
Mapping Selected Emergent Marine Toxin-Producing Organisms Using Historical Samples with Two Methods (Biosensors and Real-Time PCR): A Comparison of Resolution


Abstract
1. Introduction
2. Materials and Methods
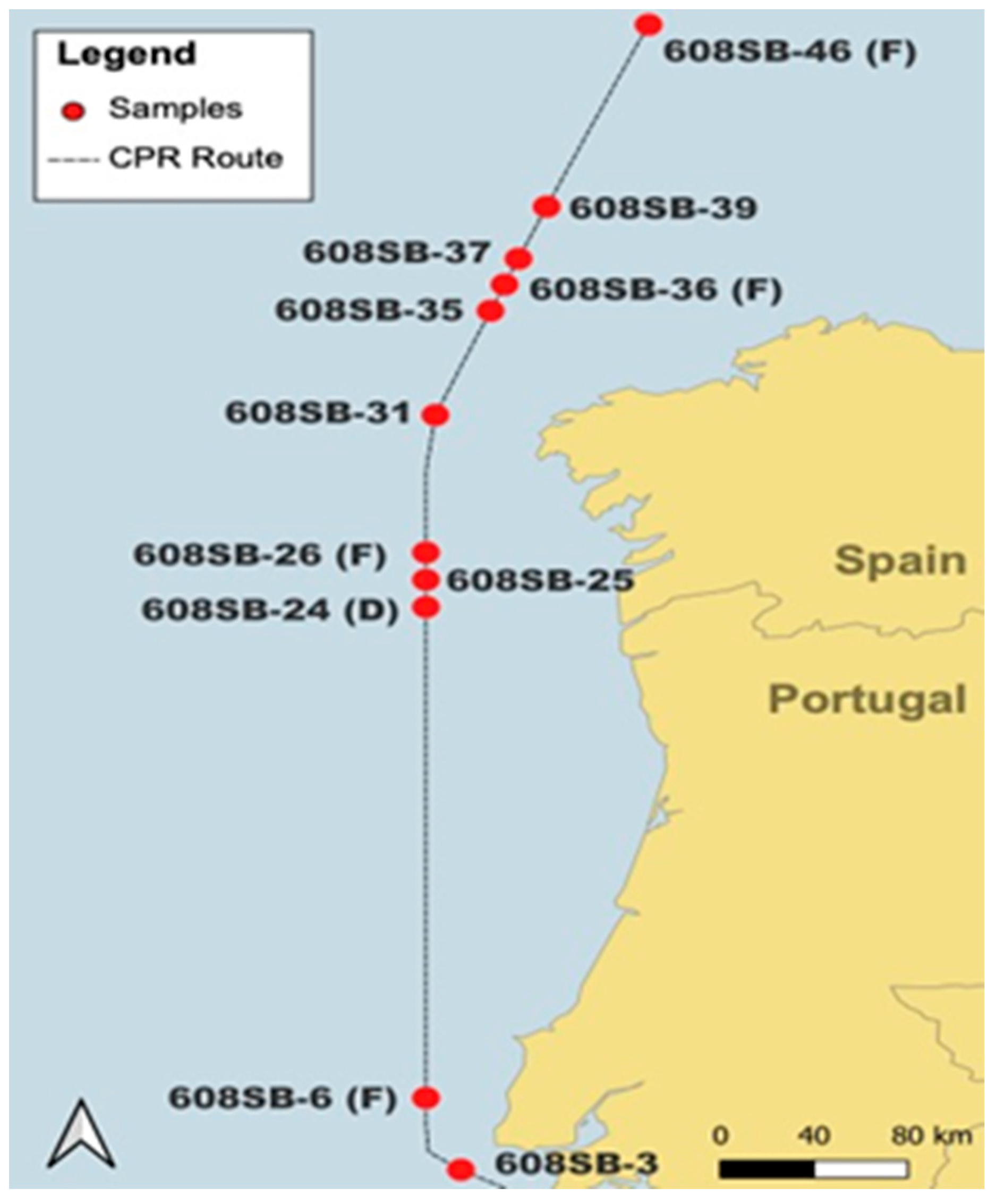
2.1. Sample Collection
2.2. Extraction of DNA from CPR Samples
2.3. PCR Reactions
2.4. Probe Hybridization
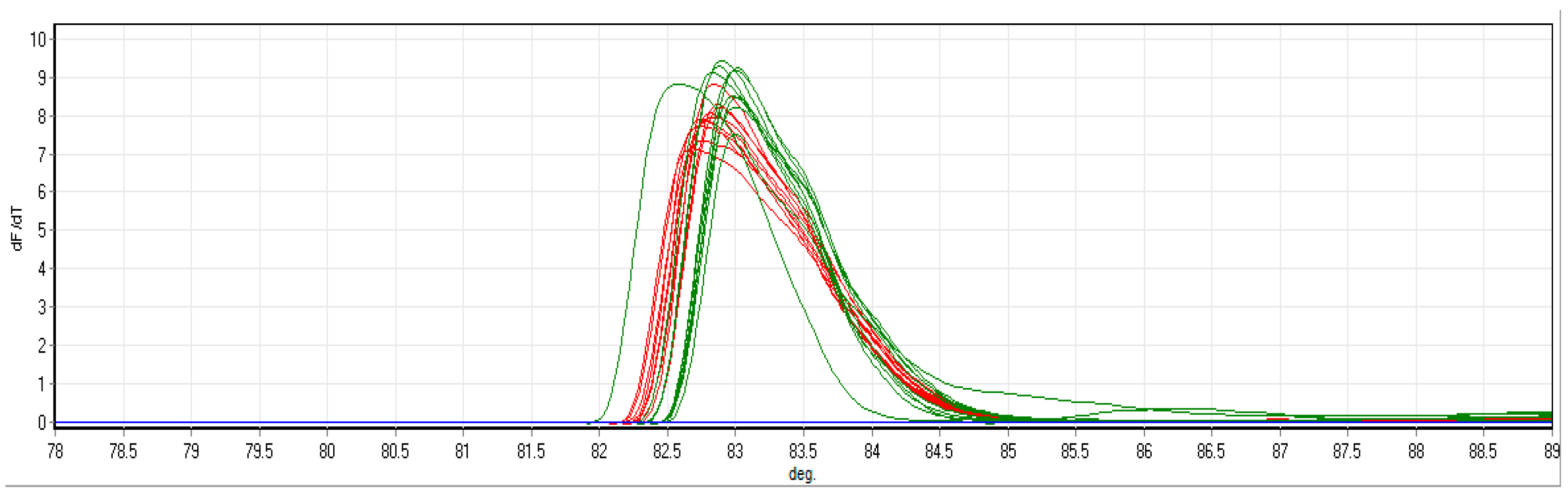
2.5. High-Resolution Melt-Curve Quantitative Real-Time PCR (HRM-RT-PCR) of P. delicatissima on CPR Samples from the 608 SB Route
3. Results
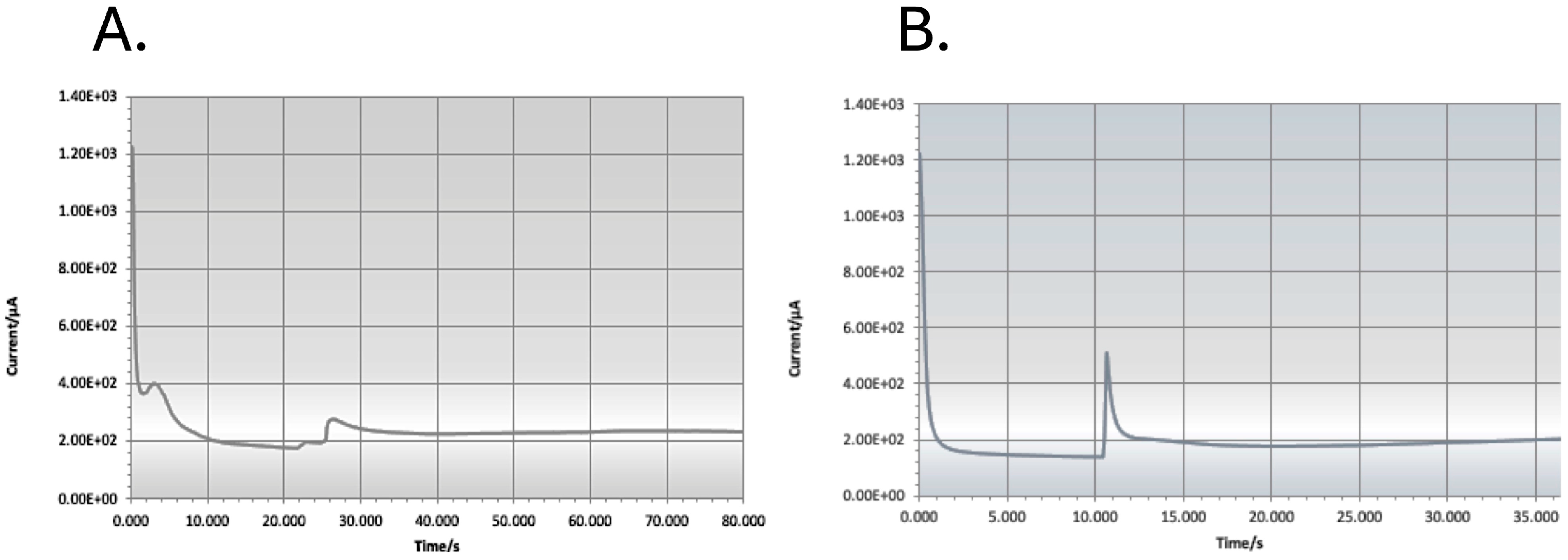
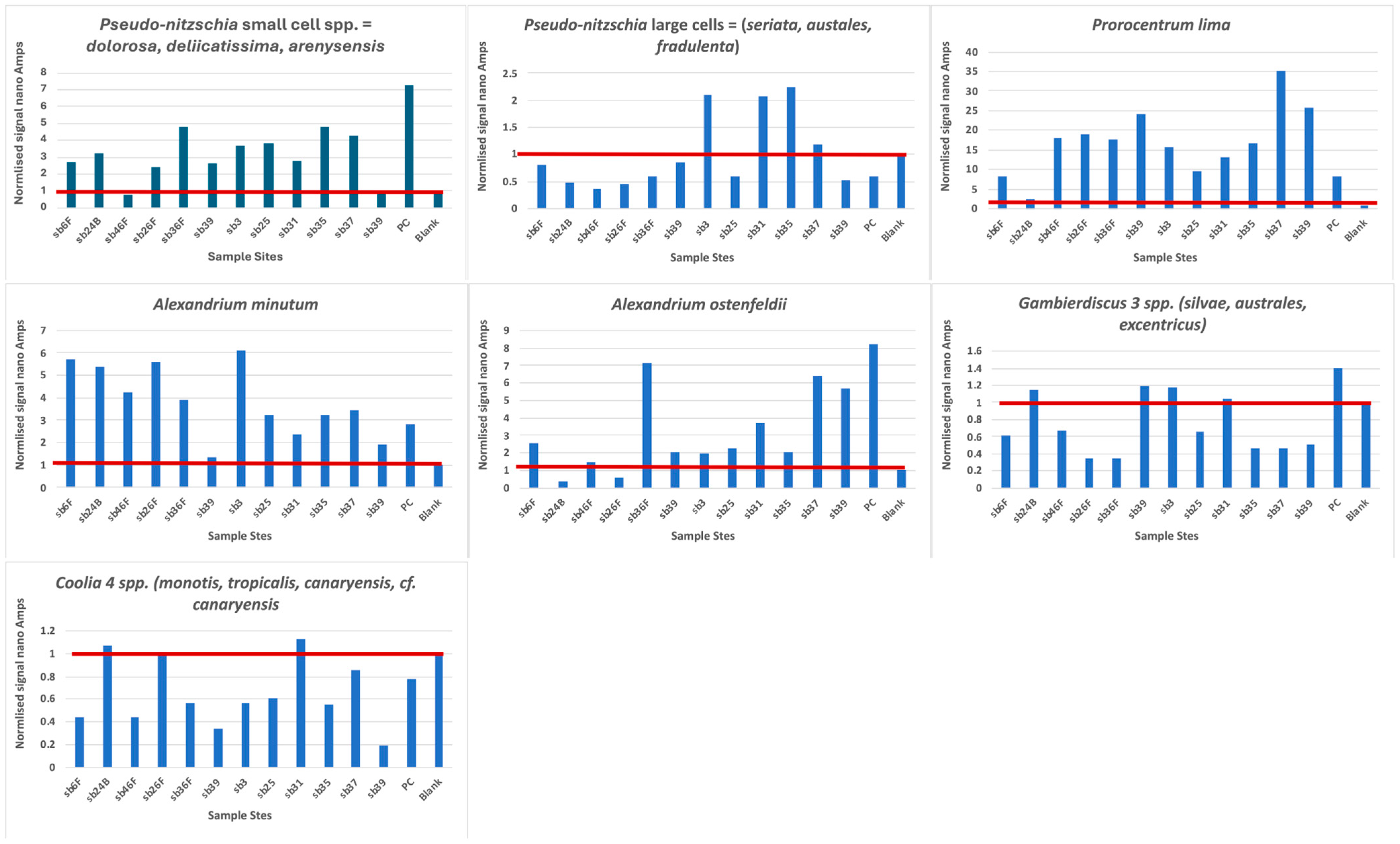
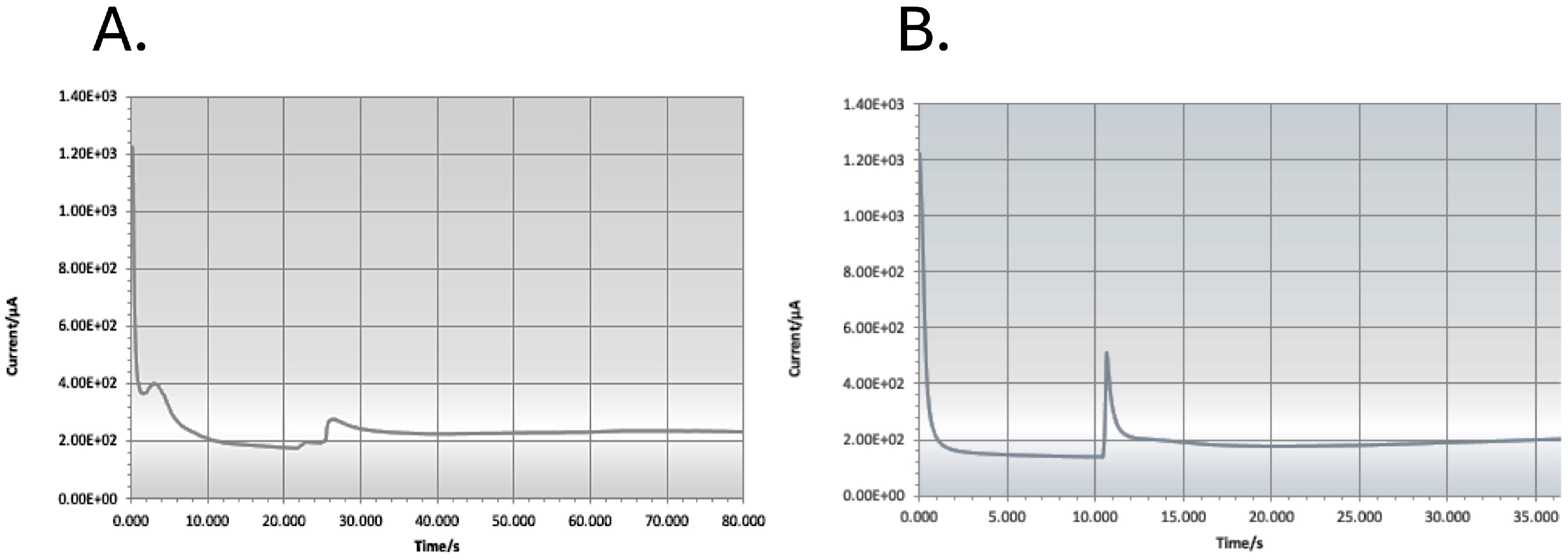
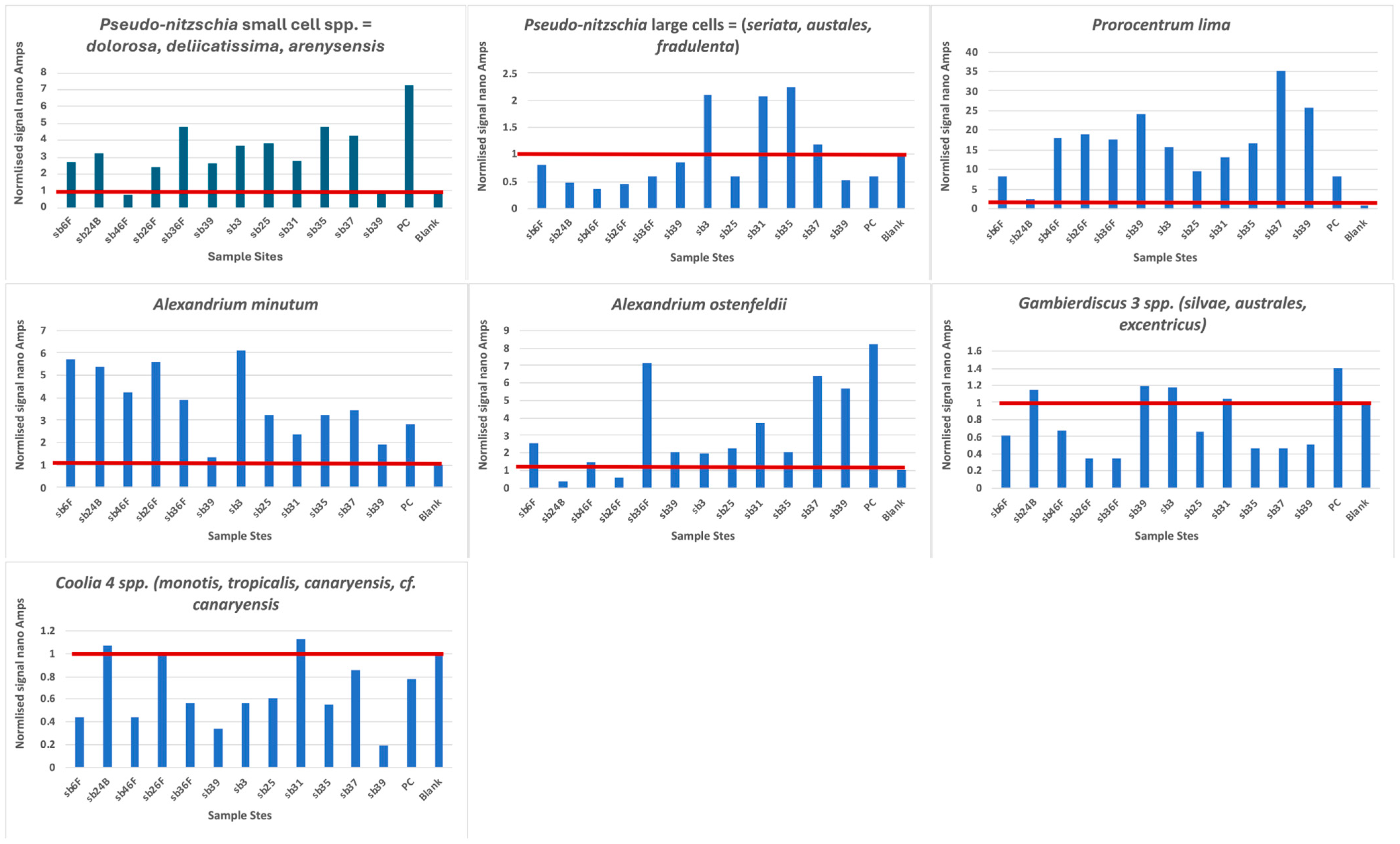
3.1. Biosensor Analysis
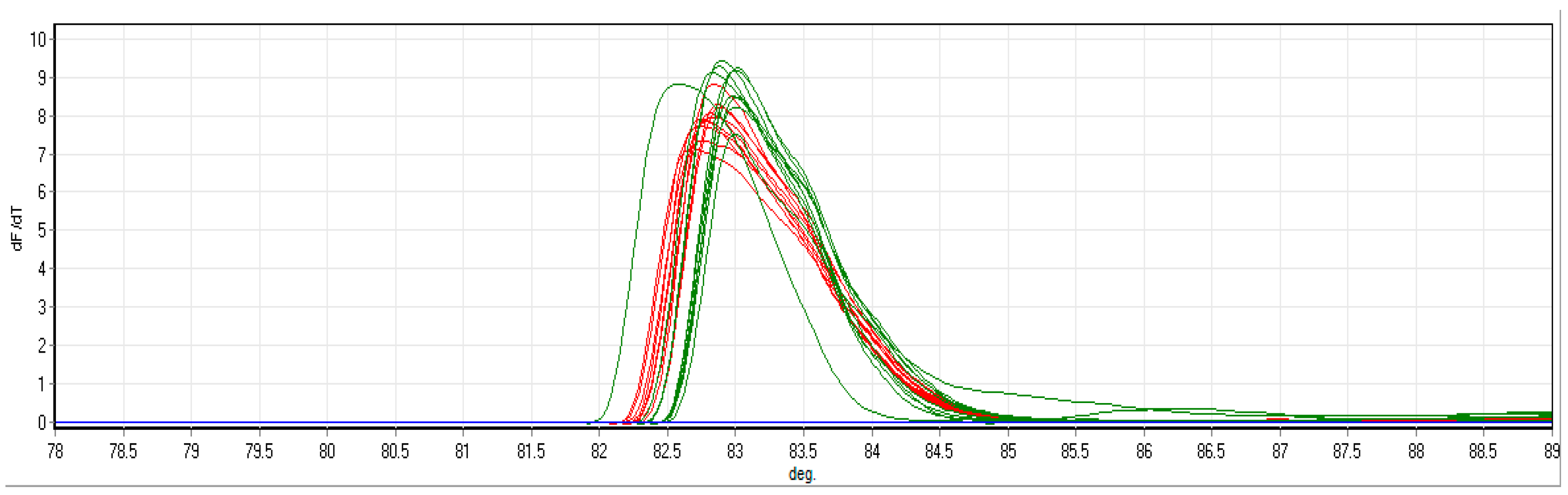
3.2. HRM-RT-PCR Analysis
3.3. Comparison of RT-PCR Time Intervals with Biosensor Detection of Pseudo-nitzschia
3.4. Distribution of Harmful Algae along the Iberian Peninsula
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vezzulli, L.; Martinez-Urtaza, J.; Stern, R. Continuous Plankton Recorder in the omics era: From marine microbiome to global ocean observations. Curr. Opin. Biotechnol. 2022, 73, 61–66. [Google Scholar] [CrossRef]
- Richardson, A.J.; Walne, A.W.; John, A.W.G.; Jonas, T.D.; Lindley, J.A.; Sims, D.W.; Stevens, D.; Witt, M. Using continuous plankton recorder data. Prog. Oceanogr. 2006, 68, 27–74. [Google Scholar] [CrossRef]
- Batten, S.D.; Abu-Alhaija, R.; Chiba, S.; Edwards, M.; Graham, G.; Jyothibabu, R.; Kitchener, J.A.; Koubbi, P.; McQuatters-Gollop, A.; Muxagata, E.; et al. A global plankton diversity monitoring program. Front. Mar. Sci. 2019, 6, 321. [Google Scholar] [CrossRef]
- Hinder, S.L.; Hays, G.C.; Edwards, M.; Roberts, E.C.; Walne, A.W.; Gravenor, M.B. Changes in marine dinoflagellate and diatom abundance under climate change. Nat. Clim. Chang. 2012, 2, 271–275. [Google Scholar] [CrossRef]
- Stern, R.; Schroeder, D.; Highfield, A.; Al-Kandari, M.; Vezzulli, L.; Richardson, A. Chapter 2—Uses of molecular taxonomy in identifying phytoplankton communities from the Continuous Plankton Recorder Survey. In Advances in Phytoplankton Ecology; Clementson, L.A., Eriksen, R.S., Willis, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2022; pp. 47–79. [Google Scholar]
- Oosting, T.; Hilario, E.; Wellenreuther, M.; Ritchie, P.A. DNA degradation in fish: Practical solutions and guidelines to improve DNA preservation for genomic research. Ecol. Evol. 2020, 8643–8651. [Google Scholar] [CrossRef] [PubMed]
- Ripley, S.J.; Baker, A.C.; Miller, P.I.; Walne, A.W.; Schroeder, D.C. Development and validation of a molecular technique for the analysis of archived formalin-preserved phytoplankton samples permits retrospective assessment of Emiliania huxleyi communities. J. Microbiol. Methods 2008, 73, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Shiozaki, T.; Itoh, F.; Hirose, Y.; Onodera, J.; Kuwata, A.; Harada, N. A DNA metabarcoding approach for recovering plankton communities from archived samples fixed in formalin. PLoS ONE 2021, 16, e0245936. [Google Scholar] [CrossRef]
- Stern, R.; Moore, S.; Trainer, V.; Bill, B.; Fischer, A.; Batten, S. Spatial and temporal patterns of Pseudo-nitzschia genetic diversity in the North Pacific Ocean from the Continuous Plankton Recorder survey. Mar. Ecol.-Prog. Ser. 2018, 606, 7–28. [Google Scholar] [CrossRef]
- Vezzulli, L.; Brettar, I.; Pezzati, E.; Reid, P.C.; Colwell, R.R.; Höfle, M.G.; Pruzzo, C. Long-term effects of ocean warming on the prokaryotic community: Evidence from the Vibrios. ISME 2012, 6, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Johns, D.; Broughton, D. The CPR Survey; v1.0, Dataset/Sampling Event; Marine Biological Association: Plymouth, UK, 2019. [Google Scholar] [CrossRef]
- Penna, A.; Galluzzi, L. The quantitative real-time PCR applications in the monitoring of marine harmful algal bloom (HAB) species. Environ. Sci. Pollut. Res. 2013, 20, 6851–6862. [Google Scholar] [CrossRef]
- Vezzulli, L.; Grande, C.; Reid, P.C.; Hélaouët, P.; Edwards, M.; Höfle, M.G.; Brettar, I.; Colwell, R.R.; Pruzzo, C. Climate influence on Vibrio and associated human diseases during the past half-century in the coastal North Atlantic. Proc. Nat. Acad. Sci. USA 2016, 113, E5062–E5071. [Google Scholar] [CrossRef]
- Clarke, J. Determining the Occurence of Non-Toxic Variants of Harmful Algal Species Using Automated Sampling Platforms. Master’s Thesis, University of Plymouth, Plymouth, UK, 2015. [Google Scholar]
- Scholin, C.A.; Herzog, M.; Sogin, M.; Anderson, D.M. Identification of group- and strain-specific genetic markers for globally distributed Alexandrium (Dinophyceae). II. Sequence analysis of a fragment of the LSU rRNA gene. J. Phycol. 1994, 30, 999–1011. [Google Scholar] [CrossRef]
- McDonald, S.M.; Sarno, D.; Zingone, A. Identifying Pseudo-nitzschia species in natural samples using genus-specific PCR primers and clone libraries. Harmful Algae 2007, 9, 849–860. [Google Scholar] [CrossRef]
- Guillou, L.; Nézan, E.; Cueff, V.; Erard-Le Denn, E.; Cambon-Bonavita, M.; Gentien, P.; Barbier, G. Genetic diversity and molecular detection of three toxic dinoflagellate genera (Alexandrium, Dinophysis, and Karenia) from French Coasts. Protist 2002, 153, 222–238. [Google Scholar] [CrossRef] [PubMed]
- Toebe, K.; Alpermann, T.J.; Tillmann, U.; Krock, B.; Cembella, A.; John, U. Molecular discrimination of toxic and non-toxic Alexandrium species (Dinophyta) in natural phytoplankton assemblages from the Scottish coast of the North Sea. Eur. J. Phycol. 2013, 48, 12–26. [Google Scholar] [CrossRef]
- Medlin, L.K.; Gamella, M.; Mengs, G.; Serafín, V.; Campuzano, S.; Pingarrón, J.M. Advances in the detection of toxic algae using electrochemical biosensors. Biosensors 2020, 10, 207. [Google Scholar] [CrossRef] [PubMed]
- Stern, R.; Picard, K.; Clarke, J.; Walker, C.E.; Martins, C.; Marshall, C.; Amorim, A.; Woodward, E.M.S.; Widdicombe, C.; Tarran, G.; et al. Composition and patterns of taxa assemblages in the western channel assessed by 18s sequencing, microscopy and flow cytometry. J. Mar. Sci. Eng. 2023, 11, 480. [Google Scholar] [CrossRef]
- Ruijter, J.M.; Pfaffl, M.W.; Zhao, S.; Spiess, A.N.; Boggy, G.; Blom, J.; Rutledge, R.G.; Sisti, D.; Lievens, A.; De Preter, K.; et al. Evaluation of qPCR curve analysis methods for reliable biomarker discovery: Bias, resolution, precision, and implications. Methods 2013, 59, 32–46. [Google Scholar] [CrossRef] [PubMed]
- Guiry, M.D. Exuviaella lima (Ehrenberg) Bütschli 1885; AlgaeBase—World-Wide Electronic Publication: Galway, Ireland, 2016. [Google Scholar]
- Ajani, P.A.; Verma, A.; Kim, J.H.; Woodcock, S.; Nishimura, T.; Farrell, H.; Zammit, A.; Brett, S.; Murray, S.A. Using qPCR and high-resolution sensor data to model a multi-species Pseudo-nitzschia (Bacillariophyceae) bloom in southeastern Australia. Harmful Algae 2021, 108, 102095. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.X.; Dong, H.C.; Lundholm, N.; Teng, S.T.; Zheng, G.C.; Tan, Z.J.; Lim, P.T.; Li, Y. Species composition and toxicity of the genus Pseudo-nitzschia in Taiwan Strait, including P. chiniana sp. nov. and P. qiana sp. nov. Harmful Algae 2019, 84, 195–209. [Google Scholar] [CrossRef]
- Einaga, N.; Yoshida, A.; Noda, H.; Suemitsu, M.; Nakayama, Y.; Sakurada, A.; Kawaji, Y.; Yamaguchi, H.; Sasaki, Y.; Tokino, T.; et al. Assessment of the quality of DNA from various formalin-fixed paraffin-embedded (FFPE) tissues and the use of this DNA for next-generation sequencing (NGS) with no artifactual mutation. PLoS ONE 2017, 12, e0176280. [Google Scholar] [CrossRef]
- Guiry, M.D.; Guiry, G.M. AlgaeBase. World-Wide Electronic Publication. 2017. Available online: https://www.algaebase.org (accessed on 14 January 2024).
- Trainer, V.L.; Bates, S.S.; Lundholm, N.; Thessen, A.E.; Cochlan, W.P.; Adams, N.G.; Trick, C.G. Pseudo-nitzschia physiological ecology, phylogeny, toxicity, monitoring and impacts on ecosystem health. Harmful Algae 2012, 14, 271–300. [Google Scholar] [CrossRef]
- Crespo, B.G.; Figueiras, F.G. A spring poleward current and its influence on microplankton assemblages and harmful dinoflagellates on the western Iberian coast. Harmful Algae 2007, 6, 686–699. [Google Scholar] [CrossRef]
- Torres Palenzuela, J.M.; González Vilas, L.; Bellas, F.M.; Garet, E.; González-Fernández, A.; Spyrakos, E. Pseudo-nitzschia blooms in a coastal upwelling system: Remote sensing detection, toxicity and environmental variables. Water 2019, 11, 1954. [Google Scholar] [CrossRef]
- Lundholm, N.; Moestrup, Ø.; Kotaki, Y.; Hoef-Emden, K.; Scholin, C.; Miller, P. Inter- and intraspecific variation of the Pseudo-nitzschia delicatissima complex (Bacillariophyceae) illustrated by rRNA probes, morphological data and phylogenetic analyses. J. Phycol. 2006, 42, 464–481. [Google Scholar] [CrossRef]
- Blanco, J.; Arévalo, F.; Moroño, Á.; Correa, J.; Rossignoli, A.E.; Lamas, J.P. Spirolides in bivalve mollusk of the Galician (NW Spain) coast: Interspecific, spatial, temporal variation and presence of an isomer of 13-desmethyl spirolide C. Toxins 2022, 15, 13. [Google Scholar] [CrossRef]
- Lamas, J.P.; Arévalo, F.; Moroño, Á.; Correa, J.; Rossignoli, A.E.; Blanco, J. Gymnodimine A in mollusks from the north Atlantic Coast of Spain: Prevalence, concentration, and relationship with spirolides. Environ. Poll. 2021, 279, 116919. [Google Scholar] [CrossRef] [PubMed]
- Bravo, I.; Isabel Figueroa, R.; Garcés, E.; Fraga, S.; Massanet, A. The intricacies of dinoflagellate pellicle cysts: The example of Alexandrium minutum cysts from a bloom-recurrent area (Bay of Baiona, NW Spain). Deep Sea Res. Part II Top. Studies Ocean. 2010, 57, 166–174. [Google Scholar] [CrossRef]
- Nogueira, E.; Bravo, I.; Montero, P.; Díaz-Tapia, P.; Calvo, S.; Ben-Gigirey, B.; Figueroa, R.I.; Garrido, J.L.; Ramilo, I.; Lluch, N.; et al. HABs in coastal upwelling systems: Insights from an exceptional red tide of the toxigenic dinoflagellate Alexandrium minutum. Ecol. Ind. 2022, 137, 108790. [Google Scholar] [CrossRef]
- Godinho, L.; Soliño, L.; Churro, C.; Timoteo, V.; Santos, C.; Gouveia, N.; Diogène, J.; Reis Costa, P. Distribution, identification and cytotoxicity of Gambierdiscus (Dinophyceae) in the Atlantic Selvagens Islands (Madeira, Portugal): A ciguatera gateway to Europe. Eur. J. Phycol. 2023, 58, 156–168. [Google Scholar] [CrossRef]
- Costa, P.R.; Churro, C.; Rodrigues, S.M.; Frazão, B.; Barbosa, M.; Godinho, L.; Soliño, L.; Timóteo, V.; Gouveia, N. A 15-Year retrospective review of ciguatera in the Madeira Islands (North-East Atlantic, Portugal). Toxins 2023, 15, 630. [Google Scholar] [CrossRef]
- Bravo, I.; Rodriguez, F.; Ramilo, I.; Rial, P.; Fraga, S. Ciguatera-causing dinoflagellate Gambierdiscus spp. (Dinophyceae) in a subtropical region of north Atlantic Ocean (Canary Islands): Morphological characterization and biogeography. Toxins 2019, 11, 423. [Google Scholar] [CrossRef] [PubMed]
- David, H.; Laza-Martínez, A.; Miguel, I.; Orive, E. Broad distribution of Coolia monotis and restricted distribution of Coolia cf. canariensis (Dinophyceae) on the Atlantic coast of the Iberian Peninsula. Phycologia 2014, 53, 342–352. [Google Scholar] [CrossRef]
- David, H.; Laza-Martínez, A.; Kromkamp, J.C.; Orive, E. Physiological response of Prorocentrum lima (Dinophyceae) to varying light intensities. FEMS Micro. Ecol. 2017, 94, fix166. [Google Scholar] [CrossRef] [PubMed]
- Torres, R.B.; Barton, E.D.; Miller, P.; Fanjul, E. Spatial patterns of wind and sea surface temperature in the Galician upwelling region. J. Geophys. Res. Space Phys. 2003, 108, 1–14. [Google Scholar] [CrossRef]
- González-Nuevo, G.; Nogueira, E. Intrusions of warm and salty waters onto the NW and N Iberian shelf in early spring and its relationship to climate variability. J. Atmos. Ocean. Sci. 2005, 10, 361–375. [Google Scholar] [CrossRef]
- Picado, A.; Vaz, N.; Alvarez, I.; Dias, J.M. Modelling coastal upwelling off NW Iberian Peninsula: New insights on the fate of phytoplankton blooms. Sci. Total. Environ. 2023, 874, 162416. [Google Scholar] [CrossRef]
- Quality Status Report 2000: Region IV—Bay of Biscay and Iberian Coast; OSPAR Commission: London, UK, 2000; Available online: https://qsr2010.ospar.org/media/assessments/QSR_2000_Region_IV.pdf (accessed on 10 January 2024).
- Rines, J.E.B.; Donaghay, P.L.; Dekshenieks, M.M.; Sullivan, J.M.; Twardowski, M.S. Thin layers and camouflage: Hidden Pseudo-nitzschia spp. (Bacillariophyceae) populations in a fjord in the San Juan Islands, Washington, USA. Mar. Ecol.-Prog. Ser. 2002, 225, 123–137. [Google Scholar] [CrossRef]
- Painter, S.C.; Finlay, M.; Hemsley, V.S.; Martin, A.P. Seasonality, phytoplankton succession and the biogeochemical impacts of an autumn storm in the northeast Atlantic Ocean. Prog. Oceanogr. 2016, 142, 72–104. [Google Scholar] [CrossRef]
- Martens, H.; Van de Waal, D.B.; Brandenburg, K.M.; Krock, B.; Tillmann, U. Salinity effects on growth and toxin production in an Alexandrium ostenfeldii (Dinophyceae) isolate from The Netherlands. J. Plank. Res. 2016, 38, 1302–1316. [Google Scholar] [CrossRef]
- Bravo, I.; Fraga, S.; Figueroa, R.I.; Pazos, Y.; Massanet, A.; Ramilo, I. Bloom dynamics and life cycle strategies of two toxic dinoflagellates in a coastal upwelling system (NW Iberian Peninsula). Deep Sea Res. Part II Top. Stud. Oceanogr. 2010, 57, 222–234. [Google Scholar] [CrossRef]
- Ruvindy, R.; Barua, A.; Bolch, C.J.S.; Sarowar, C.; Savela, H.; Murray, S.A. Genomic copy number variability at the genus, species and population levels impacts in-situ ecological analyses of dinoflagellates and harmful algal blooms. ISME Commun. 2023, 3, 70. [Google Scholar] [CrossRef] [PubMed]
- Yarimizu, K.; Sildever, S.; Hamamoto, Y.; Tazawa, S.; Oikawa, H.; Yamaguchi, H.; Basti, L.; Mardones, J.I.; Paredes-Mella, J.; Nagai, S. Development of an absolute quantification method for ribosomal RNA gene copy numbers per eukaryotic single cell by digital PCR. Harmful Algae 2021, 103, 102008. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Hu, Z.; Deng, Y.; Shang, L.; Gobler, C.J.; Tang, Y.Z. Dependence of genome size and copy number of rRNA gene on cell volume in dinoflagellates. Harmful Algae 2021, 109, 102108. [Google Scholar] [CrossRef]
- Martínez, J.; García-Ladona, E.; Ballabrera-Poy, J.; Isern-Fontanet, J.; González-Motos, S.; Allegue, J.M.; González-Haro, C. Atlas of surface currents in the Mediterranean and Canary–Iberian–Biscay waters. J. Oper. Oceanogr. 2024, 17, 40–62. [Google Scholar] [CrossRef]
- Heimann, K.; Sparrow, L. Chapter 37—Ciguatera: Tropical reef fish poisoning. In Handbook of Marine Microalgae; Kim, S.-K., Ed.; Academic Press: Boston, MA, USA, 2015; pp. 547–558. [Google Scholar]
- Ramilo, I.; Figueroa, R.I.; Rayon-Vina, F.; Cuadrado, A.; Bravo, I. Temperature-dependent growth and sexuality of the ciguatoxin producer dinoflagellate Gambierdiscus spp. in cultures established from the Canary Islands. Harmful Algae 2021, 110, 102130. [Google Scholar] [CrossRef]
- Collins, R.A.; Wangensteen, O.S.; O’Gorman, E.J.; Mariani, S.; Sims, D.W.; Genner, M.J. Persistence of environmental DNA in marine systems. Commun. Biol. 2018, 1, 185. [Google Scholar] [CrossRef]
- de la Vara, A.; Cabos, W.; Sein, D.V.; Teichmann, C.; Jacob, D. Impact of air–sea coupling on the climate change signal over the Iberian Peninsula. Clim. Dyn. 2021, 57, 2325–2349. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample ID | Year | Month | Latitude | Longitude | DNA Extraction Time Point |
|---|---|---|---|---|---|
| 608SB-3 | 2014 | 5 | 38.558 | −9.662 | 9 y |
| 608SB-4D * | 2014 | 5 | 38.667 | −9.82 | 1 m |
| 608SB-4D | 2014 | 5 | 38.667 | −9.82 | 7 m |
| 608SB-6F * | 2014 | 5 | 38.997 | −9.833 | 1 m |
| 608SB-6F | 2014 | 5 | 38.997 | −9.833 | 7 m |
| 608SB-13 | 2014 | 5 | 40.163 | −9.833 | 9 y |
| 608SB-14D * | 2014 | 5 | 40.332 | −9.833 | 1 m |
| 608SB-14D | 2014 | 5 | 40.332 | −9.833 | 7 m |
| 608SB-16F * | 2014 | 5 | 40.663 | −9.833 | 1 m |
| 608SB-16F | 2014 | 5 | 40.663 | −9.833 | 7 m |
| 608SB-19 | 2014 | 5 | 41.165 | −9.883 | 9 y |
| 608SB-24D * | 2014 | 5 | 41.997 | −9.833 | 1 m |
| 608SB-24D | 2014 | 5 | 41.997 | −9.833 | 7 m |
| 608SB-25 | 2014 | 5 | 42.163 | −9.833 | 9 y |
| 608SB-26F * | 2014 | 5 | 42.33 | −9.833 | 1 m |
| 608SB-26F | 2014 | 5 | 42.33 | −9.833 | 7 m |
| 608SB-31 | 2014 | 5 | 43.17 | −9.787 | 9 y |
| 608SB-33 | 2014 | 5 | 43.488 | −9.653 | 9 y |
| 608SB-34D * | 2014 | 5 | 43.647 | −9.585 | 1 m |
| 608SB-34D | 2014 | 5 | 43.647 | −9.585 | 7 m |
| 608SB-35 | 2014 | 5 | 43.807 | −9.517 | 9 y |
| 608SB-36F * | 2014 | 5 | 43.967 | −9.448 | 1 m |
| 608SB-36F | 2014 | 5 | 43.967 | −9.448 | 7 m |
| 608SB-39 | 2014 | 5 | 44.125 | −9.38 | 9 y |
| 608SB-37 | 2014 | 5 | 44.443 | −9.242 | 9 y |
| 608SB-44D * | 2014 | 5 | 45.238 | −8.887 | 1 m |
| 608SB-44D | 2014 | 5 | 45.238 | −8.887 | 7 m |
| 608SB-46F * | 2014 | 5 | 45.555 | −8.743 | 1 m |
| 608SB-46F | 2014 | 5 | 45.555 | −8.743 | 7 m |
| Primer Name | Sequence 5′ to 3′ | Reference |
|---|---|---|
| D1R-Eukaryote F | ACCCGCTGAATTTAAGCATA | [15] |
| D2C-Eukaryote R | CCTTGGTCCGTGTTTCAAGA | [15] |
| 186F/D1-Pseudo-nitzschia | GTTCCTTGGAAAAGGACAGCTGA | [16] |
| 548R D1-Pseudo-nitzschia | AGACATCAACTCTGACTG | [16] |
| Alex1 Alexandrium tamarense complex | ACCACCCACTTTGCATTCCA | [17] |
| Amin2 Alexandrium minutum | AGCACTGATGTGTAAGGGCT | [17] |
| 378F Alexandrium tamarense | CCATGAGGGAAATATGAAAGGA | [18] |
| Acat3 Alexandrium catenella | AAGTGCAACACTCCCACCAA | [17] |
| PdeliLSUF2 | CCTGGTGGAGTGAGTCGTTGTCAACG | [14] |
| PdeliLSUR3 | TCAACCAAAGCAAACCCACGCAAGCTC | [14] |
| Sample Number | Latitude | Longitude |
|---|---|---|
| 608SB-46 (F) | 45.555 | −8.743 |
| 608SB-39 | 44.443 | −9.242 |
| 608SB-37 | 44.125 | −9.38 |
| 608SB-36 (F) | 43.967 | −9.448 |
| 608SB-35 | 43.807 | −9.517 |
| 608SB-31 | 43.17 | −9.787 |
| 608SB-26 (F) | 42.33 | −9.833 |
| 608SB-25 | 42.163 | −9.833 |
| 608SB-24 (D) | 41.997 | −9.833 |
| 608SB-6 (F) | 38.997 | −9.833 |
| 608SB-3 | 38.558 | −9.662 |
| Species Probe | Approximate Location in LSU Using Prorocentrum micans as a Reference Sequence for the Dinoflagellate Probes. Those Positions in () Are Not Present in the Shorter PCR Fragment Amplified with 378F + D2R. |
|---|---|
| Alexandrium minutum | 583 |
| Alexandrium ostenfeldii | 629 |
| Coolia 4 spp. (monotis, tropicalis, canariensis, cf. canariensis | 379, (177), 393, (265) |
| Gambierdiscus 2 spp. (silvae, australes) | (76), (338) |
| Prorocentrum lima species complex | 554, 426 |
| Pseudo-nitzschia small cells = (delicatissima, dolorosa, arenysensis) | 579, (143), (167) |
| Pseudo-nitzschia large cells = (seriata, australes, fraudulenta) | 379, 439, 424 |
| SB46F | SB39 | SB39 Short | SB37 | SB36F | SB35 | SB31 | SB26F | SB25 | SB24D | SB6F | SB3 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Pseudo-nitzschia small cells | + | ++ | + | +++ | +++ | +++ | ++ | ++ | +++ | +++ | ++ | +++ |
| Pseudo-nitzschia large cells | + | + | + | ++ | + | ++ | ++ | + | + | + | + | ++ |
| Prorocentrum lima | +++ | +++ | +++ | +++ | +++ | +++ | +++ | +++ | +++ | ++ | +++ | +++ |
| Alexandrium ostenfeldii | + | ++ | +++ | +++ | +++ | ++ | +++ | ++ | ++ | +++ | ++ | ++ |
| Alexandrium minutum | +++ | ++ | ++ | +++ | +++ | +++ | ++ | +++ | +++ | +++ | +++ | +++ |
| Gambierdiscus 2 spp. | + | ++ | + | + | + | + | ++ | + | + | ++ | + | ++ |
| Coolia 4 spp. | + | + | + | + | + | + | ++ | ++ | + | ++ | + | + |
| Sample ID | P. delicatissima RT-PCR Time 1 m | P. delicatissima RT-PCR Time 7 m | Small Cell of Pseudo-nitzschia Biosensor Time 9 yr |
|---|---|---|---|
| 608SB-4D * | 1 (A) | 11 (A) | N/A |
| 608SB-6F * | 2 (P) | 12 (A) | P |
| 608SB-14D * | 3 (A) | 13 (A) | N/A |
| 608SB-16F * | 4 (P) | 14 (P) | N/A |
| 608SB-24D * | 5 (P) | 15 (A) | P |
| 608SB-26F * | 6 (P) | 16 (P) | P |
| 608SB-34D * | 7 (A) | 17 (A) | N/A |
| 608SB-36F * | 8 (P) | 18 (A) | P |
| 608SB-44D * | 9 (P) | 19 (A) | N/A |
| 608SB-46F * | 10 (P) | 20 (A) | P |
| Biosensor Taxa Positively Identified | Identified Historically in the Iberian Peninsula | Reference | Explanation |
|---|---|---|---|
| Large cells of Pseudo-nitzschia (>3 µM width) | Yes | [28] [27] [29] | Wide variety of larger species of P. seriata complex, commonly P. australis and also P. pungens. Mostly near coastal areas, except at surface NW point, outside Iberian Poleward current. CPR microscopic observations at 608SB-3, -35, -36, -37, -39. |
| Small cells of Pseudo-nitzschia (<3 µM width) | Yes | [27] [29] [30] | P. delicatissima complex in Rias and isolated from near-coastal Portugal. CPR microscopic observation only at 608SB-25. |
| Alexandrium ostenfeldii | Unclear | [31] [32] | Spirolides and Gymnodinine A in molluscs, also produced by A. ostenfeldii. Not recorded in CPR survey. |
| Alexandrium minutum | Yes | [33] [34] | Typically in Rias with accompanying cyst beds but found offshore in 2018. Not recorded in CPR survey. |
| Gambierdiscus 2 spp. | No | [35] [36] [37] | Found in NE Atlantic Balieric, Madiera and Selvagens Islands reported gateway to Europe. Not recorded in CPR survey. |
| Coolia 4 spp. | Yes | [38] | C. monotis broad distribution. C. canariensis near Bilbao. Not recorded in CPR survey. |
| Prorocentrum lima | Yes | [39] | Isolated in 3 regions. Slow growing taxa. CPR microscopic observation of Prorocentrum “Exuviella” at 608SB-3, -23, -25, -27, -32, -35, -36, -37, -39. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mengs, G.; Stern, R.F.; Clarke, J.L.; Faith, M.; Medlin, L.K. Mapping Selected Emergent Marine Toxin-Producing Organisms Using Historical Samples with Two Methods (Biosensors and Real-Time PCR): A Comparison of Resolution. Appl. Microbiol. 2024, 4, 312-328. https://doi.org/10.3390/applmicrobiol4010021
Mengs G, Stern RF, Clarke JL, Faith M, Medlin LK. Mapping Selected Emergent Marine Toxin-Producing Organisms Using Historical Samples with Two Methods (Biosensors and Real-Time PCR): A Comparison of Resolution. Applied Microbiology. 2024; 4(1):312-328. https://doi.org/10.3390/applmicrobiol4010021
Chicago/Turabian StyleMengs, Gerado, Rowena F. Stern, Jessica L. Clarke, Matthew Faith, and Linda K. Medlin. 2024. "Mapping Selected Emergent Marine Toxin-Producing Organisms Using Historical Samples with Two Methods (Biosensors and Real-Time PCR): A Comparison of Resolution" Applied Microbiology 4, no. 1: 312-328. https://doi.org/10.3390/applmicrobiol4010021
APA StyleMengs, G., Stern, R. F., Clarke, J. L., Faith, M., & Medlin, L. K. (2024). Mapping Selected Emergent Marine Toxin-Producing Organisms Using Historical Samples with Two Methods (Biosensors and Real-Time PCR): A Comparison of Resolution. Applied Microbiology, 4(1), 312-328. https://doi.org/10.3390/applmicrobiol4010021