




Addressing Inequality in the COVID-19 Pandemic in Africa: A Snapshot from Clinical Symptoms to Vaccine Distribution



Abstract
:1. COVID-19 in Africa
2. SARS-CoV-2
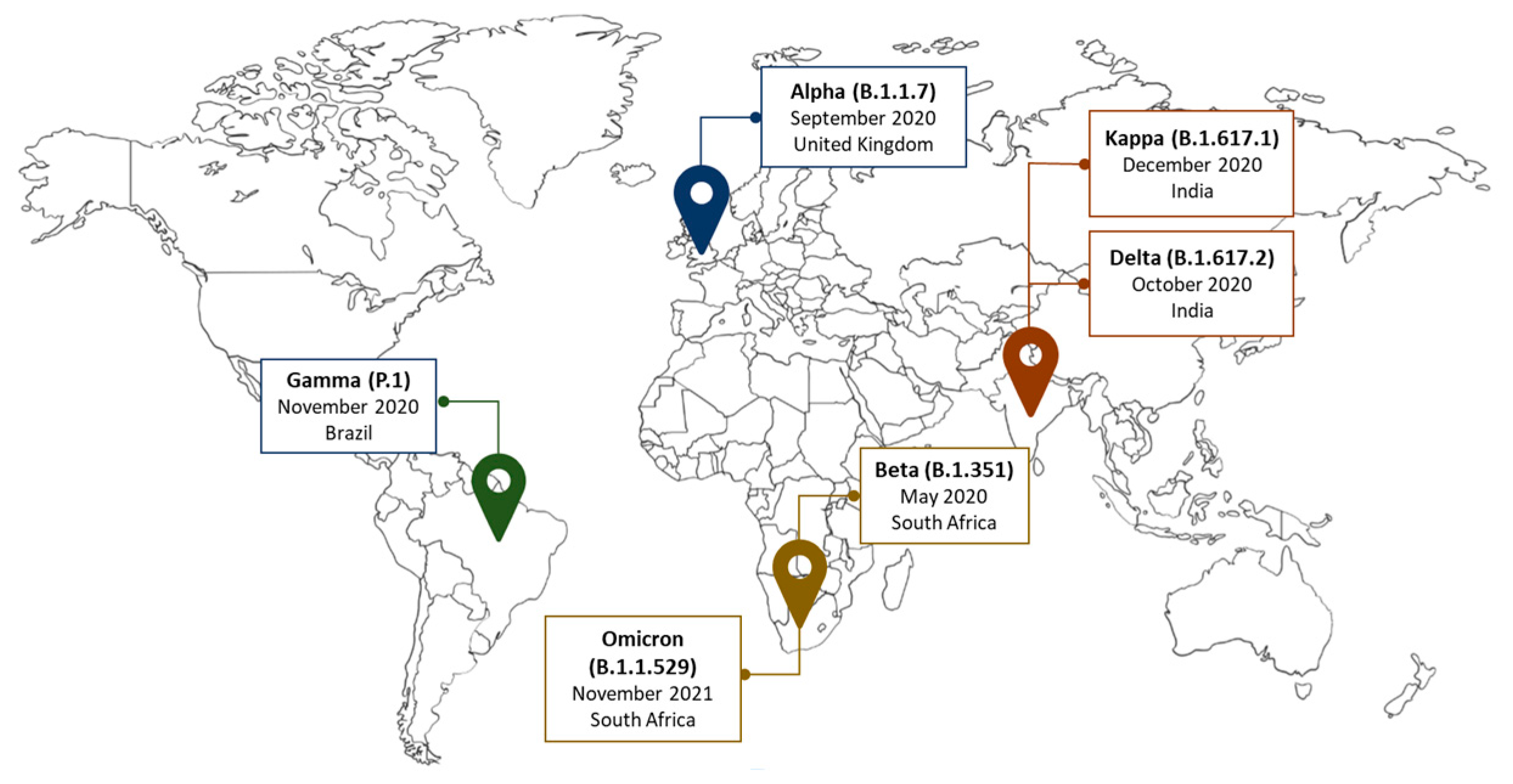
3. Genetic Variants of SARS-CoV-2 Reported in Africa
4. The Transmissibility of SARS-CoV-2 and the Difficulty of Applying Standardized Protection Measures in Africa
5. The Sequelae Caused by SARS-CoV-2
6. The Comorbidities Underlying COVID-19 Severity in Africa
7. The Immune Response to SARS-CoV-2 in Africa
8. Vaccine Development
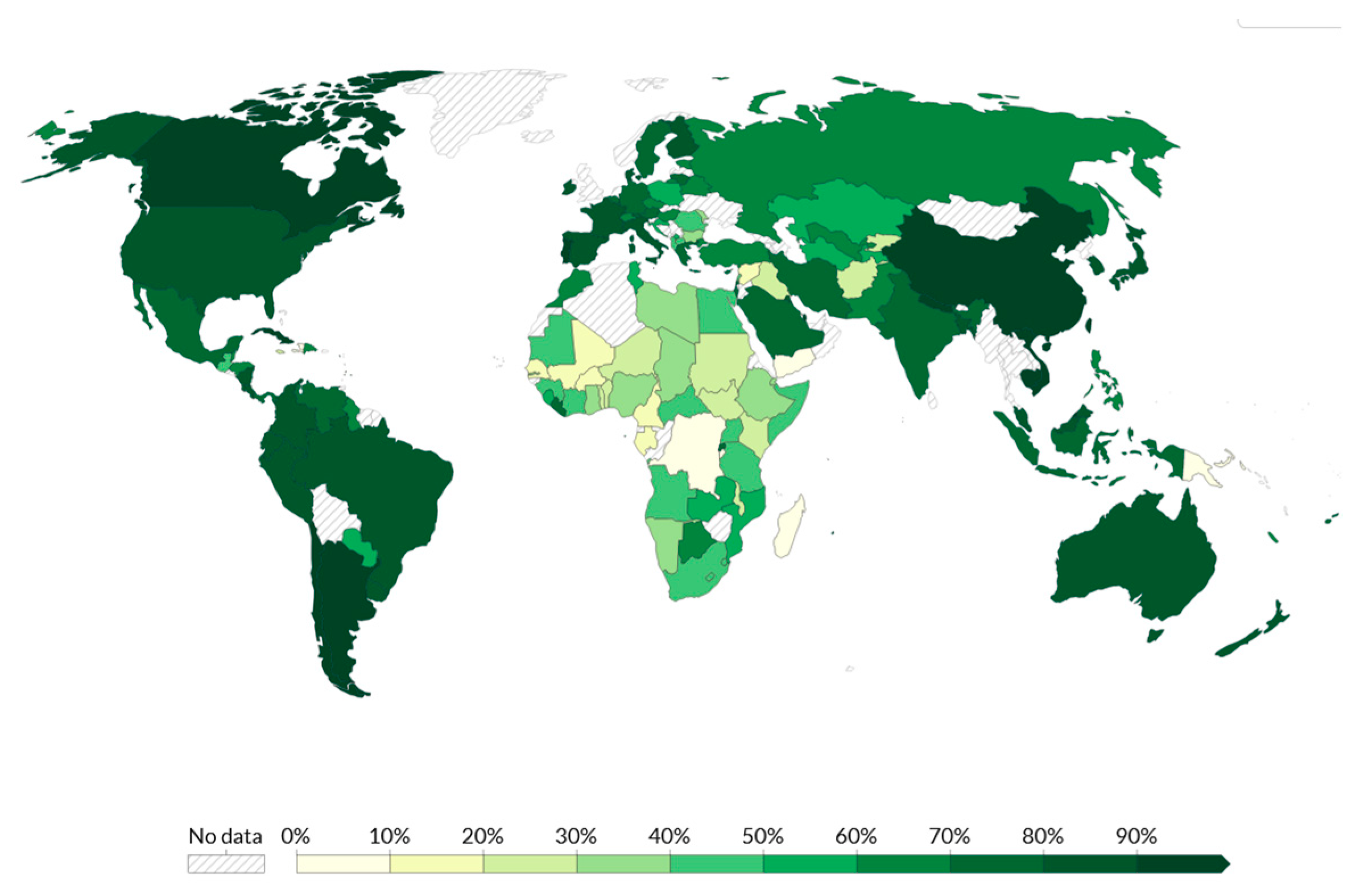
9. Vaccine Inequity Distribution in Africa
10. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- United Nations World Population Prospects 2019: Highlights. Available online: https://population.un.org/wpp/Publications/Files/wpp2019_10KeyFindings.pdf (accessed on 9 April 2023).
- The Fund for Peace Global Data. Available online: https://fragilestatesindex.org/global-data/ (accessed on 19 March 2023).
- Bwire, G.; Ario, A.R.; Eyu, P.; Ocom, F.; Wamala, J.F.; Kusi, K.A.; Ndeketa, L.; Jambo, K.C.; Wanyenze, R.K.; Talisuna, A.O. The COVID-19 Pandemic in the African Continent. BMC Med. 2022, 20, 167. [Google Scholar] [CrossRef]
- Udoakang, A.; Oboh, M.; Henry-Ajala, A.; Anyigba, C.; Omoleke, S.; Amambua-Ngwa, A.; Paemka, L.; Awandare, G.; Quashie, P. Low COVID-19 Impact in Africa: The Multifactorial Nexus. Open Res. Afr. 2021, 4, 47. [Google Scholar] [CrossRef]
- Cilliers, J.; Kwasi, S.; Yeboua, K.; Oosthuizen, M.; Alexander, K.; Pooe, T.; Moyer, J. Impact of COVID-19 in Africa: A Scenario Analysis to 2030. SSRN Electron. J. 2020. [Google Scholar] [CrossRef]
- Dong, E.; Du, H.; Gardner, L. An Interactive Web-Based Dashboard to Track COVID-19 in Real Time. Lancet Infect. Dis. 2020, 20, 533–534. [Google Scholar] [CrossRef] [PubMed]
- El-Sadr, W.M.; Justman, J. Africa in the Path of COVID-19. N. Engl. J. Med. 2020, 383, e11. [Google Scholar] [CrossRef]
- Mutombo, P.N.; Fallah, M.P.; Munodawafa, D.; Kabel, A.; Houeto, D.; Goronga, T.; Mweemba, O.; Balance, G.; Onya, H.; Kamba, R.S.; et al. COVID-19 Vaccine Hesitancy in Africa: A Call to Action. Lancet Glob. Health 2022, 10, e320–e321. [Google Scholar] [CrossRef] [PubMed]
- Hussein, M.I.H.; Albashir, A.A.D.; Elawad, O.A.M.A.; Homeida, A. Malaria and COVID-19: Unmasking Their Ties. Malar. J. 2020, 19, 457. [Google Scholar] [CrossRef] [PubMed]
- Yazdanbakhsh, M.; Kremsner, P.G.; Van Ree, R. Immunology: Allergy, Parasites, and the Hygiene Hypothesis. Science 2002, 296, 490–494. [Google Scholar] [CrossRef]
- Arts, R.J.W.; Moorlag, S.J.C.F.M.; Novakovic, B.; Li, Y.; Wang, S.Y.; Oosting, M.; Kumar, V.; Xavier, R.J.; Wijmenga, C.; Joosten, L.A.B.; et al. BCG Vaccination Protects against Experimental Viral Infection in Humans through the Induction of Cytokines Associated with Trained Immunity. Cell Host Microbe 2018, 23, 89–100.e5. [Google Scholar] [CrossRef] [PubMed]
- Rolot, M.; Dougall, A.M.; Chetty, A.; Javaux, J.; Chen, T.; Xiao, X.; Machiels, B.; Selkirk, M.E.; Maizels, R.M.; Hokke, C.; et al. Helminth-Induced IL-4 Expands Bystander Memory CD8+ T Cells for Early Control of Viral Infection. Nat. Commun. 2018, 9, 4516. [Google Scholar] [CrossRef]
- Boum, Y.; Bebell, L.M.; Bisseck, A.-C.Z.-K. Africa Needs Local Solutions to Face the COVID-19 Pandemic. Lancet 2021, 397, 1238–1240. [Google Scholar] [CrossRef] [PubMed]
- Nguimkeu, P.; Tadadjeu, S. Why Is the Number of COVID-19 Cases Lower than Expected in Sub-Saharan Africa? A Cross-Sectional Analysis of the Role of Demographic and Geographic Factors. World Dev. 2021, 138, 105251. [Google Scholar] [CrossRef] [PubMed]
- Mwiinde, A.M.; Siankwilimba, E.; Sakala, M.; Banda, F.; Michelo, C. Climatic and Environmental Factors Influencing COVID-19 Transmission—An African Perspective. Trop. Med. Infect. Dis. 2022, 7, 433. [Google Scholar] [CrossRef]
- Chan, K.H.; Peiris, J.S.M.; Lam, S.Y.; Poon, L.L.M.; Yuen, K.Y.; Seto, W.H. The Effects of Temperature and Relative Humidity on the Viability of the SARS Coronavirus. Adv. Virol. 2011, 2011, 734690. [Google Scholar] [CrossRef]
- Sharif, N.; Sarkar, M.K.; Ahmed, S.N.; Ferdous, R.N.; Nobel, N.U.; Parvez, A.K.; Talukder, A.A.; Dey, S.K. Environmental Correlation and Epidemiologic Analysis of COVID-19 Pandemic in Ten Regions in Five Continents. Heliyon 2021, 7, e06576. [Google Scholar] [CrossRef]
- Whittemore, P.B. COVID-19 Fatalities, Latitude, Sunlight, and Vitamin D. Am. J. Infect. Control 2020, 48, 1042–1044. [Google Scholar] [CrossRef]
- Shao, L.; Ge, S.; Jones, T.; Santosh, M.; Silva, L.F.O.; Cao, Y.; Oliveira, M.L.S.; Zhang, M.; BéruBé, K. The Role of Airborne Particles and Environmental Considerations in the Transmission of SARS-CoV-2. Geosci. Front. 2021, 12, 101189. [Google Scholar] [CrossRef]
- Adekunle, I.A.; Tella, S.A.; Oyesiku, K.O.; Oseni, I.O. Spatio-Temporal Analysis of Meteorological Factors in Abating the Spread of COVID-19 in Africa. Heliyon 2020, 6, e04749. [Google Scholar] [CrossRef]
- Salyer, S.J.; Maeda, J.; Sembuche, S.; Kebede, Y.; Tshangela, A.; Moussif, M.; Ihekweazu, C.; Mayet, N.; Abate, E.; Ouma, A.O.; et al. The First and Second Waves of the COVID-19 Pandemic in Africa: A Cross-Sectional Study. Lancet 2021, 397, 1265–1275. [Google Scholar] [CrossRef]
- World Health Organization (WHO). WHO Coronavirus (COVID-19) Dashboard. Available online: https://covid19.who.int/ (accessed on 3 November 2021).
- Diop, B.Z.; Ngom, M.; Pougué Biyong, C.; Pougué Biyong, J.N. The Relatively Young and Rural Population May Limit the Spread and Severity of COVID-19 in Africa: A Modelling Study. BMJ Glob. Health 2020, 5, e002699. [Google Scholar] [CrossRef]
- World Data International Tourism: The Most Popular Countries. Available online: https://www.worlddata.info/tourism.php (accessed on 9 April 2023).
- Bradshaw, D.; Dorrington, R.; Moultrie, T.; Groenewald, P.; Moultrie, H. Underestimated COVID-19 Mortality in WHO African Region. Lancet Glob. Health 2022, 10, e1559. [Google Scholar] [CrossRef]
- Chitungo, I.; Dzobo, M.; Hlongwa, M.; Dzinamarira, T. COVID-19: Unpacking the Low Number of Cases in Africa. Public Health Pract. 2020, 1, 100038. [Google Scholar] [CrossRef] [PubMed]
- Lucero-Prisno, D.E.; Adebisi, Y.A.; Lin, X. Current Efforts and Challenges Facing Responses to 2019-NCoV in Africa. Glob. Health Res. Policy 2020, 5, 21. [Google Scholar] [CrossRef]
- Jensen, N.; Kelly, A.H.; Avendano, M. The COVID-19 Pandemic Underscores the Need for an Equity-Focused Global Health Agenda. Humanit. Soc. Sci. Commun. 2021, 8, 15. [Google Scholar] [CrossRef]
- Smolen, K.K.; Cai, B.; Gelinas, L.; Edgardo, S.; Iii, F.; Larsen, M.; Speert, D.P.; Cooper, P.J.; Esser, M.; Marchant, A.; et al. Single-Cell Analysis of Innate Cytokine Responses to Pattern Recognition Receptor Stimulation in Children across Four Continents. J. Immunol. 2014, 193, 3003–3012. [Google Scholar] [CrossRef] [PubMed]
- Carlson, C.J.; Zipfel, C.M.; Garnier, R.; Bansal, S. Global Estimates of Mammalian Viral Diversity Accounting for Host Sharing. Nat. Ecol. Evol. 2019, 3, 1070–1075. [Google Scholar] [CrossRef] [PubMed]
- Olival, K.J.; Hosseini, P.R.; Zambrana-Torrelio, C.; Ross, N.; Bogich, T.L.; Daszak, P. Host and Viral Traits Predict Zoonotic Spillover from Mammals. Nature 2017, 546, 646–650. [Google Scholar] [CrossRef] [PubMed]
- Pekar, J.E.; Magee, A.; Parker, E.; Moshiri, N.; Izhikevich, K.; Havens, J.L.; Gangavarapu, K.; Malpica Serrano, L.M.; Crits-Christoph, A.; Matteson, N.L.; et al. The Molecular Epidemiology of Multiple Zoonotic Origins of SARS-CoV-2. Science 2022, 377, 960–966. [Google Scholar] [CrossRef] [PubMed]
- Weiss, S.R.; Leibowitz, J.L. Coronavirus Pathogenesis; Maramorosch, K., Shatkin, A.J., Murphy, A.F., Eds.; Academic Press: Cambridge, MA, USA, 2011; Volume 81, pp. 85–164. [Google Scholar]
- Li, H.; Liu, S.-M.; Yu, X.-H.; Tang, S.-L.; Tang, C.-K. Coronavirus Disease 2019 (COVID-19): Current Status and Future Perspectives. Int. J. Antimicrob. Agents 2020, 55, 105951. [Google Scholar] [CrossRef]
- Chauhan, S. Comprehensive Review of Coronavirus Disease 2019 (COVID-19). Biomed. J. 2020, 43, 334–340. [Google Scholar] [CrossRef]
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R.; et al. A Novel Coronavirus from Patients with Pneumonia in China, 2019. N. Engl. J. Med. 2020, 382, 727–733. [Google Scholar] [CrossRef]
- Letko, M.; Marzi, A.; Munster, V. Functional Assessment of Cell Entry and Receptor Usage for SARS-CoV-2 and Other Lineage B Betacoronaviruses. Nat. Microbiol. 2020, 5, 562–569. [Google Scholar] [CrossRef]
- Wan, Y.; Shang, J.; Graham, R.; Baric, R.S.; Li, F. Receptor Recognition by the Novel Coronavirus from Wuhan: An Analysis Based on Decade-Long Structural Studies of SARS Coronavirus. J. Virol. 2020, 94, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Wu, A.; Peng, Y.; Huang, B.; Ding, X.; Wang, X.; Niu, P.; Meng, J.; Zhu, Z.; Zhang, Z.; Wang, J.; et al. Genome Composition and Divergence of the Novel Coronavirus (2019-NCoV) Originating in China. Cell Host Microbe 2020, 27, 325–328. [Google Scholar] [CrossRef] [PubMed]
- Li, F. Receptor Recognition Mechanisms of Coronaviruses: A Decade of Structural Studies. J. Virol. 2015, 89, 1954–1964. [Google Scholar] [CrossRef] [PubMed]
- Shang, J.; Wan, Y.; Luo, C.; Ye, G.; Geng, Q.; Auerbach, A.; Li, F. Cell Entry Mechanisms of SARS-CoV-2. Proc. Natl. Acad. Sci. USA 2020, 117, 11727. [Google Scholar] [CrossRef] [PubMed]
- Harrison, A.G.; Lin, T.; Wang, P. Mechanisms of SARS-CoV-2 Transmission and Pathogenesis. Trends Immunol. 2020, 41, 1100–1115. [Google Scholar] [CrossRef] [PubMed]
- Jackson, C.B.; Farzan, M.; Chen, B.; Choe, H. Mechanisms of SARS-CoV-2 Entry into Cells. Nat. Rev. Mol. Cell Biol. 2022, 23, 3–20. [Google Scholar] [CrossRef] [PubMed]
- Artika, I.M.; Dewantari, A.K.; Wiyatno, A. Molecular Biology of Coronaviruses: Current Knowledge. Heliyon 2020, 6, e04743. [Google Scholar] [CrossRef]
- de Wit, E.; van Doremalen, N.; Falzarano, D.; Munster, V.J. SARS and MERS: Recent Insights into Emerging Coronaviruses. Nat. Rev. Microbiol. 2016, 14, 523–534. [Google Scholar] [CrossRef]
- Florindo, H.F.; Kleiner, R.; Vaskovich-Koubi, D.; Acúrcio, R.C.; Carreira, B.; Yeini, E.; Tiram, G.; Liubomirski, Y.; Satchi-Fainaro, R. Immune-Mediated Approaches against COVID-19. Nat. Nanotechnol. 2020, 15, 630–645. [Google Scholar] [CrossRef]
- Li, X.; Geng, M.; Peng, Y.; Meng, L.; Lu, S. Molecular Immune Pathogenesis and Diagnosis of COVID-19. J. Pharm. Anal. 2020, 10, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Mascola, J.R.; Graham, B.S.; Fauci, A.S. SARS-CoV-2 Viral Variants—Tackling a Moving Target. JAMA 2021, 325, 1261–1262. [Google Scholar] [CrossRef]
- Prévost, J.; Finzi, A. The Great Escape? SARS-CoV-2 Variants Evading Neutralizing Responses. Cell Host Microbe 2021, 29, 322–324. [Google Scholar] [CrossRef] [PubMed]
- Tao, K.; Tzou, P.L.; Nouhin, J.; Gupta, R.K.; de Oliveira, T.; Kosakovsky Pond, S.L.; Fera, D.; Shafer, R.W. The Biological and Clinical Significance of Emerging SARS-CoV-2 Variants. Nat. Rev. Genet. 2021, 22, 757–773. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO) Tracking SARS-CoV-2 Variants. Available online: https://www.who.int/activities/tracking-SARS-CoV-2-variants (accessed on 24 January 2024).
- Tessema, S.K.; Nkengasong, J.N. Understanding COVID-19 in Africa. Nat. Rev. Immunol. 2021, 21, 469–470. [Google Scholar] [CrossRef]
- Viana, R.; Moyo, S.; Amoako, D.G.; Tegally, H.; Scheepers, C.; Althaus, C.L.; Anyaneji, U.J.; Bester, P.A.; Boni, M.F.; Chand, M.; et al. Rapid Epidemic Expansion of the SARS-CoV-2 Omicron Variant in Southern Africa. Nature 2022, 603, 679–686. [Google Scholar] [CrossRef]
- Gao, S.-J.; Guo, H.; Luo, G. Omicron Variant (B.1.1.529) of SARS-CoV-2, a Global Urgent Public Health Alert! J. Med. Virol. 2022, 94, 1255–1256. [Google Scholar] [CrossRef]
- Chang, D.; Chang, X.; He, Y.; Tan, K.J.K. The Determinants of COVID-19 Morbidity and Mortality across Countries. Sci. Rep. 2022, 12, 5888. [Google Scholar] [CrossRef]
- Ma, Q.; Liu, J.; Liu, Q.; Kang, L.; Liu, R.; Jing, W.; Wu, Y.; Liu, M. Global Percentage of Asymptomatic SARS-CoV-2 Infections Among the Tested Population and Individuals with Confirmed COVID-19 Diagnosis: A Systematic Review and Meta-Analysis. JAMA Netw. Open 2021, 4, e2137257. [Google Scholar] [CrossRef]
- Christie, B. COVID-19: Early Studies Give Hope Omicron Is Milder than Other Variants. BMJ 2021, 375, n3144. [Google Scholar] [CrossRef]
- World Health Organization (WHO). TAG-VE Statement on the Meeting of 3 January on the COVID-19 Situation in China. Available online: https://www.who.int/news/item/04-01-2023-tag-ve-statement-on-the-3rd-january-meeting-on-the-covid-19-situation-in-china (accessed on 9 April 2023).
- Cao, Y.; Jian, F.; Wang, J.; Yu, Y.; Song, W.; Yisimayi, A.; Wang, J.; An, R.; Chen, X.; Zhang, N.; et al. Imprinted SARS-CoV-2 Humoral Immunity Induces Convergent Omicron RBD Evolution. Nature 2023, 614, 521–529. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, M.; Lin, W.; Dong, W.; Xu, J. Evolutionary Analysis of Omicron Variant BF.7 and BA.5.2 Pandemic in China. J. Biosaf. Biosecur. 2023, 5, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Yao, L.; Wei, T.; Tian, F.; Jin, D.-Y.; Chen, L.; Wang, M. Presumed Asymptomatic Carrier Transmission of COVID-19. JAMA 2020, 323, 1406–1407. [Google Scholar] [CrossRef]
- Lauer, S.A.; Grantz, K.H.; Bi, Q.; Jones, F.K.; Zheng, Q.; Meredith, H.R.; Azman, A.S.; Reich, N.G.; Lessler, J. The Incubation Period of Coronavirus Disease 2019 (COVID-19) From Publicly Reported Confirmed Cases: Estimation and Application. Ann. Intern. Med. 2020, 172, 577–582. [Google Scholar] [CrossRef] [PubMed]
- Azkur, A.K.; Akdis, M.; Azkur, D.; Sokolowska, M.; van de Veen, W.; Brüggen, M.-C.; O’Mahony, L.; Gao, Y.; Nadeau, K.; Akdis, C.A. Immune Response to SARS-CoV-2 and Mechanisms of Immunopathological Changes in COVID-19. Allergy 2020, 75, 1564–1581. [Google Scholar] [CrossRef]
- Gandhi, R.T.; Lynch, J.B.; del Rio, C. Mild or Moderate COVID-19. N. Engl. J. Med. 2020, 383, 1757–1766. [Google Scholar] [CrossRef]
- Wang, D.; Hu, B.; Hu, C.; Zhu, F.; Liu, X.; Zhang, J.; Wang, B.; Xiang, H.; Cheng, Z.; Xiong, Y.; et al. Clinical Characteristics of 138 Hospitalized Patients with 2019 Novel Coronavirus-Infected Pneumonia in Wuhan, China. JAMA-J. Am. Med. Assoc. 2020, 323, 1061–1069. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Liao, X.; Qian, S.; Yuan, J.; Wang, F.; Liu, Y.; Wang, Z.; Wang, F.-S.; Liu, L.; Zhang, Z. Community Transmission of Severe Acute Respiratory Syndrome Coronavirus 2, Shenzhen, China, 2020. Emerg. Infect. Dis. 2020, 26, 1320–1323. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.F.-W.; Yuan, S.; Kok, K.-H.; To, K.K.-W.; Chu, H.; Yang, J.; Xing, F.; Liu, J.; Yip, C.C.-Y.; Poon, R.W.-S.; et al. A Familial Cluster of Pneumonia Associated with the 2019 Novel Coronavirus Indicating Person-to-Person Transmission: A Study of a Family Cluster. Lancet 2020, 395, 514–523. [Google Scholar] [CrossRef]
- Burke, R.M.; Midgley, C.M.; Dratch, A.; Fenstersheib, M.; Haupt, T.; Holshue, M.; Ghinai, I.; Jarashow, M.C.; Lo, J.; McPherson, T.D.; et al. Active Monitoring of Persons Exposed to Patients with Confirmed COVID-19-United States, January-February 2020. MMWR Morb. Mortal. Wkly. Rep. 2020, 69, 245–246. [Google Scholar] [CrossRef] [PubMed]
- van Doremalen, N.; Bushmaker, T.; Morris, D.H.; Holbrook, M.G.; Gamble, A.; Williamson, B.N.; Tamin, A.; Harcourt, J.L.; Thornburg, N.J.; Gerber, S.I.; et al. Aerosol and Surface Stability of SARS-CoV-2 as Compared with SARS-CoV-1. N. Engl. J. Med. 2020, 382, 1564–1567. [Google Scholar] [CrossRef] [PubMed]
- Van Damme, W.; Dahake, R.; van de Pas, R.; Vanham, G.; Assefa, Y. COVID-19: Does the Infectious Inoculum Dose-Response Relationship Contribute to Understanding Heterogeneity in Disease Severity and Transmission Dynamics? Med. Hypotheses 2021, 146, 110431. [Google Scholar] [CrossRef] [PubMed]
- Gralton, J.; Tovey, E.R.; McLaws, M.-L.; Rawlinson, W.D. Respiratory Virus RNA Is Detectable in Airborne and Droplet Particles. J. Med. Virol. 2013, 85, 2151–2159. [Google Scholar] [CrossRef] [PubMed]
- Somsen, G.A.; van Rijn, C.; Kooij, S.; Bem, R.A.; Bonn, D. Small Droplet Aerosols in Poorly Ventilated Spaces and SARS-CoV-2 Transmission. Lancet Respir. Med. 2020, 8, 658–659. [Google Scholar] [CrossRef] [PubMed]
- Stadnytskyi, V.; Bax, C.E.; Bax, A.; Anfinrud, P. The Airborne Lifetime of Small Speech Droplets and Their Potential Importance in SARS-CoV-2 Transmission. Proc. Natl. Acad. Sci. USA 2020, 117, 11875. [Google Scholar] [CrossRef] [PubMed]
- UNICEF. Water Security for All; UNICEF: New York, NY, USA, 2021. [Google Scholar]
- Guan, W.; Ni, Z.; Hu, Y.; Liang, W.; Ou, C.; He, J.; Liu, L.; Shan, H.; Lei, C.; Hui, D.S.C.; et al. Clinical Characteristics of Coronavirus Disease 2019 in China. N. Engl. J. Med. 2020, 382, 1708–1720. [Google Scholar] [CrossRef]
- Peng, L.; Liu, J.; Xu, W.; Luo, Q.; Chen, D.; Lei, Z.; Huang, Z.; Li, X.; Deng, K.; Lin, B.; et al. SARS-CoV-2 Can Be Detected in Urine, Blood, Anal Swabs, and Oropharyngeal Swabs Specimens. J. Med. Virol. 2020, 92, 1676–1680. [Google Scholar] [CrossRef]
- Wu, Y.; Guo, C.; Tang, L.; Hong, Z.; Zhou, J.; Dong, X.; Yin, H.; Xiao, Q.; Tang, Y.; Qu, X.; et al. Prolonged Presence of SARS-CoV-2 Viral RNA in Faecal Samples. Lancet Gastroenterol. Hepatol. 2020, 5, 434–435. [Google Scholar] [CrossRef]
- Wang, W.; Xu, Y.; Gao, R.; Lu, R.; Han, K.; Wu, G.; Tan, W. Detection of SARS-CoV-2 in Different Types of Clinical Specimens. JAMA 2020, 323, 1843–1844. [Google Scholar] [CrossRef]
- Xiao, F.; Tang, M.; Zheng, X.; Liu, Y.; Li, X.; Shan, H. Evidence for Gastrointestinal Infection of SARS-CoV-2. Gastroenterology 2020, 158, 1831–1833.e3. [Google Scholar] [CrossRef]
- Zheng, S.; Fan, J.; Yu, F.; Feng, B.; Lou, B.; Zou, Q.; Xie, G.; Lin, S.; Wang, R.; Yang, X.; et al. Viral Load Dynamics and Disease Severity in Patients Infected with SARS-CoV-2 in Zhejiang Province, China, January-March 2020: Retrospective Cohort Study. BMJ 2020, 369, m1443. [Google Scholar] [CrossRef] [PubMed]
- UNICEF; World Health Organization (WHO). Progress on Drinking Water, Sanitation and Hygiene 2000–2017; UNICEF: New York, NY, USA, 2019. [Google Scholar]
- Sun, S.; Han, J. Open Defecation and Squat Toilets, an Overlooked Risk of Fecal Transmission of COVID-19 and Other Pathogens in Developing Communities. Environ. Chem. Lett. 2021, 19, 787–795. [Google Scholar] [CrossRef]
- World Health Organization. Breastfeeding and COVID-19: WHO Scientific Brief; World Health Organization: Geneva, Switzerland, 2020; pp. 1–3. [Google Scholar]
- Bhattacharjee, N.V.; Schaeffer, L.E.; Marczak, L.B.; Ross, J.M.; Swartz, S.J.; Albright, J.; Gardner, W.M.; Shields, C.; Sligar, A.; Schipp, M.F.; et al. Mapping Exclusive Breastfeeding in Africa between 2000 and 2017. Nat. Med. 2019, 25, 1205–1212. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharjee, N.V.; Schaeffer, L.E.; Hay, S.I.; Lu, D.; Schipp, M.F.; Lazzar-Atwood, A.; Donkers, K.M.; Abady, G.G.; Abd-Allah, F.; Abdelalim, A.; et al. Mapping Inequalities in Exclusive Breastfeeding in Low- and Middle-Income Countries, 2000–2018. Nat. Hum. Behav. 2021, 5, 1027–1045. [Google Scholar] [CrossRef]
- World Health Organization (WHO); UNICEF. Global Nutrition Targets 2025: Breastfeeding Policy Brief; World Health Organization: Geneva, Switzerland, 2014. [Google Scholar]
- Dong, L.; Tian, J.; He, S.; Zhu, C.; Wang, J.; Liu, C.; Yang, J. Possible Vertical Transmission of SARS-CoV-2 From an Infected Mother to Her Newborn. JAMA 2020, 323, 1846–1848. [Google Scholar] [CrossRef]
- Alabi, Q.K.; Oyedeji, A.S.; Kayode, O.O.; Kajewole-Alabi, D.I. Impact of COVID-19 Pandemic on Mother and Child Health in Sub-Saharan Africa—A Review. Pediatr. Res. 2023, 94, 1278–1283. [Google Scholar] [CrossRef] [PubMed]
- Desai, A.D.; Lavelle, M.; Boursiquot, B.C.; Wan, E.Y. Long-Term Complications of COVID-19. Am. J. Physiol.-Cell Physiol. 2022, 322, C1–C11. [Google Scholar] [CrossRef]
- Zeng, N.; Zhao, Y.-M.; Yan, W.; Li, C.; Lu, Q.-D.; Liu, L.; Ni, S.-Y.; Mei, H.; Yuan, K.; Shi, L.; et al. A Systematic Review and Meta-Analysis of Long Term Physical and Mental Sequelae of COVID-19 Pandemic: Call for Research Priority and Action. Mol. Psychiatry 2023, 28, 423–433. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Haupert, S.R.; Zimmermann, L.; Shi, X.; Fritsche, L.G.; Mukherjee, B. Global Prevalence of Post-Coronavirus Disease 2019 (COVID-19) Condition or Long COVID: A Meta-Analysis and Systematic Review. J. Infect. Dis. 2022, 226, 1593–1607. [Google Scholar] [CrossRef]
- Han, Q.; Zheng, B.; Daines, L.; Sheikh, A. Long-Term Sequelae of COVID-19: A Systematic Review and Meta-Analysis of One-Year Follow-Up Studies on Post-COVID Symptoms. Pathogens 2022, 11, 269. [Google Scholar] [CrossRef] [PubMed]
- Balbi, M.; Conti, C.; Imeri, G.; Caroli, A.; Surace, A.; Corsi, A.; Mercanzin, E.; Arrigoni, A.; Villa, G.; Di Marco, F.; et al. Post-Discharge Chest CT Findings and Pulmonary Function Tests in Severe COVID-19 Patients. Eur. J. Radiol. 2021, 138, 109676. [Google Scholar] [CrossRef] [PubMed]
- Nyasulu, P.S.; Tamuzi, J.L.; Erasmus, R.T. Burden, Causation, and Particularities of Long-COVID in African Populations: A Rapid Systematic Review. medRxiv 2023. [Google Scholar] [CrossRef] [PubMed]
- Jassat, W.; Mudara, C.; Vika, C.; Welch, R.; Arendse, T.; Dryden, M.; Blumberg, L.; Mayet, N.; Tempia, S.; Parker, A.; et al. A Cohort Study of Post-COVID-19 Condition across the Beta, Delta, and Omicron Waves in South Africa: 6-Month Follow-up of Hospitalized and Nonhospitalized Participants. Int. J. Infect. Dis. 2023, 128, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Lawal, Y. Africa’s Low COVID-19 Mortality Rate: A Paradox? Int. J. Infect. Dis. 2021, 102, 118–122. [Google Scholar] [CrossRef] [PubMed]
- Dalal, S.; Beunza, J.J.; Volmink, J.; Adebamowo, C.; Bajunirwe, F.; Njelekela, M.; Mozaffarian, D.; Fawzi, W.; Willett, W.; Adami, H.-O.; et al. Non-Communicable Diseases in Sub-Saharan Africa: What We Know Now. Int. J. Epidemiol. 2011, 40, 885–901. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Deaths from Noncommunicable Diseases on the Rise in Africa. Available online: https://www.afro.who.int/news/deaths-noncommunicable-diseases-rise-africa (accessed on 9 April 2023).
- Ajaero, C.K.; De Wet-Billings, N.; Atama, C.; Agwu, P.; Eze, E.J. The Prevalence and Contextual Correlates of Non-Communicable Diseases among Inter-Provincial Migrants and Non-Migrants in South Africa. BMC Public Health 2021, 21, 999. [Google Scholar] [CrossRef]
- Vos, T.; Lim, S.S.; Abbafati, C.; Abbas, K.M.; Abbasi, M.; Abbasifard, M.; Abbasi-Kangevari, M.; Abbastabar, H.; Abd-Allah, F.; Abdelalim, A.; et al. Global Burden of 369 Diseases and Injuries in 204 Countries and Territories, 1990-2019: A Systematic Analysis for the Global Burden of Disease Study 2019. Lancet 2020, 396, 1204–1222. [Google Scholar] [CrossRef]
- Cole, J.B.; Florez, J.C. Genetics of Diabetes Mellitus and Diabetes Complications. Nat. Rev. Nephrol. 2020, 16, 377–390. [Google Scholar] [CrossRef]
- World Health Organization (WHO). African Region Tops World in Undiagnosed Diabetes: WHO Analysis. Available online: https://www.afro.who.int/news/african-region-tops-world-undiagnosed-diabetes-who-analysis (accessed on 28 March 2023).
- Anjorin, A.A.; Abioye, A.I.; Asowata, O.E.; Soipe, A.; Kazeem, M.I.; Adesanya, I.O.; Raji, M.A.; Adesanya, M.; Oke, F.A.; Lawal, F.J.; et al. Comorbidities and the COVID-19 Pandemic Dynamics in Africa. Trop. Med. Int. Health 2021, 26, 2–13. [Google Scholar] [CrossRef]
- Schoen, K.; Horvat, N.; Guerreiro, N.F.C.; de Castro, I.; de Giassi, K.S. Spectrum of Clinical and Radiographic Findings in Patients with Diagnosis of H1N1 and Correlation with Clinical Severity. BMC Infect. Dis. 2019, 19, 964. [Google Scholar] [CrossRef]
- Banik, G.R.; Alqahtani, A.S.; Booy, R.; Rashid, H. Risk Factors for Severity and Mortality in Patients with MERS-CoV: Analysis of Publicly Available Data from Saudi Arabia. Virol. Sin. 2016, 31, 81–84. [Google Scholar] [CrossRef]
- Yang, J.K.; Feng, Y.; Yuan, M.Y.; Yuan, S.Y.; Fu, H.J.; Wu, B.Y.; Sun, G.Z.; Yang, G.R.; Zhang, X.L.; Wang, L.; et al. Plasma Glucose Levels and Diabetes Are Independent Predictors for Mortality and Morbidity in Patients with SARS. Diabet. Med. 2006, 23, 623–628. [Google Scholar] [CrossRef]
- Hussain, A.; Bhowmik, B.; do Vale Moreira, N.C. COVID-19 and Diabetes: Knowledge in Progress. Diabetes Res. Clin. Pract. 2020, 162, 108142. [Google Scholar] [CrossRef] [PubMed]
- Burki, T. COVID-19 and Diabetes in Africa: A Lethal Combination. Lancet Diabetes Endocrinol. 2022, 10, 23. [Google Scholar] [CrossRef] [PubMed]
- Kerolos, M.M.; Ruge, M.; Gill, A.; Planek, M.I.; Volgman, A.S.; Du-Fay-De-Lavallaz, J.M.; Gomez, J.M.D.; Suboc, T.M.; Williams, K.A.; Abusin, S. Clinical Outcomes of COVID-19 Infection in Patients with Pre-Existing Cardiovascular Disease. Am. Heart J. Plus Cardiol. Res. Pract. 2022, 20, 100189. [Google Scholar] [CrossRef] [PubMed]
- Nkomo, V.T. Epidemiology and Prevention of Valvular Heart Diseases and Infective Endocarditis in Africa. Heart 2007, 93, 1510. [Google Scholar] [CrossRef] [PubMed]
- Anabwani, G.M.; Amoa, A.B.; Muita, A.K. Epidemiology of Rheumatic Heart Disease among Primary School Children in Western Kenya. Int. J. Cardiol. 1989, 23, 249–252. [Google Scholar] [CrossRef]
- McLaren, M.J.; Hawkins, D.M.; Koornhof, H.J.; Bloom, K.R.; Bramwell-Jones, D.M.; Cohen, E.; Gale, G.E.; Kanarek, K.; Lachman, A.S.; Lakier, J.B.; et al. Epidemiology of Rheumatic Heart Disease in Black Shcoolchildren of Soweto, Johannesburg. Br. Med. J. 1975, 3, 474. [Google Scholar] [CrossRef]
- Marijon, E.; Ou, P.; Celermajer, D.S.; Ferreira, B.; Mocumbi, A.O.; Jani, D.; Paquet, C.; Jacob, S.; Sidi, D.; Jouven, X. Prevalence of Rheumatic Heart Disease Detected by Echocardiographic Screening. N. Engl. J. Med. 2007, 357, 470–476. [Google Scholar] [CrossRef]
- Ibrahim-Khalil, S.; Elhag, M.; Ali, E.; Mahgoub, F.; Hakiem, S.; Omer, N.; Shafie, S.; Mahgoub, E. An Epidemiological Survey of Rheumatic Fever and Rheumatic Heart Disease in Sahafa Town, Sudan. J. Epidemiol. Community Health 1992, 46, 477. [Google Scholar] [CrossRef] [PubMed]
- Oli, K.; Tekle-Haimanot, R.; Forsgren, L.; Ekstedt, J. Rheumatic Heart Disease Prevalence among Schoolchildren of an Ethiopian Rural Town. Cardiology 1992, 80, 152–155. [Google Scholar] [CrossRef] [PubMed]
- Margolin, E.; Burgers, W.A.; Sturrock, E.D.; Mendelson, M.; Chapman, R.; Douglass, N.; Williamson, A.-L.; Rybicki, E.P. Prospects for SARS-CoV-2 Diagnostics, Therapeutics and Vaccines in Africa. Nat. Rev. Microbiol. 2020, 18, 690–704. [Google Scholar] [CrossRef]
- Western Cape Department of Health in Collaboration with the National Institute for Communicable; Diseases, S.A. Risk Factors for Coronavirus Disease 2019 (COVID-19) Death in a Population Cohort Study from the Western Cape Province, South Africa. Clin. Infect. Dis. 2021, 73, e2005. [Google Scholar] [CrossRef]
- Motta, I.; Centis, R.; D’Ambrosio, L.; García-García, J.-M.; Goletti, D.; Gualano, G.; Lipani, F.; Palmieri, F.; Sánchez-Montalvá, A.; Pontali, E.; et al. Tuberculosis, COVID-19 and Migrants: Preliminary Analysis of Deaths Occurring in 69 Patients from Two Cohorts. Pulmonology 2020, 26, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Nägele, F.; Graber, M.; Hirsch, J.; Pölzl, L.; Sahanic, S.; Fiegl, M.; Hau, D.; Engler, C.; Lechner, S.; Stalder, A.K.; et al. Correlation between Structural Heart Disease and Cardiac SARS-CoV-2 Manifestations. Commun. Med. 2022, 2, 142. [Google Scholar] [CrossRef] [PubMed]
- Tomasoni, D.; Italia, L.; Adamo, M.; Inciardi, R.M.; Lombardi, C.M.; Solomon, S.D.; Metra, M. COVID-19 and Heart Failure: From Infection to Inflammation and Angiotensin II Stimulation. Searching for Evidence from a New Disease. Eur. J. Heart Fail. 2020, 22, 957–966. [Google Scholar] [CrossRef] [PubMed]
- Omoush, S.A.; Alzyoud, J.A.M. The Prevalence and Impact of Coinfection and Superinfection on the Severity and Outcome of COVID-19 Infection: An Updated Literature Review. Pathogens 2022, 11, 445. [Google Scholar] [CrossRef]
- Blanco, J.-R.; Cobos-Ceballos, M.-J.; Navarro, F.; Sanjoaquin, I.; Arnaiz de las Revillas, F.; Bernal, E.; Buzon-Martin, L.; Viribay, M.; Romero, L.; Espejo-Perez, S.; et al. Pulmonary Long-Term Consequences of COVID-19 Infections after Hospital Discharge. Clin. Microbiol. Infect. 2021, 27, 892–896. [Google Scholar] [CrossRef]
- Raisi-Estabragh, Z.; Cooper, J.; Salih, A.; Raman, B.; Lee, A.M.; Neubauer, S.; Harvey, N.C.; Petersen, S.E. Cardiovascular Disease and Mortality Sequelae of COVID-19 in the UK Biobank. Heart 2023, 109, 119. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Obesity Rising in Africa, WHO Analysis Finds. Available online: https://www.afro.who.int/news/obesity-rising-africa-who-analysis-finds (accessed on 12 March 2023).
- Arentz, M.; Yim, E.; Klaff, L.; Lokhandwala, S.; Riedo, F.X.; Chong, M.; Lee, M. Characteristics and Outcomes of 21 Critically Ill Patients With COVID-19 in Washington State. JAMA 2020, 323, 1612–1614. [Google Scholar] [CrossRef]
- Bhatraju, P.K.; Ghassemieh, B.J.; Nichols, M.; Kim, R.; Jerome, K.R.; Nalla, A.K.; Greninger, A.L.; Pipavath, S.; Wurfel, M.M.; Evans, L.; et al. COVID-19 in Critically Ill Patients in the Seattle Region—Case Series. N. Engl. J. Med. 2020, 382, 2012–2022. [Google Scholar] [CrossRef] [PubMed]
- Richardson, S.; Hirsch, J.S.; Narasimhan, M.; Crawford, J.M.; McGinn, T.; Davidson, K.W.; Barnaby, D.P.; Becker, L.B.; Chelico, J.D.; Cohen, S.L.; et al. Presenting Characteristics, Comorbidities, and Outcomes Among 5700 Patients Hospitalized With COVID-19 in the New York City Area. JAMA 2020, 323, 2052–2059. [Google Scholar] [CrossRef] [PubMed]
- Nakeshbandi, M.; Maini, R.; Daniel, P.; Rosengarten, S.; Parmar, P.; Wilson, C.; Kim, J.M.; Oommen, A.; Mecklenburg, M.; Salvani, J.; et al. The Impact of Obesity on COVID-19 Complications: A Retrospective Cohort Study. Int. J. Obes. 2020, 44, 1832–1837. [Google Scholar] [CrossRef] [PubMed]
- Nagy, É.; Cseh, V.; Barcs, I.; Ludwig, E. The Impact of Comorbidities and Obesity on the Severity and Outcome of COVID-19 in Hospitalized Patients—A Retrospective Study in a Hungarian Hospital. Int. J. Environ. Res. Public Health 2023, 20, 1372. [Google Scholar] [CrossRef] [PubMed]
- Katona, P.; Katona-Apte, J. The Interaction between Nutrition and Infection. Clin. Infect. Dis. 2008, 46, 1582–1588. [Google Scholar] [CrossRef]
- Mertens, E.; Peñalvo, J.L. The Burden of Malnutrition and Fatal COVID-19: A Global Burden of Disease Analysis. Front. Nutr. 2021, 7, 351. [Google Scholar] [CrossRef] [PubMed]
- Menshawey, R.; Menshawey, E.; Alserr, A.H.K.; Abdelmassih, A.F. Low Iron Mitigates Viral Survival: Insights from Evolution, Genetics, and Pandemics—A Review of Current Hypothesis. Egypt. J. Med. Hum. Genet. 2020, 21, 75. [Google Scholar] [CrossRef]
- Williams, A.M.; Ladva, C.N.; Leon, J.S.; Lopman, B.A.; Tangpricha, V.; Whitehead, R.D.; Armitage, A.E.; Wray, K.; Morovat, A.; Pasricha, S.R.; et al. Changes in Micronutrient and Inflammation Serum Biomarker Concentrations after a Norovirus Human Challenge. Am. J. Clin. Nutr. 2019, 110, 1456–1464. [Google Scholar] [CrossRef]
- Cavezzi, A.; Troiani, E.; Corrao, S. COVID-19: Hemoglobin, Iron, and Hypoxia beyond Inflammation. A Narrative Review. Clin. Pract. 2020, 10, 1271. [Google Scholar] [CrossRef]
- Chen, N.; Zhou, M.; Dong, X.; Qu, J.; Gong, F.; Han, Y.; Qiu, Y.; Wang, J.; Liu, Y.; Wei, Y.; et al. Epidemiological and Clinical Characteristics of 99 Cases of 2019 Novel Coronavirus Pneumonia in Wuhan, China: A Descriptive Study. Lancet 2020, 395, 507–513. [Google Scholar] [CrossRef]
- Chen, X.; Zhao, B.; Qu, Y.; Chen, Y.; Xiong, J.; Feng, Y.; Men, D.; Huang, Q.; Liu, Y.; Yang, B.; et al. Detectable Serum Severe Acute Respiratory Syndrome Coronavirus 2 Viral Load (RNAemia) Is Closely Correlated with Drastically Elevated Interleukin 6 Level in Critically Ill Patients with Coronavirus Disease 2019. Clin. Infect. Dis. 2020, 71, 1937–1942. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Li, M.; Zhou, Z.; Guan, X.; Xiang, Y. Can We Use Interleukin-6 (IL-6) Blockade for Coronavirus Disease 2019 (COVID-19)-Induced Cytokine Release Syndrome (CRS)? J. Autoimmun. 2020, 111, 102452. [Google Scholar] [CrossRef] [PubMed]
- Martins, A.C.; Almeida, J.I.; Lima, I.S.; Kapitão, A.S.; Gozzelino, R. Iron Metabolism and the Inflammatory Response. IUBMB Life 2017, 69, 442–450. [Google Scholar] [CrossRef] [PubMed]
- Alipour, R.; Hashemi, S.H.; Mikaeili, F. Serum Iron Level in Patients with COVID-19: A Case Report Study. Int. J. Res. Med. Sci. 2020, 8, 2658. [Google Scholar] [CrossRef]
- Nai, A.; Lorè, N.I.; Pagani, A.; De Lorenzo, R.; Di Modica, S.; Saliu, F.; Cirillo, D.M.; Rovere-Querini, P.; Manfredi, A.A.; Silvestri, L. Hepcidin Levels Predict COVID-19 Severity and Mortality in a Cohort of Hospitalized Italian Patients. Am. J. Hematol. 2021, 96, E32–E35. [Google Scholar] [CrossRef]
- Ganz, T. Anemia of Inflammation. N. Engl. J. Med. 2019, 381, 1148–1157. [Google Scholar] [CrossRef]
- Yağcı, S.; Serin, E.; Acicbe, Ö.; Zeren, M.İ.; Odabaşı, M.S. The Relationship between Serum Erythropoietin, Hepcidin, and Haptoglobin Levels with Disease Severity and Other Biochemical Values in Patients with COVID-19. Int. J. Lab. Hematol. 2021, 43, 142–151. [Google Scholar] [CrossRef]
- Litton, E.; Lim, J. Iron Metabolism: An Emerging Therapeutic Target in Critical Illness. Crit. Care 2019, 2019, 573–584. [Google Scholar]
- Shah, A.; Frost, J.N.; Aaron, L.; Donovan, K.; Drakesmith, H.; McKechnie, S.R.; Stanworth, S.J. Systemic Hypoferremia and Severity of Hypoxemic Respiratory Failure in COVID-19. Crit. Care 2020, 24, 320. [Google Scholar] [CrossRef]
- Wang, K.; Chen, W.; Zhou, Y.-S.; Lian, J.-Q.; Zhang, Z.; Du, P.; Gong, L.; Zhang, Y.; Cui, H.-Y.; Geng, J.-J.; et al. SARS-CoV-2 Invades Host Cells via a Novel Route: CD147-Spike Protein. bioRxiv 2020. [Google Scholar] [CrossRef]
- Radzikowska, U.; Ding, M.; Tan, G.; Zhakparov, D.; Peng, Y.; Wawrzyniak, P.; Wang, M.; Li, S.; Morita, H.; Altunbulakli, C.; et al. Distribution of ACE2, CD147, CD26, and Other SARS-CoV-2 Associated Molecules in Tissues and Immune Cells in Health and in Asthma, COPD, Obesity, Hypertension, and COVID-19 Risk Factors. Allergy Eur. J. Allergy Clin. Immunol. 2020, 75, 2829–2845. [Google Scholar] [CrossRef]
- Latini, A.; Agolini, E.; Novelli, A.; Borgiani, P.; Giannini, R.; Gravina, P.; Smarrazzo, A.; Dauri, M.; Andreoni, M.; Rogliani, P.; et al. COVID-19 and Genetic Variants of Protein Involved in the SARS-CoV-2 Entry into the Host Cells. Genes 2020, 11, 1010. [Google Scholar] [CrossRef] [PubMed]
- Shilts, J.; Crozier, T.W.M.; Greenwood, E.J.D.; Lehner, P.J.; Wright, G.J. No Evidence for Basigin/CD147 as a Direct SARS-CoV-2 Spike Binding Receptor. Sci. Rep. 2021, 11, 413. [Google Scholar] [CrossRef] [PubMed]
- Vankadari, N.; Wilce, J.A. Emerging WuHan (COVID-19) Coronavirus: Glycan Shield and Structure Prediction of Spike Glycoprotein and Its Interaction with Human CD26. Emerg. Microbes Infect. 2020, 9, 601–604. [Google Scholar] [CrossRef]
- O’Leary, H.; Ou, X.; Broxmeyer, H.E. The Role of Dipeptidyl Peptidase 4 in Hematopoiesis and Transplantation. Curr. Opin. Hematol. 2013, 20, 314. [Google Scholar] [CrossRef]
- Bu, X.; Yao, Y.; Li, X. Immune Checkpoint Blockade in Breast Cancer Therapy. In Advances in Experimental Medicine and Biology; Springer: Berlin/Heidelberg, Germany, 2017. [Google Scholar]
- Soumana, A.; Samaila, A.; Moustapha, L.M.; Kamaye, M.; Daouda, B.; Salifou, I.A.; Lagare, A.; Adehossi, E.O.; Ibrahim, M.L. A Fatal Case of COVID-19 in an Infant with Severe Acute Malnutrition Admitted to a Paediatric Ward in Niger. Case Rep. Pediatr. 2020, 2020, 8847415. [Google Scholar] [CrossRef]
- Savino, W.; Dardenne, M.; Velloso, L.A.; Dayse Silva-Barbosa, S. The Thymus Is a Common Target in Malnutrition and Infection. Br. J. Nutr. 2007, 98, S11–S16. [Google Scholar] [CrossRef]
- Vong, T.; Yanek, L.R.; Wang, L.; Yu, H.; Fan, C.; Zhou, E.; Oh, S.J.; Szvarca, D.; Kim, A.; Potter, J.J.; et al. Malnutrition Increases Hospital Length of Stay and Mortality among Adult Inpatients with COVID-19. Nutrients 2022, 14, 1310. [Google Scholar] [CrossRef]
- Kuvibidila, S.; Dardenne, M.; Savino, W.; Lepault, F. Influence of Iron-Deficiency Anemia on Selected Thymus Functions in Mice: Thymulin Biological Activity, T-Cell Subsets, and Thymocyte Proliferation. Am. J. Clin. Nutr. 1990, 51, 228–232. [Google Scholar] [CrossRef]
- Gwamaka, M.; Kurtis, J.D.; Sorensen, B.E.; Holte, S.; Morrison, R.; Mutabingwa, T.K.; Fried, M.; Duffy, P.E. Iron Deficiency Protects against Severe Plasmodium Falciparum Malaria and Death in Young Children. Clin. Infect. Dis. 2012, 54, 1137–1144. [Google Scholar] [CrossRef] [PubMed]
- Frost, J.N.; Tan, T.K.; Abbas, M.; Wideman, S.K.; Bonadonna, M.; Stoffel, N.U.; Wray, K.; Kronsteiner, B.; Smits, G.; Campagna, D.R.; et al. Hepcidin-Mediated Hypoferremia Disrupts Immune Responses to Vaccination and Infection. Med 2021, 2, 164–179.e12. [Google Scholar] [CrossRef]
- Stoffel, N.U.; Uyoga, M.A.; Mutuku, F.M.; Frost, J.N.; Mwasi, E.; Paganini, D.; van der Klis, F.R.M.; Malhotra, I.J.; LaBeaud, A.D.; Ricci, C.; et al. Iron Deficiency Anemia at Time of Vaccination Predicts Decreased Vaccine Response and Iron Supplementation at Time of Vaccination Increases Humoral Vaccine Response: A Birth Cohort Study and a Randomized Trial Follow-Up Study in Kenyan Infants. Front. Immunol. 2020, 11, 1313. [Google Scholar] [CrossRef] [PubMed]
- Cugno, M.; Meroni, P.L.; Consonni, D.; Griffini, S.; Grovetti, E.; Novembrino, C.; Torri, A.; Griffante, G.; Gariglio, M.; Varani, L.; et al. Effects of Antibody Responses to Pre-Existing Coronaviruses on Disease Severity and Complement Activation in COVID-19 Patients. Microorganisms 2022, 10, 1191. [Google Scholar] [CrossRef]
- Sagar, M.; Reifler, K.; Rossi, M.; Miller, N.S.; Sinha, P.; White, L.F.; Mizgerd, J.P. Recent Endemic Coronavirus Infection Is Associated with Less-Severe COVID-19. J. Clin. Investig. 2021, 131, e143380. [Google Scholar] [CrossRef]
- Namuniina, A.; Muyanja, E.S.; Biribawa, V.M.; Okech, B.A.; Price, M.A.; Hills, N.; Nanteza, A.; Ssentalo Bagaya, B.; Weiskopf, D.; Riou, C.; et al. High Proportion of Ugandans with Pre-Pandemic SARS-CoV-2 Cross-Reactive CD4+ and CD8+ T-Cell 2 Responses. medRxiv 2023. [Google Scholar] [CrossRef]
- Pedersen, J.; Koumakpayi, I.H.; Babuadze, G.; Baz, M.; Ndiaye, O.; Faye, O.; Diagne, C.T.; Dia, N.; Naghibosadat, M.; McGeer, A.; et al. Cross-Reactive Immunity against SARS-CoV-2 N Protein in Central and West Africa Precedes the COVID-19 Pandemic. Sci. Rep. 2022, 12, 12962. [Google Scholar] [CrossRef]
- Habibzadeh, F. Malaria and the Incidence of COVID-19 in Africa: An Ecological Study. BMC Infect. Dis. 2023, 23, 66. [Google Scholar] [CrossRef]
- Divangahi, M.; Aaby, P.; Khader, S.A.; Barreiro, L.B.; Bekkering, S.; Chavakis, T.; van Crevel, R.; Curtis, N.; DiNardo, A.R.; Dominguez-Andres, J.; et al. Trained Immunity, Tolerance, Priming and Differentiation: Distinct Immunological Processes. Nat. Immunol. 2021, 22, 2–6. [Google Scholar] [CrossRef] [PubMed]
- Schrum, J.E.; Crabtree, J.N.; Dobbs, K.R.; Kiritsy, M.C.; Reed, G.W.; Gazzinelli, R.T.; Netea, M.G.; Kazura, J.W.; Dent, A.E.; Fitzgerald, K.A.; et al. Plasmodium Falciparum Induces Trained Innate Immunity. J. Immunol. 2018, 200, 1243–1248. [Google Scholar] [CrossRef] [PubMed]
- Steinberg, M.H. Management of Sickle Cell Disease. N. Engl. J. Med. 1999, 340, 1021–1030. [Google Scholar] [CrossRef]
- Crowell, T.A.; Daud, I.I.; Maswai, J.; Owuoth, J.; Sing’oei, V.; Imbach, M.; Dear, N.; Sawe, F.; Eller, L.A.; Polyak, C.S.; et al. Severe Acute Respiratory Syndrome Coronavirus-2 Antibody Prevalence in People with and without HIV in Rural Western Kenya, January to March 2020. AIDS 2021, 35, 2401–2404. [Google Scholar] [CrossRef]
- Vergori, A.; Boschini, A.; Notari, S.; Lorenzini, P.; Castilletti, C.; Colavita, F.; Matusali, G.; Tartaglia, E.; Gagliardini, R.; Boschi, A.; et al. SARS-CoV-2 Specific Immune Response and Inflammatory Profile in Advanced HIV-Infected Persons during a COVID-19 Outbreak. Viruses 2022, 14, 1575. [Google Scholar] [CrossRef]
- du Bruyn, E.; Stek, C.; Daroowala, R.; Said-Hartley, Q.; Hsiao, M.; Schafer, G.; Goliath, R.T.; Abrahams, F.; Jackson, A.; Wasserman, S.; et al. Effects of Tuberculosis and/or HIV-1 Infection on COVID-19 Presentation and Immune Response in Africa. Nat. Commun. 2023, 14, 188. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wang, Y.; Fleming, J.; Yu, Y.; Gu, Y.; Liu, C.; Fan, L.; Wang, X.; Cheng, M.; Bi, L.; et al. Active or Latent Tuberculosis Increases Susceptibility to COVID-19 and Disease Severity. medRxiv 2020. [Google Scholar] [CrossRef]
- Sy, K.T.L.; Haw, N.J.L.; Uy, J. Previous and Active Tuberculosis Increases Risk of Death and Prolongs Recovery in Patients with COVID-19. medRxiv 2020. [Google Scholar] [CrossRef] [PubMed]
- Davies, M.-A. HIV and Risk of COVID-19 Death: A Population Cohort Study from the Western Cape Province, South Africa. medRxiv 2020. [Google Scholar] [CrossRef]
- Madan, M.; Baldwa, B.; Raja, A.; Tyagi, R.; Dwivedi, T.; Mohan, A.; Mittal, S.; Madan, K.; Hadda, V.; Tiwari, P.; et al. Impact of Latent Tuberculosis on Severity and Outcomes in Admitted COVID-19 Patients. Cureus 2021, 13, e19882. [Google Scholar] [CrossRef]
- Costela-Ruiz, V.J.; Illescas-Montes, R.; Puerta-Puerta, J.M.; Ruiz, C.; Melguizo-Rodríguez, L. SARS-CoV-2 Infection: The Role of Cytokines in COVID-19 Disease. Cytokine Growth Factor Rev. 2020, 54, 62–75. [Google Scholar] [CrossRef]
- Rajamanickam, A.; Kumar, N.P.; Padmapriyadarsini, C.; Nancy, A.; Selvaraj, N.; Karunanithi, K.; Munisankar, S.; BM, S.; Renji, R.M.; Ambu, T.C.; et al. Latent Tuberculosis Co-Infection Is Associated with Heightened Levels of Humoral, Cytokine and Acute Phase Responses in Seropositive SARS-CoV-2 Infection. J. Infect. 2021, 83, 339–346. [Google Scholar] [CrossRef]
- Riou, C.; du Bruyn, E.; Stek, C.; Daroowala, R.; Goliath, R.T.; Abrahams, F.; Said-Hartley, Q.; Allwood, B.W.; Hsiao, N.-Y.; Wilkinson, K.A.; et al. Relationship of SARS-CoV-2–Specific CD4 Response to COVID-19 Severity and Impact of HIV-1 and Tuberculosis Coinfection. J. Clin. Investig. 2021, 131, e149125. [Google Scholar] [CrossRef] [PubMed]
- Day, C.L.; Abrahams, D.A.; Lerumo, L.; Janse van Rensburg, E.; Stone, L.; O’rie, T.; Pienaar, B.; de Kock, M.; Kaplan, G.; Mahomed, H.; et al. Functional Capacity of Mycobacterium Tuberculosis -Specific T Cell Responses in Humans Is Associated with Mycobacterial Load. J. Immunol. 2011, 187, 2222–2232. [Google Scholar] [CrossRef]
- Zhou, F.; Yu, T.; Du, R.; Fan, G.; Liu, Y.; Liu, Z.; Xiang, J.; Wang, Y.; Song, B.; Gu, X.; et al. Clinical Course and Risk Factors for Mortality of Adult Inpatients with COVID-19 in Wuhan, China: A Retrospective Cohort Study. Lancet 2020, 395, 1054–1062. [Google Scholar] [CrossRef] [PubMed]
- Lippi, G.; Plebani, M. Laboratory Abnormalities in Patients with COVID-2019 Infection. Clin. Chem. Lab. Med. (CCLM) 2020, 58, 1131–1134. [Google Scholar] [CrossRef] [PubMed]
- Lippi, G.; Plebani, M. Procalcitonin in Patients with Severe Coronavirus Disease 2019 (COVID-19): A Meta-Analysis. Clin. Chim. Acta 2020, 505, 190–191. [Google Scholar] [CrossRef] [PubMed]
- Herold, T.; Jurinovic, V.; Arnreich, C.; Hellmuth, J.C.; von Bergwelt-Baildon, M.; Klein, M.; Weinberger, T. Level of IL-6 Predicts Respiratory Failure in Hospitalized Symptomatic COVID-19 Patients. medRxiv 2020. [Google Scholar] [CrossRef]
- Kuodi, P.; Gorelik, Y.; Zayyad, H.; Wertheim, O.; Wiegler, K.B.; Abu Jabal, K.; Dror, A.A.; Nazzal, S.; Glikman, D.; Edelstein, M. Association between BNT162b2 Vaccination and Reported Incidence of Post-COVID-19 Symptoms: Cross-Sectional Study 2020-21, Israel. NPJ Vaccines 2022, 7, 101. [Google Scholar] [CrossRef] [PubMed]
- Venkatesan, P. Do Vaccines Protect from Long COVID? Lancet Respir. Med. 2022, 10, e30. [Google Scholar] [CrossRef]
- Ayoubkhani, D.; Bermingham, C.; Pouwels, K.B.; Glickman, M.; Nafilyan, V.; Zaccardi, F.; Khunti, K.; Alwan, N.A.; Walker, A.S. Trajectory of Long COVID Symptoms after COVID-19 Vaccination: Community Based Cohort Study. BMJ 2022, 377, e069676. [Google Scholar] [CrossRef]
- Tran, V.-T.; Porcher, R.; Pane, I.; Ravaud, P. Course of Post COVID-19 Disease Symptoms over Time in the ComPaRe Long COVID Prospective e-Cohort. Nat. Commun. 2022, 13, 1812. [Google Scholar] [CrossRef]
- Pizarro-Pennarolli, C.; Sánchez-Rojas, C.; Torres-Castro, R.; Vera-Uribe, R.; Sanchez-Ramirez, D.C.; Vasconcello-Castillo, L.; Solís-Navarro, L.; Rivera-Lillo, G. Assessment of Activities of Daily Living in Patients Post COVID-19: A Systematic Review. PeerJ 2021, 9, e11026. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). COVID-19 Vaccine Tracker and Landscape. Available online: https://www.who.int/publications/m/item/draft-landscape-of-covid-19-candidate-vaccines (accessed on 1 February 2023).
- Sharma, K.; Koirala, A.; Nicolopoulos, K.; Chiu, C.; Wood, N.; Britton, P.N. Vaccines for COVID-19: Where Do We Stand in 2021? Paediatr. Respir. Rev. 2021, 39, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Kudlay, D.; Svistunov, A.; Satyshev, O. COVID-19 Vaccines: An Updated Overview of Different Platforms. Bioengineering 2022, 9, 714. [Google Scholar] [CrossRef]
- The Nobel Assembly at Karolinska Institutet. Nobel Prize Outreach AB 2023. Available online: https://www.nobelprize.org/prizes/medicine/2023/press-release/ (accessed on 24 January 2024).
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Pérez Marc, G.; Moreira, E.D.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 MRNA COVID-19 Vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef] [PubMed]
- Baden, L.R.; El Sahly, H.M.; Essink, B.; Kotloff, K.; Frey, S.; Novak, R.; Diemert, D.; Spector, S.A.; Rouphael, N.; Creech, C.B.; et al. Efficacy and Safety of the MRNA-1273 SARS-CoV-2 Vaccine. N. Engl. J. Med. 2020, 384, 403–416. [Google Scholar] [CrossRef] [PubMed]
- Sharma, O.; Sultan, A.A.; Ding, H.; Triggle, C.R. A Review of the Progress and Challenges of Developing a Vaccine for COVID-19. Front. Immunol. 2020, 11, 585354. [Google Scholar] [CrossRef] [PubMed]
- Walsh, E.E.; Frenck, R.W.; Falsey, A.R.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Neuzil, K.; Mulligan, M.J.; Bailey, R.; et al. Safety and Immunogenicity of Two RNA-Based COVID-19 Vaccine Candidates. N. Engl. J. Med. 2020, 383, 2439–2450. [Google Scholar] [CrossRef]
- NHS England. COVID-19 MRNA Vaccine (BNT162) Mechanism of Action for BNT162B2. 2021. Available online: https://www.england.nhs.uk/south/wp-content/uploads/sites/6/2021/07/phe-ref-ra-21.034-part-2-covid-19-vaccine-modernar-and-pfizer-biontech-myocarditis-and-pericarditis.pdf (accessed on 24 January 2024).
- Tavilani, A.; Abbasi, E.; Kian Ara, F.; Darini, A.; Asefy, Z. COVID-19 Vaccines: Current Evidence and Considerations. Metabol. Open 2021, 12, 100124. [Google Scholar] [CrossRef]
- Tatsis, N.; Ertl, H.C.J. Adenoviruses as Vaccine Vectors. Mol. Ther. 2004, 10, 616–629. [Google Scholar] [CrossRef]
- Hillus, D.; Schwarz, T.; Tober-Lau, P.; Vanshylla, K.; Hastor, H.; Thibeault, C.; Jentzsch, S.; Helbig, E.T.; Lippert, L.J.; Tscheak, P.; et al. Safety, Reactogenicity, and Immunogenicity of Homologous and Heterologous Prime-Boost Immunisation with ChAdOx1 NCoV-19 and BNT162b2: A Prospective Cohort Study. Lancet Respir. Med. 2021, 9, 1255–1265. [Google Scholar] [CrossRef]
- Creech, C.B.; Walker, S.C.; Samuels, R.J. SARS-CoV-2 Vaccines. JAMA 2021, 325, 1318–1320. [Google Scholar] [CrossRef]
- Sadoff, J.; Gray, G.; Vandebosch, A.; Cárdenas, V.; Shukarev, G.; Grinsztejn, B.; Goepfert, P.A.; Truyers, C.; Fennema, H.; Spiessens, B.; et al. Safety and Efficacy of Single-Dose Ad26.COV2.S Vaccine against COVID-19. N. Engl. J. Med. 2021, 384, 2187–2201. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Huang, J.; Zhang, Z.; Wu, J.; Zhang, J.; Hu, H.; Zhu, T.; Zhang, J.; Luo, L.; Fan, P.; et al. Safety, Tolerability, and Immunogenicity of an Aerosolised Adenovirus Type-5 Vector-Based COVID-19 Vaccine (Ad5-NCoV) in Adults: Preliminary Report of an Open-Label and Randomised Phase 1 Clinical Trial. Lancet Infect. Dis. 2021, 21, 1654–1664. [Google Scholar] [CrossRef]
- Bali, P.; Rafi, A. Immunological Mechanisms of Vaccination. Nat. Immunol. 2011, 12, 509–517. [Google Scholar]
- Wang, H.; Zhang, Y.; Huang, B.; Deng, W.; Quan, Y.; Wang, W.; Xu, W.; Zhao, Y.; Li, N.; Zhang, J.; et al. Development of an Inactivated Vaccine Candidate, BBIBP-CorV, with Potent Protection against SARS-CoV-2. Cell 2020, 182, 713–721.e9. [Google Scholar] [CrossRef]
- Kyriakidis, N.C.; López-Cortés, A.; González, E.V.; Grimaldos, A.B.; Prado, E.O. SARS-CoV-2 Vaccines Strategies: A Comprehensive Review of Phase 3 Candidates. NPJ Vaccines 2021, 6, 28. [Google Scholar] [CrossRef] [PubMed]
- Xia, S.; Zhang, Y.; Wang, Y.; Wang, H.; Yang, Y.; Gao, G.F.; Tan, W.; Wu, G.; Xu, M.; Lou, Z.; et al. Safety and Immunogenicity of an Inactivated SARS-CoV-2 Vaccine, BBIBP-CorV: A Randomised, Double-Blind, Placebo-Controlled, Phase 1/2 Trial. Lancet Infect. Dis. 2021, 21, 39–51. [Google Scholar] [CrossRef]
- Al Kaabi, N.; Zhang, Y.; Xia, S.; Yang, Y.; Al Qahtani, M.M.; Abdulrazzaq, N.; Al Nusair, M.; Hassany, M.; Jawad, J.S.; Abdalla, J.; et al. Effect of 2 Inactivated SARS-CoV-2 Vaccines on Symptomatic COVID-19 Infection in Adults: A Randomized Clinical Trial. JAMA 2021, 326, 35–45. [Google Scholar] [CrossRef]
- Saeed, B.Q.; Al-Shahrabi, R.; Alhaj, S.S.; Alkokhardi, Z.M.; Adrees, A.O. Side Effects and Perceptions Following Sinopharm COVID-19 Vaccination. Int. J. Infect. Dis. 2021, 111, 219–226. [Google Scholar] [CrossRef]
- Ghiasi, N.; Valizadeh, R.; Arabsorkhi, M.; Hoseyni, T.S.; Esfandiari, K.; Sadighpour, T.; Jahantigh, H.R. Efficacy and Side Effects of Sputnik V, Sinopharm and AstraZeneca Vaccines to Stop COVID-19; a Review and Discussion. Immunopathol. Persa 2021, 7, e31. [Google Scholar] [CrossRef]
- Tanriover, M.D.; Doğanay, H.L.; Akova, M.; Güner, H.R.; Azap, A.; Akhan, S.; Köse, Ş.; Erdinç, F.Ş.; Akalın, E.H.; Tabak, Ö.F.; et al. Efficacy and Safety of an Inactivated Whole-Virion SARS-CoV-2 Vaccine (CoronaVac): Interim Results of a Double-Blind, Randomised, Placebo-Controlled, Phase 3 Trial in Turkey. Lancet 2021, 398, 213–222. [Google Scholar] [CrossRef]
- Ulhaq, Z.S.; Soraya, G.V.; Indriana, K. Breakthrough COVID-19 Case after Full-Dose Administration of CoronaVac Vaccine. Indian J. Med. Microbiol. 2021, 39, 562. [Google Scholar] [CrossRef]
- Ramasamy, M.N.; Jessop, L.J. CoronaVac: More Data for Regulators and Policy Makers. Lancet 2021, 398, 186–188. [Google Scholar] [CrossRef]
- Roytenberg, R.; García-Sastre, A.; Li, W. Vaccine-Induced Immune Thrombotic Thrombocytopenia: What Do We Know Hitherto? Front. Med. 2023, 10, 1155727. [Google Scholar] [CrossRef]
- Ruggiero, R.; Balzano, N.; Di Napoli, R.; Mascolo, A.; Berrino, P.M.; Rafaniello, C.; Sportiello, L.; Rossi, F.; Capuano, A. Capillary Leak Syndrome Following COVID-19 Vaccination: Data from the European Pharmacovigilance Database Eudravigilance. Front. Immunol. 2022, 13, 956825. [Google Scholar] [CrossRef]
- Luxi, N.; Giovanazzi, A.; Arcolaci, A.; Bonadonna, P.; Crivellaro, M.A.; Cutroneo, P.M.; Ferrajolo, C.; Furci, F.; Guidolin, L.; Moretti, U.; et al. Allergic Reactions to COVID-19 Vaccines: Risk Factors, Frequency, Mechanisms and Management. BioDrugs 2022, 36, 443–458. [Google Scholar] [CrossRef] [PubMed]
- Moss, P.; Berenbaum, F.; Curigliano, G.; Grupper, A.; Berg, T.; Pather, S. Benefit–Risk Evaluation of COVID-19 Vaccination in Special Population Groups of Interest. Vaccine 2022, 40, 4348–4360. [Google Scholar] [CrossRef]
- The Lancet. COVID-19 in Africa: A Lesson in Solidarity. Lancet 2021, 398, 185. [Google Scholar] [CrossRef] [PubMed]
- Mathieu, E.; Ritchie, H.; Rodés-Guirao, L.; Appel, C.; Giattino, C.; Hasell, J.; Macdonald, B.; Dattani, S.; Beltekian, D.; Ortiz-Ospina, E.; et al. Coronavirus Pandemic (COVID-19). Available online: https://ourworldindata.org/coronavirus (accessed on 9 January 2023).
- Herzog, L.M.; Norheim, O.F.; Emanuel, E.J.; McCoy, M.S. Covax Must Go beyond Proportional Allocation of COVID Vaccines to Ensure Fair and Equitable Access. BMJ 2021, 372, m4853. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). Fair Allocation Mechanism for COVID-19 Vaccines through the COVAX Facility. Available online: https://www.who.int/publications/m/item/fair-allocation-mechanism-for-covid-19-vaccines-through-the-covax-facility (accessed on 9 April 2023).
- World Health Organization (WHO). WHO Lists Two Additional COVID-19 Vaccines for Emergency Use and COVAX Roll-Out. Available online: https://www.who.int/news/item/15-02-2021-who-lists-two-additional-covid-19-vaccines-for-emergency-use-and-covax-roll-out (accessed on 1 February 2023).
- Mwai, P. COVID-19 Africa: Who Is Getting the Vaccine? Available online: https://www.bbc.com/news/56100076 (accessed on 2 April 2021).
- Our World in Data Coronavirus (COVID-19) Vaccinations. Available online: https://ourworldindata.org/covid-vaccinations#what-share-of-the-population-has-received-at-least-one-dose-of-vaccine (accessed on 19 March 2023).
- World Health Organization (WHO). Africa COVID-19 Vaccination Dashboard. Available online: https://app.powerbi.com/view?r=eyJrIjoiOTI0ZDlhZWEtMjUxMC00ZDhhLWFjOTYtYjZlMGYzOWI4NGIwIiwidCI6ImY2MTBjMGI3LWJkMjQtNGIzOS04MTBiLTNkYzI4MGFmYjU5MCIsImMiOjh9 (accessed on 9 April 2023).
- Kabakama, S.; Konje, E.T.; Dinga, J.N.; Kishamawe, C.; Morhason-Bello, I.; Hayombe, P.; Adeyemi, O.; Chimuka, E.; Lumu, I.; Amuasi, J.; et al. Commentary on COVID-19 Vaccine Hesitancy in Sub-Saharan Africa. Trop. Med. Infect. Dis. 2022, 7, 130. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). Joint Statement on Dose Donations of COVID-19 Vaccines to African Countries. Available online: https://www.who.int/news/item/29-11-2021-joint-statement-on-dose-donations-of-covid-19-vaccines-to-african-countries (accessed on 9 April 2023).
- World Health Organization (WHO). Strategy to Achieve Global COVID-19 Vaccination by Mid-2022. Available online: https://www.who.int/publications/m/item/strategy-to-achieve-global-covid-19-vaccination-by-mid-2022 (accessed on 1 February 2023).
- Nature. Africa Is Bringing Vaccine Manufacturing Home. Nature 2022, 602, 184. [Google Scholar] [CrossRef] [PubMed]
- Paton, J. BioNTech to Send COVID Vaccine Production Units to Africa. Available online: https://www.bloomberg.com/news/articles/2022-02-16/biontech-to-send-covid-vaccine-production-units-to-africa#xj4y7vzkg (accessed on 12 March 2023).
- Wellcome. Scaling Up African Vaccine Manufacturing Capacity. Available online: https://cms.wellcome.org/sites/default/files/2023-01/Wellcome-Biovac-BCG-Scaling-up-African-vaccine-manufacturing-capacity-report-2023_0.pdf (accessed on 24 January 2024).
- World Health Organization (WHO). Africa Continued Delivery of Immunization Services in the Context of COVID-19 Pandemic. Available online: https://www.afro.who.int/news/continued-delivery-immunization-services-context-covid-19-pandemic (accessed on 28 March 2023).
- Hakizimana, D.; Ntizimira, C.; Mbituyumuremyi, A.; Hakizimana, E.; Mahmoud, H.; Birindabagabo, P.; Musanabaganwa, C.; Gashumba, D. The Impact of COVID-19 on Malaria Services in Three High Endemic Districts in Rwanda: A Mixed-Method Study. Malar. J. 2022, 21, 48. [Google Scholar] [CrossRef] [PubMed]
- Desta, A.A.; Woldearegay, T.W.; Gebremeskel, E.; Alemayehu, M.; Getachew, T.; Gebregzabiher, G.; Ghebremedhin, K.D.; Zgita, D.N.; Aregawi, A.B.; Redae, G. Impacts of COVID-19 on Essential Health Services in Tigray, Northern Ethiopia: A Pre-Post Study. PLoS ONE 2021, 16, e0256330. [Google Scholar] [CrossRef] [PubMed]
- Jensen, C.; McKerrow, N.H. Child Health Services during a COVID-19 Outbreak in KwaZulu-Natal Province, South Africa. S. Afr. Med. J. 2020, 111, 114. [Google Scholar] [CrossRef]
- Harvey, R. The Ignored Pandemic: The Dual Crises of Gender-Based Violence and COVID-19; Oxfam: Oxford, UK, 2021. [Google Scholar]
- Mednick, S. COVID-19 Brings Abuse and Other Fears to Displaced Women in South Sudan. Available online: https://www.thenewhumanitarian.org/feature/2020/05/06/coronavirus-south-sudan-women-abuse-gender-violence (accessed on 24 January 2024).
- International Labour Organization. COVID-19 Impact on Child Labour and Forced Labour: The Response of the IPEC+ Flagship Programme; International Labour Organization: Geneva, Switzerland, 2020. [Google Scholar]
- Xavier, J.S.; Moir, M.; Tegally, H.; Sitharam, N.; Abdool Karim, W.; San, J.E.; Linhares, J.; Wilkinson, E.; Ascher, D.B.; Baxter, C.; et al. SARS-CoV-2 Africa Dashboard for Real-Time COVID-19 Information. Nat. Microbiol. 2023, 8, 1–4. [Google Scholar] [CrossRef]
- Tegally, H.; Wilkinson, E.; Althaus, C.L.; Giovanetti, M.; San, J.E.; Giandhari, J.; Pillay, S.; Naidoo, Y.; Ramphal, U.; Msomi, N.; et al. Rapid Replacement of the Beta Variant by the Delta Variant in South Africa. medRxiv 2021. [Google Scholar] [CrossRef]
- Wickramasinghe, D.; Wickramasinghe, N.; Kamburugamuwa, S.A.; Arambepola, C.; Samarasekera, D.N. Correlation between Immunity from BCG and the Morbidity and Mortality of COVID-19. Trop. Dis. Travel. Med. Vaccines 2020, 6, 17. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Vaccine Type | Name | Characterization | Company | Vaccine Efficacy | Storage | Systemic Reactogenicity | References |
|---|---|---|---|---|---|---|---|
| mRNA vaccines | BNT162b2 | Nucleoside modified mRNA encoding full length S protein (stabilized in prefusion conformation) | Pfizer, BioNTech, Fosun Pharma | 95% effective in preventing symptomatic COVID-19; 100% effective in preventing severe COVID-19 | Six months at −70 °C or stored at RT for less than 2 h when undiluted. | Fatigue, headache, fever, and chills. Swelling, pain, and redness at the site of injection. | [191,196] |
| mRNA-1273 | Synthetic mRNA encoding the pre-fusion stabilized S protein | Moderna | 94.1% effective in preventing symptomatic COVID-19 100% effective in preventing severe COVID-19 | Stored for less than 12 h at RT; for 30 days at 2 to 8 °C; more than 1 month stored at −25 °C to −15 °C. | Swelling, pain, and redness at the site of injection. Fatigue, headache, fever, vomiting, chills, myalgia, urticarial, and arthralgia. | [192,196] |
| Vaccine Type | Name | Characterization | Company | Vaccine Efficacy | Storage | Systemic Reactogenicity | References |
|---|---|---|---|---|---|---|---|
| Viral Vector Vaccines | ChAdOx1 nCoV-19 (AZD1222) | Replication-deficient chimpanzee viral vector with SARS-CoV-2 S protein | University of Oxford, AstraZeneca | 64.1% after 1st dose and 70.4% after the 2nd dose preventing symptomatic COVID-19; 100% effective in preventing severe COVID-19. | 2–8 °C for 6 months | Fatigue, myalgia, headache, feverishness or chills, and fever higher than 38 °C | [198,199] |
| Ad26.COV2.S | Recombinant replication-incompetent human viral vector encoding a full-length, stabilized SARS-CoV-2 S protein | Janssen Pharmaceutical Companies (Johnson & Johnson) | 64% efficacy against moderate-to-severe COVID-19; 81.7% efficacy against severe COVID-19 | Stored at 2 to 8 °C for less than 6 months; more than 6 months −20 °C | Injection site pain, headache, fatigue, myalgia, nausea, venous thromboembolic events, seizures, and tinnitus | [188,200] | |
| Ad5-nCoV | Replication-defective adenovirus type-5 vectored vaccine that encodes the SARS-CoV-2 spike protein | CanSinoBIO | 65 to 69% efficacy against symptomatic COVID-19; 90 to 95% efficacy against severe COVID-19 | Stored at 2–8 °C | Fever, fatigue, and headache | [201] | |
| Sputnik V | Full-length SARS-CoV-2 glycoprotein S carried by adenoviral vector | The Gamaleya National Center | 91.6% efficacy against symptomatic COVID-19; 100% effective in preventing severe COVID-19. | Stored at 2 to 8 °C for less than 6 months; −18 °C in liquid form. | Mild pain at the injection site, fever, headache, fatigue, and muscle aches | [199] |
| Vaccine Type | Name | Characterization | Company | Vaccine Efficacy | Storage | Systemic Reactogenicity | References |
|---|---|---|---|---|---|---|---|
| Inactivated Vaccines | BBIBP-CorV | Inactivated HB02 strain with aluminum hydroxide adjuvant | Sinopharm, Beijing Institute of Biological Products | 78.1% efficacy against symptomatic COVID-19; 90 to 95% efficacy against severe COVID-19 | 2–8 °C (unknown lifespan) | Pain, fatigue, headache, lethargy, and tenderness. | [188,206,207] |
| CoronaVac | Inactivated CN02 strain with aluminum hydroxide adjuvant | Sinovac Biotech | Variable efficacy when comparing different countries (50.7–83.5%); 100% efficacy against severe COVID-19 | 2–8 °C (unknown lifespan) | Injection site pain and fatigue are the most common events; adverse events include an allergic reaction | [188,208,209,210,211] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pêgo, A.C.; Lima, I.S.; Gozzelino, R. Addressing Inequality in the COVID-19 Pandemic in Africa: A Snapshot from Clinical Symptoms to Vaccine Distribution. COVID 2024, 4, 170-190. https://doi.org/10.3390/covid4020014
Pêgo AC, Lima IS, Gozzelino R. Addressing Inequality in the COVID-19 Pandemic in Africa: A Snapshot from Clinical Symptoms to Vaccine Distribution. COVID. 2024; 4(2):170-190. https://doi.org/10.3390/covid4020014
Chicago/Turabian StylePêgo, Ana Catarina, Illyane Sofia Lima, and Raffaella Gozzelino. 2024. "Addressing Inequality in the COVID-19 Pandemic in Africa: A Snapshot from Clinical Symptoms to Vaccine Distribution" COVID 4, no. 2: 170-190. https://doi.org/10.3390/covid4020014
APA StylePêgo, A. C., Lima, I. S., & Gozzelino, R. (2024). Addressing Inequality in the COVID-19 Pandemic in Africa: A Snapshot from Clinical Symptoms to Vaccine Distribution. COVID, 4(2), 170-190. https://doi.org/10.3390/covid4020014