
Green Macroalgae Hydrolysate for Biofuel Production: Potential of Ulva rigida


 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Macroalgae and Determination of Their Composition
2.2. Microorganisms and Inoculum Preparation
2.3. Endogenous Biomass Identification
2.4. Preparation of Algal Hydrolysates
2.5. Synthetic Medium
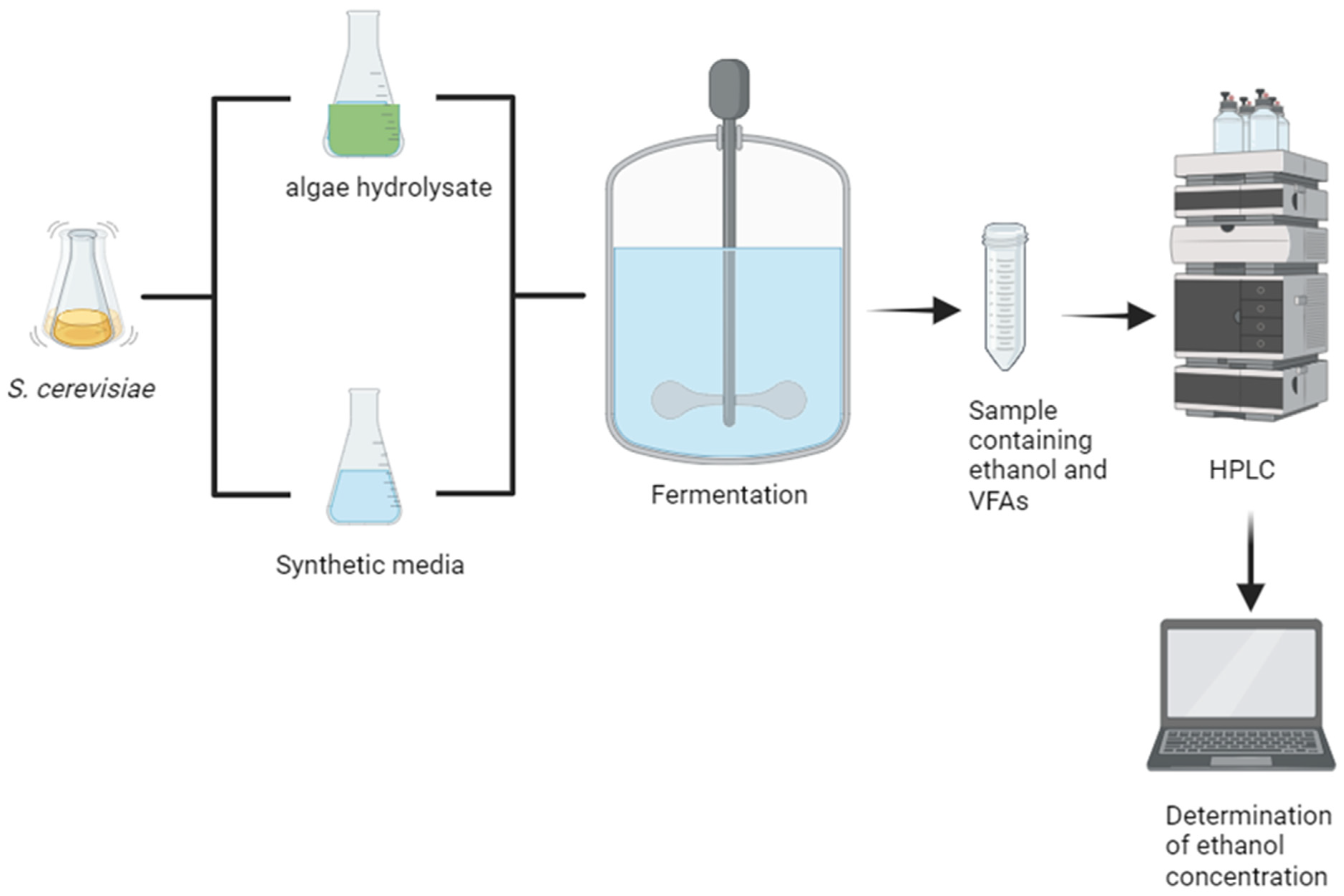
2.6. Ethanol Fermentation
2.7. Analytical Methods
3. Results and Discussion
3.1. Global Composition of Ulva sp.
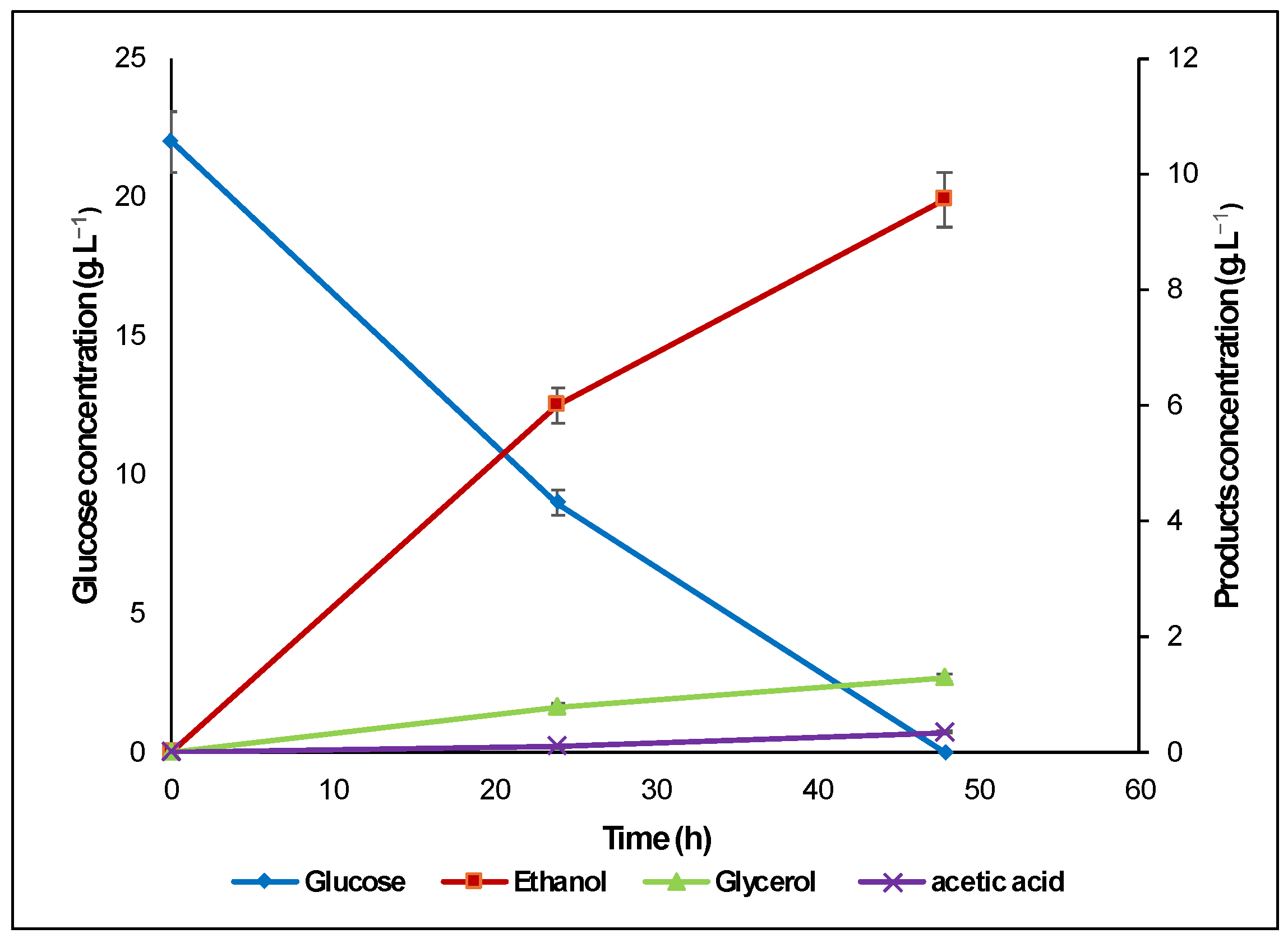
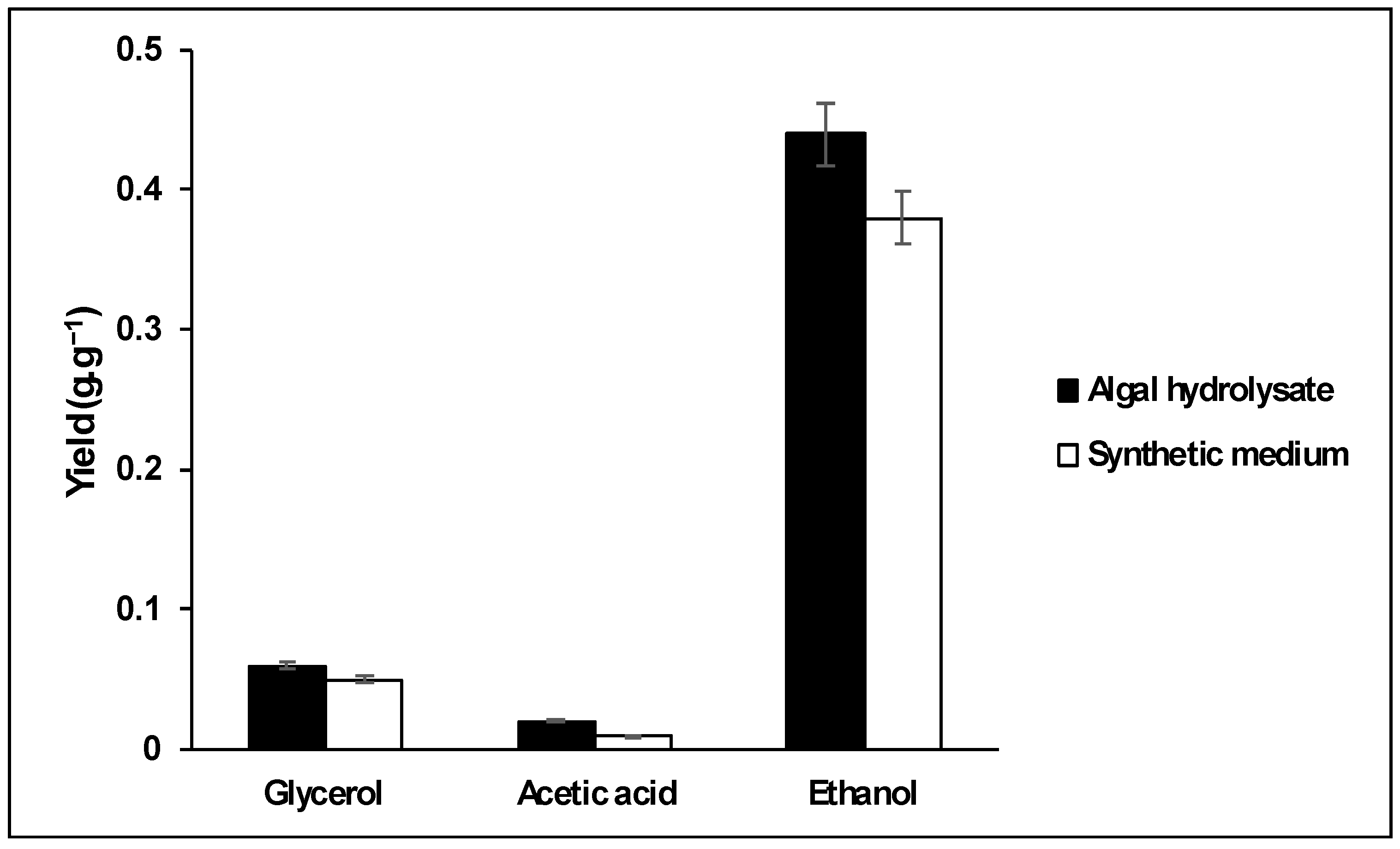
3.2. Fermentation of Algal Hydrolysate with S. cerevisiae
3.3. Optimization of the Fermentation with S. cerevisiae
3.3.1. Effect of Medium Enrichment
3.3.2. Inoculum Size Effect
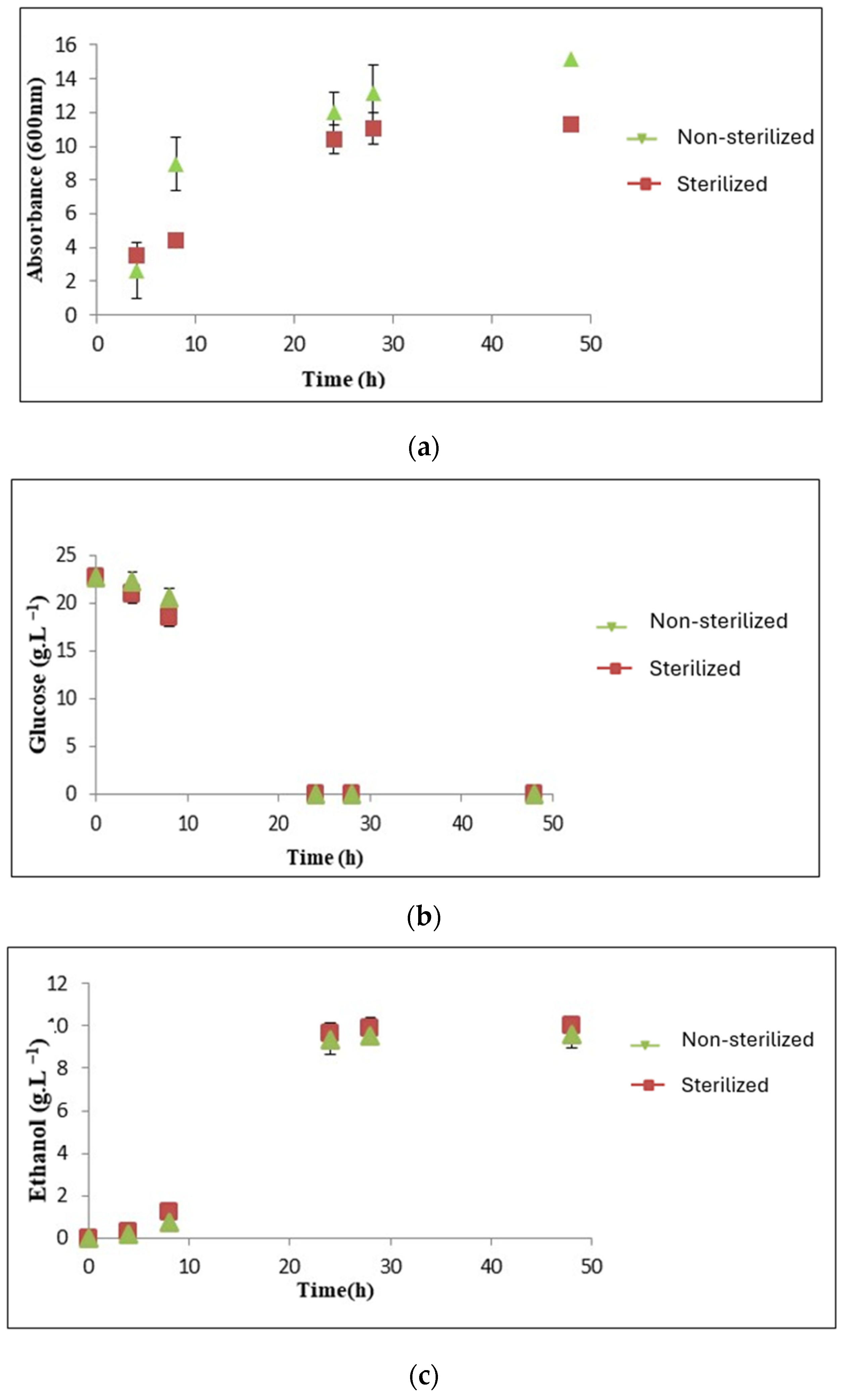
3.3.3. Sterilization Effect
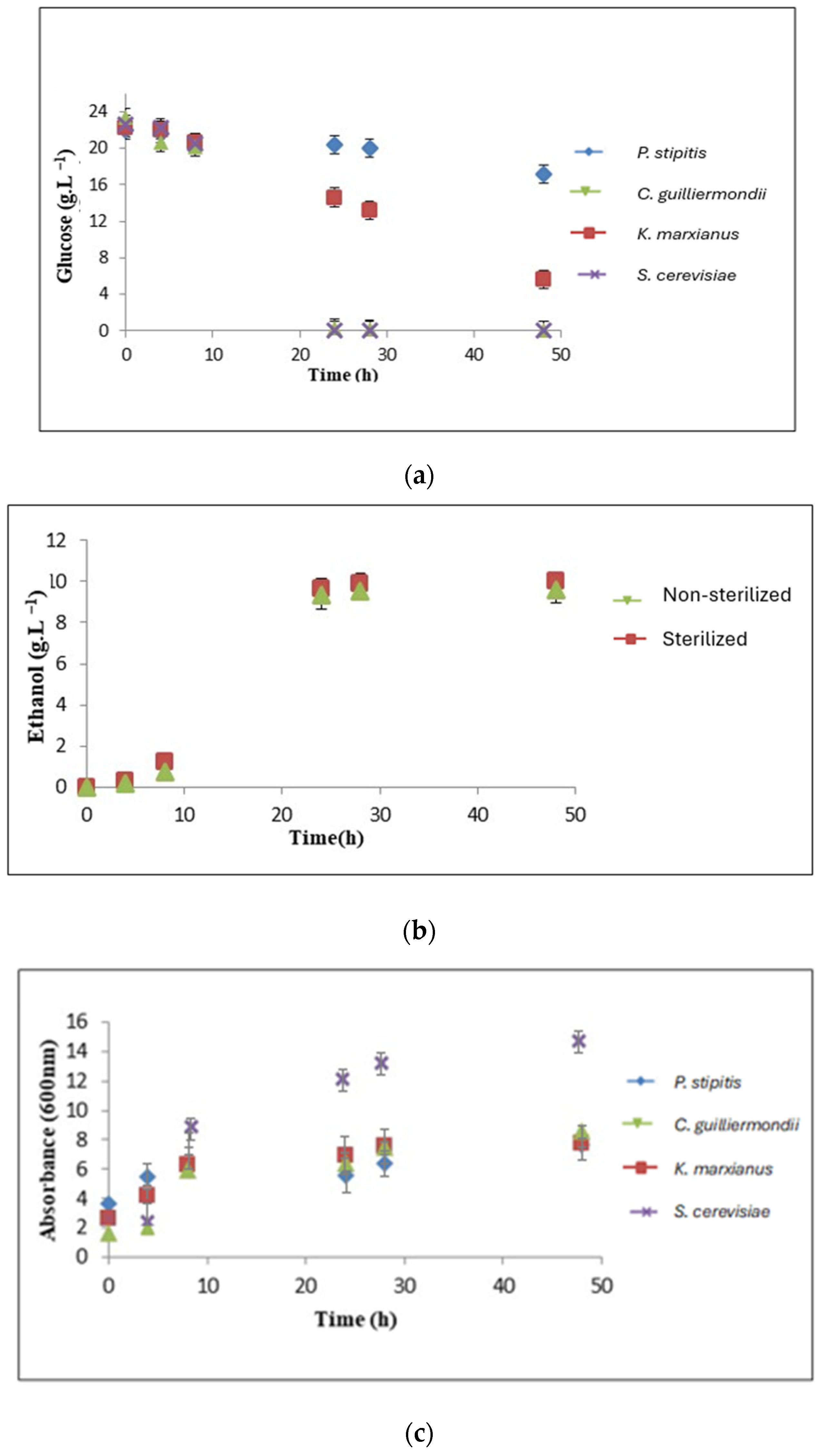
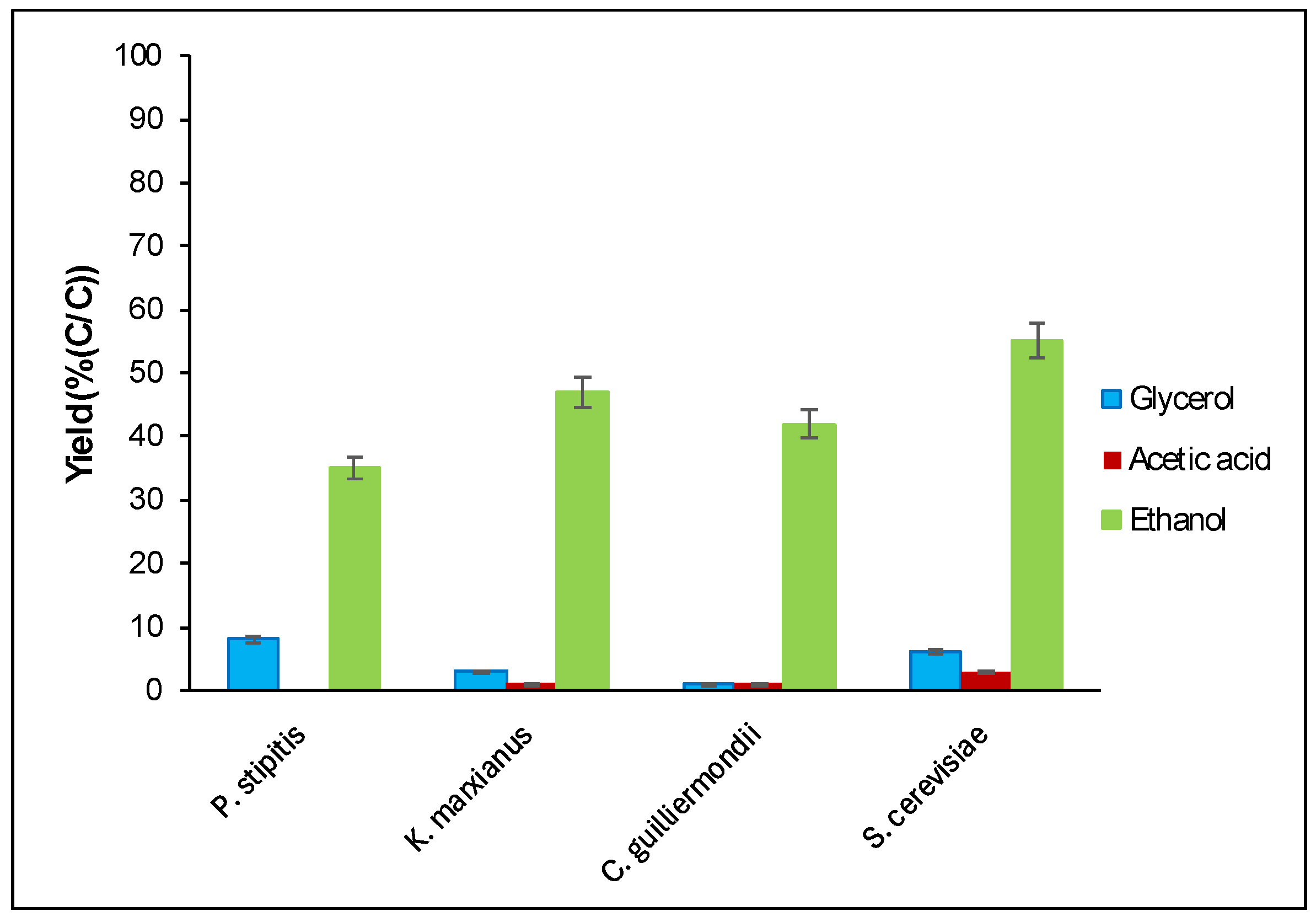
3.4. Comparison with Three Other Yeast Strains
3.5. Comparison of Ethanol Yields for Different Green Algal Feedstocks
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Ruangrit, K.; Chaipoot, S.; Phongphisutthinant, R.; Kamopas, W.; Jeerapan, I.; Pekkoh, J.; Srinuanpan, S. Environmental-Friendly Pretreatment and Process Optimization of Macroalgal Biomass for Effective Ethanol Production as an Alternative Fuel Using Saccharomyces cerevisiae. Biocatal. Agric. Biotechnol. 2021, 31, 101919. [Google Scholar] [CrossRef]
- Farobie, O.; Amrullah, A.; Anis, L.A.; Hartulistiyoso, E.; Syaftika, N.; Saefurahman, G.; Bayu, A. Valorization of Brown Macroalgae sargassum Plagiophyllum for Biogas Production under Different Salinity Conditions. Bioresour. Technol. Rep. 2023, 22, 101403. [Google Scholar] [CrossRef]
- Ullah, K.; Ahmad, M.; Sofia; Sharma, V.K.; Lu, P.M.; Harvey, A.; Zafar, M.; Sultana, S. Assessing the Potential of Algal Biomass Opportunities for Bioenergy Industry: A Review. Fuel 2015, 143, 414–423. [Google Scholar] [CrossRef]
- Demirbas, A.; Fatih Demirbas, M. Importance of Algae Oil as a Source of Biodiesel. Energy Convers. Manag. 2011, 52, 163–170. [Google Scholar] [CrossRef]
- Gielen, D.; Boshell, F.; Saygin, D.; Bazilian, M.D.; Wagner, N.; Gorini, R. The Role of Renewable Energy in the Global Energy Transformation. Energy Strategy Rev. 2019, 24, 38–50. [Google Scholar] [CrossRef]
- Arias, A.; Feijoo, G.; Moreira, M.T. Macroalgae Biorefineries as a Sustainable Resource in the Extraction of Value-Added Compounds. Algal Res. 2023, 69, 102954. [Google Scholar] [CrossRef]
- Bai, F.; Yan, W.; Zhang, S.; Yu, D.; Bai, L. Immobilized Lipase of Reconstructed Oil Bodies and Its Potential Application in Biodiesel Production. Fuel 2014, 128, 340–346. [Google Scholar] [CrossRef]
- Shah, I.H.; Manzoor, M.A.; Jinhui, W.; Li, X.; Hameed, M.K.; Rehaman, A.; Li, P.; Zhang, Y.; Niu, Q.; Chang, L. Comprehensive Review: Effects of Climate Change and Greenhouse Gases Emission Relevance to Environmental Stress on Horticultural Crops and Management. J. Environ. Manag. 2024, 351, 119978. [Google Scholar] [CrossRef]
- Antar, M.; Lyu, D.; Nazari, M.; Shah, A.; Zhou, X.; Smith, D.L. Biomass for a Sustainable Bioeconomy: An Overview of World Biomass Production and Utilization. Renew. Sustain. Energy Rev. 2021, 139, 110691. [Google Scholar] [CrossRef]
- Malla, F.A.; Bandh, S.A.; Wani, S.A.; Hoang, A.T.; Sofi, N.A. Biofuels: Potential Alternatives to Fossil Fuels. In Biofuels in Circular Economy; Bandh, S.A., Malla, F.A., Eds.; Springer Nature: Singapore, 2022; pp. 1–15. ISBN 978-981-19583-7-3. [Google Scholar]
- Wang, P.; Lü, X. Chapter 1—General Introduction to Biofuels and Bioethanol. In Advances in 2nd Generation of Bioethanol Production; Lü, X., Ed.; Woodhead Publishing Series in Energy; Woodhead Publishing: Salzton, UK, 2021; pp. 1–7. ISBN 978-0-12-818862-0. [Google Scholar]
- Loulergue, P.; Balannec, B.; Fouchard-Le Graët, L.; Cabrol, A.; Sayed, W.; Djelal, H.; Amrane, A.; Szymczyk, A. Air-Gap Membrane Distillation for the Separation of Bioethanol from Algal-Based Fermentation Broth. Sep. Purif. Technol. 2019, 213, 255–263. [Google Scholar] [CrossRef]
- Chniti, S.; Djelal, H.; Hassouna, M.; Amrane, A. Residue of Dates from the Food Industry as a New Cheap Feedstock for Ethanol Production. Biomass Bioenergy 2014, 69, 66–70. [Google Scholar] [CrossRef]
- Yue, Z.; Ma, D.; Peng, S.; Zhao, X.; Chen, T.; Wang, J. Integrated Utilization of Algal Biomass and Corn Stover for Biofuel Production. Fuel 2016, 168, 1–6. [Google Scholar] [CrossRef]
- Cripwell, R.A.; Favaro, L.; Viljoen-Bloom, M.; van Zyl, W.H. Consolidated Bioprocessing of Raw Starch to Ethanol by Saccharomyces Cerevisiae: Achievements and Challenges. Biotechnol. Adv. 2020, 42, 107579. [Google Scholar] [CrossRef]
- Barboza-Rodríguez, R.; Rodríguez-Jasso, R.M.; Rosero-Chasoy, G.; Rosales Aguado, M.L.; Ruiz, H.A. Photobioreactor Configurations in Cultivating Microalgae Biomass for Biorefinery. Bioresour. Technol. 2024, 394, 130208. [Google Scholar] [CrossRef]
- Renzaho, A.M.N.; Kamara, J.K.; Toole, M. Biofuel Production and Its Impact on Food Security in Low and Middle Income Countries: Implications for the Post-2015 Sustainable Development Goals. Renew. Sustain. Energy Rev. 2017, 78, 503–516. [Google Scholar] [CrossRef]
- Malode, S.J.; Prabhu, K.K.; Mascarenhas, R.J.; Shetti, N.P.; Aminabhavi, T.M. Recent Advances and Viability in Biofuel Production. Energy Convers. Manag. X 2021, 10, 100070. [Google Scholar] [CrossRef]
- Bušić, A.; Marđetko, N.; Kundas, S.; Morzak, G.; Belskaya, H.; Ivančić Šantek, M.; Komes, D.; Novak, S.; Šantek, B. Bioethanol Production from Renewable Raw Materials and Its Separation and Purification: A Review. Food Technol. Biotechnol. 2018, 56, 289–311. [Google Scholar] [CrossRef]
- Jatoi, A.S.; Abbasi, S.A.; Hashmi, Z.; Shah, A.K.; Alam, M.S.; Bhatti, Z.A.; Maitlo, G.; Hussain, S.; Khandro, G.A.; Usto, M.A.; et al. Recent Trends and Future Perspectives of Lignocellulose Biomass for Biofuel Production: A Comprehensive Review. Biomass Conv. Bioref. 2023, 13, 6457–6469. [Google Scholar] [CrossRef]
- Zhang, K.; Zhang, F.; Wu, Y.-R. Emerging Technologies for Conversion of Sustainable Algal Biomass into Value-Added Products: A State-of-the-Art Review. Sci. Total Environ. 2021, 784, 147024. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Ge, Y.; Xiao, M. Towards Biofuel Generation III+: A Sustainable Industrial Symbiosis Design of Co-Producing Algal and Cellulosic Biofuels. J. Clean. Prod. 2021, 306, 127144. [Google Scholar] [CrossRef]
- Das, P.; Chandramohan, V.P.; Mathimani, T.; Pugazhendhi, A. Recent Advances in Thermochemical Methods for the Conversion of Algal Biomass to Energy. Sci. Total Environ. 2021, 766, 144608. [Google Scholar] [CrossRef] [PubMed]
- Eloka-Eboka, A.C.; Maroa, S.; Behera, S. 16—Algal Biofuels—Technologies, Scope, Opportunities, Challenges, and Applications. In Sustainable Biofuels; Ray, R.C., Ed.; Applied Biotechnology Reviews; Academic Press: Cambridge, MA, USA, 2021; pp. 449–470. ISBN 978-0-12-820297-5. [Google Scholar]
- Kumar, M.; Sun, Y.; Rathour, R.; Pandey, A.; Thakur, I.S.; Tsang, D.C.W. Algae as Potential Feedstock for the Production of Biofuels and Value-Added Products: Opportunities and Challenges. Sci. Total Environ. 2020, 716, 137116. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, Y.; Yoshikuni, Y. Metabolic Engineering for Valorization of Macroalgae Biomass. Metab. Eng. 2022, 71, 42–61. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Zhang, J.; Cui, H. The Isolation and Performance Studies of an Alginate Degrading and Ethanol Producing Strain. Chem. Biochem. Eng. Q. 2014, 28, 391–398. [Google Scholar] [CrossRef]
- Wong, K.H.; Tan, I.S.; Foo, H.C.Y.; Chin, L.M.; Cheah, J.R.N.; Sia, J.K.; Tong, K.T.X.; Lam, M.K. Third-Generation Bioethanol and L-Lactic Acid Production from Red Macroalgae Cellulosic Residue: Prospects of Industry 5.0 Algae. Energy Convers. Manag. 2022, 253, 115155. [Google Scholar] [CrossRef]
- Broda, M.; Yelle, D.J.; Serwańska, K. Bioethanol Production from Lignocellulosic Biomass—Challenges and Solutions. Molecules 2022, 27, 8717. [Google Scholar] [CrossRef]
- Stiger, V.; Bourgougnon, N.; Deslandes, E. Carbohydrates from Seaweeds. In Seaweed in Health and Disease Prevention; Academic Press: Cambridge, MA, USA, 2016; pp. 223–274. ISBN 978-0-12-802772-1. [Google Scholar]
- Guidara, M.; Yaich, H.; Amor, I.B.; Fakhfakh, J.; Gargouri, J.; Lassoued, S.; Blecker, C.; Richel, A.; Attia, H.; Garna, H. Effect of Extraction Procedures on the Chemical Structure, Antitumor and Anticoagulant Properties of Ulvan from Ulva Lactuca of Tunisia Coast. Carbohydr. Polym. 2021, 253, 117283. [Google Scholar] [CrossRef]
- Li, C.; Tang, T.; Du, Y.; Jiang, L.; Yao, Z.; Ning, L.; Zhu, B. Ulvan and Ulva Oligosaccharides: A Systematic Review of Structure, Preparation, Biological Activities and Applications. Bioresour. Bioprocess. 2023, 10, 66. [Google Scholar] [CrossRef]
- Andrade, C.; Martins, P.L.; Duarte, L.C.; Oliveira, A.C.; Carvalheiro, F. Development of an Innovative Macroalgae Biorefinery: Oligosaccharides as Pivotal Compounds. Fuel 2022, 320, 123780. [Google Scholar] [CrossRef]
- Wahlström, N.; Edlund, U.; Pavia, H.; Toth, G.; Jaworski, A.; Pell, A.J.; Choong, F.X.; Shirani, H.; Nilsson, K.P.R.; Richter-Dahlfors, A. Cellulose from the Green Macroalgae Ulva Lactuca: Isolation, Characterization, Optotracing, and Production of Cellulose Nanofibrils. Cellulose 2020, 27, 3707–3725. [Google Scholar] [CrossRef]
- Tan, I.S.; Lam, M.K.; Foo, H.C.Y.; Lim, S.; Lee, K.T. Advances of Macroalgae Biomass for the Third Generation of Bioethanol Production. Chin. J. Chem. Eng. 2020, 28, 502–517. [Google Scholar] [CrossRef]
- Morales-Contreras, B.E.; Flórez-Fernández, N.; Dolores Torres, M.; Domínguez, H.; Rodríguez-Jasso, R.M.; Ruiz, H.A. Hydrothermal Systems to Obtain High Value-Added Compounds from Macroalgae for Bioeconomy and Biorefineries. Bioresour. Technol. 2022, 343, 126017. [Google Scholar] [CrossRef] [PubMed]
- Kazemi Shariat Panahi, H.; Dehhaghi, M.; Aghbashlo, M.; Karimi, K.; Tabatabaei, M. Shifting Fuel Feedstock from Oil Wells to Sea: Iran Outlook and Potential for Biofuel Production from Brown Macroalgae (Ochrophyta; Phaeophyceae). Renew. Sustain. Energy Rev. 2019, 112, 626–642. [Google Scholar] [CrossRef]
- Hou, X.; Hansen, J.H.; Bjerre, A.-B. Integrated Bioethanol and Protein Production from Brown Seaweed Laminaria digit. Bioresour. Technol. 2015, 197, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.Y.; Ra, C.-H.; Kim, S.-K. Ethanol Production from the Seaweed, Gelidium amansii Using Specific Sugar Acclimated Yeasts. J. Microbiol. Biotechnol. 2014, 24, 264–269. [Google Scholar] [CrossRef] [PubMed]
- Kostas, E.T.; White, D.A.; Du, C.; Cook, D.J. Selection of Yeast Strains for Bioethanol Production from UK Seaweeds. J. Appl. Phycol. 2016, 28, 1427–1441. [Google Scholar] [CrossRef]
- Lara, A.; Rodríguez-Jasso, R.M.; Loredo-Treviño, A.; Aguilar, C.N.; Meyer, A.S.; Ruiz, H.A. Chapter 17—Enzymes in the Third Generation Biorefinery for Macroalgae Biomass. In Biomass, Biofuels, Biochemicals; Singh, S.P., Pandey, A., Singhania, R.R., Larroche, C., Li, Z., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 363–396. ISBN 978-0-12-819820-9. [Google Scholar]
- Zhang, R.; Wang, Q.; Shen, H.; Yang, Y.; Liu, P.; Dong, Y. Environmental Benefits of Macroalgae Products: A Case Study of Agar Based on Life Cycle Assessment. Algal Res. 2024, 78, 103384. [Google Scholar] [CrossRef]
- Sudhakar, M.P.; Kumar, B.R.; Mathimani, T.; Arunkumar, K. A Review on Bioenergy and Bioactive Compounds from Microalgae and Macroalgae-Sustainable Energy Perspective. J. Clean. Prod. 2019, 228, 1320–1333. [Google Scholar] [CrossRef]
- Sun, H.; Zhao, W.; Mao, X.; Li, Y.; Wu, T.; Chen, F. High-Value Biomass from Microalgae Production Platforms: Strategies and Progress Based on Carbon Metabolism and Energy Conversion. Biotechnol. Biofuels 2018, 11, 227. [Google Scholar] [CrossRef]
- Kostas, E.T.; Adams, J.M.M.; Ruiz, H.A.; Durán-Jiménez, G.; Lye, G.J. Macroalgal Biorefinery Concepts for the Circular Bioeconomy: A Review on Biotechnological Developments and Future Perspectives. Renew. Sustain. Energy Rev. 2021, 151, 111553. [Google Scholar] [CrossRef]
- Lawton, R.J.; Cole, A.J.; Roberts, D.A.; Paul, N.A.; de Nys, R. The Industrial Ecology of Freshwater Macroalgae for Biomass Applications. Algal Res. 2017, 24, 486–491. [Google Scholar] [CrossRef]
- Chemodanov, A.; Robin, A.; Golberg, A. Design of Marine Macroalgae Photobioreactor Integrated into Building to Support Seagriculture for Biorefinery and Bioeconomy. Bioresour. Technol. 2017, 241, 1084–1093. [Google Scholar] [CrossRef] [PubMed]
- Sebök, S.; Herppich, W.B.; Hanelt, D. Development of an Innovative Ring-Shaped Cultivation System for a Land-Based Cultivation of Marine Macroalgae. Aquac. Eng. 2017, 77, 33–41. [Google Scholar] [CrossRef]
- Dahiya, A. Chapter 13—Algae Biomass Cultivation for Advanced Biofuel Production. In Bioenergy, 2nd ed.; Dahiya, A., Ed.; Academic Press: Cambridge, MA, USA, 2020; pp. 245–266. ISBN 978-0-12-815497-7. [Google Scholar]
- Elsayed, K.N.M.; Kolesnikova, T.A.; Noke, A.; Klöck, G. Imaging the Accumulated Intracellular Microalgal Lipids as a Response to Temperature Stress. 3 Biotech 2017, 7, 41. [Google Scholar] [CrossRef] [PubMed]
- Traugott, H.; Zollmann, M.; Cohen, H.; Chemodanov, A.; Liberzon, A.; Golberg, A. Aeration and Nitrogen Modulated Growth Rate and Chemical Composition of Green Macroalgae Ulva sp. Cultured in a Photobioreactor. Algal Res. 2020, 47, 101808. [Google Scholar] [CrossRef]
- Korzen, L.; Pulidindi, I.N.; Israel, A.; Abelson, A.; Gedanken, A. Single Step Production of Bioethanol from the Seaweed Ulva rigida Using Sonication. RSC Adv. 2015, 5, 16223–16229. [Google Scholar] [CrossRef]
- Korzen, L.; Israel, A. Growth, Protein and Carbohydrate Contents in Ulva Rigida and Gracilaria Bursa-Pastoris Integrated with an Offshore Fish Farm. J. Appl. Phycol. 2015, 28, 1835–1845. [Google Scholar] [CrossRef]
- Dickson, R.; Liu, J.J. A Strategy for Advanced Biofuel Production and Emission Utilization from Macroalgal Biorefinery Using Superstructure Optimization. Energy 2021, 221, 119883. [Google Scholar] [CrossRef]
- Su, T.; Zhao, D.; Khodadadi, M.; Len, C. Lignocellulosic Biomass for Bioethanol: Recent Advances, Technology Trends, and Barriers to Industrial Development. Curr. Opin. Green Sustain. Chem. 2020, 24, 56–60. [Google Scholar] [CrossRef]
- Kechkar, M.; Sayed, W.; Cabrol, A.; Aziza, M.; Ahmed Zaid, T.; Amrane, A.; Djelal, H. Isolation and Identification of Yeast Strains from Sugarcane Molasses Dates and Figs for Ethanol Production Under Conditions Simulating Algal Hydrolysate. Braz. J. Chem. Eng. 2019, 36, 157–169. [Google Scholar] [CrossRef]
- Dave, N.; Selvaraj, R.; Varadavenkatesan, T.; Vinayagam, R. A Critical Review on Production of Bioethanol from Macroalgal Biomass. Algal Res. 2019, 42, 101606. [Google Scholar] [CrossRef]
- Polprasert, S.; Choopakar, O.; Elefsiniotis, P. Bioethanol Production from Pretreated Palm Empty Fruit Bunch (PEFB) Using Sequential Enzymatic Hydrolysis and Yeast Fermentation. Biomass Bioenergy 2021, 149, 106088. [Google Scholar] [CrossRef]
- Phaiboonsilpa, N.; Chysirichote, T.; Champreda, V.; Laosiripojana, N. Fermentation of Xylose, Arabinose, Glucose, Their Mixtures and Sugarcane Bagasse Hydrolyzate by Yeast Pichia stipitis for Ethanol Production. Energy Rep. 2020, 6, 710–713. [Google Scholar] [CrossRef]
- Sayed, W.; Cabrol, A.; Abdallah, R.; Taha, S.; Amrane, A.; Djelal, H. Enhancement of Ethanol Production from Synthetic Medium Model of Hydrolysate of Macroalgae. Renew. Energy 2018, 124, 3–10. [Google Scholar] [CrossRef]
- Acourene, A.; Ammouche, A. Optimization of Ethanol, Citric Acid, and α-Amylase Production from Date Wastes by Strains of Saccharomyces cerevisiae, Aspergillus niger, and Candida guilliermondii. J. Ind. Microbiol. Biotechnol. 2012, 39, 759–766. [Google Scholar] [CrossRef]
- Cunha-Pereira, F.d.; Hickert, L.R.; Rech, R.; Dillon, A.P.; Ayub, M.A.Z.; Cunha-Pereira, F.d.; Hickert, L.R.; Rech, R.; Dillon, A.P.; Ayub, M.A.Z. Fermentation of Hexoses and Pentoses from Hydrolyzed Soybean Hull into Ethanol and Xylitol by Candida guilliermondii BL 13. Brazilian J. Chem. Eng. 2017, 34, 927–936. [Google Scholar] [CrossRef]
- Prosky, L.; Asp, N.G.; Schweizer, T.F.; DeVries, J.W.; Furda, I. Determination of Insoluble, Soluble, and Total Dietary Fiber in Foods and Food Products: Interlaboratory Study. J. Assoc. Off. Anal. Chem. 1988, 71, 1017–1023. [Google Scholar] [CrossRef]
- Sáez-Plaza, P.; Navas, M.J.; Wybraniec, S.; Michałowski, T.; Garcia Asuero, A. An Overview of the Kjeldahl Method of Nitro-gen Determination. Part II. Sample Preparation, Working Scale, Instrumental Finish, and Quality Control. Crit. Rev. Anal. Chem. 2013, 43, 224–272. [Google Scholar] [CrossRef]
- Anechiţei, L.; Cojocaru, T.; Munteanu, G.; Bulgariu, L. Simple Methods for Quantitative Determination of Sulphate Ions from Aqueous Media with Industrial Applications. Bull. Polytech. Inst. Jassy Constructions. Archit. Sect. 2019, 65, 27–37. [Google Scholar]
- Djelal, H.; Larher, F.; Martin, G.; Amrane, A. Effect of the Dissolved Oxygen on the Bioproduction of Glycerol and Ethanol by Hansenula anomala Growing under Salt Stress Conditions. J. Biotechnol. 2006, 125, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Clark, A.E.; Kaleta, E.J.; Arora, A.; Wolk, D.M. Matrix-Assisted Laser Desorption Ionization–Time of Flight Mass Spectrometry: A Fundamental Shift in the Routine Practice of Clinical Microbiology. Clin. Microbiol. Rev. 2013, 26, 547–603. [Google Scholar] [CrossRef]
- Schultz-Jensen, N.; Thygesen, A.; Leipold, F.; Thomsen, S.T.; Roslander, C.; Lilholt, H.; Bjerre, A.B. Pretreatment of the Macroalgae Chaetomorpha linum for the Production of Bioethanol—Comparison of Five Pretreatment Technologies. Bioresour. Technol. 2013, 140, 36–42. [Google Scholar] [CrossRef]
- Nova, P.; Pimenta-Martins, A.; Maricato, É.; Nunes, C.; Abreu, H.; Coimbra, M.A.; Freitas, A.C.; Gomes, A.M. Chemical Composition and Antioxidant Potential of Five Algae Cultivated in Fully Controlled Closed Systems. Molecules 2023, 28, 4588. [Google Scholar] [CrossRef] [PubMed]
- Kidgell, J.T.; Magnusson, M.; de Nys, R.; Glasson, C.R.K. Ulvan: A Systematic Review of Extraction, Composition and Function. Algal Res. 2019, 39, 101422. [Google Scholar] [CrossRef]
- Paulert, R.; Ebbinghaus, D.; Urlass, C.; Moerschbacher, B.M. Priming of the Oxidative Burst in Rice and Wheat Cell Cultures by Ulvan, a Polysaccharide from Green Macroalgae, and Enhanced Resistance against Powdery Mildew in Wheat and Barley Plants. Plant Pathol. 2010, 59, 634–642. [Google Scholar] [CrossRef]
- Paradossi, G.; Cavalieri, F.; Chiessi, E. A Conformational Study on the Algal Polysaccharide Ulvan. Macromolecules 2002, 35, 6404–6411. [Google Scholar] [CrossRef]
- Trivedi, N.; Gupta, V.; Reddy, C.R.K.; Jha, B. Enzymatic Hydrolysis and Production of Bioethanol from Common Macrophytic Green Alga Ulva fasciata Delile. Bioresour. Technol. 2013, 150, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Toth, G.B.; Harrysson, H.; Wahlström, N.; Olsson, J.; Oerbekke, A.; Steinhagen, S.; Kinnby, A.; White, J.; Albers, E.; Edlund, U.; et al. Effects of Irradiance, Temperature, Nutrients, and pCO2 on the Growth and Biochemical Composition of Cultivated Ulva fenestrata. J. Appl. Phycol. 2020, 32, 3243–3254. [Google Scholar] [CrossRef]
- Bird, M.I.; Wurster, C.M.; de Paula Silva, P.H.; Bass, A.M.; de Nys, R. Algal Biochar—Production and Properties. Bioresour. Technol. 2011, 102, 1886–1891. [Google Scholar] [CrossRef]
- Tan, I.S.; Lam, M.K.; Lee, K.T. Hydrolysis of Macroalgae Using Heterogeneous Catalyst for Bioethanol Production. Carbohydr. Polym. 2013, 94, 561–566. [Google Scholar] [CrossRef]
- Yeon, J.-H.; Lee, S.-E.; Choi, W.Y.; Kang, D.H.; Lee, H.-Y.; Jung, K.-H. Repeated-Batch Operation of Surface-Aerated Fermentor for Bioethanol Production from the Hydrolysate of Seaweed Sargassum sagamianum. J. Microbiol. Biotechnol. 2011, 21, 323–331. [Google Scholar] [CrossRef]
- Kumar, S.; Gupta, R.; Kumar, G.; Sahoo, D.; Kuhad, R.C. Bioethanol Production from Gracilaria verrucosa, a Red Alga, in a Biorefinery Approach. Bioresour. Technol. 2013, 135, 150–156. [Google Scholar] [CrossRef]
- Lee, S.-E.; Lee, J.-E.; Shin, G.-Y.; Choi, W.Y.; Kang, D.H.; Lee, H.-Y.; Jung, K.-H. Development of a Practical and Cost-Effective Medium for Bioethanol Production from the Seaweed Hydrolysate in Surface-Aerated Fermentor by Repeated-Batch Operation. J. Microbiol. Biotechnol. 2012, 22, 107–113. [Google Scholar] [CrossRef]
- Trivedi, N.; Reddy, C.R.K.; Radulovich, R.; Jha, B. Solid State Fermentation (SSF)-Derived Cellulase for Saccharification of the Green Seaweed Ulva for Bioethanol Production. Algal Res. 2015, 9, 48–54. [Google Scholar] [CrossRef]
- Choi, W.Y.; Han, J.G.; Lee, C.G.; Song, C.H.; Kim, J.S.; Seo, Y.C.; Lee, S.E.; Jung, K.H.; Kang, D.H.; Heo, S.J.; et al. Bioethanol Production from Ulva Pertusa Kjellman by High-Temperature Liquefaction. Chem. Biochem. Eng. Q. 2012, 26, 15–21. [Google Scholar]
- Valente, L.M.P.; Gouveia, A.; Rema, P.; Matos, J.; Gomes, E.F.; Pinto, I.S. Evaluation of Three Seaweeds Gracilaria Bursa-Pastoris, Ulva rigida and Gracilaria cornea as Dietary Ingredients in European Sea Bass (Dicentrarchus labrax) Juveniles. Aquaculture 2006, 252, 85–91. [Google Scholar] [CrossRef]
- Garg, S.K.; Jain, A. Fermentative Production of 2,3-Butanediol: A Review. Bioresour. Technol. 1995, 51, 103–109. [Google Scholar] [CrossRef]
- Grover, B.P.; Garg, S.K.; Verma, J. Production of 2,3-Butanediol from Wood Hydrolysate by Klebsiella pneumoniae. World J. Microbiol. Biotechnol. 1990, 6, 328–332. [Google Scholar] [CrossRef]
- Perego, P.; Converti, A.; Del Borghi, M. Effects of Temperature, Inoculum Size and Starch Hydrolyzate Concentration on Butanediol Production by Bacillus licheniformis. Bioresour. Technol. 2003, 89, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Maiorella, B.L.; Blanch, H.W.; Wilke, C.R. Economic Evaluation of Alternative Ethanol Fermentation Processes. Biotechnol. Bioeng. 1984, 26, 1003–1025. [Google Scholar] [CrossRef] [PubMed]
- Sakai, K.; Ezaki, Y. Open L-Lactic Acid Fermentation of Food Refuse Using Thermophilic Bacillus coagulans and Fluorescence in Situ Hybridization Analysis of Microflora. J. Biosci. Bioeng. 2006, 101, 457–463. [Google Scholar] [CrossRef]
- Sarris, D.; Giannakis, M.; Philippoussis, A.; Komaitis, M.; Koutinas, A.A.; Papanikolaou, S. Conversions of Olive Mill Wastewater-Based Media by Saccharomyces cerevisiae through Sterile and Non-Sterile Bioprocesses. J. Chem. Technol. Biotechnol. 2013, 88, 958–969. [Google Scholar] [CrossRef]
- Tao, F.; Miao, J.Y.; Shi, G.Y.; Zhang, K.C. Ethanol Fermentation by an Acid-Tolerant Zymomonas mobilis under Non-Sterilized Condition. Process Biochem. 2005, 40, 183–187. [Google Scholar] [CrossRef]
- Lin, Y.; Tanaka, S. Ethanol Fermentation from Biomass Resources: Current State and Prospects. Appl. Microbiol. Biotechnol. 2006, 69, 627–642. [Google Scholar] [CrossRef]
- Wu, W.-H.; Hung, W.-C.; Lo, K.-Y.; Chen, Y.-H.; Wan, H.-P.; Cheng, K.-C. Bioethanol Production from Taro Waste Using Thermo-Tolerant Yeast Kluyveromyces marxianus K21. Bioresour. Technol. 2016, 201, 27–32. [Google Scholar] [CrossRef]
- Schirmer-Michel, Â.C.; Flôres, S.H.; Hertz, P.F.; Matos, G.S.; Ayub, M.A.Z. Production of Ethanol from Soybean Hull Hydrolysate by Osmotolerant Candida guilliermondii NRRL Y-2075. Bioresour. Technol. 2008, 99, 2898–2904. [Google Scholar] [CrossRef]
- Silva, V.F.; Arruda, P.V.; Felipe, M.G.; Gonçalves, A.R.; Rocha, G.J. Fermentation of Cellulosic Hydrolysates Obtained by Enzymatic Saccharification of Sugarcane Bagasse Pretreated by Hydrothermal Processing. J. Ind. Microbiol. Biotechnol. 2011, 38, 809–817. [Google Scholar] [CrossRef] [PubMed]
- Grootjen, D.R.J.; Meijlink, L.H.H.M.; van der Lans, R.G.J.M.; Luyben, K.C.A.M. Cofermentation of Glucose and Xylose with Immobilized Pichia stipitis and Saccharomyces cerevisiae. Enzym. Microb. Technol. 1990, 12, 860–864. [Google Scholar] [CrossRef]
- Hahn-Hägerdal, B.; Jeppsson, H.; Skoog, K.; Prior, B.A. Biochemistry and Physiology of Xylose Fermentation by Yeasts. Enzym. Microb. Technol. 1994, 16, 933–943. [Google Scholar] [CrossRef]
- Ligthelm, M.E.; Prior, B.A.; Preez, J.C. du The Oxygen Requirements of Yeasts for the Fermentation of D-Xylose and d-Glucose to Ethanol. Appl. Microbiol. Biotechnol. 1988, 28, 63–68. [Google Scholar] [CrossRef]
- Skoog, K.; Hahn-Hägerdal, B. Effect of Oxygenation on Xylose Fermentation by Pichia stipitis. Appl. Environ. Microbiol. 1990, 56, 3389–3394. [Google Scholar] [CrossRef] [PubMed]
- Rouhollah, H.; Iraj, N.; Giti, E.; Sorah, A. Mixed Sugar Fermentation by Pichia stipitis, Sacharomyces cerevisiaea, and an Isolated Xylose fermenting Kluyveromyces marxianus and Their Cocultures. Afr. J. Biotechnol. 2007, 6, 9. [Google Scholar]
- Yanagisawa, M.; Nakamura, K.; Ariga, O.; Nakasaki, K. Production of High Concentrations of Bioethanol from Seaweeds That Contain Easily Hydrolyzable Polysaccharides. Process Biochem. 2011, 46, 2111–2116. [Google Scholar] [CrossRef]
- Borines, M.G.; de Leon, R.L.; Cuello, J.L. Bioethanol Production from the macroalgae Sargassum spp. Bioresour. Technol. 2013, 138, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Kuhad, R.C.; Mehta, G.; Gupta, R.; Sharma, K.K. Fed Batch Enzymatic Saccharification of Newspaper Cellulosics Improves the Sugar Content in the Hydrolysates and Eventually the Ethanol Fermentation by Saccharomyces cerevisiae. Biomass Bioenergy 2010, 34, 1189–1194. [Google Scholar] [CrossRef]
- Mishra, A.; Sharma, A.K.; Sharma, S.; Bagai, R.; Mathur, A.S.; Gupta, R.P.; Tuli, D.K. Lignocellulosic Ethanol Production Employing Immobilized Saccharomyces cerevisiae in Packed Bed Reactor. Renew. Energy 2016, 98, 57–63. [Google Scholar] [CrossRef]
- El Harchi, M.; Fakihi Kachkach, F.Z.; El Mtili, N. Optimization of Thermal Acid Hydrolysis for Bioethanol Production from Ulva Rigida with Yeast Pachysolen tannophilus. S. Afr. J. Bot. 2018, 115, 161–169. [Google Scholar] [CrossRef]
- Khambhaty, Y.; Mody, K.; Gandhi, M.R.; Thampy, S.; Maiti, P.; Brahmbhatt, H.; Eswaran, K.; Ghosh, P.K. Kappaphycus alvarezii as a Source of Bioethanol. Bioresour. Technol. 2012, 103, 180–185. [Google Scholar] [CrossRef]
- Ben Yahmed, N.; Jmel, M.A.; Ben Alaya, M.; Bouallagui, H.; Marzouki, M.N.; Smaali, I. A Biorefinery Concept Using the Green Macroalgae Chaetomorpha linum for the Coproduction of Bioethanol and Biogas. Energy Convers. Manag. 2016, 119, 257–265. [Google Scholar] [CrossRef]
- Zhang, W.; Mao, Y.; Liu, Z.; Wang, M. Ethanol Production from Colpomenia sinuosa by an Alginate Fermentation Strain Meyerozyma guilliermondii. Indian J. Microbiol. 2022, 62, 112–122. [Google Scholar] [CrossRef]
- Wu, F.-C.; Wu, J.-Y.; Liao, Y.-J.; Wang, M.-Y.; Shih, I.-L. Sequential Acid and Enzymatic Hydrolysis in Situ and Bioethanol Production from Gracilaria Biomass. Bioresour. Technol. 2014, 156, 123–131. [Google Scholar] [CrossRef] [PubMed]

 ) and the synthetic medium (
) and the synthetic medium ( ).
) and the synthetic medium ().
).
) and the synthetic medium ().




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Proximate Composition | Relative % (Dry Weight Basis/Brut) |
|---|---|
| Dry matter | 94.9 ± 0.05 |
| Fibers | 34.90 ± 0.05 |
| Protein | 5.68 ± 0.05 |
| Ash | 31.9 ± 0.05 |
| N Kjeldahl | 0.91 ± 0.05 |
| Total sulfates | 7.8 ± 0.05 |
| Carbohydrate Content | Glucose | Rhamnose | Xylose | Iduronic Acid | Glucuronic Acid |
|---|---|---|---|---|---|
| Relative % (dry weight basis/brut) | 25.6 ± 0.05 | 6.5 ± 0.05 | 2.1 ± 0.05 | 0.6 ± 0.05 | 2.8 ± 0.05 |
| Conditions | Cell Density (600 nm) | Ethanol (g·L−1) | Productivity (g·L−1·h−1) | Glycerol Yield (%) (c/c) | Ethanol Yields (%) (c/c) | Efficiency (%) |
|---|---|---|---|---|---|---|
| Without nutrients, ammonium chloride and peptone | 9.35 ±0.05 | 9.55 ± 0.05 | 0.20 ± 0.05 | 5.87 ± 0.05 | 56.83 ± 0.05 | 85.33 ± 0.05 |
| With nutrients and ammonium chloride | 9.10 ±0.05 | 9.41 ± 0.05 | 0.20 ± 0.05 | 6.87 ± 0.05 | 55.92 ± 0.05 | 83.96 ± 0.05 |
| With peptone | 9.30 ±0.05 | 9.40 ± 0.05 | 0.20 ± 0.05 | 6.03 ± 0.05 | 55.79 ± 0.05 | 83.76 ± 0.05 |
| Inoculum Size % (v/v) | Ethanol (g·L−1) | Sugar Consumed % | Productivity g·L−1·h−1 | Glucose Consumption Rate g·L−1·h−1 | Y Ethanol (%) (c/c) | (Ethanol)o/t Ratio (%) |
|---|---|---|---|---|---|---|
| 0.1 | 5.54 ± 0.05 | 58 ± 0.05 | 0.23 ± 0.05 | 0.54 ± 0.05 | 56.19 ± 0.05 | 84.38 ± 0.05 |
| 1 | 9.64 ± 0.05 | 100 ± 0.05 | 0.40 ± 0.05 | 0.95 ± 0.05 | 55.20 ± 0.05 | 82.89 ± 0.05 |
| Composition % (Dry Weight/Dry Weight) | ||||||
|---|---|---|---|---|---|---|
| Ash | Moisture | Lipids | Total Nitrogen | Carbohydrates | Calcium (mg/100 g) | |
| Hydrolysate | 5.63 ± 0.05 | 88.8 ± 0.05 | 0.3 ± 0.05 | 0.4 ± 0.05 | 4.9 ± 0.05 | 55.2 ± 0.05 |
| Autoclaved hydrolysate | 5.52 ± 0.05 | 89.2 ± 0.05 | 0.1 ± 0.05 | 0.4 ± 0.05 | 4.8 ± 0.05 | 54.0 ± 0.05 |
| Glucose Consumption Rate at 48 h (g·L−1·h−1) | Ethanol Production Rate (g·L−1·h−1) | (Ethanol) o/t Ratio | ||||
|---|---|---|---|---|---|---|
| Hydrolysate | Synthetic Medium | Hydrolysate | Synthetic Medium | Hydrolysate | Synthetic Medium | |
| K. marxianus | 0.49 ± 0.05 | 0.43 ± 0.05 | 0.18 ± 0.05 | 0.14 ± 0.05 | 70.49 ± 0.05 | 64.10 ± 0.05 |
| P. stipitis | 0.10 ± 0.05 | 0.41 ± 0.05 | 0.03 ± 0.05 | 0.10 ± 0.05 | 52.25 ± 0.05 | 49.37 ± 0.05 |
| C. guilliermondii | 0.35 ± 0.05 | 0.43 ± 0.05 | 0.11 ± 0.05 | 0.09 ± 0.05 | 63.20 ± 0.05 | 42.97 ± 0.05 |
| S. cerevisiae | 0.47 ± 0.05 | 0.42 ± 0.05 | 0.20 ± 0.05 | 0.16 ± 0.05 | 82.87 ± 0.05 | 74.35 ± 0.05 |
| Algae | Fermenting Strain | Enrichment | Size of Inoculum | Ethanol Yield | References |
|---|---|---|---|---|---|
| U. rigida | S. cerevisiae CLIB 95 | Without enrichment | 1% v/v (0.12 g) | 0.54% (c/c) (0.41 g·g−1 reducing sugar) | Present study |
| U. rigida | Pachysolen tannophilus | - | 5% | 0.37 g·g−1 | [103] |
| Colpomenia sinuosa | Meyerozyma guilliermondii, | - | 10% | 0.26 g·g−1 | [106] |
| Kappaphycus alvarezii | S. cerevisiae (NCIM 3455) | With nitrogen source | 5% v/v | 0.39 g·g−1 | [104] |
| Chaetomorpha linum | S. cerevisiae (Baker’s yeast) | Yeast extract Peptone medium | 10% v/v | 0.41 g·g−1 | [105] |
| U. rigida | S. cerevisiae (beaker) | - | 0.5 g | 333.3 mg·g−1 | [52] |
| Gracilaria sp. | S. cerevisiae | 2 g·L−1 yeast extract | 10% v/v | 0.47 g·g−1 | [107] |
| Gracilaria verrucosa | S. cerevisiae | 3 g·L−1 yeast extract and 0.25 g·L−1 (NH4)2HPO4 | 6% v/v | 0.43 g·g-−1 | [78] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sayed, W.; Cabrol, A.; Salma, A.; Amrane, A.; Benoit, M.; Pierre, R.; Djelal, H. Green Macroalgae Hydrolysate for Biofuel Production: Potential of Ulva rigida. Appl. Microbiol. 2024, 4, 563-581. https://doi.org/10.3390/applmicrobiol4020039
Sayed W, Cabrol A, Salma A, Amrane A, Benoit M, Pierre R, Djelal H. Green Macroalgae Hydrolysate for Biofuel Production: Potential of Ulva rigida. Applied Microbiology. 2024; 4(2):563-581. https://doi.org/10.3390/applmicrobiol4020039
Chicago/Turabian StyleSayed, Walaa, Audrey Cabrol, Alaa Salma, Abdeltif Amrane, Maud Benoit, Ronan Pierre, and Hayet Djelal. 2024. "Green Macroalgae Hydrolysate for Biofuel Production: Potential of Ulva rigida" Applied Microbiology 4, no. 2: 563-581. https://doi.org/10.3390/applmicrobiol4020039
APA StyleSayed, W., Cabrol, A., Salma, A., Amrane, A., Benoit, M., Pierre, R., & Djelal, H. (2024). Green Macroalgae Hydrolysate for Biofuel Production: Potential of Ulva rigida. Applied Microbiology, 4(2), 563-581. https://doi.org/10.3390/applmicrobiol4020039