Abstract
The global pandemic sparked by the emergence of SARS-CoV-2 and its variants has imposed a substantial burden of morbidity and mortality. Central to the battle against these viral threats is the immune response, with a spotlight on the pivotal role played by neutralizing antibodies. This comprehensive review delves into current research, unravelling the dual functionality of neutralizing antibodies acting as formidable barriers to viral replication and crucial facilitators of adaptive immune memory. Beyond this dual purpose, the review illuminates the nuanced variability characterizing neutralizing antibody responses to SARS-CoV-2. Emphasizing the dynamic nature of these responses, the review advocates for the plausible challenges in targeted therapeutic interventions. This review also attempts to compare various vaccination approaches and their impact on SARS-CoV-2, as well as offer insights into various Omicron variations. Recognizing the ever-evolving viral landscape, this exploration underscores the necessity of flexible approaches to address the diverse challenges posed by SARS-CoV-2 and its variants, contributing valuable insights to the ongoing global efforts in pandemic mitigation and public health safeguarding.
1. Introduction
The severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) is the source of COVID-19. Since its initial discovery in Wuhan, China in December 2019, the epidemic has spread throughout the world. When an infected individual coughs or sneezes, the virus mainly spreads by respiratory droplets. It can also spread by contacting contaminated surfaces. The immune system’s involvement in SARS-CoV-2 infection is critical to understanding the dynamics of COVID-19, from early identification and containment to the formation of memory responses [1]. To contain and eliminate this novel coronavirus, the immune system must synchronize both an innate and adaptive defense. One of the intricate and dynamic interactions between the immune system in SARS-CoV-2 infection is the Innate Immune Response, which comprises the Recognition and Early Response phase, when the innate immune system acts as the first line of defense against SARS-CoV-2 [2]. This interaction involves a variety of immune cell types and their reactions. Pattern recognition receptors (PRRs), particularly Toll-like receptors (TLRs) and RIG-I-like receptors (RLRs), set off a sequence of events that recognize components of viruses. Following their identification of the virus, dendritic cells and macrophages produce chemokines and cytokines that elicit inflammation and attract immune cells to the site of infection. Natural killer cells, or NK cells, are also crucial for early antiviral defense. They locate and eliminate infected cells by inducing apoptosis, stopping the virus from spreading [3]. NK cells function as a bridge between the innate and adaptive immune responses by influencing the activation of other immune cells. Initially, B cells are exposed to viral antigens by antigen-presenting cells, specifically dendritic cells, which activate B cells and start the adaptive immune response against the pathogen. This link determines when a robust immune response begins. Then, B cell-mediated immunity starts to work, which helps to target several viral proteins, most notably the spike protein [4]. Neutralizing antibodies could suppress an infection by preventing the virus from attaching to host cells. It also consists of helper T cells, or CD4+ T cells, which facilitate B cells’ production of antibodies and the activation of cytotoxic T cells [5]. They are essential to the immune response’s maintenance and coordination. Cytotoxic T cells, or CD8+ T cells, subsequently identify and eliminate the infected cells. They play a crucial role in controlling the virus’s reproduction and halting its propagation throughout the host [6]. Following an injection or infection, B lymphocytes for memory are generated. These cells “remember” the virus and respond fast when it is encountered again [7]. Furthermore, following re-infection, memory B cells differentiate into plasma cells that generate antibodies, leading to a more potent and quick antibody response. Moreover, memory T cells—both CD4+ and CD8+—help to maintain protection over the long run. They can recognize viral antigens and react more rapidly and effectively in the event of a re-infection [8]. However, an excess of pro-inflammatory cytokines and chemokines that trigger a cytokine storm and intensify severe COVID-19 symptoms, ultimately leading to tissue destruction and organ failure, deteriorates the patient’s state. As a result, by supporting immune response control, preventing excessive inflammation and immunopathology, and balancing pro- and anti-inflammatory signals, T regulatory (Treg) cells serve a crucial role in preserving the delicate balance between immunopathology and protective immunity. New variations of SARS-CoV-2 can evade immune response, even though they have been shown to be effective against several strains of COVID-19 and are employed by our bodies as a defense mechanism.
As of right now, neutralization antibodies are being developed against novel strains, like the JN.1 variation, which was discovered in August 2023. Since then, it has been spreading rapidly and infecting a sizable portion of the population. Based on the data currently available, JN.1 is the most effective virus in terms of evading immune response. This is because of the antigenic diversity it has acquired from the Omicron subvariant and the addition of the RBD (S-L455) mutation [9], which allows it to become less dependent on ACE 2 binding in humans. S-L455 is located between the ACE 2 and RBD domain. Additionally, three mutations in non-S proteins have been discovered in it [10], which is concerning because it can elude immune response in the methods indicated above. Therefore, the emphasis of this review is to clarify the difficulties therapeutic antibodies have in neutralizing SARS-CoV-2 as well as the conceivable ways in which neutralizing antibodies can control an illness. This review also attempts to compare various vaccination approaches and their impact on SARS-CoV-2, as well as offer insights into various Omicron variations [11].
2. Materials and Methods
The purpose of conducting a literature review on neutralizing antibody response against SARS-CoV-2 was to evaluate the treatment options and the risk factors associated with SARS-CoV-2, as well as to provide insight on the challenges of therapeutic monoclonal antibodies in neutralizing SARS-CoV-2 response along with its mechanism of action. All the literature that was found in the searches was critically reviewed, both in terms of quality reporting and usefulness to policymakers and decision-makers. The literature searches dated from 1 August 2012 to 4 July 2023 were included from different journals and publishers. The review aims to provide a comprehensive analysis of the neutralizing antibody response against SARS-CoV-2, thereby enhancing our understanding of COVID-19 immunity.
3. Role of Neutralizing Antibodies in Immune Response
Neutralizing antibodies are a class of antibodies that play a crucial role in the immune response against viral infections, including SARS-CoV-2. These specialized antibodies are a part of the adaptive immune system [12]. When a virus, such as SARS-CoV-2, enters the body, the immune system recognizes it as a foreign invader and triggers an immune response. B cells, a type of white blood cell, are activated to produce the antibodies specific to virus through the Identification of Antigens during their first encounter; i.e., when B cells encounter the virus or any of its constituent parts, including the spike protein, during an infection or immunization, they become activated. They also have sites for Antigen Binding known as B cell receptors (BCRs) on their surface. The activation mechanism of the virus is started when these BCRs attach to antigens on its surface, after which the virus is internalized by the process of endocytosis and breakdown of the viral antigen is done inside the B cell. Then, helper T cells interact with the presentation of antigen through major histocompatibility complex class II (MHC-II) molecules on the surface of B lymphocytes that are used to present the processed viral antigens [13]. The helper T cell is activated by this contact in addition to co-stimulatory signals. Further, cytokines are released by activated helper T cells, thereby giving vital signals to B cells and encouraging their activation and differentiation. A population of memory B cells and plasma cells are thus produced because of the activated B cell’s clonal growth. A portion of the B cells that have been stimulated undergoes differentiation into plasma cells. These cells have been trained to produce antibodies. During an infection or vaccination, plasma cells generate a significant number of antibodies that are particular to the viral antigens that were encountered. Antibodies are then secreted by plasma cells into the circulation and other body fluids. By this process the spike protein and other components of SARS-CoV-2 are selectively recognized and bound to by the antibodies. Antibodies are responsible for neutralization, which is achieved by stopping the virus from clinging to or invading host cells, through opsonization and through triggering the complement system [14].
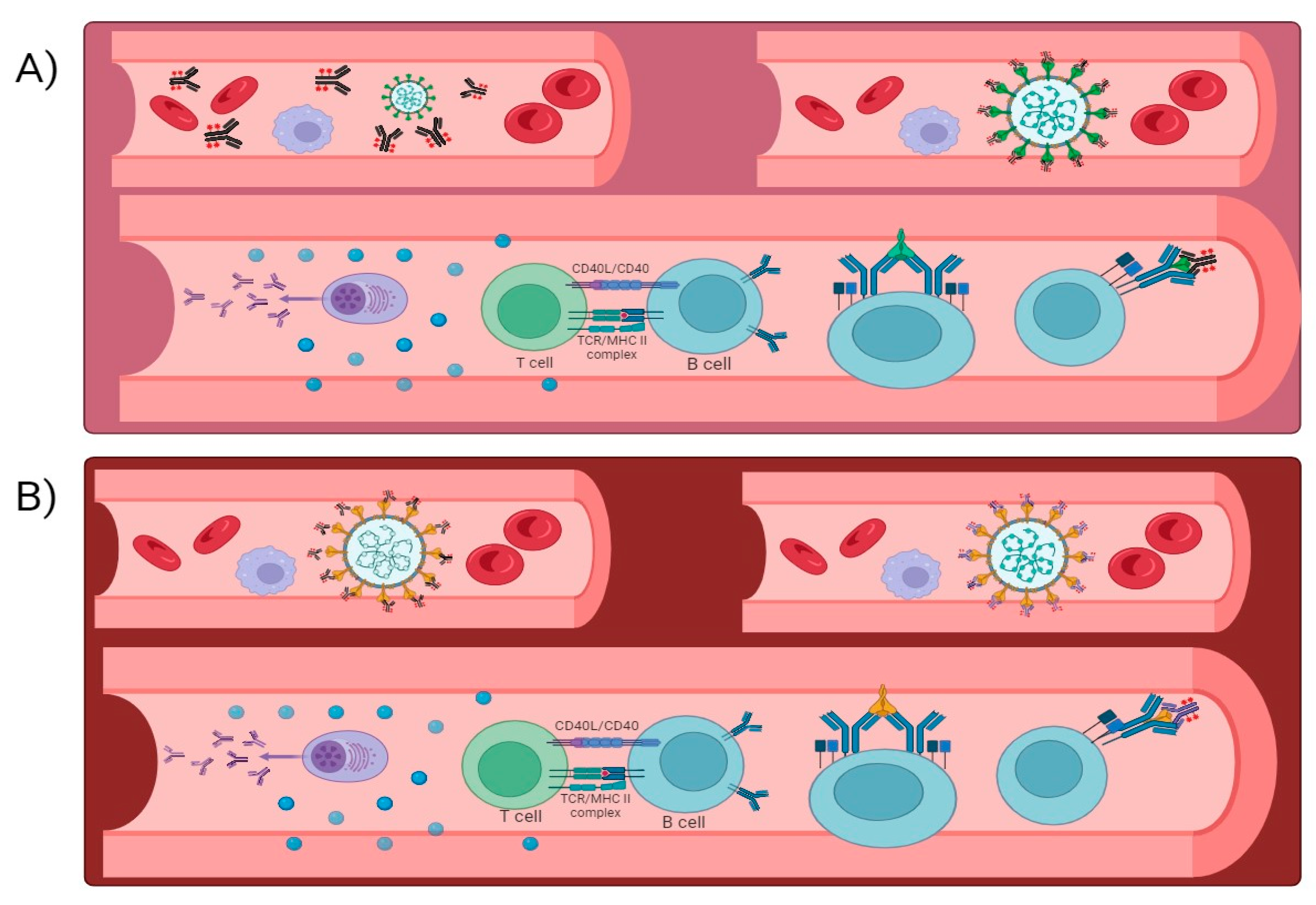
Neutralizing antibodies have been used as therapeutic agents to treat COVID-19 patients, either by isolating antibodies from convalescent individuals or by generating them through monoclonal antibody therapies [15]. Neutralizing antibodies contribute to the immune response through targeting the pathogen, as neutralizing antibodies are highly specialized proteins which binds to the antigen. For example, in the case of SARS-CoV-2, the spike protein on the virus’s surface is a primary target for neutralizing antibodies. By binding to these antigens, neutralizing antibodies directly recognize and tag the virus for destruction [16]; they can also neutralize the infectivity of the virus by binding to viral antigens and preventing their attachment and entry into the host cells, they recruit other immune cells, such as macrophages and natural killer (NK) cells, through a process called antibody-dependent cellular cytotoxicity (ADCC), and they activate complementary system leading to the formation of membrane attack complexes which punctures the viral membrane, causing the virus to lyse or rupture [17] (Figure 1). Neutralizing antibodies that are responsible for controlling infections are mentioned in Table 1. Vaccination strategies leverage this immune response by priming the body to generate neutralizing antibodies without causing severe illness. This proactive approach helps in the establishment of immunity from the virus, protecting vulnerable populations, and ultimately reducing the impact of infectious diseases like COVID-19. As research continues, a deeper understanding of neutralizing antibodies’ dynamics and interactions with different pathogens will aid in developing more effective vaccines and treatments for various viral infections [18]. However, it is essential to understand that the neutralizing antibody response to SARS-CoV-2 can vary between individuals and may decline over time. Factors such as the severity of the infection, age, underlying health conditions, and the emergence of viral variants can influence the level and potency of neutralizing antibodies [13].
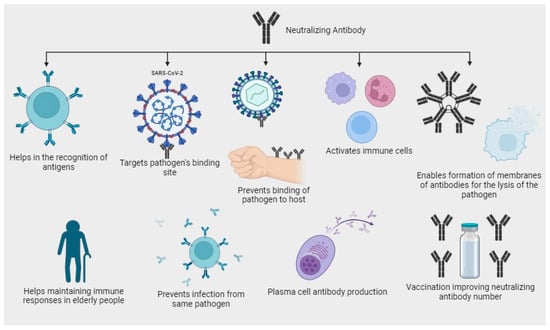
Figure 1.
Diagram depicting the potential antibody neutralization mechanisms targeting SARS-CoV-2.

Table 1.
List of neutralizing antibodies in controlling an infection.
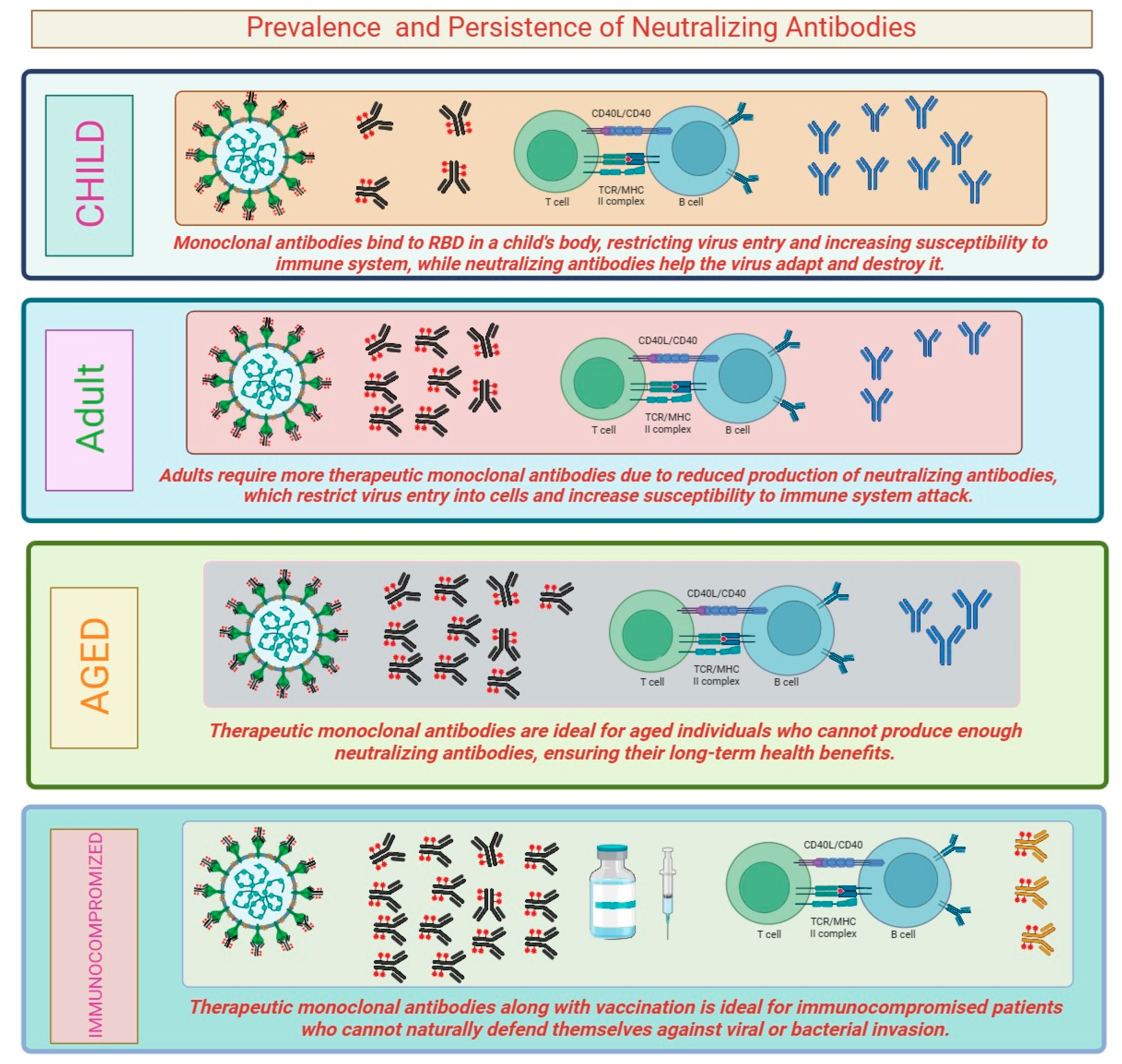
3.1. Prevalence and Persistence of Neutralizing Antibodies
The prevalence and persistence of neutralizing antibodies can vary depending on several factors, including the pathogen, the individual’s immune response, and the duration of follow-up. In the case of viral infections, including SARS-CoV-2, neutralizing antibodies are typically detectable after an individual has been infected or vaccinated. The prevalence of neutralizing antibodies can be high during the acute phase of the infection or shortly after vaccination. However, the levels of these antibodies may decline over time. Studies have shown that neutralizing antibodies against SARS-CoV-2 can persist for several months, providing some level of protection against reinfection. Vaccination plays a crucial role in inducing neutralizing antibodies. In case of many viral vaccines, including measles, mumps, and rubella (MMR) and hepatitis B vaccines, neutralizing antibodies can persist for years and even decades, providing long-lasting protection against the respective infections [25], antibodies can also be generated in response to bacterial infections. For example, tetanus and diphtheria vaccines induce the production of neutralizing antibodies against the toxins produced by these bacteria. The variation in the response of neutralizing antibodies in this case is because Clostridium tetani is a bacterial infection which produces stable toxins and does not itself undergo any frequent genetic changes, while SARS-CoV-2, a new coronavirus, can evade the immune response and undergoes genetic mutations over time. In immunocompromised individuals, the prevalence and persistence of neutralizing antibodies is affected. These individuals have a reduced ability to produce and maintain a robust immune response, leading to lower levels of neutralizing antibodies and potentially increased susceptibility to infections [26]. Prevalence and persistence of neutralizing antibodies can influence the risk of reinfection and the impact of viral variants. Studies have shown that individuals with higher levels of neutralizing antibodies are less likely to experience severe disease upon reinfection. However, the duration of protection provided by neutralizing antibodies may not be uniform for all infections (Figure 2). In some cases, waning levels of neutralizing antibodies over time may lead to a higher susceptibility to reinfection, particularly with new variants of the virus that might partially evade the immune response [27]. Some infections, such as influenza (flu), are caused by viruses that undergo frequent antigenic changes, leading to seasonal outbreaks. The prevalence and persistence of neutralizing antibodies against influenza viruses can vary between different strains and may not always confer complete protection. As a result, seasonal flu vaccines are updated regularly to match circulating strains and to stimulate the production of strain-specific neutralizing antibodies [28]. In chronic viral infections, such as HIV and hepatitis C, neutralizing antibodies may be generated but the virus can evade the immune response and persist in the body. These viruses have evolved mechanisms to evade neutralization by mutating their surface proteins, thus making it challenging to maintain the persistence of neutralizing antibody responses [29]. The timing of vaccination can influence the persistence of neutralizing antibodies. Some vaccines may require booster doses to maintain protective levels of neutralizing antibodies over an extended period. For example, tetanus and diphtheria vaccines requires booster shots after every ten years to ensure sustained protection [30]. Neutralizing antibodies can vary with age. In some cases, older individuals may experience reduced immune responses and a decline in neutralizing antibody production, which may affect the effectiveness of certain vaccines. It is essential to note that the prevalence and persistence of neutralizing antibodies are dynamic and may change over time. Immune responses can be influenced by various factors including age, health status, and the presence of coexisting medical conditions.
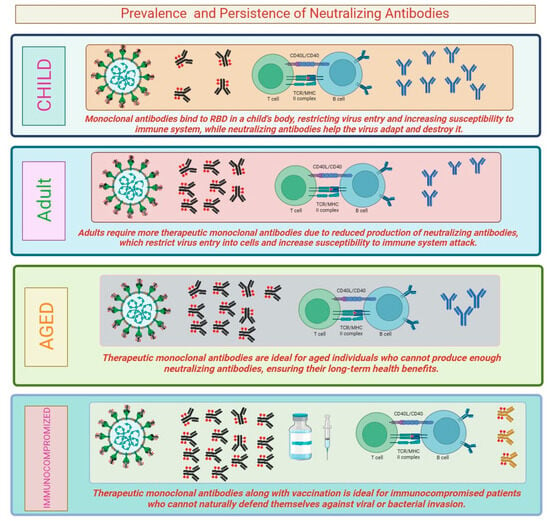
Figure 2.
Persistence level of neutralizing antibody in persons of different categories.
3.2. Effectiveness of Vaccination on Neutralizing Antibodies
Effectiveness of vaccination on neutralizing antibody is a critical aspect of immune response generated by vaccination. By the introduction of harmless form of the virus or its components (e.g., viral proteins), vaccines prime the immune system to recognize the virus as a threat and mount a targeted response [31]. Here is how vaccination improves the effectiveness of neutralizing antibodies: (a) Production of Neutralizing Antibodies: Vaccination prompts the immune system, specifically B cells, to produce neutralizing antibodies against the virus’s specific antigens. For example, in the case of COVID-19 mRNA vaccines (Pfizer-BioNTech, Moderna), the vaccines contain genetic instructions to produce the spike protein found on the surface of the SARS-CoV-2 virus. The immune system recognizes the spike protein as foreign and start producing neutralizing antibodies against it [32]. (b) Increase in Neutralizing Antibody Levels: Vaccination leads to an increase in the levels of neutralizing antibodies in the bloodstream. Although vaccination aids the functioning of neutralizing antibodies, it may not be helpful for all, hence requiring us to observe every aspect possible. Some of the aspects are mentioned in Table 2.
Table 2.
Comparison of COVID-19 vaccination strategies and their effects.
4. Omicron Variant and Immune Escape
The emergence of the SARS-CoV-2 Omicron variant has raised significant concerns due to its high number of mutations. The spike protein is a key target for neutralizing antibodies, and the substantial mutations in this region have led to questions about the variant’s potential to evade the immune response [64]. The rapid spread of the Omicron variant has raised concerns about its ability to evade immune response. The Omicron variant, also known as B.1.1.529, was first detected in November 2021 and since then has spread to numerous countries [65]. This variant’s spike protein has more than 30 mutations, including the key mutation in receptor-binding domain (RBD) which may influence its interaction with the host’s immune system [66]. Several mutations in RBD and other regions of the spike protein are thought to influence the variant’s ability to partially evade neutralizing antibodies produced in response to prior infections or vaccinations [67]. Studies have suggested that certain monoclonal antibodies and convalescent plasma may have reduced effectiveness against the Omicron due to certain mutations on the spike protein on which monoclonal antibodies bind for virus neutralization. Additionally, breakthrough infections in fully vaccinated individuals have been reported, indicating partial immune escape from existing vaccines [68]. Initial reports on the Omicron variant’s impact on vaccine-induced immunity have been concerning. Despite this, vaccines have still demonstrated a degree of protection against the disease, including the situation of hospitalization and death caused by the Omicron variant. Vaccine developers have begun to modify existing vaccines to better match the Omicron variant’s spike protein. Early data from these adapted vaccines have shown improved neutralizing activity against the variant, offering hope for maintaining protection against emerging variants [69]. Further information on omicron variants along with their site of action is summarized in Table 3.
Table 3.
Omicron variant overview: Mechanism of immune escape, challenges, immune response, site of action, and drawbacks.
5. Therapeutic Monoclonal Antibodies and Neutralization
Development of therapeutic monoclonal antibodies targeting the viral spike protein’s RBD, and other critical regions has offered a promising avenue for treatment. These monoclonal antibodies aim to neutralize the virus and limit its replication, thereby mitigating the severity of the disease and preventing hospitalizations [81]. Therapeutic monoclonal antibodies neutralize SARS-CoV-2 through various mechanisms. They typically target the RBD, which is essential for viral entry into host cells. Casirivimab and Imdevimab form a cocktail that binds to the RBD, blocking viral attachment and entry [82]. Similarly, Sotrovimab targets a conserved epitope on the RBD, hindering viral entry and replication. REGN-COV2, a combination of casirivimab and imdevimab, and tixagevimab with cilgavimab each target non-overlapping epitopes on the RBD to reduce the likelihood of escape mutants. Therapeutic monoclonal antibodies not only directly neutralize the virus but also trigger an immune response (Figure 3). The spike protein’s RBD is the potential site of action for a variety of therapeutic monoclonal antibodies. By engaging the immune system, these antibodies aid in viral clearance and limits viral replication [83] (please refer Table 4 for more information). The immune response may include the activation of natural killer (NK) cells, phagocytes, and other components of the immune system to mount an antiviral defense. The primary site of action for most therapeutic monoclonal antibodies is the RBD. However, the virus’s continuous evolution and emergence of new variants pose challenges in targeting specific epitopes effectively [84]. Variants like B.1.1.529 have mutations in the RBD that may reduce the binding efficacy of certain monoclonal antibodies, leading to potential breakthrough infections. Monoclonal antibody production entails advanced biotechnological procedures, commonly employing mammalian cell cultures. These procedures demand a significant number of resources and necessitate strict quality control measures to guarantee the uniformity, purity, and effectiveness of the end product. Expanding manufacturing to satisfy worldwide demand is a difficult task. Constructing and running the necessary infrastructure for mAb production is costly, and the process of developing these manufacturing capabilities can cause significant delays in making them widely accessible. Supply chain constraints arise due to the intricate nature of the mAb supply chain, spanning from the procurement of raw ingredients to the ultimate distribution, leading to potential shortages. This is particularly troublesome in low- and middle-income nations, where access to these medicines may be limited [85]. The production expenses contribute to the high price of mAbs, making them less accessible to broad portions of the worldwide population. Health systems in many regions of the world may struggle to fund these therapies, limiting their distribution and usage. Administration of mAbs can induce immunological reactions, ranging from moderate symptoms like fever and chills to severe anaphylaxis, a life-threatening allergic reaction. These potential dangers require careful observation before and after the administration of the treatment, which can make it more difficult to carry out in environments without sufficient medical oversight. In order to reduce these risks, patients frequently need to take pre-medications (such as antihistamines and corticosteroids) and follow precise infusion protocols. This can increase the overall burden and complexity of the treatment [86]. Viruses, especially RNA viruses like SARS-CoV-2, mutate rapidly. Variants with mutations in the areas targeted by mAbs can develop, potentially lowering the efficiency of these antibodies. This needs continual surveillance and the creation of novel mAbs to keep pace with viral evolution. The necessity for variant-specific treatments means that mAbs may have a limited useful lifespan, requiring continual research and development to meet new strains. This dynamic further complicates the manufacturing and distribution processes, as fresh formulas must be regularly developed and disseminated.
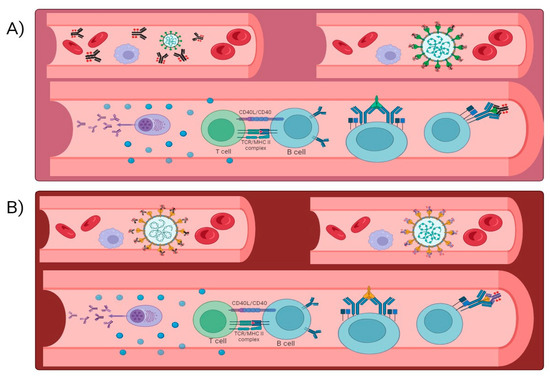
Figure 3.
(A,B) When an antigen enters the human body, it elicits an immune response after which plasma cells are formed and the antigen is memorized. This process generally takes time and can be fastened with the help of therapeutic antibodies.
Table 4.
List of therapeutic monoclonal antibodies, depicting possible challenges in neutralizing SARS-CoV-2 response along with its mechanism of action.
6. Strategies for Adaptive Pandemic Control
Adaptive pandemic control strategies for handling COVID-19 and getting ready for future pandemics depend on adaptive pandemic control plans. These techniques call for a flexible, evidence-based strategy that one can change depending on the situation. Early detection and surveillance constitute the fundamental components. Early discovery of new diseases depends on strong worldwide surveillance systems being established. Networks should combine data from digital platforms, labs, and hospitals to track illness patterns instantly. Furthermore, thorough genome sequencing is essential to find and monitor viral mutations and variations, enabling quick changes in control strategies. Also, rapid response and containment techniques are quite important. Developing and improving methods for the quick isolation of sick people and quarantining exposed people will greatly help to stop the virus from spreading. This calls for building surge capacity for quarantine facilities and guaranteeing compliance via social support systems and legal systems [117]. Quickly identifying and managing epidemics depends on improving contact tracing capacity, through conventional techniques as well as digital tools including mobile apps. By separating instances before they spread to others, good contact tracing helps to stop general transmission. Using localized data, adaptive public health measures entail conducting focused actions. For instance, selective application of regional lockdowns, mask requirements, and gathering restrictions help to balance limiting the transmission of the infection by reducing social effects. Real-time data constantly evaluating transmission hazards lets public health policies be dynamically changed. By helping to predict possible epidemics, predictive models allow proactive scaling-up of treatments to prevent catastrophes [118]. Keystones of pandemic control are vaccination development and distribution. Investing in fast-developing platforms for vaccines, such mRNA technology, helps to create vaccines for new diseases quickly. Establishing pre-existing vaccination candidates that can be rapidly altered for new challenges is absolutely vital. Vaccine equitable distribution under coordinated international initiatives such as COVAX is essential to stop the virus from spreading and evolving among unprotected people, therefore undermining world attempts to manage the epidemic. Public compliance with health standards depends on public communication and engagement [119]. Maintaining open and honest communication fosters confidence and promotes following of policies. Giving the public correct and timely information on hazards, policies, and the justification for decisions allows them to appreciate the significance of the actions. Especially in diverse communities, using behavioral science techniques such nudges and culturally customized messaging helps to increase compliance even more. Health system resilience is improving healthcare systems’ ability to manage major epidemic outbreaks. This includes increasing ICU beds, ventilator availability, and staffing of healthcare workers through pre-emptive investments and planning [120]. Creating integrated care networks that links hospital systems, primary care, and public health guarantees coordinated and effective responses [121]. Using digital health tools and telemedicine can help to preserve continuity of treatment even in cases of strained healthcare systems. The emphasis of research and development should be on building structures for quick mobilization of ideas. This helps pathogen research, pharmaceutical development, and intervention testing right before an epidemic start. Crucially are adaptive clinical studies capable of pivoting fast to assess newly developed treatments as they become accessible. Comprehensive solutions addressing all facets of pandemic control depend on multidisciplinary collaboration among virologists, epidemiologists, social scientists, and other experts [122]. Pandemic readiness and response depend critically on international cooperation. By means of institutions such as the World Health Organization (WHO), strengthening worldwide coordination guarantees that reactions are coordinated internationally. By use of data, tools, and best practices, sharing helps present a cohesive front against the epidemic. Ensuring a worldwide coordinated response depends on negotiations and application of pandemic preparation agreements establishing criteria for openness, resource sharing, and mutual aid during crises. Managing the difficult issues that develop during pandemics requires both ethical and legal frameworks. Clearly defining legislative frameworks helps to establish the extent of emergency authority including resource allocation, mandated vaccines, and lockdowns. These systems ought to reconcile public health requirements with personal liberties. Especially with relation to resource allocation, treatment prioritizing, and balancing individual freedoms with community health, developing ethical rules for decision-making during pandemics is also vital [123]. Finally, pandemic readiness depends mostly on education and training. Improving readiness is achieved by means of bettering public health campaigns, training courses for public health officials, healthcare professionals, and the general population. Frequent drills, simulations, and ongoing education on newly developing infectious diseases guarantee that every participant is ready to react. Including communities in pandemic preparedness planning and response initiatives guarantees that policies are culturally relevant and generally accepted, therefore strengthening the public health response by means of more resilient and cooperative approach.
7. Conclusions
SARS-CoV-2, a formidable adversary since its emergence on 12 December 2019, has led to an ongoing global challenge. Neutralizing antibodies, crucial in curtailing viral efficiency, face new challenges with emerging variants capable of modulating their receptor-binding domains (RBD). This adaptability threatens the efficacy of existing neutralizing antibodies, potentially necessitating additional booster doses for robust viral detection. As we confront the evolving virus, our response must also evolve. It is imperative to explore innovative strategies to effectively manage the mutating strains. Current therapeutic monoclonal antibodies offer promise, but their potential breakthrough hinges on a strategic synthesis of their characteristics. Combining the strengths of available therapeutic monoclonal antibodies into a unified approach could yield a more potent and versatile solution. This underscores the importance of ongoing research and collaborative efforts to navigate the intricate landscape of viral evolution, ensuring that our strategies for combating SARS-CoV-2 align with the dynamic challenges posed by its mutating strains. In the face of uncertainty, adaptability and innovation are key to staying ahead in the ongoing battle against this resilient virus.
Author Contributions
Conceptualization: P.D.; data curation: V.A.I. and A.M.; formal analysis: V.A.I.; investigation: A.M.; supervision: P.D.; writing—original manuscript: V.A.I. and A.M.; writing—review and editing: P.D., D.K., V.A.I. and A.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Vianello, C.; Strozzi, F.; Mocellin, P.; Cimetta, E.; Fabiano, B.; Manenti, F.; Pozzi, R.; Maschio, G. A perspective on early detection systems models for COVID-19 spreading. Biochem. Biophys. Res. Commun. 2021, 538, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Bhargava, A.; Dahiya, P. COVID-19 Pandemic: Assessment of current strategies and socio-economic Impact. J. Health Manag. 2022, 24, 466–477. [Google Scholar] [CrossRef]
- Li, D.; Wu, M. Pattern recognition receptors in health and diseases. Signal Transduct. Target. Ther. 2021, 6, 291. [Google Scholar] [CrossRef] [PubMed]
- Raskov, H.; Orhan, A.; Salanti, A.; Gaggar, S.; Gögenur, I. Natural killer cells in cancer and cancer immunotherapy. Cancer Lett. 2021, 520, 233–242. [Google Scholar] [CrossRef]
- Furukawa, K.; Tjan, L.H.; Sutandhio, S.; Kurahashi, Y.; Iwata, S.; Tohma, Y.; Sano, S.; Nakamura, S.; Nishimura, M.; Arii, J.; et al. Cross-neutralizing activity against SARS-CoV-2 variants in COVID-19 patients: Comparison of 4 waves of the pandemic in Japan. InOpen Forum Infect. Dis. 2021, 8, 10. [Google Scholar] [CrossRef]
- Sun, L.; Su, Y.; Jiao, A.; Wang, X.; Zhang, B. T cells in health and disease. Signal Transduct. Target. Ther. 2023, 8, 235. [Google Scholar] [CrossRef]
- Cañete, P.F.; Vinuesa, C.G. COVID-19 makes B cells forget, but T cells remember. Cell 2020, 183, 13–15. [Google Scholar] [CrossRef]
- Laidlaw, B.J.; Cyster, J.G. Transcriptional regulation of memory B cell differentiation. Nat. Rev. Immunol. 2021, 21, 209–220. [Google Scholar] [CrossRef]
- Yang, S.; Yu, Y.; Xu, Y.; Jian, F.; Song, W.; Yisimayi, A.; Wang, P.; Wang, J.; Liu, J.; Yu, L.; et al. Fast evolution of SARS-CoV-2 BA. 2· 86 to JN. 1 under heavy immune pressure. Lancet Infect. Dis. 2024, 24, e70–e72. [Google Scholar] [CrossRef]
- Kaku, Y.; Okumura, K.; Padilla-Blanco, M.; Kosugi, Y.; Uriu, K.; Hinay, A.A., Jr.; Chen, L.; Plianchaisuk, A.; Kobiyama, K.; Ishii, K.J. Genotype to phenotype Japan (G2P-Japan) consortium. Virological characteristics of the SARS-CoV-2 JN.1 variant. bioRxiv 2023. [Google Scholar] [CrossRef]
- Yang, L.; Xie, X.; Tu, Z.; Fu, J.; Xu, D.; Zhou, Y. The signal pathways and treatment of cytokine storm in COVID-19. Signal Transduct. Target. Ther. 2021, 6, 255. [Google Scholar] [CrossRef] [PubMed]
- National Institutes of Health. (n.d.). Long COVID. In NIH COVID-19 Research. Available online: https://covid19.nih.gov/covid-19-topics/long-covid (accessed on 1 September 2024).
- Huang, C.; Huang, L.; Wang, Y.; Li, X.; Ren, L.; Gu, X.; Kang, L.; Guo, L.; Liu, M.; Zhou, X.; et al. 6-month consequences of COVID-19 in patients discharged from hospital: A cohort study. Lancet 2021, 397, 220–232. [Google Scholar] [CrossRef] [PubMed]
- Shah, K.; Al-Haidari, A.; Sun, J.; Kazi, J.U. T cell receptor (TCR) signaling in health and disease. Signal Transduct. Target. Ther. 2021, 6, 412. [Google Scholar] [CrossRef] [PubMed]
- Logue, J.K.; Franko, N.M.; McCulloch, D.J.; McDonald, D.; Magedson, A.; Wolf, C.R.; Chu, H.Y. Sequelae in adults at 6 months after COVID-19 infection. JAMA Netw. Open 2021, 4, e210830. [Google Scholar] [CrossRef]
- Raveendran, A.V.; Jayadevan, R.; Sashidharan, S. Long COVID: An overview. Diabetes Metab. Syndr. Clin. Res. Rev. 2021, 15, 869–875. [Google Scholar] [CrossRef]
- Callard, F.; Perego, E. How and why patients made Long Covid. Soc. Sci. Med. 2021, 268, 113426. [Google Scholar] [CrossRef]
- Garg, M.; Maralakunte, M.; Garg, S.; Dhooria, S.; Sehgal, I.; Bhalla, A.S.; Vijayvergiya, R.; Grover, S.; Bhatia, V.; Jagia, P.; et al. The conundrum of ‘long-COVID-19: A narrative review. Int. J. Gen. Med. 2021, 14, 2491–2506. [Google Scholar] [CrossRef]
- Mohandas, S.; Jagannathan, P.; Henrich, T.J.; Sherif, Z.A.; Bime, C.; Quinlan, E.; Portman, M.A.; Gennaro, M.; Rehman, J. Recover Mechanistic Pathways Task Force. Immune mechanisms underlying COVID-19 pathology and post-acute sequelae of SARS-CoV-2 infection (PASC). Elife 2023, 12, e86014. [Google Scholar] [CrossRef]
- Files, J.K.; Boppana, S.; Perez, M.D.; Sarkar, S.; Lowman, K.E.; Qin, K.; Sterrett, S.; Carlin, E.; Bansal, A.; Sabbaj, S.; et al. Sustained cellular immune dysregulation in individuals recovering from SARS-CoV-2 infection. J. Clin. Investig. 2021, 131, e140491. [Google Scholar] [CrossRef]
- Dai, W.; Zhang, B.; Jiang, X.M.; Su, H.; Li, J.; Zhao, Y.; Xie, X.; Jin, Z.; Peng, J.; Liu, F.; et al. Structure-based design of antiviral drug candidates targeting the SARS-CoV-2 main protease. Science 2020, 368, 1331–1335. [Google Scholar] [CrossRef]
- Crook, H.; Raza, S.; Nowell, J.; Young, M.; Edison, P. Long covid-mechanisms, risk factors, and management. BMJ (Clin. Res. Ed.) 2021, 374, n1648. [Google Scholar] [CrossRef] [PubMed]
- Altmann, D.M.; Boyton, R.J. SARS-CoV-2 T cell immunity: Specificity, function, durability, and role in protection. Sci. Immunol. 2021, 6, eabg6347. [Google Scholar] [CrossRef] [PubMed]
- Baig, A.M. Chronic COVID syndrome: Need for an appropriate medical terminology for long-COVID and COVID long-haulers. J. Med. Virol. 2021, 93, 2555–2556. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Leon, S.; Wegman-Ostrosky, T.; Perelman, C.; Sepulveda, R.; Rebolledo, P.A.; Cuapio, A.; Villapol, S. More than 50 Long-term effects of COVID-19: A systematic review and meta-analysis. Sci. Rep. 2021, 11, 1–12. [Google Scholar] [CrossRef]
- Bliddal, S.; Banasik, K.; Pedersen, O.B.; Nissen, J.; Cantwell, L.; Schwinn, M.; Tulstrup, M.; Westergaard, D.; Ullum, H.; Brunak, S.; et al. Acute and persistent symptoms in non-hospitalized PCR-confirmed COVID-19 patients. Sci. Rep. 2021, 11, 19765. [Google Scholar] [CrossRef]
- Nabavi, N. Long COVID: How to define it and how to manage it. Clevel. Clin. J. Med. 2021. [Google Scholar] [CrossRef]
- Dweck, M.R.; Bularga, A.; Hahn, R.T.; Bing, R.; Lee, K.K.; Chapman, A.R.; White, A.; Salvo, G.D.; Sade, L.E.; Pearce, K.; et al. Global evaluation of echocardiography in patients with COVID-19. Eur. Heart J.—Cardiovasc. Imaging 2021, 22, 116–126. [Google Scholar] [CrossRef]
- Dennis, A.; Wamil, M.; Kapur, S.; Alberts, J.; Badley, A.D.; Decker, G.A.; Rizza, S.A.; Banerjee, R.; Banerjee, A. Multi-organ impairment in low-risk individuals with long COVID. bioRxiv 2020. [Google Scholar] [CrossRef]
- Huang, L.; Yao, Q.; Gu, X.; Wang, Q.; Ren, L.; Wang, Y.; Hu, P.; Guo, L.; Liu, M.; Xu, J.; et al. 1-year outcomes in hospital survivors with COVID-19: A longitudinal cohort study. Lancet 2021, 398, 747–758. [Google Scholar] [CrossRef]
- Paces, J.; Strizova, Z.; Daniel, S.M.R.Z.; Cerny, J. COVID-19 and the immune system. Physiol. Res. 2021, 69, 379–388. [Google Scholar] [CrossRef]
- Sudre, C.H.; Murray, B.; Varsavsky, T.; Graham, M.S.; Penfold, R.S.; Bowyer, R.C.; Pujol, J.C.; Klaser, K.; Antonelli, M.; Canas, L.S.; et al. Attributes and predictors of long COVID. Nat. Med. 2021, 27, 626–631. [Google Scholar] [CrossRef] [PubMed]
- Buonsenso, D.; Munblit, D.; De Rose, C.; Sinatti, D.; Ricchiuto, A.; Carfi, A.; Valentini, P. Preliminary evidence on long COVID in children. Acta Paediatr. 2021, 110, 2208. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Guo, R.; Lei, L.; Liu, H.; Wang, Y.; Wang, Y.; Qian, H.; Dai, T.; Zhang, T.; Lai, Y.; et al. COVID-19 infection induces readily detectable morphological and inflammation-related phenotypic changes in peripheral blood monocytes. J. Leukoc. Biol. 2020, 109, 13–22. [Google Scholar] [CrossRef]
- Lancet, T. Facing up to long COVID. Lancet 2020, 396, 1861. [Google Scholar] [CrossRef] [PubMed]
- Alwan, N.A. The road to addressing Long Covid. Science 2021, 373, 491–493. [Google Scholar] [CrossRef]
- Grifoni, A.; Weiskopf, D.; Ramirez, S.I.; Mateus, J.; Dan, J.M.; Moderbacher, C.R.; Rawlings, S.A.; Sutherland, A.; Premkumar, L.; Jadi, R.S.; et al. Targets of T cell responses to SARS-CoV-2 coronavirus in humans with COVID-19 disease and unexposed individuals. Cell 2020, 181, 1489–1501.e15. [Google Scholar] [CrossRef]
- Long, Q.-X.; Tang, X.-J.; Shi, Q.-L.; Li, Q.; Deng, H.-J.; Yuan, J.; Hu, J.-L.; Xu, W.; Zhang, Y.; Lv, F.-J.; et al. Clinical and immunological assessment of asymptomatic SARS-CoV-2 infections. Nat. Med. 2020, 26, 1200–1204. [Google Scholar] [CrossRef]
- Gallais, F.; Velay, A.; Wendling, M.-J.; Nazon, C.; Partisani, M.; Sibilia, J.; Candon, S.; Fafi-Kremer, S. Intrafamilial exposure to SARS-CoV-2 induces cellular immune response without seroconversion. bioRxiv 2020. [Google Scholar] [CrossRef]
- Varnaitė, R.; García, M.; Glans, H.; Maleki, K.T.; Sandberg, J.T.; Tynell, J.; Christ, W.; Lagerqvist, N.; Asgeirsson, H.; Ljunggren, H.G.; et al. Expansion of SARS-CoV-2-specific antibody-secreting cells and generation of neutralizing antibodies in hospitalized COVID-19 patients. J. Immunol. 2021, 206, 2017–2026. [Google Scholar] [CrossRef]
- Perego, E.; Callard, F.; Stras, L.; Melville-Jóhannesson, B.; Pope, R.; Alwan, N.A. Why the patient-made term ‘long Covid’ is needed. Wellcome Open Res. 2020, 5, 224. [Google Scholar] [CrossRef]
- Halpin, S.; O’Connor, R.; Sivan, M. Long COVID and chronic COVID syndromes. J. Med. Virol. 2021, 93, 1242–1243. [Google Scholar] [CrossRef] [PubMed]
- Wang, E.Y.; Mao, T.; Klein, J.; Dai, Y.; Huck, J.D.; Jaycox, J.R.; Liu, F.; Zhou, T.; Israelow, B.; Wong, P.; et al. Diverse functional autoantibodies in patients with COVID-19. Nature 2021, 595, 283–288. [Google Scholar] [CrossRef] [PubMed]
- Venkatesan, P. NICE guideline on long COVID. Lancet Respir. Med. 2021, 9, 129. [Google Scholar] [CrossRef] [PubMed]
- Avendaño-Solá, C.; Ramos-Martínez, A.; Muñez-Rubio, E.; Ruiz-Antorán, B.; Malo de Molina, R.; Torres, F.; Fernández-Cruz, A.; Calderón-Parra, J.; Payares-Herrera, C.; Díaz de Santiago, A.; et al. ConPlas-19 Study Group A multicenter randomized open-label clinical trial for convalescent plasma in patients hospitalized with COVID-19 pneumonia. J. Clin. Investig. 2021, 131, e152740. [Google Scholar] [CrossRef]
- Xia, H.; Cao, Z.; Xie, X.; Zhang, X.; Chen, J.Y.; Wang, H.; Menachery, V.D.; Rajsbaum, R.; Shi, P.Y. Evasion of Type I Interferon by SARS-CoV-2. Cell Rep. 2020, 33, 108234. [Google Scholar] [CrossRef]
- Gaebler, C.; Wang, Z.; Lorenzi, J.C.C.; Muecksch, F.; Finkin, S.; Tokuyama, M.; Cho, A.; Jankovic, M.; Schaefer-Babajew, D.; Oliveira, T.Y.; et al. Evolution of antibody immunity to SARS-CoV-2. Nature 2021, 591, 639–644. [Google Scholar] [CrossRef]
- Pellegrini, D.; Kawakami, R.; Guagliumi, G.; Sakamoto, A.; Kawai, K.; Gianatti, A.; Nasr, A.; Kutys, R.; Guo, L.; Cornelissen, A.; et al. Microthrombi as a Major Cause of Cardiac Injury in COVID-19: A Pathologic Study. Circulation 2021, 143, 1031–1042. [Google Scholar] [CrossRef]
- Michelen, M.; Manoharan, L.; Elkheir, N.; Cheng, V.; Dagens, A.; Hastie, C.; O’Hara, M.; Suett, J.; Dahmash, D.; Bugaeva, P.; et al. Characterising long COVID: A living systematic review. BMJ Glob. Health 2021, 6, e005427. [Google Scholar] [CrossRef]
- Faraj, S.S.; Jalal, P.J. IL1β, IL-6, and TNF-α cytokines cooperate to modulate a complicated medical condition among COVID-19 patients: Case-control study. Ann. Med. Surg. 2023, 85, 2291–2297. [Google Scholar] [CrossRef]
- Sun, Y.; Zou, Y.; Wang, H.; Cui, G.; Yu, Z.; Ren, Z. Immune response induced by novel coronavirus infection. Front. Cell. Infect. Microbiol. 2022, 12, 988604. [Google Scholar] [CrossRef]
- Zuo, J.; Dowell, A.C.; Pearce, H.; Verma, K.; Long, H.M.; Begum, J.; Aiano, F.; Amin-Chowdhury, Z.; Hoschler, K.; Brooks, T.; et al. Robust SARS-CoV-2-specific T cell immunity is maintained at 6 months following primary infection. Nat. Immunol. 2021, 22, 620–626. [Google Scholar] [CrossRef] [PubMed]
- Choi, B.; Choudhary, M.C.; Regan, J.; Sparks, J.A.; Padera, R.F.; Qiu, X.; Solomon, I.H.; Kuo, H.H.; Boucau, J.; Bowman, K.; et al. Persistence and Evolution of SARS-CoV-2 in an Immunocompromised Host. N. Engl. J. Med. 2020, 383, 2291–2293. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Leon, S.; Wegman-Ostrosky, T.; Ayuzo Del Valle, N.C.; Perelman, C.; Sepulveda, R.; Rebolledo, P.A.; Cuapio, A.; Villapol, S. Long-COVID in children and adolescents: A systematic review and meta-analyses. Sci. Rep. 2022, 12, 9950. [Google Scholar] [CrossRef] [PubMed]
- Rodda, L.B.; Netland, J.; Shehata, L.; Pruner, K.B.; Morawski, P.A.; Thouvenel, C.D.; Takehara, K.K.; Eggenberger, J.; Hemann, E.A.; Waterman, H.R.; et al. Functional SARS-CoV-2-Specific Immune Memory Persists after Mild COVID-19. Cell 2021, 184, 169–183.e17. [Google Scholar] [CrossRef]
- Goel, R.R.; Apostolidis, S.A.; Painter, M.M.; Mathew, D.; Pattekar, A.; Kuthuru, O.; Gouma, S.; Hicks, P.; Meng, W.; Rosenfeld, A.M.; et al. Distinct antibody and memory B cell responses in SARS-CoV-2 naïve and recovered individuals following mRNA vaccination. Sci. Immunol. 2021, 6, eabi6950. [Google Scholar] [CrossRef]
- Yong, S.J. Long COVID or post-COVID-19 syndrome: Putative pathophysiology, risk factors, and treatments. Infect. Dis. 2021, 53, 737–754. [Google Scholar] [CrossRef]
- Mateus, J.; Grifoni, A.; Tarke, A.; Sidney, J.; Ramirez, S.I.; Dan, J.M.; Burger, Z.C.; Rawlings, S.A.; Smith, D.M.; Phillips, E.; et al. Selective and cross-reactive SARS-CoV-2 T cell epitopes in unexposed humans. Science 2020, 370, 89–94. [Google Scholar] [CrossRef]
- Thompson, M.G.; Burgess, J.L.; Naleway, A.L.; Tyner, H.; Yoon, S.K.; Meece, J.; Olsho, L.E.W.; Caban-Martinez, A.J.; Fowlkes, A.L.; Lutrick, K.; et al. Prevention and Attenuation of COVID-19 with the BNT162b2 and mRNA-1273 Vaccines. N. Engl. J. Med. 2021, 385, 320–329. [Google Scholar] [CrossRef]
- Asadi-Pooya, A.A.; Nemati, H.; Shahisavandi, M.; Akbari, A.; Emami, A.; Lotfi, M.; Rostamihosseinkhani, M.; Barzegar, Z.; Kabiri, M.; Zeraatpisheh, Z.; et al. Long COVID in children and adolescents. World J. Pediatr. 2021, 17, 495–499. [Google Scholar] [CrossRef]
- Feng, Z.; Diao, B.; Wang, R.; Wang, G.; Wang, C.; Tan, Y.; Liu, L.; Wang, C.; Liu, Y.; Liu, Y.; et al. The novel severe acute respiratory syndrome Coronavirus 2 (SARS-CoV-2) directly decimates human spleens and lymph nodes. medRxiv 2020, 2020-03. [Google Scholar] [CrossRef]
- Mostafa, M.; Barhoum, A.; Sehit, E.; Gewaid, H.; Mostafa, E.; Omran, M.M.; Abdalla, M.S.; Abdel-Haleem, F.M.; Altintas, Z.; Forster, R.J. Current trends in COVID-19 diagnosis and its new variants in physiological fluids: Surface antigens, antibodies, nucleic acids, and RNA sequencing. Trends Anal. Chem. 2022, 157, 116750. [Google Scholar] [CrossRef]
- Rogers, T.F.; Zhao, F.; Huang, D.; Beutler, N.; Burns, A.; He, W.T.; Limbo, O.; Smith, C.; Song, G.; Woehl, J.; et al. Isolation of potent SARS-CoV-2 neutralizing antibodies and protection from disease in a small animal model. Science 2020, 369, 956–963. [Google Scholar] [CrossRef] [PubMed]
- Fact Sheet for Healthcare Providers: Emergency Use Authorization for EvusheldTM (Tixagevimab Co-Packaged with Cilgavimab) Highlights of Emergency Use Authorization (EUA) These Highlights of the EUA Do not Include All the Information Needed to Use EvusheldTM under the EUA. See the Full Fact Sheet for Healthcare Providers for Evusheld. (n.d.). Available online: https://www.fda.gov/media/154701/ (accessed on 22 May 2023).
- Liu, S.; Galat, V.; Galat, Y.; Lee, Y.K.A.; Wainwright, D.; Wu, J. NK cell-based cancer immunotherapy: From basic biology to clinical development. J. Hematol. Oncol. 2021, 14, 7. [Google Scholar] [CrossRef] [PubMed]
- Introna, M.; Franceschetti, M.; Ciocca, A.; Borleri, G.; Conti, E.; Golay, J.; Rambaldi, A. Rapid and massive expansion of cord blood-derived cytokine-induced killer cells: An innovative proposal for the treatment of leukemia relapse after cord blood transplantation. Bone Marrow Transplant. 2006, 38, 621–627. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Zeng, S.; Gong, Z.; Yan, Y. Exosome-based immunotherapy: A promising approach for cancer treatment. Mol. Cancer 2020, 19, 160. [Google Scholar] [CrossRef]
- Morse, M.A.; Nair, S.K.; Mosca, P.J.; Hobeika, A.C.; Clay, T.M.; Deng, Y.; Boczkowski, D.; Proia, A.; Neidzwiecki, D.; Clavien, P.A.; et al. Immunotherapy with autologous, human dendritic cells transfected with carcinoembryonic antigen mRNA. Cancer Investig. 2003, 21, 341–349. [Google Scholar] [CrossRef]
- RECOVERY Collaborative Group; Horby, P.; Lim, W.S.; Emberson, J.R.; Mafham, M.; Bell, J.L.; Linsell, L.; Staplin, N.; Brightling, C.; Ustianowski, A.; et al. Dexamethasone in Hospitalized Patients with COVID-19. N. Engl. J. Med. 2021, 384, 693–704. [Google Scholar] [CrossRef]
- Salama, C.; Han, J.; Yau, L.; Reiss, W.G.; Kramer, B.; Neidhart, J.D.; Criner, G.J.; Kaplan-Lewis, E.; Baden, R.; Pandit, L.; et al. Tocilizumab in patients hospitalized with COVID-19 pneumonia. N. Engl. J. Med. 2021, 384, 20–30. [Google Scholar] [CrossRef]
- Liu, Y.; Sawalha, A.H.; Lu, Q. COVID-19 and autoimmune diseases. Curr. Opin. Rheumatol. 2021, 33, 155–162. [Google Scholar] [CrossRef]
- Tan, C.K.; Wong, Y.J.; Wang, L.M.; Ang, T.L.; Kumar, R. Autoimmune hepatitis following COVID-19 vaccination: True causality or mere association? J. Hepatol. 2021, 75, 1250–1252. [Google Scholar] [CrossRef]
- Zuo, Y.; Yalavarthi, S.; Shi, H.; Gockman, K.; Zuo, M.; Madison, J.A.; Blair, C.; Weber, A.; Barnes, B.J.; Egeblad, M.; et al. Neutrophil extracellular traps in COVID-19. JCI Insight 2020, 5, e138999. [Google Scholar] [CrossRef] [PubMed]
- Zuo, Y.; Estes, S.K.; Ali, R.A.; Gandhi, A.A.; Yalavarthi, S.; Shi, H.; Sule, G.; Gockman, K.; Madison, J.A.; Zuo, M.; et al. Prothrombotic autoantibodies in serum from patients hospitalized with COVID-19. Sci. Transl. Med. 2020, 12, eabd3876. [Google Scholar] [CrossRef] [PubMed]
- Abu-Rumeileh, S.; Abdelhak, A.; Foschi, M.; Tumani, H.; Otto, M. Guillain-Barré syndrome spectrum associated with COVID-19: An up-to-date systematic review of 73 cases. J. Neurol. 2021, 268, 1133–1170. [Google Scholar] [CrossRef] [PubMed]
- Marjot, T.; Buescher, G.; Sebode, M.; Barnes, E.; Barritt, A.S.; Armstrong, M.J., 4th; Baldelli, L.; Kennedy, J.; Mercer, C.; Ozga, A.-K.; et al. SARS-CoV-2 infection in patients with autoimmune hepatitis. J. Hepatol. 2021, 74, 1335–1343. [Google Scholar] [CrossRef]
- Woodruff, M.C.; Ramonell, R.P.; Nguyen, D.C.; Cashman, K.S.; Saini, A.S.; Haddad, N.S.; Ley, A.M.; Kyu, S.; Howell, J.C.; Ozturk, T.; et al. Extrafollicular B cell responses correlate with neutralizing antibodies and morbidity in COVID-19. Nat. Immunol. 2020, 21, 1506–1516. [Google Scholar] [CrossRef]
- Bastard, P.; Rosen, L.B.; Zhang, Q.; Michailidis, E.; Hoffmann, H.H.; Zhang, Y.; Dorgham, K.; Philippot, Q.; Rosain, J.; Béziat, V.; et al. Autoantibodies against type I IFNs in patients with life-threatening COVID-19. Science 2020, 370, eabd4585. [Google Scholar] [CrossRef]
- McMahon, D.E.; Amerson, E.; Rosenbach, M.; Lipoff, J.B.; Moustafa, D.; Tyagi, A.; Desai, S.R.; French, L.E.; Lim, H.W.; Thiers, B.H.; et al. Cutaneous reactions reported after Moderna and Pfizer COVID-19 vaccination: A registry-based study of 414 cases. J. Am. Acad. Dermatol. 2021, 85, 46–55. [Google Scholar] [CrossRef]
- Cunningham, M.W. Molecular Mimicry, Autoimmunity, and Infection: The Cross-Reactive Antigens of Group A Streptococci and their Sequelae. Microbiol. Spectr. 2019, 7, 10-1128. [Google Scholar] [CrossRef]
- Del Valle, D.M.; Kim-Schulze, S.; Huang, H.H.; Beckmann, N.D.; Nirenberg, S.; Wang, B.; Lavin, Y.; Swartz, T.H.; Madduri, D.; Stock, A.; et al. An inflammatory cytokine signature predicts COVID-19 severity and survival. Nat. Med. 2020, 26, 1636–1643. [Google Scholar] [CrossRef]
- Dotan, A.; Muller, S.; Kanduc, D.; David, P.; Halpert, G.; Shoenfeld, Y. The SARS-CoV-2 as an instrumental trigger of autoimmunity. Autoimmun. Rev. 2021, 20, 102792. [Google Scholar] [CrossRef]
- Morad, G.; Helmink, B.A.; Sharma, P.; Wargo, J.A. Hallmarks of response, resistance, and toxicity to immune checkpoint blockade. Cell 2021, 184, 5309–5337. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Han, T.; Chen, J.; Hou, C.; Hua, L.; He, S.; Guo, Y.; Zhang, S.; Wang, Y.; Yuan, J.; et al. Clinical and Autoimmune Characteristics of Severe and Critical Cases of COVID-19. Clin. Transl. Sci. 2020, 13, 1077–1086. [Google Scholar] [CrossRef] [PubMed]
- Nalbandian, A.; Sehgal, K.; Gupta, A.; Madhavan, M.V.; McGroder, C.; Stevens, J.S.; Cook, J.R.; Nordvig, A.S.; Shalev, D.; Sehrawat, T.S.; et al. Post-acute COVID-19 syndrome. Nat. Med. 2021, 27, 601–615. [Google Scholar] [CrossRef] [PubMed]
- Groff, D.; Sun, A.; Ssentongo, A.E.; Ba, D.M.; Parsons, N.; Poudel, G.R.; Lekoubou, A.; Oh, J.S.; Ericson, J.E.; Ssentongo, P.; et al. Short-term and Long-term Rates of Postacute Sequelae of SARS-CoV-2 Infection: A Systematic Review. JAMA Netw. Open 2021, 4, e2128568. [Google Scholar] [CrossRef]
- WHO Rapid Evidence Appraisal for COVID-19 Therapies (REACT) Working Group; Sterne, J.A.C.; Murthy, S.; Diaz, J.V.; Slutsky, A.S.; Villar, J.; Angus, D.C.; Annane, D.; Azevedo, L.C.P.; Berwanger, O.; et al. Association Between Administration of Systemic Corticosteroids and Mortality among Critically Ill Patients With COVID-19: A Meta-analysis. JAMA 2020, 324, 1330–1341. [Google Scholar] [CrossRef]
- Bartone, P.T.; McDonald, K.; Hansma, B.J.; Solomon, J. Hardiness moderates the effects of COVID-19 stress on anxiety and depression. J. Affect. Disord. 2022, 317, 236–244. [Google Scholar] [CrossRef]
- Polastri, M.; Nava, S.; Clini, E.; Vitacca, M.; Gosselink, R. COVID-19 and pulmonary rehabilitation: Preparing for phase three. Eur. Respir. J. 2020, 55, 2001822. [Google Scholar] [CrossRef]
- Spruit, M.A.; Singh, S.J.; Garvey, C.; ZuWallack, R.; Nici, L.; Rochester, C.; Hill, K.; Holland, A.E.; Lareau, S.C.; Man, W.D.; et al. ATS/ERS Task Force on Pulmonary Rehabilitation An official American Thoracic Society/European Respiratory Society statement: Key concepts and advances in pulmonary rehabilitation. Am. J. Respir. Crit. Care Med. 2013, 188, e13–e64. [Google Scholar] [CrossRef]
- Holland, A.E.; Spruit, M.A.; Troosters, T.; Puhan, M.A.; Pepin, V.; Saey, D.; McCormack, M.C.; Carlin, B.W.; Sciurba, F.C.; Pitta, F.; et al. An official European Respiratory Society/American Thoracic Society technical standard: Field walking tests in chronic respiratory disease. Eur. Respir. J. 2014, 44, 1428–1446. [Google Scholar] [CrossRef]
- Larson, J.L.; Covey, M.K.; Wirtz, S.E.; Berry, J.K.; Alex, C.G.; Langbein, W.E.; Edwards, L. Cycle ergometer and inspiratory muscle training in chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 1999, 160, 500–507. [Google Scholar] [CrossRef]
- Nici, L.; Donner, C.; Wouters, E.; Zuwallack, R.; Ambrosino, N.; Bourbeau, J.; Carone, M.; Celli, B.; Engelen, M.; Fahy, B.; et al. ATS/ERS Pulmonary Rehabilitation Writing Committee American Thoracic Society/European Respiratory Society statement on pulmonary rehabilitation. Am. J. Respir. Crit. Care Med. 2006, 173, 1390–1413. [Google Scholar] [CrossRef]
- Mazza, M.G.; De Lorenzo, R.; Conte, C.; Poletti, S.; Vai, B.; Bollettini, I.; Melloni, E.M.T.; Furlan, R.; Ciceri, F.; Rovere-Querini, P.; et al. Anxiety and depression in COVID-19 survivors: Role of inflammatory and clinical predictors. Brain Behav. Immun. 2020, 89, 594–600. [Google Scholar] [CrossRef] [PubMed]
- Rogers, J.P.; Chesney, E.; Oliver, D.; Pollak, T.A.; McGuire, P.; Fusar-Poli, P.; Zandi, M.S.; Lewis, G.; David, A.S. Psychiatric and neuropsychiatric presentations associated with severe coronavirus infections: A systematic review and meta-analysis with comparison to the COVID-19 pandemic. Lancet Psychiatry 2020, 7, 611–627. [Google Scholar] [CrossRef]
- Hampshire, A.; Trender, W.; Chamberlain, S.R.; Jolly, A.E.; Grant, J.E.; Patrick, F.; Mazibuko, N.; Williams, S.C.; Barnby, J.M.; Hellyer, P.; et al. Cognitive deficits in people who have recovered from COVID-19. EClinicalMedicine 2021, 39, 101044. [Google Scholar] [CrossRef]
- Zheng, K.I.; Feng, G.; Liu, W.Y.; Targher, G.; Byrne, C.D.; Zheng, M.H. Extrapulmonary complications of COVID-19: A multisystem disease? J. Med. Virol. 2021, 93, 323–335. [Google Scholar] [CrossRef]
- Barker-Davies, R.M.; O’Sullivan, O.; Senaratne, K.P.P.; Baker, P.; Cranley, M.; Dharm-Datta, S.; Ellis, H.; Goodall, D.; Gough, M.; Lewis, S.; et al. The Stanford Hall consensus statement for post-COVID-19 rehabilitation. Br. J. Sports Med. 2020, 54, 949–959. [Google Scholar] [CrossRef] [PubMed]
- Andrenelli, E.; Negrini, F.; de Sire, A.; Arienti, C.; Patrini, M.; Negrini, S.; Ceravolo, M.G. International Multiprofessional Steering Committee of Cochrane Rehabilitation REH-COVER action Systematic rapid living review on rehabilitation needs due to COVID-19: Update to May 31st, 2020. Eur. J. Phys. Rehabil. Med. 2020, 56, 508–514. [Google Scholar] [CrossRef]
- Lee, M.J.; Sayers, A.E.; Drake, T.M.; Singh, P.; Bradburn, M.; Wilson, T.R.; Murugananthan, A.; Walsh, C.J.; Fearnhead, N.S.; NASBO Steering Group and NASBO Collaborators. Malnutrition, nutritional interventions and clinical outcomes of patients with acute small bowel obstruction: Results from a national, multicentre, prospective audit. BMJ Open 2019, 9, e029235. [Google Scholar] [CrossRef] [PubMed]
- Alam, S.; Bhuiyan, F.R.; Emon, T.H.; Hasan, M. Prospects of nutritional interventions in the care of COVID-19 patients. Heliyon 2021, 7, e06285. [Google Scholar] [CrossRef]
- Calder, P.C.; Carr, A.C.; Gombart, A.F.; Eggersdorfer, M. Optimal Nutritional Status for a Well-Functioning Immune System Is an Important Factor to Protect against Viral Infections. Nutrients 2020, 12, 1181. [Google Scholar] [CrossRef]
- World Health Organization. Water, Sanitation, Hygiene, and Waste Management for SARS-CoV-2, the Virus That Causes COVID-19. Available online: https://www.who.int/publications/i/item/WHO-2019-nCoV-IPC-WASH-2020.4 (accessed on 1 September 2024).
- Shah, K.; Saxena, D.; Mavalankar, D. Secondary attack rate of COVID-19 in household contacts: A systematic review. QJM Mon. J. Assoc. Physicians 2020, 113, 841–850. [Google Scholar] [CrossRef] [PubMed]
- Ceban, F.; Ling, S.; Lui, L.M.W.; Lee, Y.; Gill, H.; Teopiz, K.M.; Rodrigues, N.B.; Subramaniapillai, M.; Di Vincenzo, J.D.; Cao, B.; et al. Fatigue and cognitive impairment in Post-COVID-19 Syndrome: A systematic review and meta-analysis. Brain Behav. Immun. 2022, 101, 93–135. [Google Scholar] [CrossRef] [PubMed]
- Naliboff, B.D.; Wu, S.M.; Schieffer, B.; Bolus, R.; Pham, Q.; Baria, A.; Aragaki, D.; Van Vort, W.; Davis, F.; Shekelle, P. A randomized trial of 2 prescription strategies for opioid treatment of chronic nonmalignant pain. J. Pain 2011, 12, 288–296. [Google Scholar] [CrossRef] [PubMed]
- Katon, W.; Ciechanowski, P. Impact of major depression on chronic medical illness. J. Psychosom. Res. 2002, 53, 859–863. [Google Scholar] [CrossRef]
- Alsubaie, M.; Abbott, R.; Dunn, B.; Dickens, C.; Keil, T.F.; Henley, W.; Kuyken, W. Mechanisms of action in mindfulness-based cognitive therapy (MBCT) and mindfulness-based stress reduction (MBSR) in people with physical and/or psychological conditions: A systematic review. Clin. Psychol. Rev. 2017, 55, 74–91. [Google Scholar] [CrossRef]
- Dennis, C.-L. Peer support within a health care context: A concept analysis. Int. J. Nurs. Stud. 2003, 40, 321–332. [Google Scholar] [CrossRef]
- Carfì, A.; Bernabei, R.; Landi, F.; Gemelli Against COVID-19 Post-Acute Care Study Group. Persistent Symptoms in Patients After Acute COVID-19. JAMA 2020, 324, 603–605. [Google Scholar] [CrossRef]
- Greenhalgh, T.; Jimenez, J.L.; Prather, K.A.; Tufekci, Z.; Fisman, D.; Schooley, R. Ten scientific reasons in support of airborne transmission of SARS-CoV-2. Lancet 2021, 397, 1603–1605. [Google Scholar] [CrossRef]
- Mazza, M.G.; Palladini, M.; De Lorenzo, R.; Magnaghi, C.; Poletti, S.; Furlan, R.; Ciceri, F.; COVID-19 BioB Outpatient Clinic Study Group; Rovere-Querini, P.; Benedetti, F. Persistent psychopathology and neurocognitive impairment in COVID-19 survivors: Effect of inflammatory biomarkers at three-month follow-up. Brain Behav. Immun. 2021, 94, 138–147. [Google Scholar] [CrossRef]
- Menni, C.; Valdes, A.M.; Freidin, M.B.; Sudre, C.H.; Nguyen, L.H.; Drew, D.A.; Ganesh, S.; Varsavsky, T.; Cardoso, M.J.; El-Sayed Moustafa, J.S.; et al. Real-time tracking of self-reported symptoms to predict potential COVID-19. Nat. Med. 2020, 26, 1037–1040. [Google Scholar] [CrossRef]
- Halpin, S.J.; McIvor, C.; Whyatt, G.; Adams, A.; Harvey, O.; McLean, L.; Walshaw, C.; Kemp, S.; Corrado, J.; Singh, R.; et al. Postdischarge symptoms and rehabilitation needs in survivors of COVID-19 infection: A cross-sectional evaluation. J. Med. Virol. 2021, 93, 1013–1022. [Google Scholar] [CrossRef]
- Cajanding, R.J.M. Comprehensive Review of Cardiovascular Involvement in COVID-19. AACN Adv. Crit. Care 2021, 32, 169–187. [Google Scholar] [CrossRef] [PubMed]
- Meckawy, R.; Stuckler, D.; Mehta, A.; Al-Ahdal, T.; Doebbeling, B.N. Effectiveness of early warning systems in the detection of infectious diseases outbreaks: A systematic review. BMC Public Health 2022, 22, 2216. [Google Scholar] [CrossRef] [PubMed]
- Haldane, V.; Jung, A.S.; De Foo, C.; Bonk, M.; Jamieson, M.; Wu, S.; Verma, M.; Abdalla, S.M.; Singh, S.; Nordström, A.; et al. Strengthening the basics: Public health responses to prevent the next pandemic. BMJ 2021, 375, e067510. [Google Scholar] [CrossRef] [PubMed]
- Excler, J.L.; Saville, M.; Berkley, S.; Kim, J.H. Vaccine development for emerging infectious diseases. Nat. Med. 2021, 27, 591–600. [Google Scholar] [CrossRef] [PubMed]
- Varkey, B. Principles of Clinical Ethics and Their Application to Practice. Med. Princ. Pract. Int. J. Kuwait Univ. Health Sci. Cent. 2021, 30, 17–28. [Google Scholar] [CrossRef]
- Dai, H.; Saccardo, S.; Han, M.A.; Roh, L.; Raja, N.; Vangala, S.; Modi, H.; Pandya, S.; Sloyan, M.; Croymans, D.M. Behavioural nudges increase COVID-19 vaccinations. Nature 2021, 597, 404–409. [Google Scholar] [CrossRef]
- Kinder, K.; Bazemore, A.; Taylor, M.; Mannie, C.; Strydom, S.; George, J.; Goodyear-Smith, F. Integrating primary care and public health to enhance response to a pandemic. Prim. Health Care Res. Dev. 2021, 22, e27. [Google Scholar] [CrossRef]
- Mumtaz, H.; Riaz, M.H.; Wajid, H.; Saqib, M.; Zeeshan, M.H.; Khan, S.E.; Chauhan, Y.R.; Sohail, H.; Vohra, L.I. Current challenges and potential solutions to the use of digital health technologies in evidence generation: A narrative review. Front. Digit. Health 2023, 5, 1203945. [Google Scholar] [CrossRef]
- WHO. Launches New Initiative to Improve Pandemic Preparedness. Available online: https://www.who.int/news/item/26-04-2023-who-launches-new-initiative-to-improve-pandemic-preparedness (accessed on 1 September 2024).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).