
Kefir Culture-Mediated Fermentation to Improve Phenolic-Linked Antioxidant, Anti-Hyperglycemic and Human Gut Health Benefits in Sprouted Food Barley



Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Sprouted Barley Flour and Aqueous Slurries
2.2. Kefir Culture Preparation, Inoculation, and Fermentation
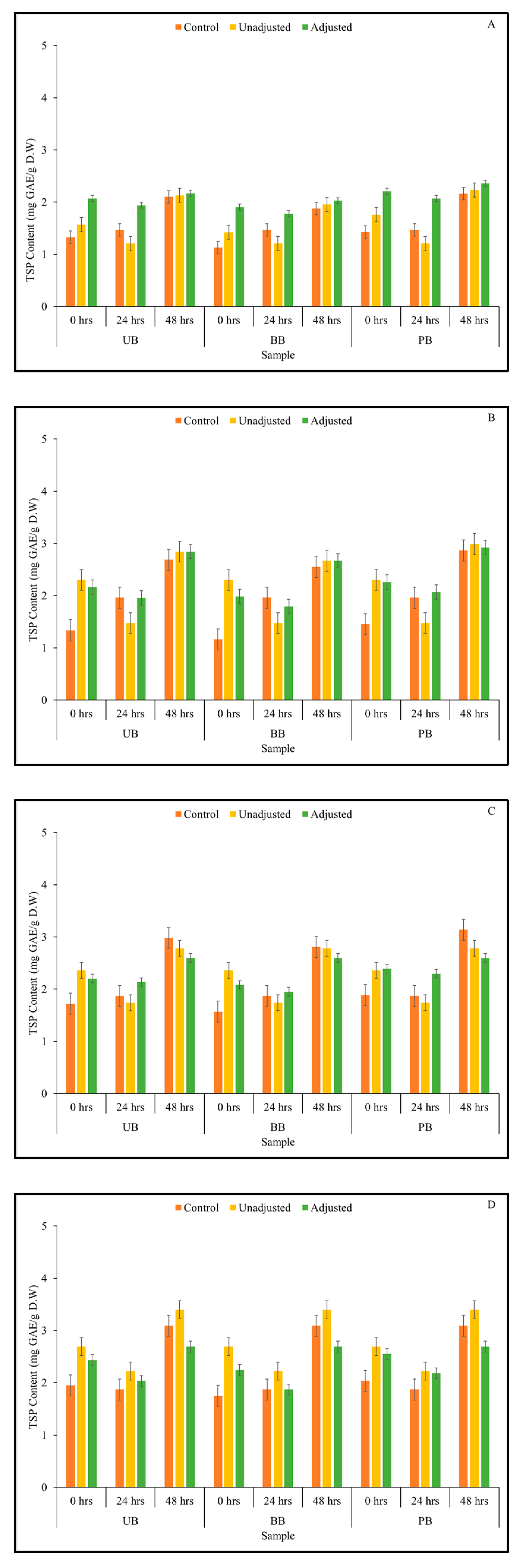
2.3. Total Soluble Phenolic (TSP) Content
2.4. Quantification of Major Phenolic Compounds
2.5. Total Antioxidant Activity Assays
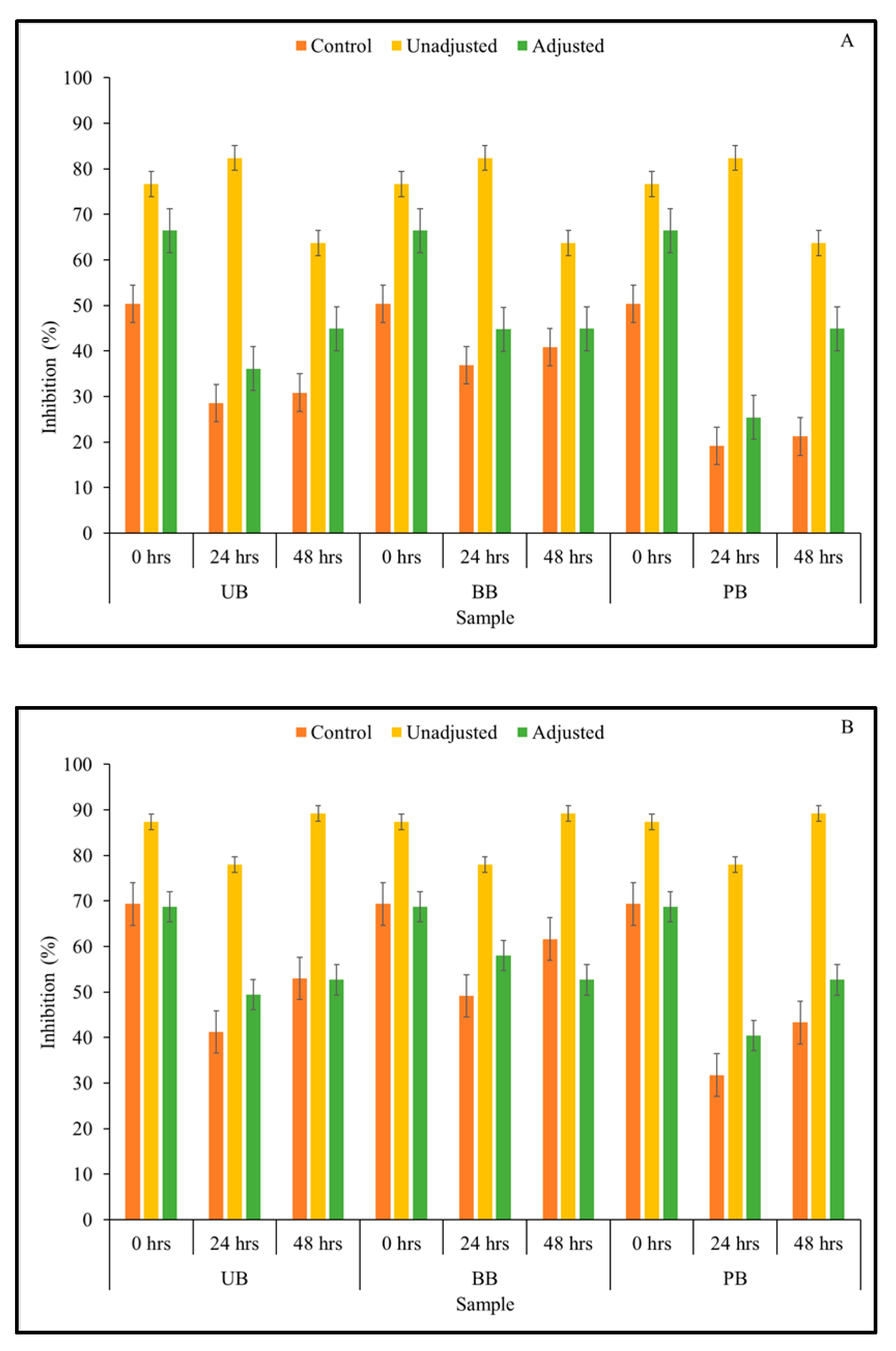
2.6. α-Amylase Inhibitory Activity
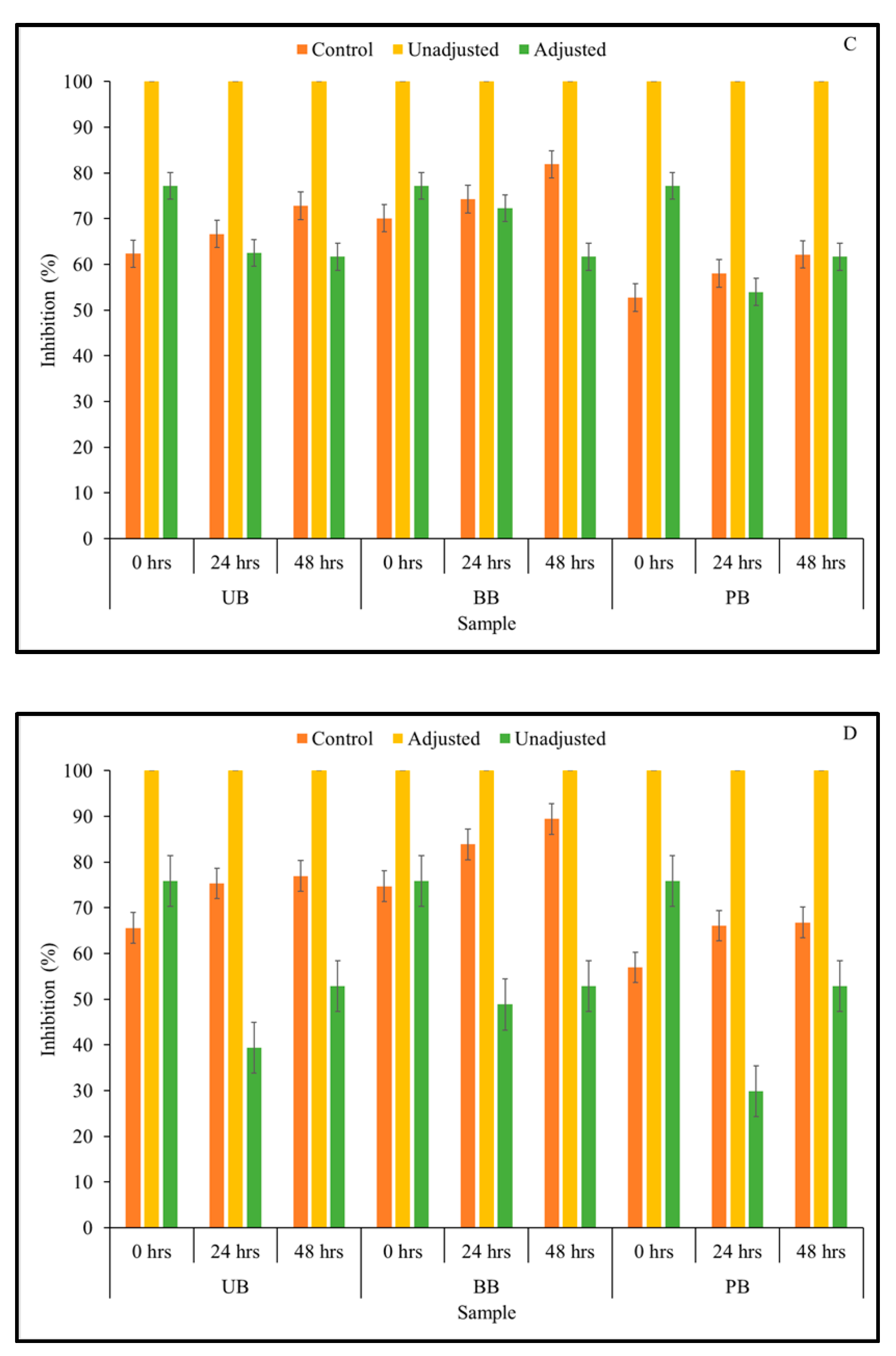
2.7. α-Glucosidase Inhibitory Activity
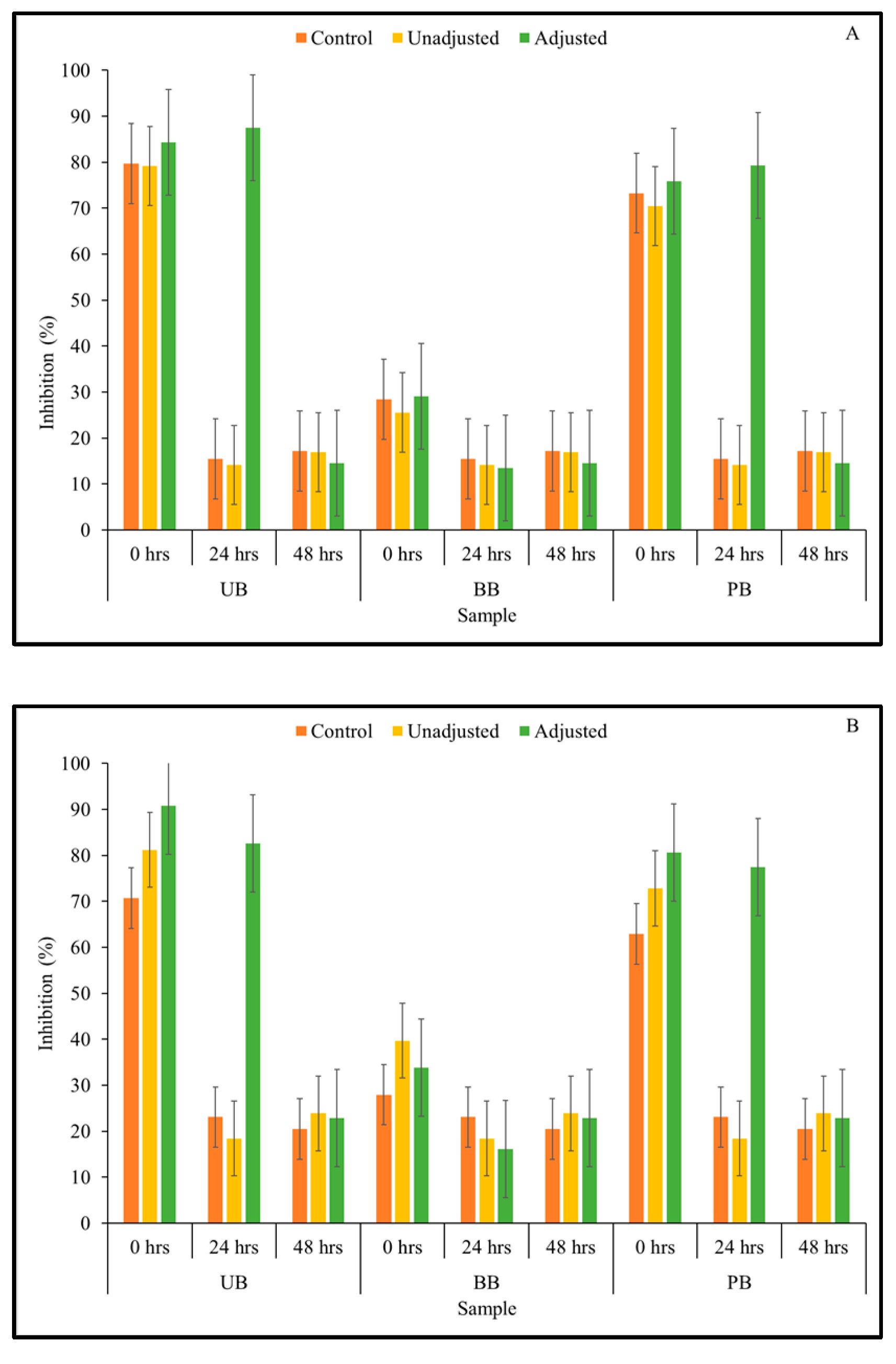
2.8. Helicobacter Pylori Inhibition Assay
2.9. Viable Cell Count of Beneficial LAB Strains
2.10. Statistical Analysis
3. Results and Discussion
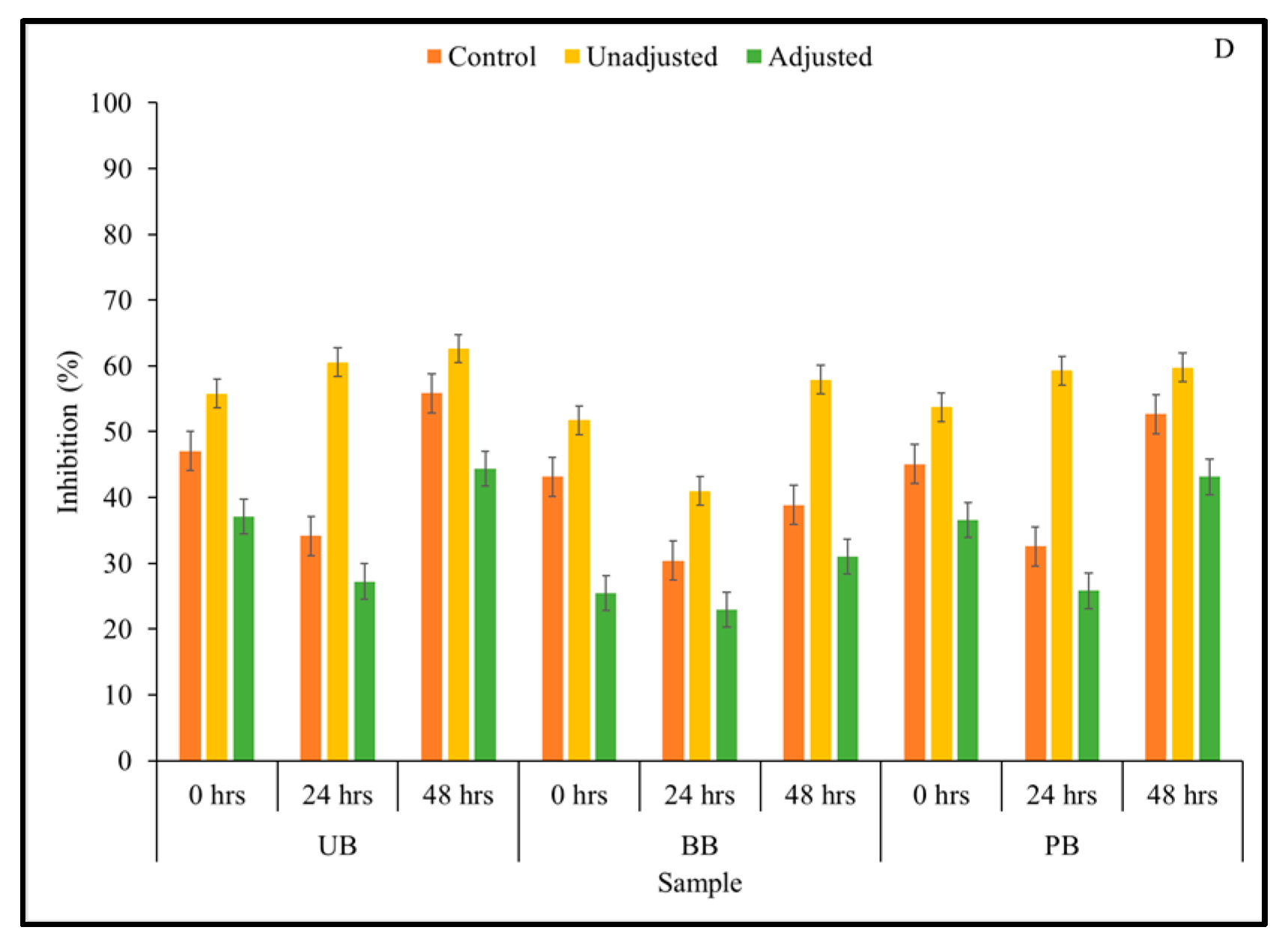
3.1. Total Soluble Phenolic Content and Phenolic Compound Profile
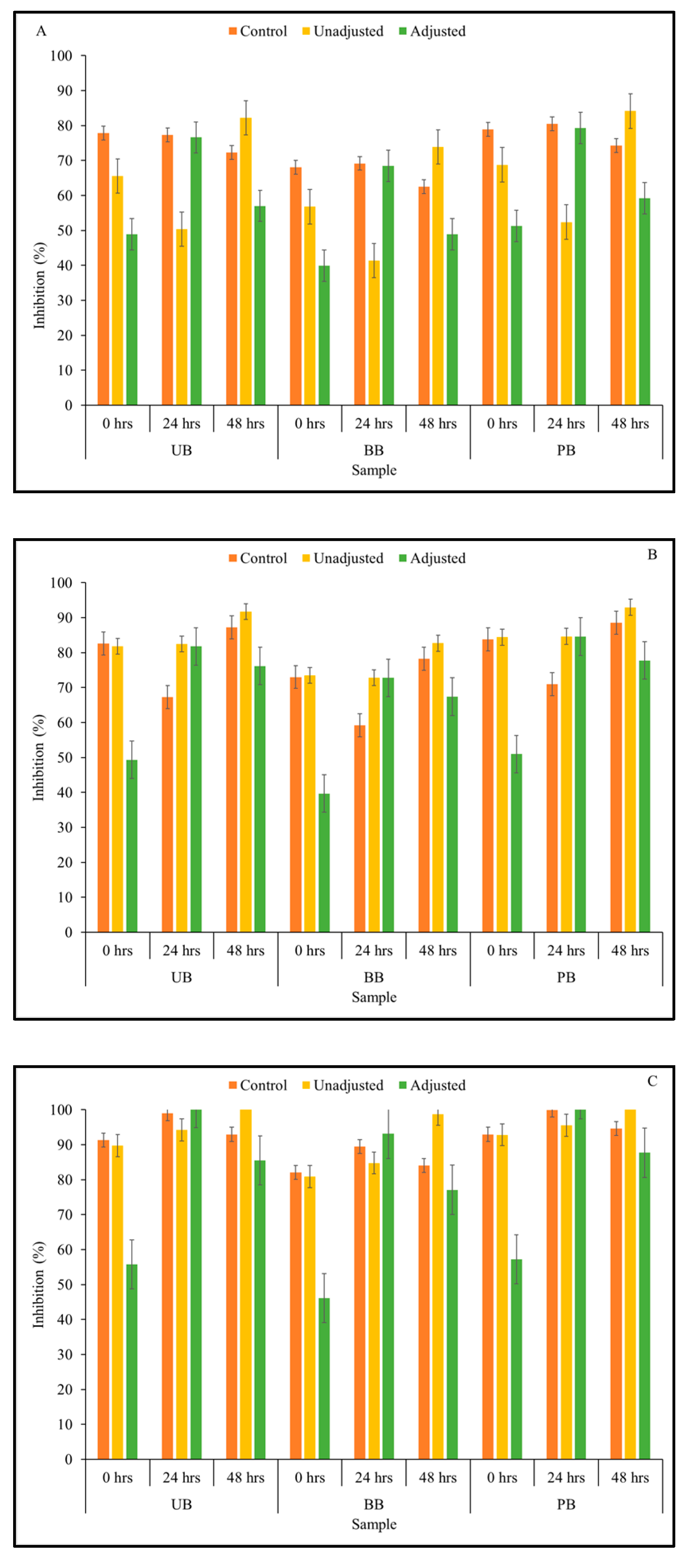
3.2. Total Antioxidant Activity
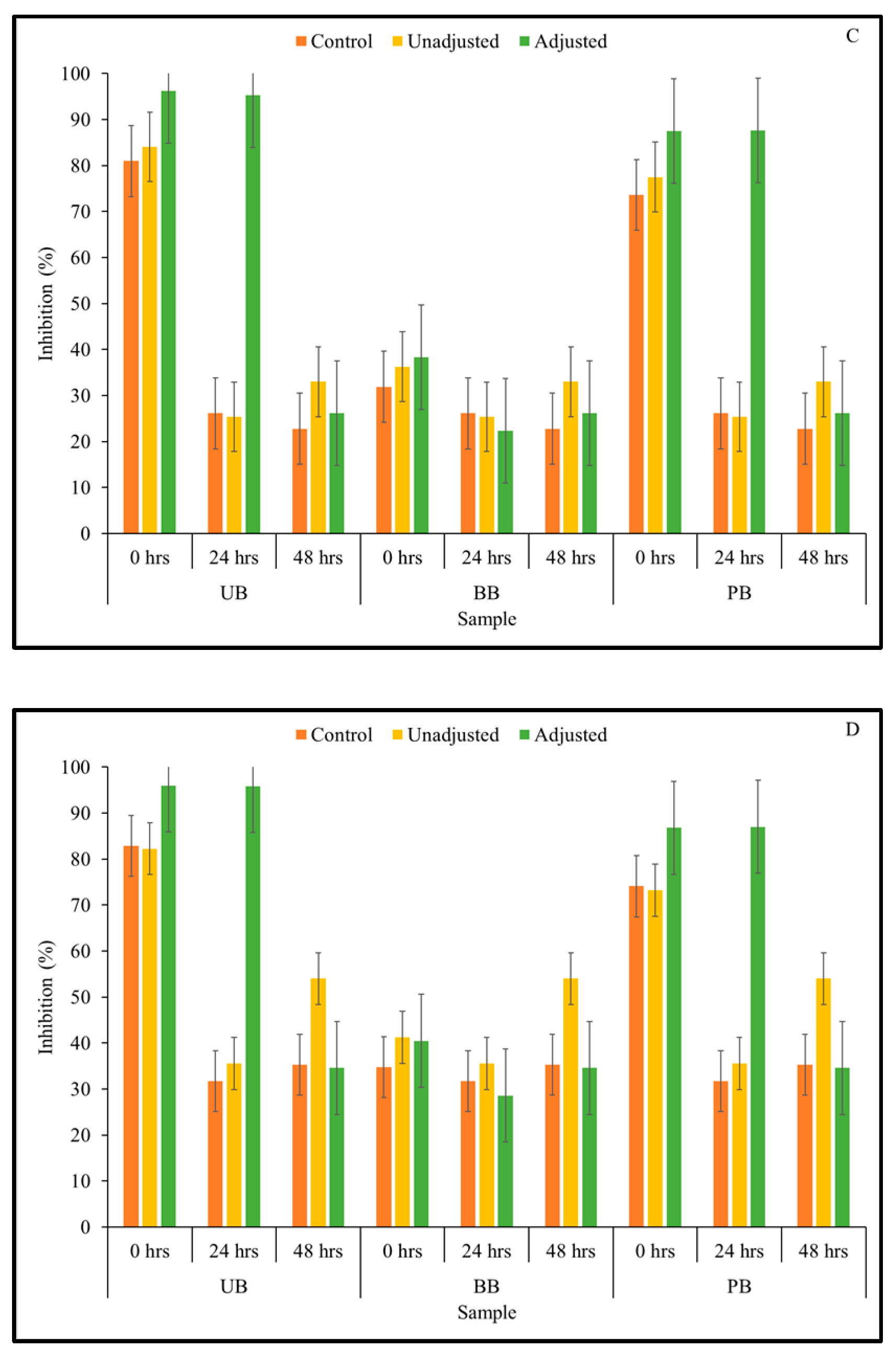
3.3. α-Amylase Enzyme Inhibitory Activity
3.4. α-Glucosidase Enzyme Inhibitory Activity
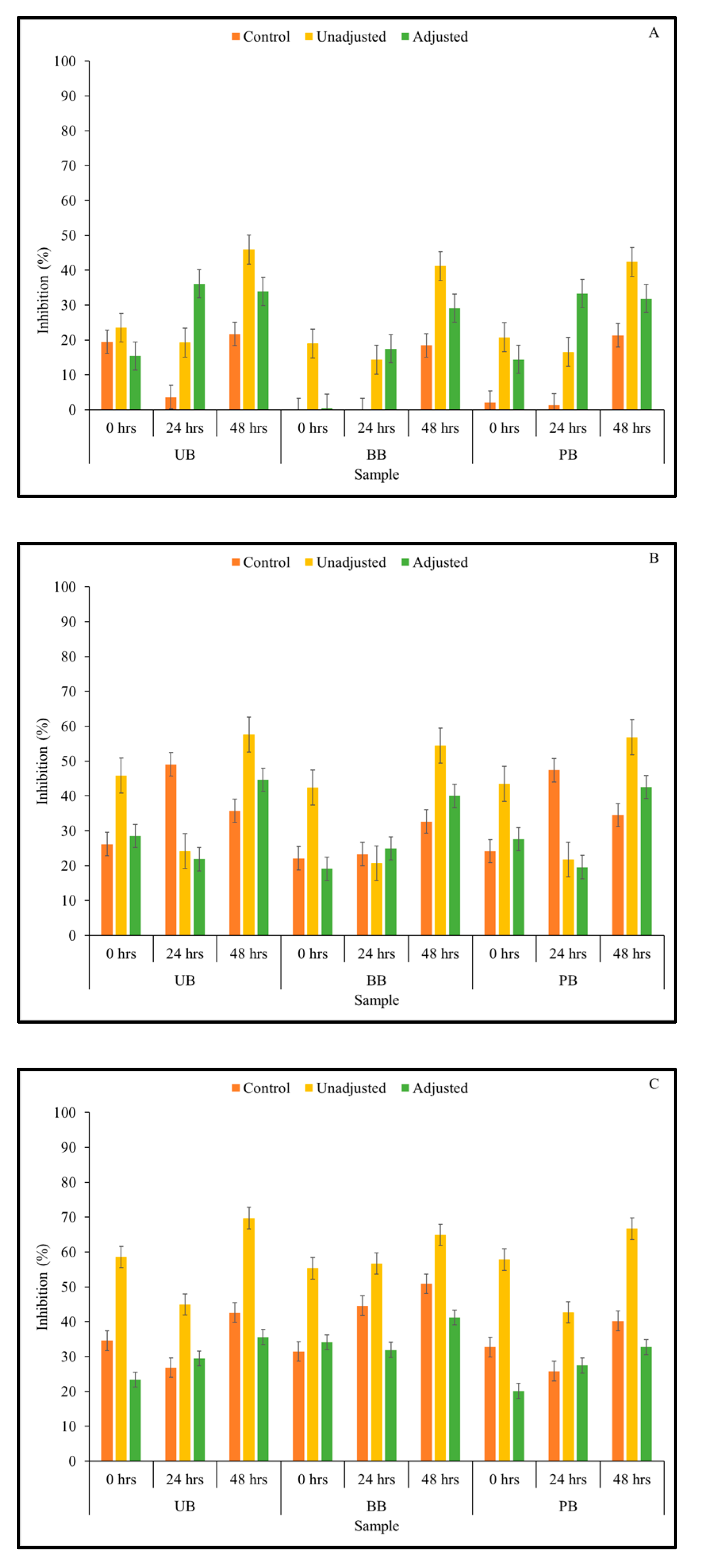
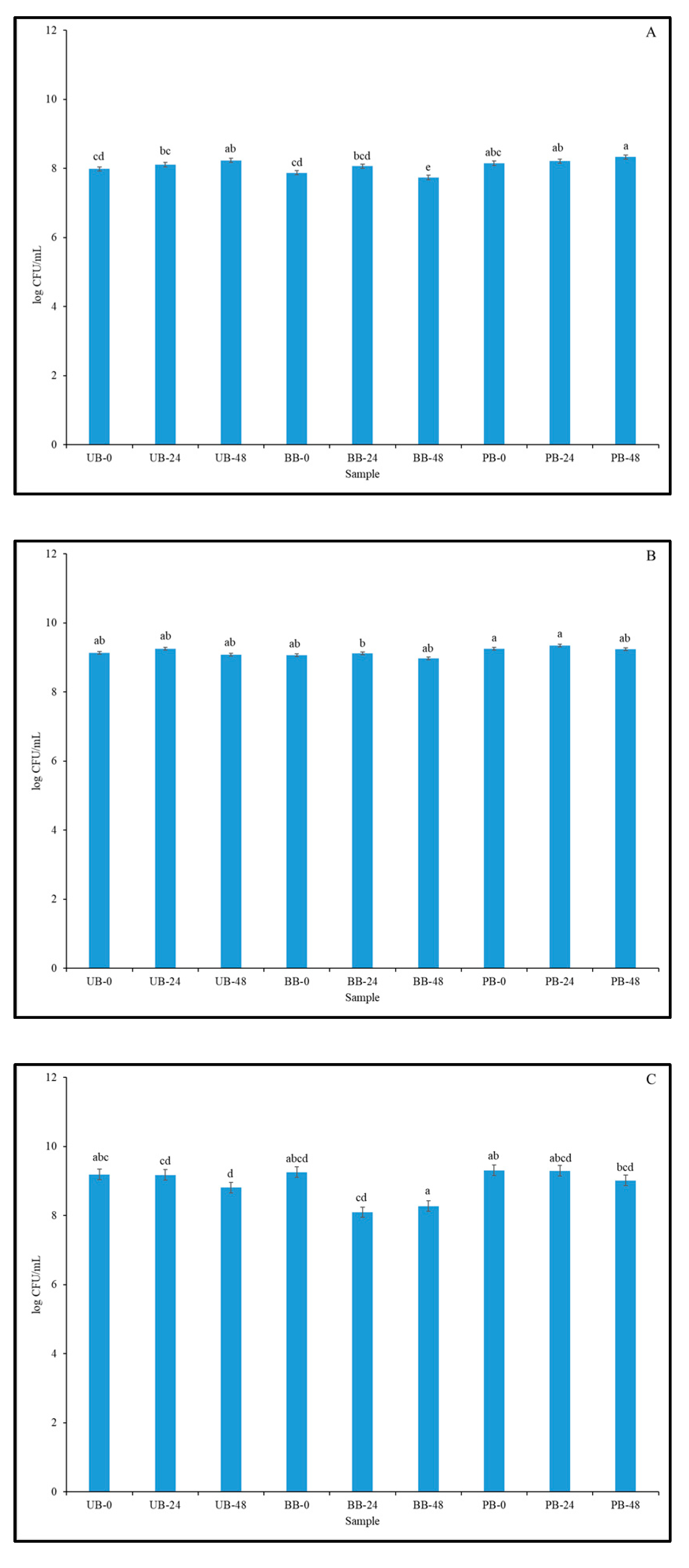
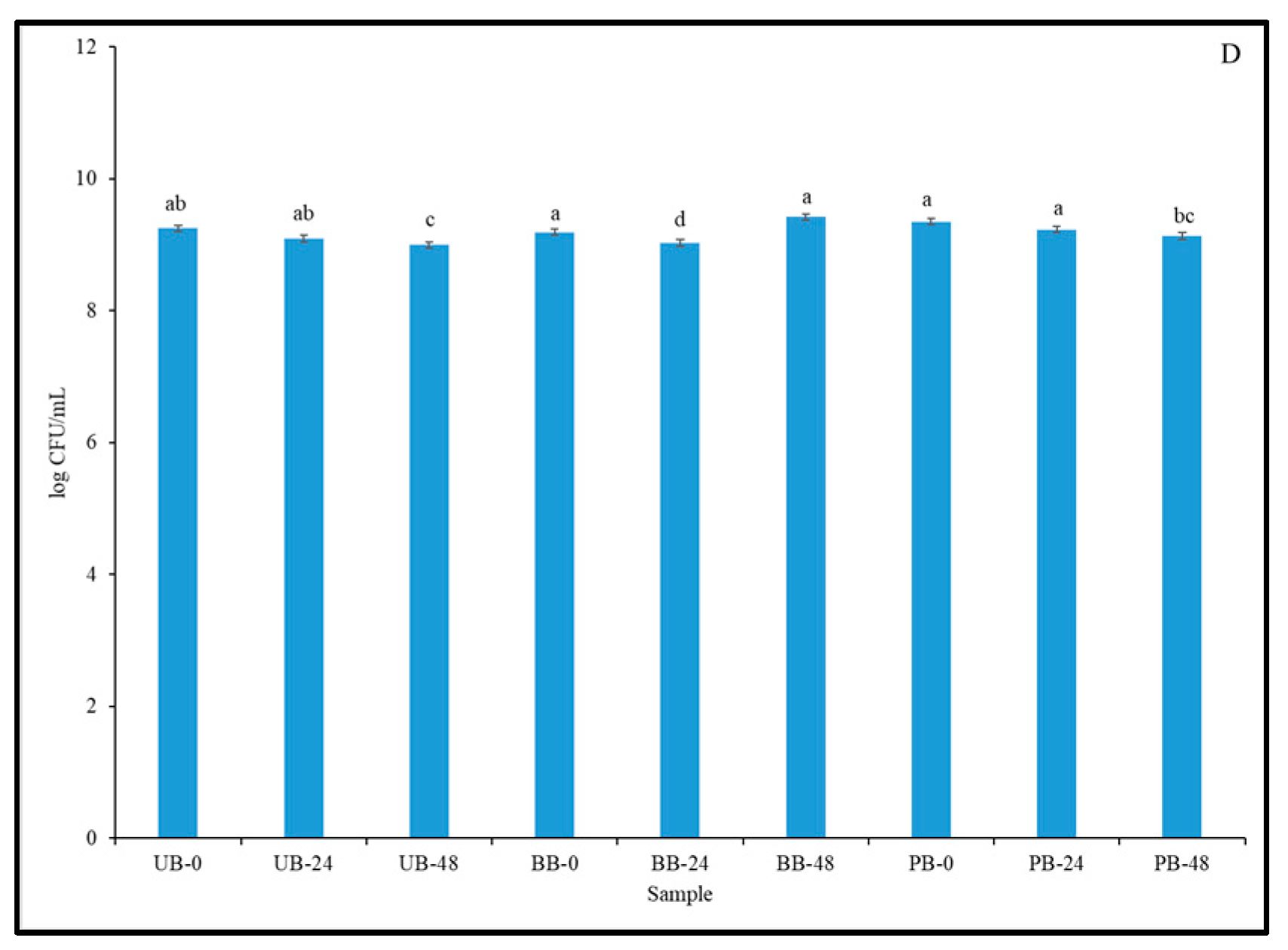
3.5. Viable Cell Count of LAB Strains in Fermented Barley Flour and Helicobacter Pylori Inhibition
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fuller, R. Probiotics in man and animals. J. Appl. Bacteriol. 1989, 66, 365–378. [Google Scholar] [CrossRef]
- Havenaar, R.; Brink, B.T.; Veld, J.H.J.H.I. Selection of strains for probiotic use. In Probiotics—The Scientific Basis; Fuller, R., Ed.; Springer: Amsterdam, The Netherlands, 1992; pp. 209–224. [Google Scholar] [CrossRef]
- Alander, M.; De Smet, I.; Nollet, L.; Verstraete, W.; von Wright, A.; Mattila-Sandholm, T. The effect of probiotic strains on the microbiota of the Simulator of the Human Intestinal Microbial Ecosystem (SHIME). Int. J. Food Microbiol. 1999, 46, 71–79. [Google Scholar] [CrossRef]
- Shori, A.B. Influence of food matrix on the viability of probiotic bacteria: A review based on dairy and non-dairy beverages. Food Biosci. 2016, 13, 1–8. [Google Scholar] [CrossRef]
- Aspri, M.; Papademas, P.; Tsaltas, D. Review on Non-Dairy Probiotics and Their Use in Non-Dairy Based Products. Fermentation 2020, 6, 30. [Google Scholar] [CrossRef] [Green Version]
- Bansal, S.; Mangal, M.; Sharma, S.K.; Gupta, R.K. Non-dairy Based Probiotics: A Healthy Treat for Intestine. Crit. Rev. Food Sci. Nutr. 2014, 56, 1856–1867. [Google Scholar] [CrossRef] [PubMed]
- Panghal, A.; Janghu, S.; Virkar, K.; Gat, Y.; Kumar, V.; Chhikara, N. Potential non-dairy probiotic products—A healthy approach. Food Biosci. 2018, 21, 80–89. [Google Scholar] [CrossRef]
- Charalampopoulos, D.; Wang, R.; Pandiella, S.; Webb, C. Application of cereals and cereal components in functional foods: A review. Int. J. Food Microbiol. 2002, 79, 131–141. [Google Scholar] [CrossRef]
- Sharma, R.; Mokhtari, S.; Jafari, S.M.; Sharma, S. Barley-based probiotic food mixture: Health effects and future prospects. Crit. Rev. Food Sci. Nutr. 2021, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Holtekjølen, A.K.; Kinitz, C.; Knutsen, S.H. Flavanol and Bound Phenolic Acid Contents in Different Barley Varieties. J. Agric. Food Chem. 2006, 54, 2253–2260. [Google Scholar] [CrossRef] [PubMed]
- Dykes, L. Phenolic Compounds in Cereal Grains and Their Health Benefits. Cereal Foods World 2007, 52, 105–111. [Google Scholar] [CrossRef]
- Okarter, N.; Liu, R.H. Health Benefits of Whole Grain Phytochemicals. Crit. Rev. Food Sci. Nutr. 2010, 50, 193–208. [Google Scholar] [CrossRef]
- Van Hung, P. Phenolic Compounds of Cereals and Their Antioxidant Capacity. Crit. Rev. Food Sci. Nutr. 2014, 56, 25–35. [Google Scholar] [CrossRef]
- Idehen, E.; Tang, Y.; Sang, S. Bioactive phytochemicals in barley. J. Food Drug Anal. 2016, 25, 148–161. [Google Scholar] [CrossRef] [Green Version]
- Ramakrishna, R.; Sarkar, D.; Shetty, K. Functional Bioactives from Barley for Human Health Benefits. In Functional Foods and Biotechnology; Shetty, K., Sarkar, D., Eds.; CRC Press: Boca Raton, FL, USA, 2019; pp. 61–85. [Google Scholar] [CrossRef]
- Ha, K.-S.; Jo, S.-H.; Mannam, V.; Kwon, Y.-I.; Apostolidis, E. Stimulation of Phenolics, Antioxidant and α-Glucosidase Inhibitory Activities During Barley (Hordeum vulgare L.) Seed Germination. Plant. Foods Hum. Nutr. 2016, 71, 211–217. [Google Scholar] [CrossRef]
- Ramakrishna, R.; Sarkar, D.; Schwarz, P.; Shetty, K. Phenolic linked anti-hyperglycemic bioactives of barley (Hordeum vulgare L.) cultivars as nutraceuticals targeting type 2 diabetes. Ind. Crop Prod. 2017, 107, 509–517. [Google Scholar] [CrossRef]
- Ramakrishna, R.; Sarkar, D.; Manduri, A.; Iyer, S.G.; Shetty, K. Improving phenolic bioactive-linked anti-hyperglycemic functions of dark germinated barley sprouts (Hordeum vulgare L.) using seed elicitation strategy. J. Food Sci. Technol. 2017, 54, 3666–3678. [Google Scholar] [CrossRef] [PubMed]
- Belobrajdic, D.P.; Bird, A.R. The potential role of phytochemicals in wholegrain cereals for the prevention of type-2 diabetes. Nutr. J. 2013, 12, 1–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panhwar, R.B.; Akbar, A.; Ali, M.F.; Yang, Q.; Feng, B. Phytochemical Components of Some Minor Cereals Associated with Diabetes Prevention and Management. J. Biosci. Med. 2018, 06, 9–22. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; He, F.; Chen, G. Improving bioaccessibility and bioavailability of phenolic compounds in cereal grains through processing technologies: A concise review. J. Funct. Foods 2014, 7, 101–111. [Google Scholar] [CrossRef]
- Donkor, O.; Stojanovska, L.; Ginn, P.; Ashton, J.; Vasiljevic, T. Germinated grains—Sources of bioactive compounds. Food Chem. 2012, 135, 950–959. [Google Scholar] [CrossRef]
- Singh, A.; Sharma, S. Bioactive components and functional properties of biologically activated cereal grains: A bibliographic review. Crit. Rev. Food Sci. Nutr. 2015, 57, 3051–3071. [Google Scholar] [CrossRef]
- Vitaglione, P.; Napolitano, A.; Fogliano, V. Cereal dietary fibre: A natural functional ingredient to deliver phenolic compounds into the gut. Trends Food Sci. Technol. 2008, 19, 451–463. [Google Scholar] [CrossRef]
- Espín, J.C.; González-Sarrías, A.; Tomás-Barberán, F.A. The gut microbiota: A key factor in the therapeutic effects of (poly)phenols. Biochem. Pharmacol. 2017, 139, 82–93. [Google Scholar] [CrossRef] [PubMed]
- Kanauchi, O.; Mitsuyama, K.; Andoh, A.; Iwanaga, T. Modulation of intestinal environment by prebiotic germinated barley foodstuff prevents chemo-induced colonic carcinogenesis in rats. Oncol. Rep. 1994, 20, 793–801. [Google Scholar] [CrossRef] [Green Version]
- Bamba, T.; Kanauchi, O.; Andoh, A.; Fujiyama, Y. A new prebiotic from germinated barley for nutraceutical treatment of ulcerative colitis. J. Gastroenterol. Hepatol. 2002, 17, 818–824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slavin, J. Whole Grains and Digestive Health. Cereal Chem. J. 2010, 87, 292–296. [Google Scholar] [CrossRef]
- Ullrich, S.E. Barley: Production, Improvement, and Uses; Wiley-Blackwell: Oxford, UK, 2011. [Google Scholar]
- Newman, R.K.; Newman, C.W. Barley for Food and Health: Science, Technology, and Products; John Wiley and Sons: Hoboken, NJ, USA, 2008. [Google Scholar]
- Nkhata, S.G.; Ayua, E.; Kamau, E.H.; Shingiro, J.-B. Fermentation and germination improve nutritional value of cereals and legumes through activation of endogenous enzymes. Food Sci. Nutr. 2018, 6, 2446–2458. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Jiménez, J.; Díaz-Rubio, M.E.; Calixto, F.D.S. Non-extractable polyphenols, a major dietary antioxidant: Occurrence, metabolic fate and health effects. Nutr. Res. Rev. 2013, 26, 118–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- E El Hag, M.; El Tinay, A.H.; E Yousif, N. Effect of fermentation and dehulling on starch, total polyphenols, phytic acid content and in vitro protein digestibility of pearl millet. Food Chem. 2002, 77, 193–196. [Google Scholar] [CrossRef]
- Anson, N.M.; Havenaar, R.; Vaes, W.; Coulier, L.; Venema, K.; Selinheimo, E.; Bast, A.; Haenen, G.R. Effect of bioprocessing of wheat bran in wholemeal wheat breads on the colonic SCFA production in vitro and postprandial plasma concentrations in men. Food Chem. 2011, 128, 404–409. [Google Scholar] [CrossRef] [PubMed]
- Mårtensson, O.; Öste, R.; Holst, O. The effect of yoghurt culture on the survival of probiotic bacteria in oat-based, non-dairy products. Food Res. Int. 2002, 35, 775–784. [Google Scholar] [CrossRef]
- McCue, P.; Kwon, Y.-I.; Shetty, K. Anti-amylase, anti-glucosidase and anti-angiotensin i-converting enzyme potential of selected foods. J. Food Biochem. 2005, 29, 278–294. [Google Scholar] [CrossRef]
- Kwon MS, Y.I.I.; Vattem, D.A.; Shetty, K. Evaluation of clonal herbs of lamiaceae species for management of diabetes and hypertension. Asia Pac. J. Clin. Nutr. 2006, 15, 107–118. [Google Scholar]
- Fujita, A.; Sarkar, D.; Genovese, M.I.; Shetty, K. Improving anti-hyperglycemic and anti-hypertensive properties of camu-camu (Myriciaria dubia Mc. Vaugh) using lactic acid bacterial fermentation. Process. Biochem. 2017, 59, 133–140. [Google Scholar] [CrossRef]
- Shetty, K.; Curtis, O.F.; Levin, R.E.; Witkowsky, R.; Ang, W. Prevention of vitrification associated with in vitro shoot culture of Oregano. (Origanum vulgare) by Pseudomonas spp. J. Plant Physiol. 1995, 147, 447–451. [Google Scholar] [CrossRef]
- Cervato, G.; Carabelli, M.; Gervasio, S.; Cittera, A.; Cazzola, R.; Cestaro, B. Antioxidant properties of oregano (Origanum vulgare) leaf extracts. J. Food Biochem. 2000, 24, 453–465. [Google Scholar] [CrossRef]
- Kwon, Y.-I.; Apostolidis, E.; Kim, Y.-C.; Shetty, K. Health Benefits of Traditional Corn, Beans, and Pumpkin: In Vitro Studies for Hyperglycemia and Hypertension Management. J. Med. Food 2007, 10, 266–275. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- McCue, P.P.; Shetty, K. Phenolic antioxidant mobilization during yogurt production from soymilk using Kefir cultures. Process. Biochem. 2005, 40, 1791–1797. [Google Scholar] [CrossRef]
- Stevenson, T.H.; Lucia, L.M.; Acuff, G.R. Development of a Selective Medium for Isolation of Helicobacter pylori from Cattle and Beef Samples. Appl. Environ. Microbiol. 2000, 66, 723–727. [Google Scholar] [CrossRef] [Green Version]
- Ranilla, L.G.; Huamán-Alvino, C.; Flores-Báez, O.; Aquino-Méndez, E.M.; Chirinos, R.; Campos, D.; Sevilla, R.; Fuentealba, C.; Pedreschi, R.; Sarkar, D.; et al. Evaluation of phenolic antioxidant-linked in vitro bioactivity of Peruvian corn (Zea mays L.) diversity targeting for potential management of hyperglycemia and obesity. J. Food Sci. Technol. 2019, 56, 2909–2924. [Google Scholar] [CrossRef]
- Sarkar, D.; Ankolekar, C.; Shetty, K. Beneficial Lactic Acid Bacteria (LAB)-Based Biotransformation of Plant and Dairy Substrates to Enhance Type 2 Diabetes-Relevant Health Benefits. In Functional Foods and Biotechnology: Biotransformation and Analysis of Functional Foods and Ingredients; Shetty, K., Sarkar, D., Eds.; CRC Press: Boca Raton, FL, USA, 2020; pp. 345–360. [Google Scholar] [CrossRef]
- Sánchez-Maldonado, A.; Schieber, A.; Gänzle, M. Structure-function relationships of the antibacterial activity of phenolic acids and their metabolism by lactic acid bacteria. J. Appl. Microbiol. 2011, 111, 1176–1184. [Google Scholar] [CrossRef]
- Curiel, J.A.; Rodríguez, H.; Landete, J.M.; Rivas, B.D.L.; Muñoz, R. Ability of Lactobacillus brevis strains to degrade food phenolic acids. Food Chem. 2010, 120, 225–229. [Google Scholar] [CrossRef] [Green Version]
- Svensson, L.; Sekwati-Monang, B.; Lutz, D.L.; Schieber, A.; Gänzle, M. Phenolic Acids and Flavonoids in Nonfermented and Fermented Red Sorghum (Sorghum bicolor (L.) Moench). J. Agric. Food Chem. 2010, 58, 9214–9220. [Google Scholar] [CrossRef] [PubMed]
- Hole, A.S.; Rud, I.; Grimmer, S.; Sigl, S.; Narvhus, J.; Sahlstrøm, S. Improved Bioavailability of Dietary Phenolic Acids in Whole Grain Barley and Oat Groat following Fermentation with Probiotic Lactobacillus acidophilus, Lactobacillus johnsonii, and Lactobacillus reuteri. J. Agric. Food Chem. 2012, 60, 6369–6375. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.-J.; Hyun, J.-N.; Kim, J.-A.; Park, J.-C.; Kim, M.-Y.; Kim, J.-G.; Lee, S.-J.; Chun, S.-C.; Chung, I.-M. Relationship between Phenolic Compounds, Anthocyanins Content and Antioxidant Activity in Colored Barley Germplasm. J. Agric. Food Chem. 2007, 55, 4802–4809. [Google Scholar] [CrossRef] [PubMed]
- Ge, X.; Jing, L.; Zhao, K.; Su, C.; Zhang, B.; Zhang, Q.; Han, L.; Yu, X.; Li, W. The phenolic compounds profile, quantitative analysis and antioxidant activity of four naked barley grains with different color. Food Chem. 2020, 335, 127655. [Google Scholar] [CrossRef] [PubMed]
- Rice-Evans, C.; Miller, N.; Paganga, G. Antioxidant properties of phenolic compounds. Trend Plant Sci. 1997, 2, 152–159. [Google Scholar] [CrossRef]
- Kähkönen, M.P.; Hopia, A.I.; Vuorela, H.J.; Rauha, J.-P.; Pihlaja, K.; Kujala, A.T.S.; Heinonen, M. Antioxidant Activity of Plant Extracts Containing Phenolic Compounds. J. Agric. Food Chem. 1999, 47, 3954–3962. [Google Scholar] [CrossRef] [PubMed]
- Balasundram, N.; Sundram, K.; Samman, S. Phenolic compounds in plants and agri-industrial by-products: Antioxidant activity, occurrence, and potential uses. Food Chem. 2006, 99, 191–203. [Google Scholar] [CrossRef]
- Sarkar, D.; Shetty, K. Metabolic Stimulation of Plant Phenolics for Food Preservation and Health. Annu. Rev. Food Sci. Technol. 2014, 5, 395–413. [Google Scholar] [CrossRef]
- Shahidi, F.; Ambigaipalan, P. Phenolics and polyphenolics in foods, beverages and spices: Antioxidant activity and health effects—A review. J. Funct. Food 2015, 18, 820–897. [Google Scholar] [CrossRef]
- Adebo, O.A.; Medina-Meza, I.G. Impact of Fermentation on the Phenolic Compounds and Antioxidant Activity of Whole Cereal Grains: A Mini Review. Molecules 2020, 25, 927. [Google Scholar] [CrossRef] [Green Version]
- Đorđević, T.M.; Šiler-Marinković, S.S.; Dimitrijevic-Brankovic, S. Effect of fermentation on antioxidant properties of some cereals and pseudo cereals. Food Chem. 2010, 119, 957–963. [Google Scholar] [CrossRef]
- Liu, Q.; Tang, G.-Y.; Zhao, C.-N.; Gan, R.-Y.; Li, H.-B. Antioxidant Activities, Phenolic Profiles, and Organic Acid Contents of Fruit Vinegars. Antioxidants 2019, 8, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Güzel-Seydim, Z.B.; Seydim, A.C.; Greene, A.; Bodine, A. Determination of Organic Acids and Volatile Flavor Substances in Kefir during Fermentation. J. Food Compos. Anal. 2000, 13, 35–43. [Google Scholar] [CrossRef]
- Shalaby, E.A.; Shanab, S.M.M. Comparison of DPPH and ABTS assays for determining antioxidant potential of water and methanol extracts of Spirulina platensis. Ind. J. Mar. Sci. 2013, 42, 556–564. [Google Scholar]
- Morón, E.B.; Abad-Jiménez, Z.; De Marañón, A.M.; Iannantuoni, F.; López, E.; López-Domènech, S.; Salom, C.; Jover, A.; Mora, V.; Roldan, I.; et al. Relationship Between Oxidative Stress, ER Stress, and Inflammation in Type 2 Diabetes: The Battle Continues. J. Clin. Med. 2019, 8, 1385. [Google Scholar] [CrossRef] [Green Version]
- McCue, P.; Shetty, K. Health Benefits of Soy Isoflavonoids and Strategies for Enhancement: A Review. Crit. Rev. Food Sci. Nutr. 2004, 44, 361–367. [Google Scholar] [CrossRef]
- Ansari, P.; Afroz, N.; Jalil, S.; Azad, S.B.; Mustakim, M.G.; Anwar, S.; Haque, S.N.; Hossain, S.M.; Tony, R.R.; Hannan, J.M.A. Anti-hyperglycemic activity of Aegle marmelos (L.) Corr. is partly mediated by increased insulin secretion, α-amylase inhibition, and retardation of glucose absorption. J. Pediatr. Endocrinol. Metab. 2017, 30, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Ankolekar, C.; Pinto, M.; Greene, D.; Shetty, K. In vitro bioassay based screening of antihyperglycemia and antihypertensive activities of Lactobacillus acidophilus fermented pear juice. Innov. Food Sci. Emerg. Technol. 2012, 13, 221–230. [Google Scholar] [CrossRef]
- Ag, H.B.B. Pharmacology of α-glucosidase inhibition. Eur. J. Clin. Investig. 2010, 24, 3–10. [Google Scholar] [CrossRef]
- Rosenstock, J.; Brown, A.; Fischer, J.; Jain, A.; Littlejohn, T.; Nadeau, D.; Sussman, A.; Taylor, T.; Krol, A.; Magner, J. Efficacy and Safety of Acarbose in Metformin-Treated Patients with Type 2 Diabetes. Diabetes Care 1998, 21, 2050–2055. [Google Scholar] [CrossRef]
- Hedrington, M.S.; Davis, S.N. Considerations when using alpha-glucosidase inhibitors in the treatment of type 2 diabetes. Expert Opin. Pharmacother. 2019, 20, 2229–2235. [Google Scholar] [CrossRef]
- Kandylis, P.; Pissaridi, K.; Bekatorou, A.; Kanellaki, M.; A Koutinas, A. Dairy and non-dairy probiotic beverages. Curr. Opin. Food Sci. 2016, 7, 58–63. [Google Scholar] [CrossRef]
- Enujiugha, V.N.; Badejo, A.A. Probiotic potentials of cereal-based beverages. Crit. Rev. Food Sci. Nutr. 2015, 57, 790–804. [Google Scholar] [CrossRef] [PubMed]
- Mridula, D.; Sharma, M. Development of non-dairy probiotic drink utilizing sprouted cereals, legume and soymilk. LWT 2015, 62, 482–487. [Google Scholar] [CrossRef]
- Sharma, M.; Mridula, D.; Gupta, R.K. Development of sprouted wheat based probiotic beverage. J. Food Sci. Technol. 2013, 51, 3926–3933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chavan, M.; Gat, Y.; Harmalkar, M.; Waghmare, R. Development of non-dairy fermented probiotic drink based on germinated and ungerminated cereals and legume. LWT 2018, 91, 339–344. [Google Scholar] [CrossRef]
- Midolo, P.D.; Lambert, J.R.; Hull, R.; Luo, F.; Grayson, M.L. In vitro inhibition of Helicobacter pylori NCTC 11637 by organic acids and lactic acid bacteria. J. Appl. Bacteriol. 1995, 79, 475–479. [Google Scholar] [CrossRef]
- Alakomi, H.-L.; Skyttä, E.; Saarela, M.; Mattila-Sandholm, T.; Latva-Kala, K.; Helander, I.M. Lactic Acid Permeabilizes Gram-Negative Bacteria by Disrupting the Outer Membrane. Appl. Environ. Microbiol. 2000, 66, 2001–2005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chun, S.-S.; Vattem, D.A.; Lin, Y.-T.; Shetty, K. Phenolic antioxidants from clonal oregano (Origanum vulgare) with antimicrobial activity against Helicobacter pylori. Process. Biochem. 2005, 40, 809–816. [Google Scholar] [CrossRef]
- Ankolekar, C.; Johnson, D.; Pinto, M.D.S.; Johnson, K.; Labbe, R.; Shetty, K. Inhibitory Potential of Tea Polyphenolics and Influence of Extraction Time Against Helicobacter pylori and Lack of Inhibition of Beneficial Lactic Acid Bacteria. J. Med. Food 2011, 14, 1321–1329. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (A) | ||||||||||
| Source | df | TSP † | ABTS | DPPH | α-Amylase Inhibition | α-Glucosidase Inhibition | ||||
| Undiluted | Half Diluted | One-Fifth Diluted | Undiluted | Half Diluted | One-Fifth Diluted | |||||
| P | 2 | 3.23 * | 9956.1 * | 1268.2 * | 2996.0 * | 9551.5 * | 1467.8 * | 4811.7 * | 2650.3 * | 951.4 * |
| S | 2 | 1.26 * | 4904.4 * | 48,742.0 * | 1816.1 * | 1174.4 * | 2739.9 * | 542.5 * | 224.0 * | 108.9 * |
| P × S | 4 | 0.01 ns | 0.81 ns | 264.9 * | 0.92 ns | 0.30 ns | 2.93 ns | 45.5 * | 36.0 * | 58.6 * |
| G | 2 | 3.92 * | 6770.5 * | 1905.9 * | 598.9 * | 11,744.0 * | 11,294.0 * | 5163.0 * | 3104.6 * | 2449.9 * |
| P × G | 4 | 0.47 * | 1695.0 * | 199.0 * | 4085.2 * | 8834.3 * | 2861.2 * | 1566.6 * | 785.1 * | 580.4 * |
| S × G | 4 | 0.00 ns | 1.22 ns | 32.2 ns | 1.57 ns | 0.82 ns | 8.86 ns | 136.8 * | 67.9 * | 16.7 ** |
| P × S × G | 8 | 0.00 ns | 3.04 ns | 84.15 * | 3.99 ns | 3.00 ns | 5.84 ns | 72.4 * | 59.3 * | 33.6 * |
| (B) | ||||||||||
| Source | df | TSP | ABTS | DPPH | α-Amylase Inhibition | α-Glucosidase Inhibition | ||||
| Undiluted | Half Diluted | One-Fifth Diluted | Undiluted | Half Diluted | One-Fifth Diluted | |||||
| P | 2 | 8.69 * | 7018.9 * | 1746.7 * | 3851.8 * | 1874.6 * | 1268.2 * | 1653.9 * | 795.7 * | 412.5 * |
| S | 2 | 1.19 * | 2986.5 * | 44,174.0 * | 2036.71 * | 1207.8 * | 3109.4 * | 228.1 * | 76.0 * | 19.8 ** |
| P × S | 4 | 0.00 ns | 2720.7 * | 633.0 * | 6.71 ns | 8.51 ns | 0.63 ns | 9.51 ns | 2.56 ns | 5.05 ns |
| G | 2 | 10.1 * | 639.0 ** | 1095.2 * | 2642. * | 2500.6 ** | 429.9 * | 3188.5 * | 2530.1 * | 917 * |
| P × G | 4 | 2.25 * | 670.0 * | 245.5 * | 2777.7 * | 1724.7 ns | 206.3 * | 3463.0 * | 859.9 * | 289.6 * |
| S × G | 4 | 0.00 ns | 87.6 ns | 194.7 * | 10.4 ** | 5.39 ns | 1.57 ns | 1.65 ns | 0.80 ns | 6.05 ns |
| P × S × G | 8 | 0.00 ns | 548.7 ns | 12.1 ns | 8.49 ** | 3.18 ns | 4.39 ns | 6.66 ns | 6.09 ns | 2.29 ns |
| (C) | ||||||||||
| Source | df | TSP | ABTS | DPPH | α-Amylase Inhibition | α-Glucosidase Inhibition | ||||
| Undiluted | Half Diluted | One-Fifth Diluted | Undiluted | Half Diluted | One-Fifth Diluted | |||||
| P | 2 | 1.17 * | 8643.1 * | 673.8 * | 3804.2 * | 8505.2 * | 3395.8 * | 12,400.0 * | 4075.4 * | 721.6 * |
| S | 2 | 1.32 * | 2016.4 * | 51,148.0 * | 1936.0 * | 1307.1 * | 2827.0 * | 191.4 * | 51.4 * | 17.2 ** |
| P × S | 4 | 0.00 ns | 724.6 * | 362.9 * | 4.34 ns | 3.77 ns | 3.93 ns | 1.63 ns | 1.72 ns | 2.60 ns |
| G | 2 | 12.28 * | 22.4 ns | 427.1 * | 5666.6 * | 3249.2 * | 2003.0 * | 3279.1 * | 2183.5 * | 1090.1 * |
| P × G | 4 | 0.87 * | 793.8 * | 371.2 * | 3067.2 * | 2536.6 * | 1993.6 * | 710.0 * | 411.4 * | 65.0 * |
| S × G | 4 | 0.00 ns | 1.39 ns | 232.3 * | 2.68 ns | 3.70 ns | 5.24 ns | 7.62 ns | 1.45 ns | 3.00 ns |
| P × S × G | 8 | 0.00 ns | 6.07 ns | 68.7 * | 5.66 ns | 4.52 ns | 4.12 ns | 4.21 ns | 2.98 ns | 4.14 ns |
| (D) | ||||||||||
| Source | df | TSP | ABTS | DPPH | α-Amylase Inhibition | α-Glucosidase Inhibition | ||||
| Undiluted | Half Diluted | One-fifth Diluted | Undiluted | Half Diluted | One-Fifth Diluted | |||||
| P | 2 | 0.47 * | 18762.0 * | 2822.7 * | 4156.5 * | 70,172.0 * | 6889.8 * | 6749.5 * | 1567.6 * | 480.3 * |
| S | 2 | 1.27 * | 2602.7 * | 35,911.0 * | 1080.5 * | 1798.1 * | 2778.4 * | 197.5 * | 35.6 * | 3.31 ns |
| P × S | 4 | 0.00 ns | 404.6 * | 785.2 * | 189.9 * | 8.45 ** | 10.9 ** | 7.16 ns | 8.63 ns | 11.5 ** |
| G | 2 | 17.2 * | 509.3 * | 404.7 * | 668.3 * | 5729.4 * | 1277.4 * | 2361.5 * | 1435.1 * | 437.5 * |
| P × G | 4 | 2.65 * | 2064.8 * | 596.3 * | 343.3 * | 2104.3 * | 268.2 * | 565.3 * | 349.7 * | 65.63 * |
| S × G | 4 | 0.00 ns | 24.6 ns | 63.3 * | 28.3 * | 43.6 * | 2.56 ns | 0.80 ns | 2.07 ns | 0.55 ns |
| P × S × G | 8 | 0.00 ns | 13.2 ns | 52.8 * | 27.0 * | 36.4 * | 2.54 ns | 2.09 ns | 7.52 ns | 4.66 ns |
| (A) | |||||||||||||
| Food Barley Type | Germination Time (h) | Dihydroxybenzoic Acid | Catechin | Cinnamic Acid | Protocatechuic Acid | Gallic Acid | Benzoic Acid | ||||||
| C | F | C | F | C | F | C | F | C | F | C | F | ||
| UB | 0 | 0.33 ± 0.00 | 0.37 ± 0.00 | 0.17 ± 0.01 | 0.12 ± 0.00 | n.d. | n.d. | 2.11 ± 0.02 | 2.63 ± 0.01 | 1.28 ± 0.01 | 1.42 ± 0.01 | 0.04 ± 0.01 | 0.4 ± 0.21 |
| 24 | 0.59 ± 0.01 | 0.66 ± 0.00 | 0.22 ± 0.05 | 0.17 ± 0.04 | n.d. | n.d. | 2.49 ± 0.05 | 3.31 ± 0.01 | 1.71 ± 0.06 | 1.78 ± 0.01 | 0.2 ± 0.03 | 0.3 ± 0.05 | |
| 48 | 0.97 ± 0.00 | 0.95 ± 0.00 | 0.29 ± 0.01 | 0.27 ± 0.01 | n.d. | n.d. | 4.19 ± 0.01 | 4.43 ± 0.01 | 2.42 ± 0.03 | 2.98 ± 0.14 | 0.22 ± 0.04 | 0.37 ± 0.05 | |
| BB | 0 | 0.38 ± 0.00 | 0.6 ± 0.17 | 0.25 ± 0.02 | 0.14 ± 0.04 | 2.56 ± 0.04 | 11.6 ± 0.35 | 1.94 ± 0.48 | 3.69 ± 0.22 | 1.01 ± 0.00 | 1.62 ± 0.37 | n.d. | n.d. |
| 24 | 0.87 ± 0.01 | 1.05 ± 0.28 | 0.23 ± 0.01 | 0.25 ± 0.06 | 5.81 ± 0.04 | 13.9 ± 0.09 | 3.92 ± 0.03 | 5.99 ± 0.16 | 2.32 ± 0.12 | 2.79 ± 0.75 | n.d. | n.d. | |
| 48 | 1.55 ± 0.01 | 1.46 ± 0.01 | 0.3 ± 0.02 | 0.33 ± 0.01 | 6.56 ± 0.21 | 16.7 ± 0.03 | 7.4 ± 0.20 | 7.85 ± 0.05 | 3.68 ± 0.05 | 5.16 ± 0.02 | n.d. | n.d. | |
| PB | 0 | 0.2 ± 0.01 | 0.28 ± 0.00 | 0.3 ± 0.00 | 0.16 ± 0.01 | 2.67 ± 0.00 | 8.23 ± 0.07 | 2.1 ± 0.01 | 2.75 ± 0.02 | 2.14 ± 0.06 | 2.23 ± 0.01 | 0.04 ± 0.00 | 0.31 ± 0.08 |
| 24 | 0.84 ± 0.00 | 0.93 ± 0.01 | 0.26 ± 0.01 | 0.21 ± 0.01 | 5.34 ± 0.06 | 14.8 ± 0.17 | 4.18 ± 0.00 | 4.93 ± 0.06 | 2.99 ± 0.01 | 3.53 ± 0.04 | 0.02 ± 0.00 | 0.26 ± 0.01 | |
| 48 | 1.39 ± 0.01 | 1.32 ± 0.01 | 0.36 ± 0.09 | 0.55 ± 0.01 | 7.63 ± 0.28 | 23.1 ± 0.17 | 6.81 ± 0.04 | 7.69 ± 0.06 | 4.26 ± 0.03 | 4.95 ± 0.03 | 0.03 ± 0.01 | 0.32 ± 0.02 | |
| (B) | |||||||||||||
| Food Barley Type | Germination Time (h) | Dihydroxybenzoic Acid | Catechin | Cinnamic Acid | Protocatechuic Acid | Gallic Acid | |||||||
| C | F | C | F | C | F | C | F | C | F | ||||
| UB | 0 | 0.45 ± 0.00 | 0.56 ± 0.00 | 0.26 ± 0.01 | 0.18 ± 0.00 | 4.66 ± 0.01 | 12.6 ± 0.74 | 3.06 ± 0.01 | 5.98 ± 0.15 | 1.19 ± 0.05 | 2.35 ± 0.00 | ||
| 24 | 0.9 ± 0.01 | 0.87 ± 0.00 | 0.3 ± 0.10 | 0.29 ± 0.00 | 6.47 ± 0.33 | 13.5 ± 0.04 | 4.86 ± 0.02 | 7.4 ± 0.01 | 1.95 ± 0.58 | 2.54 ± 0.88 | |||
| 48 | 1.61 ± 0.00 | 1.35 ± 0.00 | 0.39 ± 0.02 | 0.45 ± 0.00 | 7.69 ± 0.03 | 20.3 ± 0.03 | 8.61 ± 0.52 | 11.0 ± 0.01 | 4.65 ± 0.01 | 5.49 ± 0.03 | |||
| BB | 0 | 0.74 ± 0.00 | 0.9 ± 0.00 | 0.19 ± 0.01 | 0.41 ± 0.01 | 5.88 ± 0.01 | 4.73 ± 0.01 | 1.32 ± 0.02 | 8.69 ± 0.01 | 0.97 ± 0.01 | 1.22 ± 0.00 | ||
| 24 | 1.44 ± 0.01 | 1.27 ± 0.00 | 0.32 ± 0.01 | 0.48 ± 0.01 | 5.47 ± 0.02 | 5.22 ±0.01 | 20.8 ± 0.83 | 10.5 ± 0.02 | 3.61 ± 0.04 | 1.83 ± 0.01 | |||
| 48 | 2.29 ± 0.01 | 1.97 ± 0.78 | 0.40 ±0.04 | 0.62 ± 0.16 | 7.09 ± 1.0 | 6.09 ± 0.01 | 13.6 ± 0.03 | 18.3 ± 0.02 | 6.93 ± 0.10 | 5.25 ± 0.07 | |||
| PB | 0 | 0.49 ± 0.00 | 0.39 ± 0.00 | 0.21 ± 0.03 | 0.31 ± 0.01 | 6.61 ± 0.18 | 2.93 ± 0.03 | 1.52 ± 0.04 | 5.56 ± 0.01 | 4.01 ± 0.03 | 1.51 ± 0.00 | ||
| 24 | 1.35 ± 0.01 | 1.27 ± 0.00 | 0.37 ± 0.01 | 0.29 ± 0.12 | 7.93 ± 0.04 | 5.04 ± 0.27 | 0.5 ± 0.14 | 11.0 ± 0.05 | 4.14 ± 0.63 | 3.62 ± 0.27 | |||
| 48 | 1.95 ±0.08 | 1.99 ± 0.00 | 0.68 ± 0.02 | 0.68 ± 0.18 | 7.36 ± 0.34 | 7.49 ± 0.50 | 2.18 ± 0.22 | 16.4 ± 0.12 | 6.70 ± 0.02 | 8.61 ± 0.03 | |||
| (C) | |||||||||||||
| Food Barley Type | Germination Time (h) | Dihydroxybenzoic Acid | Catechin | Cinnamic Acid | Protocatechuic Acid | Gallic Acid | |||||||
| C | F | C | F | C | F | C | F | C | F | ||||
| UB | 0 | 0.66 ± 0.00 | 0.65 ± 0.01 | 0.29 ± 0.02 | 0.64 ± 0.16 | 4.25 ± 0.02 | 4.48 ± 0.71 | 4.75 ± 0.01 | 8.33 ± 0.01 | 1.12 ± 0.00 | 1.56 ± 0.01 | ||
| 24 | 1.36 ± 0.02 | 0.87 ± 0.00 | 0.26 ± 0.08 | 0.33 ± 0.02 | 4.6 ± 0.04 | 4.44 ± 0.03 | 6.69 ± 0.15 | 10 ± 0.37 | 1.85 ± 0.01 | 1.93 ± 0.00 | |||
| 48 | 2.32 ± 0.01 | 1.44 ± 0.02 | 0.54 ± 0.06 | 0.28 ± 0.02 | 5.86 ± 0.06 | 5.93 ± 0.33 | 13.3 ± 0.01 | 14 ± 0.02 | 6.51 ± 0.00 | 3.1 ± 0.03 | |||
| BB | 0 | 1.08 ± 0.00 | 0.96 ± 0.01 | 0.24 ± 0.08 | 0.43 ± 0.03 | 0.84 ± 0.01 | 4.18 ± 0.05 | 0.15 ± 0.00 | 1.34 ± 0.00 | 0.99 ± 0.00 | 1.72 ± 0.00 | ||
| 24 | 1.75 ± 0.69 | 1.15 ± 0.01 | 0.36 ± 0.28 | 0.19 ± 0.01 | 0.77 ± 0.03 | 6.57 ± 0.04 | 0.09 ± 0.01 | 0.68 ± 0.00 | 0.43 ±0.00 | 2.54 ± 0.00 | |||
| 48 | 3.14 ± 0.01 | 2.38 ± 0.01 | 0.39 ± 0.18 | 0.26 ± 0.01 | 1.46 ± 0.09 | 7.22 ± 0.03 | 0.04 ± 0.00 | 1.13 ± 0.00 | 0.48 ± 0.05 | 4.73 ± 0.01 | |||
| PB | 0 | 0.79 ± 0.01 | 0.57 ± 0.03 | 0.26 ± 0.14 | 0.14 ± 0.01 | 0.44 ± 0.01 | 3.35 ± 0.04 | 1.82 ± 0.10 | 9.26 ± 0.01 | 1.86 ± 0.08 | 2.3 ± 0.00 | ||
| 24 | 1.78 ± 0.03 | 1.14 ± 0.01 | 1.12 ± 0.75 | 0.21 ± 0.01 | 2.65 ± 0.82 | 5.07 ± 0.03 | 1.61 ± 0.01 | 12.2 ± 0.01 | 5.5 ± 0.09 | 2.97 ±0.06 | |||
| 48 | 2.62 ± 0.20 | 2.11 ± 0.17 | 0.32 ± 0.23 | 0.36 ± 0.03 | 5.75 ± 0.89 | 7.54 ± 0.04 | 3.2 ± 0.81 | 18.7 ± 0.03 | 3.17 ± 0.14 | 9.2 ± 0.02 | |||
| (D) | |||||||||||||
| Food Barley Type | Germination Time (h) | Dihydroxybenzoic Acid | Catechin | Cinnamic Acid | Protocatechuic Acid | Gallic Acid | |||||||
| C | F | C | F | C | F | C | F | C | F | ||||
| UB | 0 | 0.92 ± 0.00 | 0.61 ± 0.03 | 0.3 ± 0.05 | 0.67 ± 0.03 | 19.1 ± 0.09 | 28.4 ± 0.14 | 0.64 ± 0.00 | 0.79 ± 0.10 | 0.74 ± 0.60 | 2.51 ± 0.23 | ||
| 24 | 1.25 ± 0.02 | 0.85 ± 0.03 | 0.21 ± 0.08 | 0.25 ± 0.00 | 20.9 ± 0.58 | 30.5 ± 0.13 | 0.7 ± 0.05 | 0.27 ± 0.00 | 2.46 ± 0.01 | 2.75 ± 0.02 | |||
| 48 | 2.7 ± 0.06 | 1.64 ± 0.07 | 0.1 ± 0.03 | 0.37 ± 0.02 | 48.1 ± 0.02 | 38.6 ± 0.18 | 0.16 ± 0.00 | 0.53 ± 0.07 | 7.3 ± 0.01 | 4.44 ± 0.19 | |||
| BB | 0 | 1.48 ± 0.01 | 1.07 ± 0.02 | 0.21 ± 0.01 | 0.19 ± 0.00 | 1.41 ± 0.90 | 1.96 ± 0.00 | 0.21 ± 0.00 | 0.79 ± 0.00 | 1.53 ± 0.00 | 3.09 ± 0.14 | ||
| 24 | 2.23 ± 0.04 | 133 ± 0.08 | 0.06 ± 0.00 | 0.23 ± 0.00 | 0.54 ± 0.01 | 3.57 ± 0.00 | 0.14 ± 0.00 | 1.55 ± 0.01 | 5.95 ± 0.08 | 3.76 ± 0.01 | |||
| 48 | 2.97 ± 0.02 | 2.76 ± 0.05 | 0.09 ± 0.00 | 0.15 ± 0.00 | 0.6 ± 0.01 | 3.6 ± 0.00 | 0.22 ± 0.00 | 2.21 ± 0.01 | 9.72 ± 0.01 | 6.9 ± 0.01 | |||
| PB | 0 | 0.82 ± 0.01 | 0.59 ± 0.01 | 0.24 ± 0.01 | 0.15 ± 0.00 | 0.2 ± 0.00 | 1.79 ± 0.01 | 4.59 ± 0.03 | 8.67 ± 0.01 | 2.21 ± 0.10 | 3.42 ± 0.01 | ||
| 24 | 2.27 ± 0.05 | 1.18 ± 0.02 | 0.44 ± 0.01 | 0.29 ± 0.01 | 0.45 ± 0.00 | 4.24 ± 0.09 | 4.53 ± 0.12 | 13.4 ± 0.01 | 6.73 ± 0.06 | 4.38 ± 0.13 | |||
| 48 | 3.48 ± 0.81 | 2.07 ± 0.03 | 0.09 ± 0.00 | 0.4 ± 0.08 | 1.27 ± 0.01 | 5.76 ± 0.12 | 16.6 ± 0.03 | 20.1 ± 0.06 | 9.02 ± 0.01 | 9.13 ± 0.12 | |||
| (A) | |||||||
| Source | df | Dihydroxybenzoic Acid | Catechin | Cinnamic Acid | Gallic Acid | Protocatechuic Acid | Benzoic Acid |
| P | 1 | 0.09 * | 0.01 * | 1110.1 ns | 7.23 * | 22.0 * | 0.66 * |
| S | 2 | 2.10 * | 0.18 * | 2365.3 * | 36.6 * | 75.9 * | 1.20 * |
| P × S | 2 | 0.03 | 0.01 * | 560.0 ns | 1.67 * | 3.86 * | 0.35 ** |
| G | 2 | 15.0 * | 0.5 ns | 417.1 * | 95.9 * | 270.8 * | 0.02 * |
| P × G | 2 | 0.17 * | 0.11 ** | 64.6 * | 2.11 * | 2.19 * | 0.04 ** |
| S × G | 4 | 0.95 * | 0.09 ns | 297.2 * | 9.56 * | 33.3 * | 0.03 * |
| P × S × G | 4 | 0.06 | 0.09 ** | 90.2 * | 0.68 * | 2.83 ** | 0.07 ns |
| (B) | |||||||
| Source | df | Dihydroxybenzoic Acid | Catechin | Cinnamic Acid | Gallic Acid | Protocatechuic Acid | |
| P | 1 | 0.14 | 0.12 * | 115.8 * | 0.99 ns | 22.0 * | |
| S | 2 | 4.16 * | 0.26 * | 57.0 * | 62.9 * | 75.9 * | |
| P × S | 2 | 0.02 | 0.24 * | 687.2 * | 17.2 * | 3.86 ** | |
| G | 2 | 29.3 * | 1.42 * | 179.7 * | 378.1 * | 270.8 * | |
| P × G | 2 | 0.24 | 0.02 ns | 45.7 * | 4.28 ** | 2.19 ** | |
| S × G | 4 | 1.19 * | 0.27 * | 63.8 * | 11.8 * | 33.3 * | |
| P × S × G | 4 | 0.33 | 0.07 ** | 33.8 * | 33.3 * | 2.83 ns | |
| (C) | |||||||
| Source | df | Dihydroxybenzoic Acid | Catechin | Cinnamic Acid | Gallic Acid | Protocatechuic Acid | |
| P | 1 | 5.97 * | 2.76 * | 517.7 * | 22.2 * | 644.4 * | |
| S | 2 | 4.99 * | 1.62 * | 115.6 * | 101.9 * | 1619.5 * | |
| P × S | 2 | 0.01 | 6.85 * | 343.8 ** | 52.0 * | 544.2 * | |
| G | 2 | 44.5 * | 1.05 * | 421.1 * | 162.3 * | 335.3 * | |
| P × G | 2 | 1.81 * | 9.22 * | 4.22 ** | 27.8 * | 13.0 * | |
| S × G | 4 | 1.55 * | 13.2 * | 130.9 * | 33.6 * | 181.6 * | |
| P × S × G | 4 | 0.31 | 12.0 * | 40.0 * | 131.6 * | 103.0 * | |
| (D) | |||||||
| Source | df | Dihydroxybenzoic Acid | Catechin | Cinnamic Acid | Gallic Acid | Protocatechuic Acid | |
| P | 1 | 12.1 * | 0.31 * | 221.7 * | 9.35 * | 140.8 * | |
| S | 2 | 7.62 * | 0.49 * | 1995.0 * | 115.2 * | 2716.3 * | |
| P × S | 2 | 0.80 * | 0.21 * | 6.23 ns | 4.28 * | 146.8 * | |
| G | 2 | 52.8 * | 0.17 * | 1171.6 * | 55.6 * | 325.1 * | |
| P × G | 2 | 1.67 * | 0.18 * | 180.4 * | 60.3 * | 14.3 * | |
| S × G | 4 | 2.07 * | 0.59 * | 1671.8 * | 17.3 * | 601.7 * | |
| P × S × G | 4 | 2.28 * | 0.44 * | 560.1 * | 26.7 * | 42.0 * | |
| Sample | Duration of Fermentation (h) | |||
|---|---|---|---|---|
| 0 | 24 | 48 | 72 | |
| UB-0 | n.d. | n.d. | n.d. | n.d. |
| UB-24 | n.d. | 12.9 | 12.9 | 13.0 |
| UB-48 | 13.8 | 13.1 | 14.0 | 14.2 |
| BB-0 | n.d. | n.d. | n.d. | n.d. |
| BB-24 | n.d. | 12.9 | 13.1 | 13.0 |
| BB-48 | 12.9 | 13.0 | 13.3 | 13.2 |
| PB-0 | n.d. | n.d. | n.d. | n.d. |
| PB-24 | 13.0 | 13.2 | 13.1 | 14.0 |
| PB-48 | 14.3 | 15.1 | 15.8 | 16.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramakrishna, R.; Sarkar, D.; Dogramaci, M.; Shetty, K. Kefir Culture-Mediated Fermentation to Improve Phenolic-Linked Antioxidant, Anti-Hyperglycemic and Human Gut Health Benefits in Sprouted Food Barley. Appl. Microbiol. 2021, 1, 377-407. https://doi.org/10.3390/applmicrobiol1020026
Ramakrishna R, Sarkar D, Dogramaci M, Shetty K. Kefir Culture-Mediated Fermentation to Improve Phenolic-Linked Antioxidant, Anti-Hyperglycemic and Human Gut Health Benefits in Sprouted Food Barley. Applied Microbiology. 2021; 1(2):377-407. https://doi.org/10.3390/applmicrobiol1020026
Chicago/Turabian StyleRamakrishna, Ramnarain, Dipayan Sarkar, Munevver Dogramaci, and Kalidas Shetty. 2021. "Kefir Culture-Mediated Fermentation to Improve Phenolic-Linked Antioxidant, Anti-Hyperglycemic and Human Gut Health Benefits in Sprouted Food Barley" Applied Microbiology 1, no. 2: 377-407. https://doi.org/10.3390/applmicrobiol1020026
APA StyleRamakrishna, R., Sarkar, D., Dogramaci, M., & Shetty, K. (2021). Kefir Culture-Mediated Fermentation to Improve Phenolic-Linked Antioxidant, Anti-Hyperglycemic and Human Gut Health Benefits in Sprouted Food Barley. Applied Microbiology, 1(2), 377-407. https://doi.org/10.3390/applmicrobiol1020026