
Psychosocial Stress Induces Orofacial Mechanical Allodynia Due to the Enhancement of Transient Receptor Potential Ankyrin 1 Expression in Trigeminal Ganglion Neurons via the Increment of the Trace Amine-Associated Receptor 7f Expression


 , ,
, ,
Abstract
:1. Introduction
2. Results
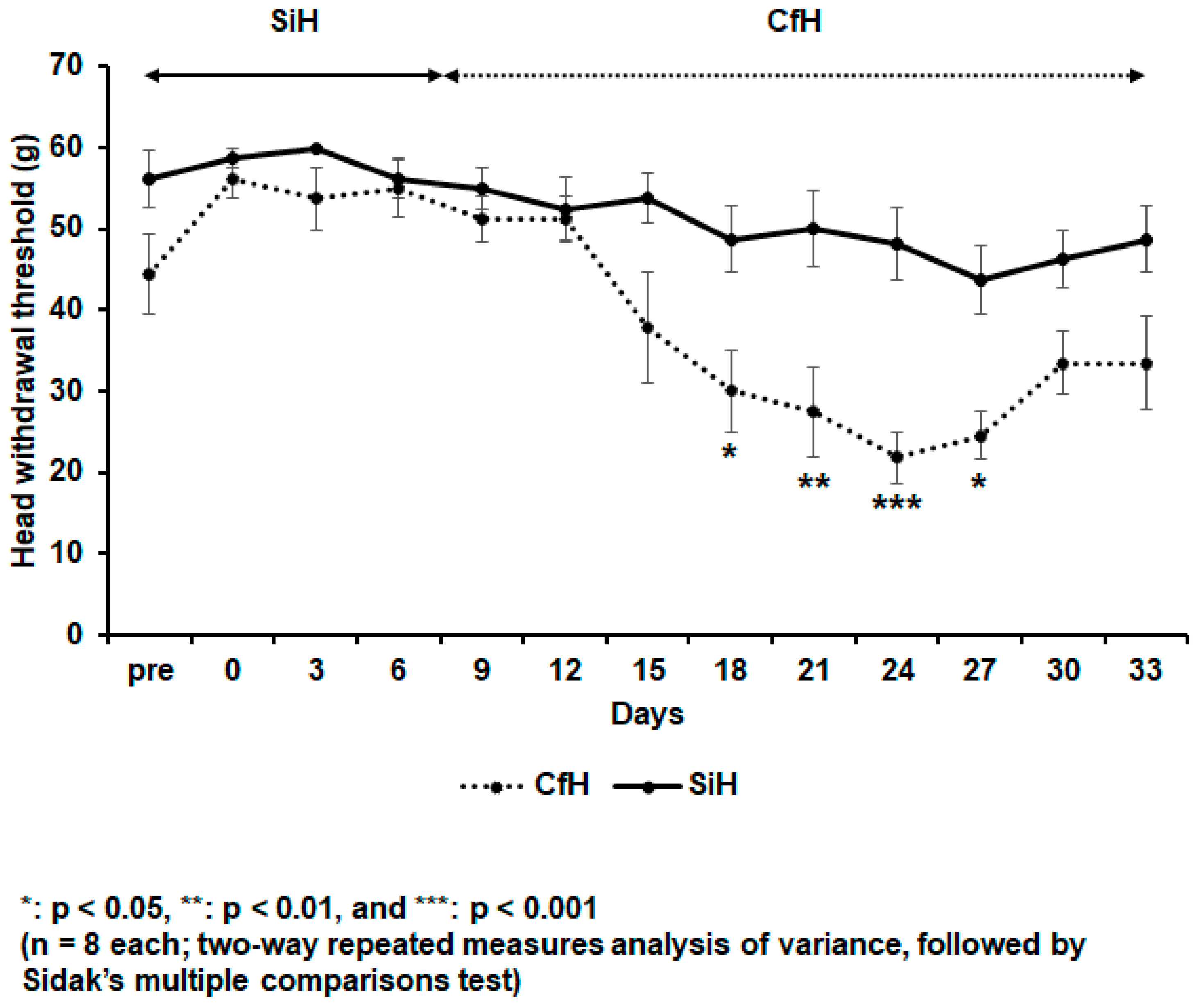
2.1. The Psychosocial Stress Decreased the Orofacial HWT
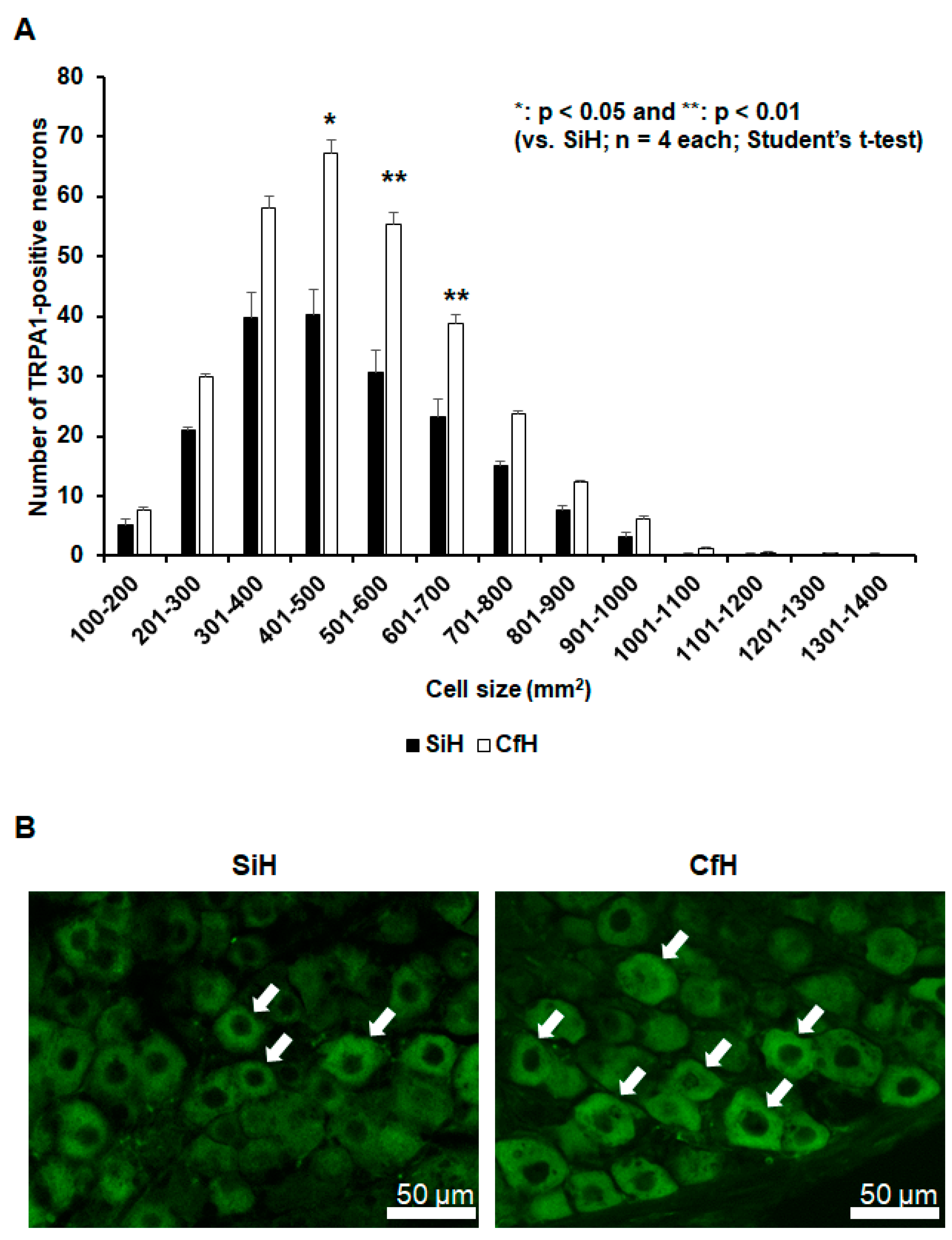
2.2. TRPA1-Positive Trigeminal Ganglion Neurons Increased in Mice under Psychosocial Stress Conditions
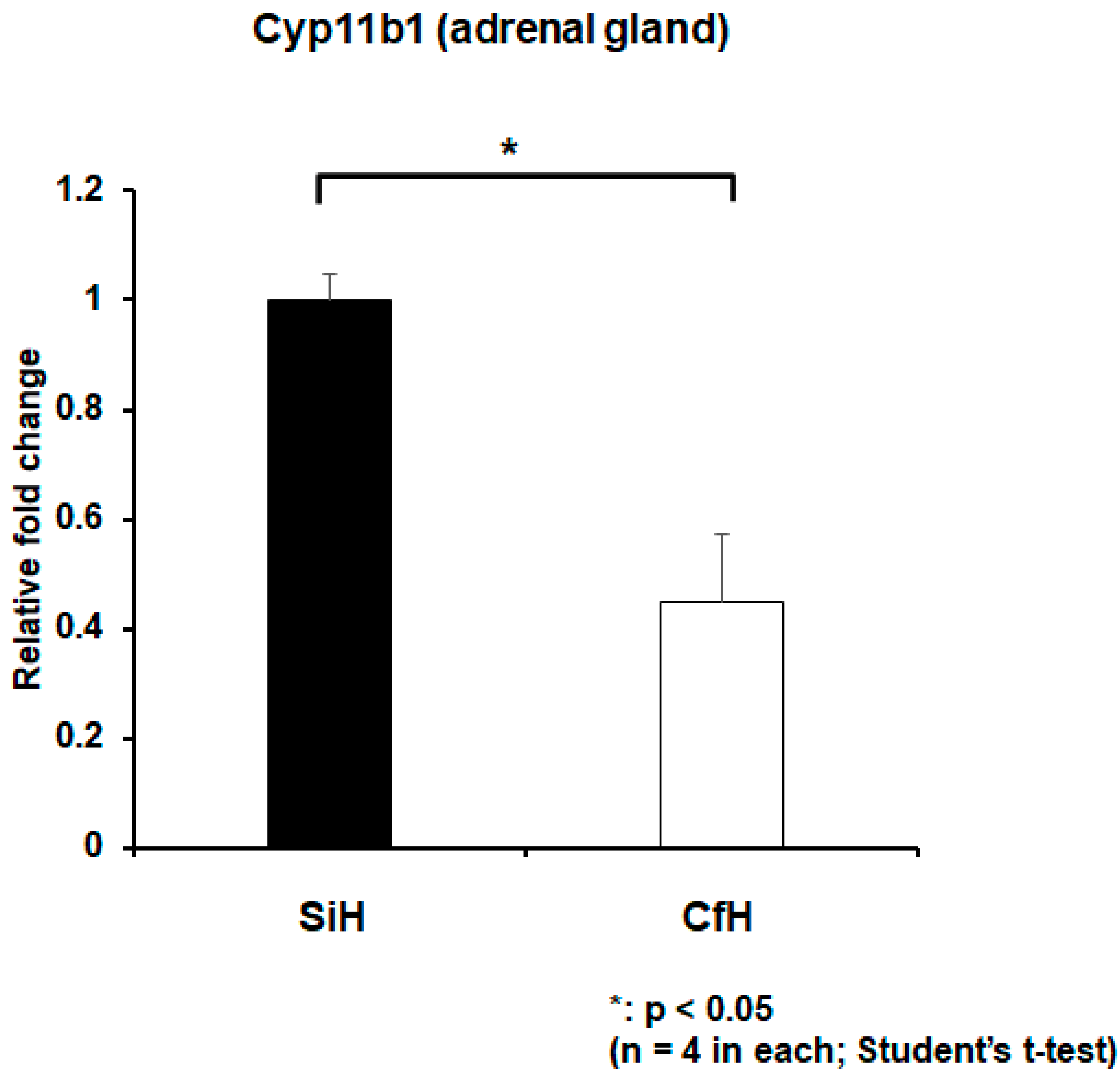
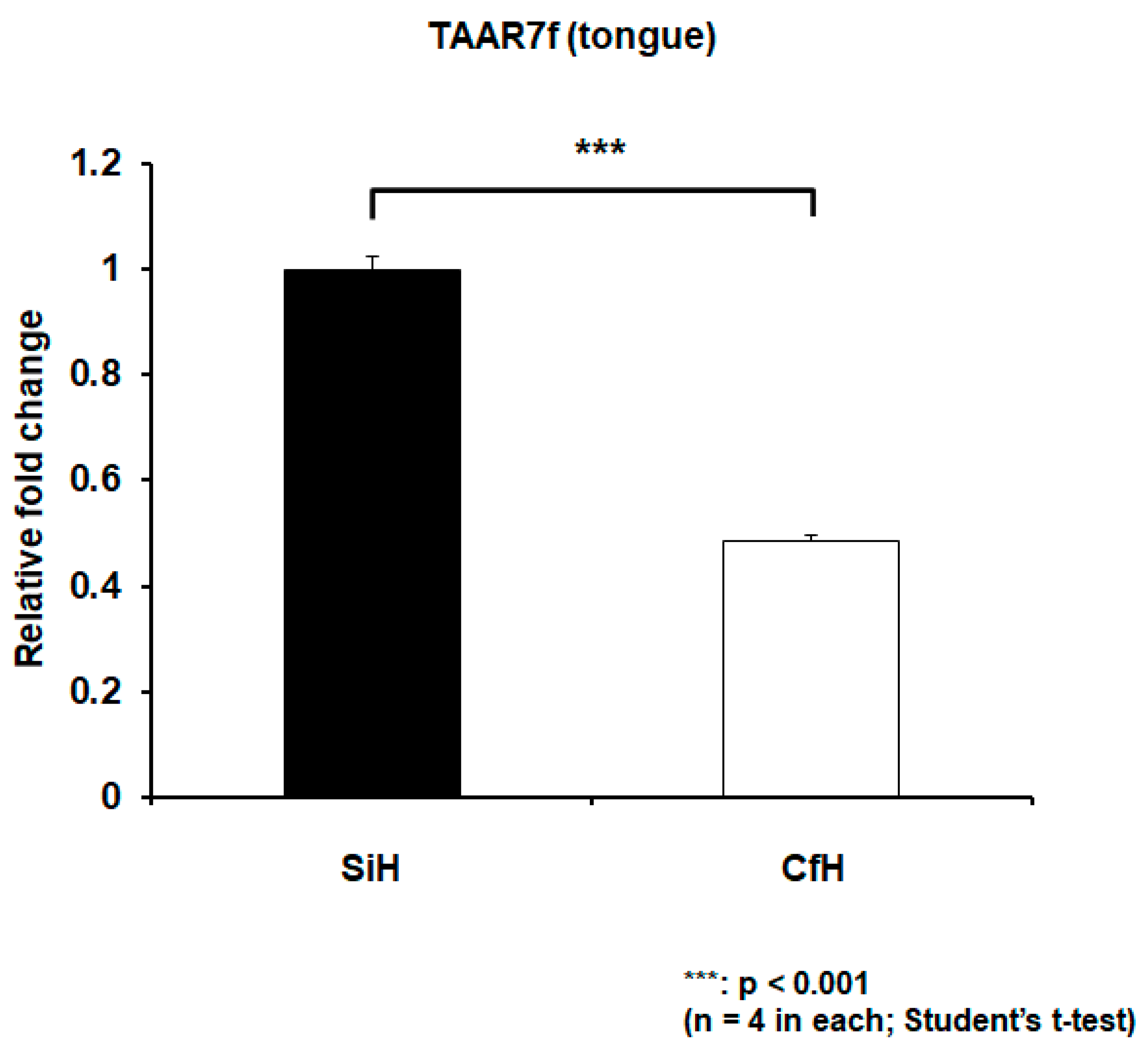
2.3. TAAR7f Expression Increased in Circulating Cells and Decreased in Tongue under Psychosocial Stress Conditions
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Psychosocial Stress Experiment
4.3. Blood, Adrenal Gland, and Tongue Sampling
4.4. RNA Isolation
4.5. Mechanical Pain Sensitivity in Whisker Pad Skin
4.6. Immunohistochemistry
4.7. DNA Microarray Analysis
4.8. Quantitative Reverse Transcription Polymerase Chain Reaction Analysis
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McEwen, B.S. Stress, adaptation, and disease. Allostasis and allostatic load. Ann. N. Y. Acad. Sci. 1998, 840, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Woda, A.; Picard, P.; Dutheil, F. Dysfunctional stress responses in chronic pain. Psychoneuroendocrinology 2016, 71, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Lamey, P.J.; Freeman, R.; Eddie, S.A.; Pankhurst, C.; Rees, T. Vulnerability and presenting symptoms in burning mouth syndrome. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 2005, 99, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Femiano, F.; Scully, C. Burning mouth syndrome (BMS): Double blind controlled study of alpha-lipoic acid (thioctic acid) therapy. J. Oral Pathol. Med. 2002, 31, 267–269. [Google Scholar] [CrossRef] [PubMed]
- Carlson, C.R.; Miller, C.S.; Reid, K.I. Psychosocial profiles of patients with burning mouth syndrome. J. Orofac. Pain 2000, 14, 59–64. [Google Scholar]
- Ohga, N.; Yamazaki, T.; Sato, J.; Hata, H.; Murata, T.; Sakata, K.; Inoue, T.; Kitagawa, Y. Dose escalation effectiveness and tolerability of paroxetine in patients with burning mouth syndrome and depressive conditions. J. Oral Maxillofac. Surg. Med. Pathol. 2015, 27, 402–406. [Google Scholar] [CrossRef]
- Maina, G.; Vitalucci, A.; Gandolfo, S.; Bogetto, F. Comparative efficacy of SSRIs and amisulpride in burning mouth syndrome: A single-blind study. J. Clin. Psychiatry 2002, 63, 38–43. [Google Scholar] [CrossRef]
- Puhakka, A.; Forssell, H.; Soinila, S.; Virtanen, A.; Röyttä, M.; Laine, M.; Tenovuo, O.; Teerijoki-Oksa, T.; Jääskeläinen, S.K. Peripheral nervous system involvement in primary burning mouth syndrome--results of a pilot study. Oral Dis. 2016, 22, 338–344. [Google Scholar] [CrossRef]
- Lauria, G.; Majorana, A.; Borgna, M.; Lombardi, R.; Penza, P.; Padovani, A.; Sapelli, P. Trigeminal small-fiber sensory neuropathy causes burning mouth syndrome. Pain 2005, 115, 332–337. [Google Scholar] [CrossRef]
- Marone, I.M.; De Logu, F.; Nassini, R.; De Carvalho Goncalves, M.; Benemei, S.; Ferreira, J.; Jain, P.; Li Puma, S.; Bunnett, N.W.; Geppetti, P.; et al. TRPA1/NOX in the soma of trigeminal ganglion neurons mediates migraine-related pain of glyceryl trinitrate in mice. Brain 2018, 141, 2312–2328. [Google Scholar] [CrossRef] [Green Version]
- Story, G.M.; Peier, A.M.; Reeve, A.J.; Eid, S.R.; Mosbacher, J.; Hricik, T.R.; Earley, T.J.; Hergarden, A.C.; Andersson, D.A.; Hwang, S.W.; et al. ANKTM1, a TRP-like channel expressed in nociceptive neurons, is activated by cold temperatures. Cell 2003, 112, 819–829. [Google Scholar] [CrossRef] [PubMed]
- Obata, K.; Katsura, H.; Mizushima, T.; Yamanaka, H.; Kobayashi, K.; Dai, Y.; Fukuoka, T.; Tokunaga, A.; Tominaga, M.; Noguchi, K. TRPA1 induced in sensory neurons contributes to cold hyperalgesia after inflammation and nerve injury. J. Clin. Investig. 2005, 115, 2393–2401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koivisto, A.; Hukkanen, M.; Saarnilehto, M.; Chapman, H.; Kuokkanen, K.; Wei, H.; Viisanen, H.; Akerman, K.E.; Lindstedt, K.; Pertovaara, A. Inhibiting TRPA1 ion channel reduces loss of cutaneous nerve fiber function in diabetic animals: Sustained activation of the TRPA1 channel contributes to the pathogenesis of peripheral diabetic neuropathy. Pharmacol. Res. 2012, 65, 149–158. [Google Scholar] [CrossRef]
- Trevisan, G.; Materazzi, S.; Fusi, C.; Altomare, A.; Aldini, G.; Lodovici, M.; Patacchini, R.; Geppetti, P.; Nassini, R. Novel therapeutic strategy to prevent chemotherapy-induced persistent sensory neuropathy by TRPA1 blockade. Cancer Res. 2013, 73, 3120–3131. [Google Scholar] [CrossRef] [Green Version]
- Elitt, C.M.; Malin, S.A.; Koerber, H.R.; Davis, B.M.; Albers, K.M. Overexpression of artemin in the tongue increases expression of TRPV1 and TRPA1 in trigeminal afferents and causes oral sensitivity to capsaicin and mustard oil. Brain Res. 2008, 1230, 80–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Unno, K.; Fujitani, K.; Takamori, N.; Takabayashi, F.; Maeda, K.; Miyazaki, H.; Tanida, N.; Iguchi, K.; Shimoi, K.; Hoshino, M. Theanine intake improves the shortened lifespan, cognitive dysfunction and behavioural depression that are induced by chronic psychosocial stress in mice. Free Radic. Res. 2011, 45, 966–974. [Google Scholar] [CrossRef]
- Unno, K.; Iguchi, K.; Tanida, N.; Fujitani, K.; Takamori, N.; Yamamoto, H.; Ishii, N.; Nagano, H.; Nagashima, T.; Hara, A.; et al. Ingestion of theanine, an amino acid in tea, suppresses psychosocial stress in mice. Exp. Physiol. 2013, 98, 290–303. [Google Scholar] [CrossRef] [PubMed]
- Bugda Gwilt, K.; González, D.P.; Olliffe, N.; Oller, H.; Hoffing, R.; Puzan, M.; El Aidy, S.; Miller, G.M. Actions of Trace Amines in the Brain-Gut-Microbiome Axis via Trace Amine-Associated Receptor-1 (TAAR1). Cell. Mol. Neurobiol. 2020, 40, 191–201. [Google Scholar] [CrossRef]
- Souza Monteiro de Araujo, D.; Nassini, R.; Geppetti, P.; De Logu, F. TRPA1 as a therapeutic target for nociceptive pain. Expert. Opin. Ther. Targets 2020, 24, 997–1008. [Google Scholar] [CrossRef]
- Konkoly, J.; Kormos, V.; Gaszner, B.; Sándor, Z.; Kecskés, A.; Alomari, A.; Szilágyi, A.; Szilágyi, B.; Zelena, D.; Pintér, E. The Role of TRPA1 Channels in the Central Processing of Odours Contributing to the Behavioural Responses of Mice. Pharmaceuticals 2021, 14, 1336. [Google Scholar] [CrossRef]
- Manchikanti, L.; Fellows, B.; Singh, V. Understanding psychological aspects of chronic pain in interventional pain management. Pain Physician 2002, 5, 57–82. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Suttle, A.; Zhang, Q.; Wang, P.; Chen, Y. Transient Receptor Potential (TRP) Ion Chan-nels in Orofacial Pain. Mol. Neurobiol. 2021, 58, 2836–2850. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.H.; Chang, Y.S.; Hsieh, Y.L. Transient receptor potential vanilloid subtype 1 deple-tion mediates mechanical allodynia through cellular signal alterations in small-fiber neuropathy. Pain Rep. 2021, 6, e922. [Google Scholar] [CrossRef] [PubMed]
- Ito, M.; Ono, K.; Hitomi, S.; Nodai, T.; Sago, T.; Yamaguchi, K.; Harano, N.; Gunnjigake, K.; Hosokawa, R.; Kawamoto, T.; et al. Prostanoid-dependent spontaneous pain and PAR2-dependent mechanical allodynia following oral mucosal trauma: Involvement of TRPV1, TRPA1 and TRPV4. Mol. Pain 2017, 13, 1744806917704138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuo, X.; Ling, J.X.; Xu, G.Y.; Gu, J.G. Operant behavioral responses to orofacial cold stimuli in rats with chronic constrictive trigeminal nerve injury: Effects of menthol and capsazepine. Mol. Pain 2013, 9, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borowsky, B.; Adham, N.; Jones, K.A.; Raddatz, R.; Artymyshyn, R.; Ogozalek, K.L.; Durkin, M.M.; Lakhlani, P.P.; Bonini, J.A.; Pathirana, S.; et al. Trace amines: Identification of a family of mammalian G protein-coupled receptors. Proc. Natl. Acad. Sci. USA 2001, 98, 8966–8971. [Google Scholar] [CrossRef] [Green Version]
- Regard, J.B.; Kataoka, H.; Cano, D.A.; Camerer, E.; Yin, L.; Zheng, Y.W.; Scanlan, T.S.; Hebrok, M.; Coughlin, S.R. Probing cell type-specific functions of Gi in vivo identifies GPCR regulators of insulin secretion. J. Clin. Investig. 2007, 117, 4034–4043. [Google Scholar] [CrossRef] [Green Version]
- Raab, S.; Wang, H.; Uhles, S.; Cole, N.; Alvarez-Sanchez, R.; Künnecke, B.; Ullmer, C.; Matile, H.; Bedoucha, M.; Norcross, R.D.; et al. Incretin-like effects of small molecule trace amine-associated receptor 1 agonists. Mol. Metab. 2016, 5, 47–56. [Google Scholar] [CrossRef]
- Liberles, S.D.; Buck, L.B. A second class of chemosensory receptors in the olfactory epithelium. Nature 2006, 442, 645–650. [Google Scholar] [CrossRef]
- Chiellini, G.; Erba, P.; Carnicelli, V.; Manfredi, C.; Frascarelli, S.; Ghelardoni, S.; Mariani, G.; Zucchi, R. Distribution of exogenous [125I]-3-iodothyronamine in mouse in vivo: Relationship with trace amine-associated receptors. J. Endocrinol. 2012, 213, 223–230. [Google Scholar] [CrossRef]
- Babusyte, A.; Kotthoff, M.; Fiedler, J.; Krautwurst, D. Biogenic amines activate blood leukocytes via trace amine-associated receptors TAAR1 and TAAR2. J. Leukoc. Biol. 2013, 93, 387–394. [Google Scholar] [CrossRef] [PubMed]
- Sriram, U.; Cenna, J.M.; Haldar, B.; Fernandes, N.C.; Razmpour, R.; Fan, S.; Ramirez, S.H.; Potula, R. Methamphetamine induces trace amine-associated receptor 1 (TAAR1) expression in human T lymphocytes: Role in immunomodulation. J. Leukoc. Biol. 2016, 99, 213–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, D.A.; Tolbert, M.D.; Singh, S.J.; Bost, K.L. Expression of neuronal trace amine-associated receptor (Taar) mRNAs in leukocytes. J. Neuroimmunol. 2007, 192, 21–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, P.; Weber, C.; Hummel, T. Brain response to intranasal trimethylamine stimulation: A preliminary human fMRI study. Neurosci. Lett. 2020, 735, 135166. [Google Scholar] [CrossRef]
- Shi, X.; Swanson, T.L.; Miner, N.B.; Eshleman, A.J.; Janowsky, A. Activation of Trace Amine-Associated Receptor 1 Stimulates an Antiapoptotic Signal Cascade via Extracellular Signal-Regulated Kinase 1/2. Mol. Pharmacol. 2019, 96, 493–504. [Google Scholar] [CrossRef]
- Santoni, G.; Nabissi, M.; Amantini, C.; Santoni, M.; Ricci-Vitiani, L.; Pallini, R.; Maggi, F.; Morelli, M.B. ERK Phosphorylation Regulates the Aml1/Runx1 Splice Variants and the TRP Channels Expression during the Differentiation of Glioma Stem Cell Lines. Cells 2021, 10, 2052. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Description | Fold Change |
|---|---|---|
| Taar7f | Trace amine-associated receptor 7F | 2.43 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ito, K.; Shinoda, M.; Soma, K.; Ikutame, D.; Ikami, E.; Mizuno, Y.; Usui, M.; Asoda, S.; Sato, T. Psychosocial Stress Induces Orofacial Mechanical Allodynia Due to the Enhancement of Transient Receptor Potential Ankyrin 1 Expression in Trigeminal Ganglion Neurons via the Increment of the Trace Amine-Associated Receptor 7f Expression. Stresses 2023, 3, 58-68. https://doi.org/10.3390/stresses3010006
Ito K, Shinoda M, Soma K, Ikutame D, Ikami E, Mizuno Y, Usui M, Asoda S, Sato T. Psychosocial Stress Induces Orofacial Mechanical Allodynia Due to the Enhancement of Transient Receptor Potential Ankyrin 1 Expression in Trigeminal Ganglion Neurons via the Increment of the Trace Amine-Associated Receptor 7f Expression. Stresses. 2023; 3(1):58-68. https://doi.org/10.3390/stresses3010006
Chicago/Turabian StyleIto, Ko, Masamichi Shinoda, Kumi Soma, Daisuke Ikutame, Eiji Ikami, Yosuke Mizuno, Michihiko Usui, Seiji Asoda, and Tsuyoshi Sato. 2023. "Psychosocial Stress Induces Orofacial Mechanical Allodynia Due to the Enhancement of Transient Receptor Potential Ankyrin 1 Expression in Trigeminal Ganglion Neurons via the Increment of the Trace Amine-Associated Receptor 7f Expression" Stresses 3, no. 1: 58-68. https://doi.org/10.3390/stresses3010006
APA StyleIto, K., Shinoda, M., Soma, K., Ikutame, D., Ikami, E., Mizuno, Y., Usui, M., Asoda, S., & Sato, T. (2023). Psychosocial Stress Induces Orofacial Mechanical Allodynia Due to the Enhancement of Transient Receptor Potential Ankyrin 1 Expression in Trigeminal Ganglion Neurons via the Increment of the Trace Amine-Associated Receptor 7f Expression. Stresses, 3(1), 58-68. https://doi.org/10.3390/stresses3010006