of Lipase from Candida Antarctica B (CALB) by Sol–Gel Technique Using Rice Husk Ash as Silic Source and Ionic Liquid as Additive. Appl. Biochem. Biotechnol. 2022, 194, 6270–6286. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.-P.; Xia, Y.-P.; Gu, X.-F.; Han, L.; Chen, Q.; Zhi, G.-Y.; Zhang, D.-H. PEI-Crosslinked Lipase on the Surface of Magnetic Microspheres and Its Characteristics. Colloids Surf. B Biointerfaces 2020, 189, 110874. [Google Scholar] [CrossRef] [PubMed]
- Najim, A.A.; Ismail, Z.Z.; Hummadi, K.K. Biodegradation Potential of Sodium Dodecyl Sulphate (SDS) by Mixed Cells in Domestic and Non-Domestic Actual Wastewaters: Experimental and Kinetic Studies. Biochem. Eng. J. 2022, 180, 108374. [Google Scholar] [CrossRef]
- Wang, H.; Andersen, K.K.; Sehgal, P.; Hagedorn, J.; Westh, P.; Borch, K.; Otzen, D.E. pH Regulation of the Kinetic Stability of the Lipase from Thermomyces Lanuginosus. Biochemistry 2013, 52, 264–276. [Google Scholar] [CrossRef]
- Willems, N.; Lelimousin, M.; Skjold-Jørgensen, J.; Svendsen, A.; Sansom, M.S.P. The Effect of Mutations in the Lid Region of Thermomyces Lanuginosus Lipase on Interactions with Triglyceride Surfaces: A Multi-Scale Simulation Study. Chem. Phys. Lipids 2018, 211, 4–15. [Google Scholar] [CrossRef]
- Cavalcante, A.L.G.; Chaves, A.V.; Fechine, P.B.A.; Alexandre, J.Y.N.H.; Freire, T.M.; Davi, D.M.B.; Neto, F.S.; Sousa, I.G.d.; Moreira, K.d.S.; Oliveira, A.L.B.d.; et al. Chemical Modification of Clay Nanocomposites for the Improvement of the Catalytic Properties of Lipase A from Candida Antarctica. Process Biochem. 2022, 120, 1–14. [Google Scholar] [CrossRef]
- mgl-admin. Manual. AutoDock Vina. Available online: https://vina.scripps.edu/manual/ (accessed on 12 April 2023).
- Rasmussen, H.Ø.; Wollenberg, D.T.W.; Wang, H.; Andersen, K.K.; Oliveira, C.L.P.; Jørgensen, C.I.; Jørgensen, T.J.D.; Otzen, D.E.; Pedersen, J.S. The Changing Face of SDS Denaturation: Complexes of Thermomyces Lanuginosus Lipase with SDS at pH 4.0, 6.0 and 8.0. J. Colloid Interface Sci. 2022, 614, 214–232. [Google Scholar] [CrossRef]
- Tah, B.; Pal, P.; Mahato, M.; Talapatra, G.B. Aggregation Behavior of SDS/CTAB Catanionic Surfactant Mixture in Aqueous Solution and at the Air/Water Interface. J. Phys. Chem. B 2011, 115, 8493–8499. [Google Scholar] [CrossRef]
- Fuguet, E.; Ràfols, C.; Rosés, M.; Bosch, E. Critical Micelle Concentration of Surfactants in Aqueous Buffered and Unbuffered Systems. Anal. Chim. Acta 2005, 548, 95–100. [Google Scholar] [CrossRef]
- Mukherjee, A.K. Hydrophobic-Hydrophilic Interaction in Lipase Catalytic Triad and Possibility of a Cofactor Mediated Catalysis. Int. J. Agric. Food Sci. 2014, 4, 84–89. [Google Scholar]
- Mateos-Diaz, E.; Amara, S.; Roussel, A.; Longhi, S.; Cambillau, C.; Carrière, F. Probing Conformational Changes and Interfacial Recognition Site of Lipases with Surfactants and Inhibitors, 1st ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2017; Volume 583. [Google Scholar]
- Santini, S.; Crowet, J.M.; Thomas, A.; Paquot, M.; Vandenbol, M.; Thonart, P.; Wathelet, J.P.; Blecker, C.; Lognay, G.; Brasseur, R.; et al. Study of Thermomyces Lanuginosa Lipase in the Presence of Tributyrylglycerol and Water. Biophys. J. 2009, 96, 4814–4825. [Google Scholar] [CrossRef]
- Yilmaz, A. The Portevin–Le Chatelier Effect: A Review of Experimental Findings. Sci. Technol. Adv. Mater. 2011, 12, 063001. [Google Scholar] [CrossRef] [PubMed]
- Banasaz, S.; Morozova, K.; Ferrentino, G.; Scampicchio, M. Encapsulation of Lipid-Soluble Bioactives by Nanoemulsions. Molecules 2020, 25, 3966. [Google Scholar] [CrossRef] [PubMed]
- Goswami, D. Lipase Catalysis in Presence of Nonionic Surfactants. Appl. Biochem. Biotechnol. 2020, 191, 744–762. [Google Scholar] [CrossRef] [PubMed]
- Adriano, W.S.; Mendonça, D.B.; Rodrigues, D.S.; Mammarella, E.J.; Giordano, R.L.C. Improving the Properties of Chitosan as Support for the Covalent Multipoint Immobilization of Chymotrypsin. Biomacromolecules 2008, 9, 2170–2179. [Google Scholar] [CrossRef]
- Carvalho de Melo, J.J.; Passos da Silva, G.L.; Mota, D.A.; de Souza Brandão, L.M.; de Souza, R.L.; Pereira, M.M.; Lima, Á.S.; Soares, C.M.F. Use of Bioprinted Lipases in Microwave-Assisted Esterification Reactions. Catalysts 2023, 13, 299. [Google Scholar] [CrossRef]
- Rivero Berti, I.; Rodenak-Kladniew, B.E.; Katz, S.F.; Arrua, E.C.; Alvarez, V.A.; Duran, N.; Castro, G.R. Enzymatic Active Release of Violacein Present in Nanostructured Lipid Carrier by Lipase Encapsulated in 3D-Bioprinted Chitosan-Hydroxypropyl Methylcellulose Matrix With Anticancer Activity. Front. Chem. 2022, 10, 1–13. [Google Scholar] [CrossRef]
- Thongchai, W. Ion-Pairing Liquid Chromatographic Method Determination of Ketoconazole and Stress Degradation in Pharmaceutical Formulations with Fluorescence Detection. Braz. J. Pharm. Sci. 2020, 56, 1–11. [Google Scholar] [CrossRef]
- Fernandez-Lorente, G.; Rocha-Martín, J.; Guisan, J.M. Immobilization of Lipases by Adsorption on Hydrophobic Supports: Modulation of Enzyme Properties in Biotransformations in Anhydrous Media. In Immobilization of Enzymes and Cells: Methods and Protocols; Guisan, J.M., Bolivar, J.M., López-Gallego, F., Rocha-Martín, J., Eds.; Springer: New York, NY, USA, 2020; pp. 143–158. ISBN 978-1-07-160215-7. [Google Scholar]
- Mateo, C.; Abian, O.; Fernandez-Lafuente, R.; Guisan, J.M. Reversible Enzyme Immobilization via a Very Strong and Nondistorting Ionic Adsorption on Support-Polyethylenimine Composites. Biotechnol. Bioeng. 2000, 68, 98–105. [Google Scholar] [CrossRef]
- McPherson, A.; Larson, B.S.; Kalasky, A. The Crystal Structures of Thermomyces (Humicola) Lanuginosa Lipase in Complex with Enzymatic Reactants. Curr. Enzym. Inhib. 2020, 16, 199–213. [Google Scholar] [CrossRef]
- Yang, H.; Zhang, W. Surfactant Imprinting Hyperactivated Immobilized Lipase as Efficient Biocatalyst for Biodiesel Production from Waste Cooking Oil. Catalysts 2019, 9, 914–926. [Google Scholar] [CrossRef]
- Kahveci, D.; Xu, X. Bioimprinted Immobilization of Candida Antarctica Lipase A for Concentration of Omega-3 Polyunsaturated Fatty Acids. J. Am. Oil Chem. Soc. 2012, 89, 1839–1845. [Google Scholar] [CrossRef]
- Mahbub, S.; Molla, M.R.; Saha, M.; Shahriar, I.; Hoque, M.A.; Halim, M.A.; Rub, M.A.; Khan, M.A.; Azum, N. Conductometric and Molecular Dynamics Studies of the Aggregation Behavior of Sodium Dodecyl Sulfate (SDS) and Cetyltrimethylammonium Bromide (CTAB) in Aqueous and Electrolytes Solution. J. Mol. Liq. 2019, 283, 263–275. [Google Scholar] [CrossRef]
- Sun, M.Y.; Bienkowski, P.R.; Davison, B.H.; Nghiem, N.P.; Webb, O.F. Predictive Model of Co-Immobilized Biocatalyst in Three-Phase Fluidized Bed Reactor; Report Number: CONF-970443-; American Chemical Society: Washington, DC, USA, 1997. [Google Scholar]
- Miložič, N.; Lubej, M.; Lakner, M.; Žnidaršič-Plazl, P.; Plazl, I. Theoretical and Experimental Study of Enzyme Kinetics in a Microreactor System with Surface-Immobilized Biocatalyst. Chem. Eng. J. 2017, 313, 374–381. [Google Scholar] [CrossRef]
- Gupta, J.; Agarwal, M.; Dalai, A.K. An Overview on the Recent Advancements of Sustainable Heterogeneous Catalysts and Prominent Continuous Reactor for Biodiesel Production. J. Ind. Eng. Chem. 2020, 88, 58–77. [Google Scholar] [CrossRef]
- Cesarini, S.; Pastor, F.I.J.; Nielsen, P.M.; Diaz, P. Moving towards a Competitive Fully Enzymatic Biodiesel Process. Sustainability 2015, 7, 7884–7903. [Google Scholar] [CrossRef]
- Suzihaque, M.U.H.; Alwi, H.; Ibrahim, U.K.; Abdullah, S.; Haron, N. Biodiesel Production from Waste Cooking Oil: A Brief Review. Mater. Today Proc. 2022, 63, S490–S495. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Support | Abbr. | Matrix | Enzyme | Immobilization pH | % Activity Immobilized a |
|---|---|---|---|---|---|
| Sulfopropyl Sepharose® (−) | SP | Crosslinked agarose Ligand: Sulfopropyl Type: Anionic, hydrophilic | TLL | 3.5 | 87.9 |
| CALB | 3.5 | 81.3 | |||
| RML | 3.5 | 87.7 | |||
| Lecitase® | 3.5 | 74.3 | |||
| Q-Sepharose® (+) | Q | Crosslinked agarose Ligand: Quaternary amine Type: Cationic, hydrophilic | TLL | 8.0 | 85.9 |
| CALB | 10.0 | 70.6 | |||
| Lecitase® | 8.5 | 41.5 | |||
| Dextran Sulfate agarose (−) | DexSO4 | Crosslinked agarose Ligand: Sulfonic acid Type: Anionic, hydrophilic | TLL | 3.5 | 85.9 |
| CALB | 3.5 | 79.6 | |||
| RML | 3.5 | 69.6 | |||
| Lecitase® | 3.5 | 58.0 | |||
| Octyl-Sepharose® * | OC | Crosslinked agarose Ligand: Octyl groups Type: Hydrophobic, hydrophilic | TLL | 7.0 | 45.0 [38] |
| CALB | 7.0 | 79.0 [39] | |||
| Lecitase® | 7.0 | 45.0 [38] | |||
| Polyethyleneimine—agarose (+) | PEI | Crosslinked agarose Ligand: Ethyleneimine Type: Cationic, hydrophilic | TLL | 8.0 | 67.2 |
| CALB | 9.5 | 25.2 | |||
| Lecitase® | 8.5 | 94.1 | |||
| Nekrolith® (+) | NK | DVB/styrene tertiary amine groups [40] Type: Cationic, hydrophobic | TLL | 8.5 | 74.5 |
| CALB | 9.5 | 72.6 | |||
| RML | 8.0 | 80.7 | |||
| Lecitase® | 8.5 | 94.1 | |||
| Lewatit® VPOC1600 | VPOC | DVB/Crosslinked polymethacrylate Ligand: None Type: Hydrophobic, Hydrophobic | TLL | 8.0 | 87.7 |
| CALB | 8.0 | 97.0 | |||
| RML | 8.0 | 82.0 | |||
| Lecitase® | 8.0 | 90.0 | |||
| Lewatit® MPSP112H | MPSP | DVB/Crosslinked polystyrene Ligand: Sulfonic acid Type: Anionic, hydrophobic | TLL | 3.5 | 72.0 |
| CALB | 3.5 | 30.0 | |||
| Lecitase® | 7.0 | 80.0 | |||
| Lewatit® MP800 | MP800 | DVB/Crosslinked polystyrene Ligand: Quaternary amine Type: Cationic, hydrophobic | TLL | 8.0 | 94.0 |
| CALB | 9.5 | 27.0 | |||
| Lecitase® | 8.5 | 82.0 | |||
| Purolite® ECR1604 | PL® | Polystyrene Ligand: Quaternary amine Type: Cationic, hydrophobic | TLL | 8.0 | 80.0 |
| CALB | 9.5 | 26.0 | |||
| Lecitase® | 8.5 | 83.2 |
| Support | Enzyme | %EE | CI |
|---|---|---|---|
| Sulfopropyl Sepharose® (−) | TLL | 3.3 | 0.4 |
| CALB | 4.4 | 0.6 | |
| RML | 2.9 | 0.4 | |
| Lecitase® | 3.6 | 0.5 | |
| Q-Sepharose (+) | TLL | 70.1 | 5.6 |
| CALB | 5.7 | 0.5 | |
| Lecitase® | 50.7 | 4.1 | |
| Dextran Sulfate agarose (−) | TLL | 3.2 | 0.4 |
| CALB | 4.4 | 0.6 | |
| RML | 3.4 | 0.4 | |
| Lecitase® | 3.7 | 0.5 | |
| Octyl-Sepharose® | CALB | 32.4 | 2.6 |
| Polyethyleneimine-agarose (+) | TLL | 4.7 | 0.4 |
| CALB | 35.8 | 2.9 | |
| Lecitase® | 68.4 | 5.5 | |
| Nekrolith® (+) | TLL | 51.3 | 4.1 |
| CALB | 30.4 | 2.4 | |
| RML | 20.6 | 1.6 | |
| Lecitase® | 22.8 | 1.8 | |
| Lewatit® VPOC1600 | TLL | 86.2 | 6.9 |
| CALB | 47.4 | 3.8 | |
| RML | 70.7 | 5.7 | |
| Lecitase® | 11.4 | 0.9 | |
| Lewatit® MPSP112H (−) | TLL | 11.0 | 0.9 |
| CALB | 9.0 | 0.7 | |
| Lecitase® | 10.0 | 0.8 | |
| Lewatit® MP800 (+) | TLL | 65.1 | 5.2 |
| CALB | 27.2 | 2.2 | |
| Lecitase® | 55.0 | 4.4 | |
| Purolite® ECR1604 | TLL | 54.2 | 4.3 |
| CALB | 10.1 | 0.8 | |
| Lecitase® | 16.3 | 1.3 | |
| Novozyme® 435 | 52.7 | 4.2 | |
| Lipozyme® TL IM | 71.3 | 5.7 | |
| Derivative (Support/Additive during Immobilization) | Surfactant-Free Derivative Esterase Activity (UI) | Ratio between Expressed and Initial Activity |
|---|---|---|
| Q-Sepharose®/WA | 0.09 ± 0.01 | 8.2 |
| Q-Sepharose®/CTAB | 0.048 ± 0.007 | 4.4 |
| Q-Sepharose®/SDS | 1.02 ± 0.09 | 92.7 |
| Derivative (Support/Condition of Immobilization) | Additive Added to the Esterase Reaction Medium (Concentration Yielding Maximum Activity.) | Maximum Activity Observed | |
|---|---|---|---|
| Relative | (UI) | ||
| Q-Sepharose®/WA | Without additives | 1 | 0.09± 0.01 |
| CTAB (0.1%) | 33 | 2.98 ± 0.06 | |
| SDS (0.01%) | 14 | 1.24 ± 0.06 | |
| PEI (0.1%) | 1.8 | 0.161 ± 0.006 | |
| CMC (0.1%) | 1.4 | 0.13 ± 0.01 | |
| Q-Sepharose®/CTAB (0.005%) | Without additives | 1 | 0.048 ± 0.007 |
| CTAB (0.1%) | 61 | 2.92 ± 0.06 | |
| SDS (0.01%) | 30 | 1.43 ± 0.05 | |
| PEI (0.1%) | 2 | 0.095 ± 0.007 | |
| CMC (0.1%) | 2 | 0.086 ± 0.006 | |
| Q-Sepharose®/SDS (0.1%) | Without additives | 1 | 1.02 ± 0.09 |
| CTAB (0.01%) | 0.51 | 0.52 ± 0.1 | |
| SDS (0.01%) | 0.64 | 0.65 ± 0.01 | |
| PEI (0.01%) | 0.60 | 0.61 ± 0.03 | |
| CMC (0.01%) | 0.65 | 0.66 ± 0.02 | |
| Unrefined Palm Oil | Used Palm Oil | |||
|---|---|---|---|---|
| Time (h) | Q-SDS-TLL | Commercial Derivative Lipozyme® | Q-SDS-TLL | Commercial Derivativelipozyme® |
| 1 | 28.5 ± 2.7 | 17.7 ± 0.3 | 28.1 ± 2.9 | 34.5 ± 3.2 |
| 6 | 45.9 ± 6.0 | 50.0 ± 5.8 | 62.6 ± 1.8 | 64.4 ± 1.7 |
| 24 | 67.5 ± 2.6 | 64.6 ± 4.0 | 79.3 ± 0.2 | 77.8 ± 0.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pardo-Tamayo, J.S.; Arteaga-Collazos, S.; Domínguez-Hoyos, L.C.; Godoy, C.A. Biocatalysts Based on Immobilized Lipases for the Production of Fatty Acid Ethyl Esters: Enhancement of Activity through Ionic Additives and Ion Exchange Supports. BioTech 2023, 12, 67. https://doi.org/10.3390/biotech12040067
Pardo-Tamayo JS, Arteaga-Collazos S, Domínguez-Hoyos LC, Godoy CA. Biocatalysts Based on Immobilized Lipases for the Production of Fatty Acid Ethyl Esters: Enhancement of Activity through Ionic Additives and Ion Exchange Supports. BioTech. 2023; 12(4):67. https://doi.org/10.3390/biotech12040067
Chicago/Turabian StylePardo-Tamayo, Juan S., Sebastián Arteaga-Collazos, Laura C. Domínguez-Hoyos, and César A. Godoy. 2023. "Biocatalysts Based on Immobilized Lipases for the Production of Fatty Acid Ethyl Esters: Enhancement of Activity through Ionic Additives and Ion Exchange Supports" BioTech 12, no. 4: 67. https://doi.org/10.3390/biotech12040067