
The Apicomplexan Parasite Toxoplasma gondii

 ,
,
Definition
:1. Toxoplasma gondii: A Successful Parasite
2. A Prevalent and Silent Infection
2.1. Epidemiology and Transmission
2.2. Infection and Disease
3. An Apicomplexa Parasite
4. Genetic Diversity: One Species, Many Strains
5. One Parasite, Several Specialized Eukaryotic Cells
5.1. Zoites, Motile Stages
5.2. Non-Motile and Gamete Stages
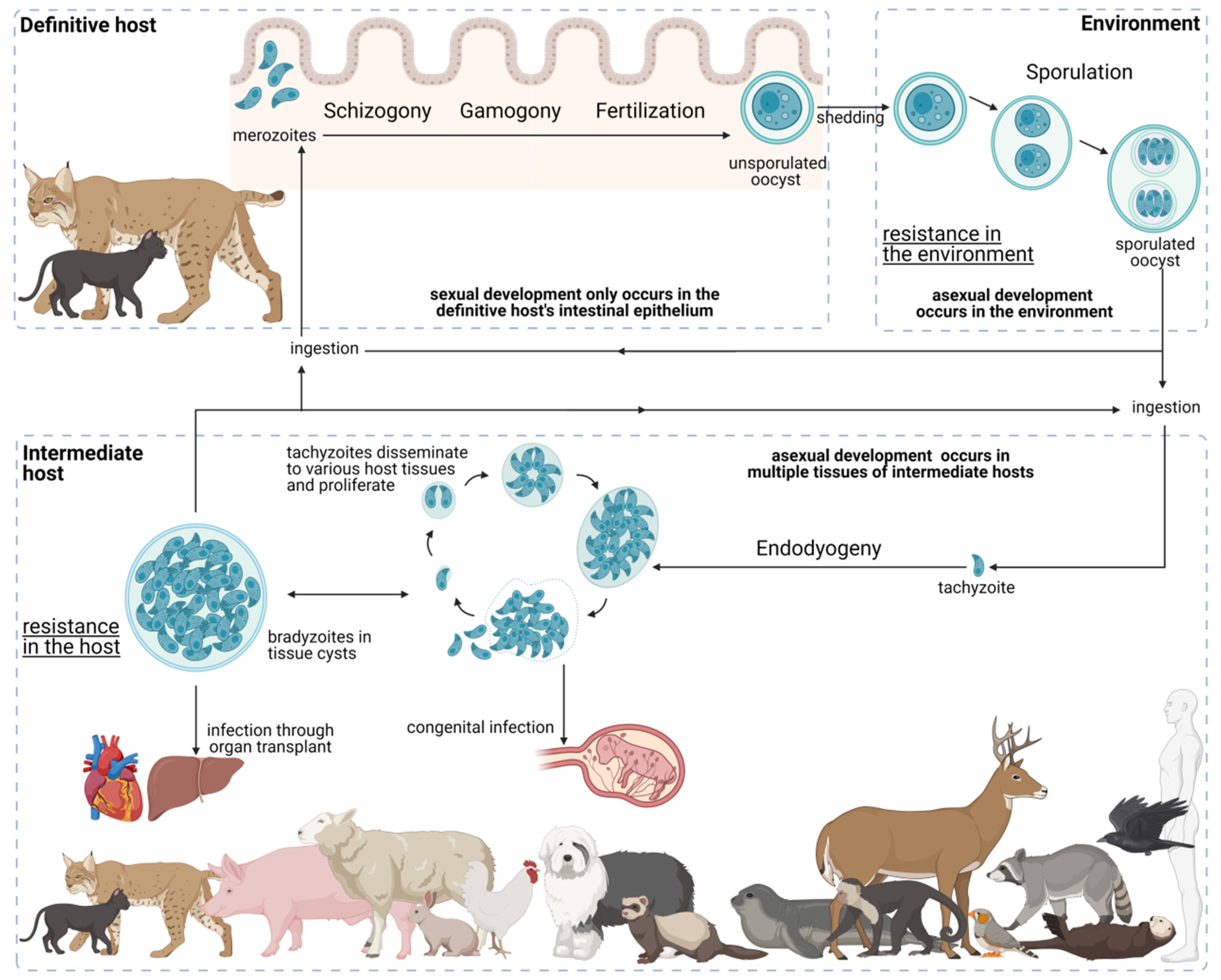
6. The Complex Toxoplasma gondii Life Cycle: A Path to Success
6.1. Development in the Definitive Host
6.2. Environmental Development
6.3. Development in the Intermediate Host
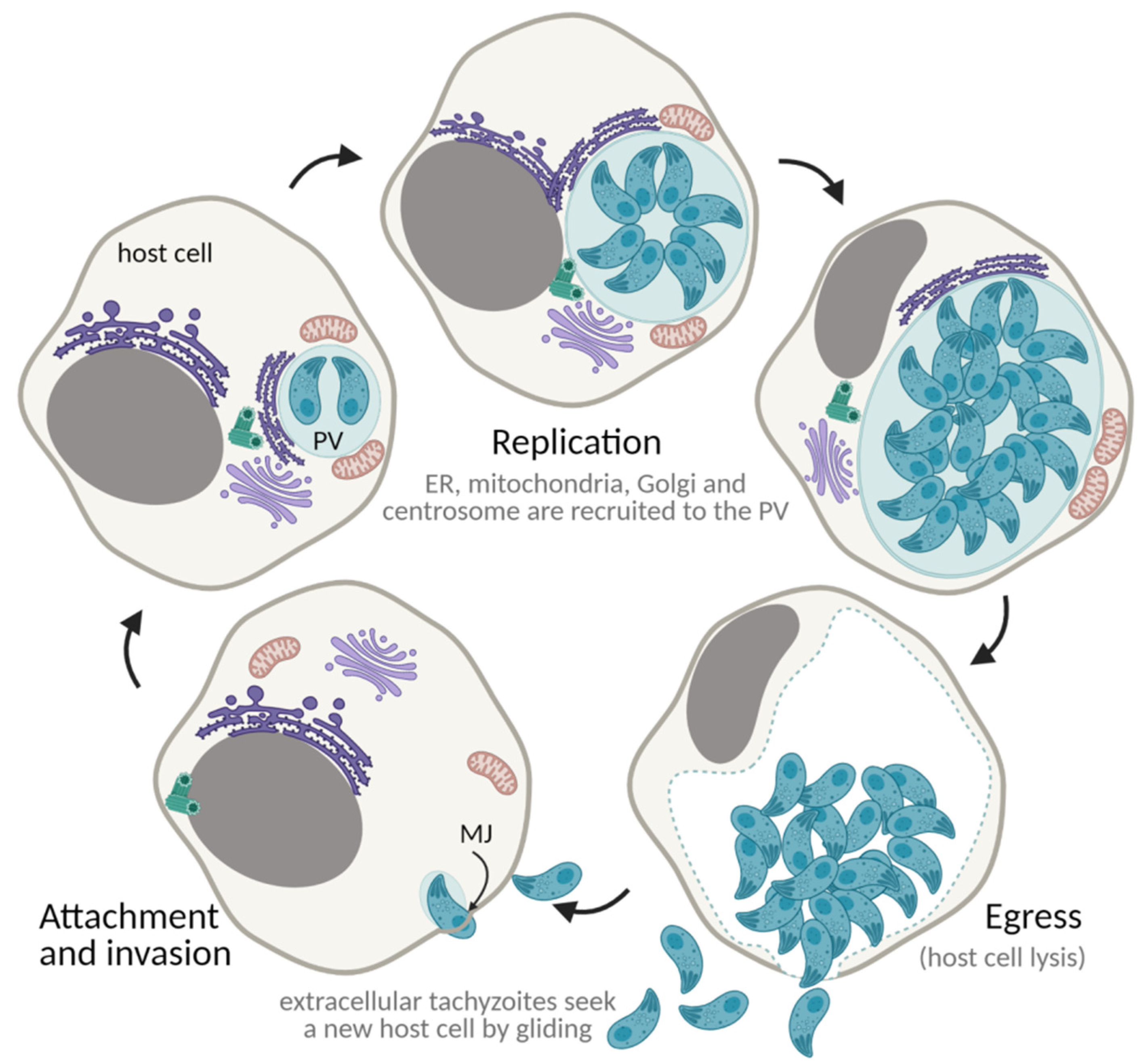
7. The Toxoplasma gondii Lytic Cycle: An Efficient Proliferation Strategy
7.1. Gliding Motility
7.2. Attachment and Invasion
7.3. Establishment of the Parasitophorous Vacuole
7.4. Proliferation through Endodyogeny
7.5. Egress and Repeat
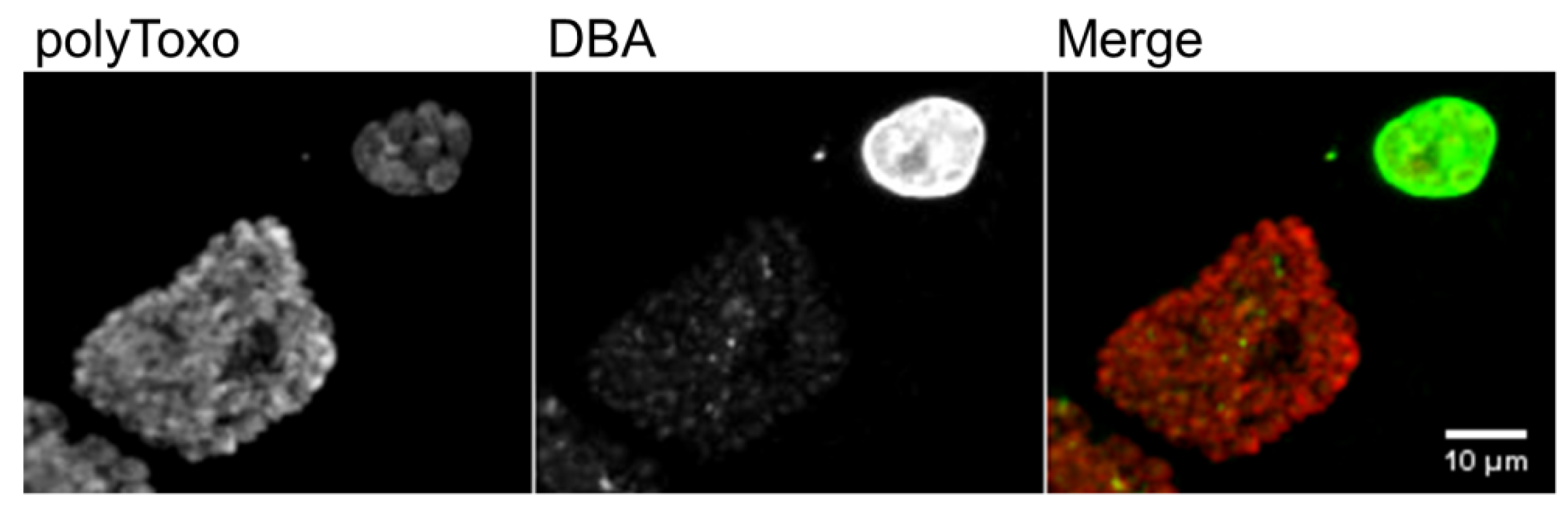
8. The Tissue Cyst
9. Conclusions and Prospects
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Entry Link on the Encyclopedia Platform
References
- Jiménez-Ruiz, E.; Wong, E.H.; Pall, G.S.; Meissner, M. Advantages and disadvantages of conditional systems for characterization of essential genes in Toxoplasma gondii. Parasitology 2014, 141, 1390–1398. [Google Scholar] [CrossRef] [PubMed]
- Nicolle, C.; Manceaux, L. Sur Une Infection à Corps de Leishman (Ou Organismes Voisins) Du Gondi. Comptes Rendus Séances L’académie Sci. 1908, 147, 763–766. [Google Scholar]
- Splendore, A. Un Nuovo Protozoa Parassita de’ Conigli. Rev. Soc. Sci. 1908, 3, 109–112. [Google Scholar]
- Nicolle, C.; Manceaux, L. Sur Un Protozoaire Nouveau Du Gondi. Comptes Rendus Séances L’académie Sci. 1909, 148, 369–372. [Google Scholar]
- McLeod, R.; Van Tubbergen, C.; Montoya, J.G.; Petersen, E. Human Toxoplasma Infection, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2014; ISBN 9780123964816. [Google Scholar]
- Stelzer, S.; Basso, W.; Silván, J.B.; Ortega-Mora, L.-M.; Maksimov, P.; Gethmann, J.; Conraths, F.; Schares, G. Toxoplasma gondii infection and toxoplasmosis in farm animals: Risk factors and economic impact. Food Waterborne Parasitol. 2019, 15, e00037. [Google Scholar] [CrossRef]
- Black, M.W.; Boothroyd, J.C. Lytic Cycle of Toxoplasma Gondii. Microbiol. Mol. Biol. Rev. 2000, 64, 607–623. [Google Scholar] [CrossRef] [PubMed]
- Aguirre, A.A.; Longcore, T.; Barbieri, M.; Dabritz, H.; Hill, D.; Klein, P.N.; Lepczyk, C.; Lilly, E.L. The one health approach to toxoplasmosis: Epidemiology, Control, and Prevention Strategies. Ecohealth 2019, 16, 378–390. [Google Scholar] [CrossRef]
- Wang, J.-L.; Zhang, N.-Z.; Li, T.-T.; He, J.-J.; Elsheikha, H.M.; Zhu, X.-Q. Advances in the development of Anti-Toxoplasma Gondii Vaccines: Challenges, opportunities, and perspectives. Trends Parasitol. 2019, 35, 239–253. [Google Scholar] [CrossRef]
- World Malaria Report 2020: 20 Years of Global Progress and Challenges; World Health Organization: Geneva, Switzerland, 2020; Licence: CC BY-NC-SA 3.0 IGO.
- Gray, J.; Zintl, A.; Hildebrandt, A.; Hunfeld, K.P.; Weiss, L. Zoonotic babesiosis: Overview of the disease and novel aspects of pathogen identity. Ticks Tick Borne Dis. 2010, 1, 3–10. [Google Scholar] [CrossRef]
- Beugnet, F.; Moreau, Y. Babesiosis. OIE Rev. Sci. Tech. 2015, 34, 627–639. [Google Scholar] [CrossRef] [PubMed]
- Tzipori, S.; Ward, H. Cryptosporidiosis: Biology, pathogenesis and disease. Microbes Infect. 2002, 4, 1047–1058. [Google Scholar] [CrossRef]
- Reichel, M.P.; Alejandra Ayanegui-Alcérreca, M.; Gondim, L.F.P.; Ellis, J.T. What is the global economic impact of neospora caninum in cattle-The Billion Dollar Question. Int. J. Parasitol. 2013, 43, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-García, G.; García-Lunar, P.; Gutiérrez-Expósito, D.; Shkap, V.; Ortega-Mora, L.M. Dynamics of Besnoitia Besnoiti infection in cattle. Parasitology 2014, 141, 1419–1435. [Google Scholar] [CrossRef] [PubMed]
- Robert-Gangneux, F.; Dardé, M.L. Epidemiology of and diagnostic strategies for toxoplasmosis. Clin. Microbiol. Rev. 2012, 25, 264–296. [Google Scholar] [CrossRef]
- Parise, M.E.; Hotez, P.J.; Slutsker, L. Neglected parasitic infections in the United States: Needs and opportunities. Am. J. Trop. Med. Hyg. 2014, 90, 783–785. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention and Control. Congenital toxoplasmosis. In ECDC. Annual Epidemiological Report for 2018; ECDC: Stockholm, Sweden, 2021. [Google Scholar]
- Flegr, J.; Prandota, J.; Sovičková, M.; Israili, Z.H. Toxoplasmosis-A global threat. Correlation of latent toxoplasmosis with specific disease burden in a set of 88 countries. PLoS ONE 2014, 9, e90203. [Google Scholar] [CrossRef]
- Smith, D.D.; Frenkel, J.K. Prevalence of antibodies to toxoplasma gondii in wild mammals of missouri and east central kansas: Biologic and ecologic considerations of transmission. J. Wildl. Dis. 1995, 31, 15–21. [Google Scholar] [CrossRef]
- De Thoisy, B.; Demar, M.; Aznar, C.; Carme, B. Ecological Correlates of Toxoplasma Gondii Exposure in Free-Ranging Neotropical Mammals. J. Wildl. Dis. 2003, 39, 456–459. [Google Scholar] [CrossRef]
- Dabritz, H.A.; Miller, M.A.; Gardner, I.A.; Packham, A.E.; Atwill, E.R.; Conrad, P.A. Risk factors for Toxoplasma Gondii Infection in Wild Rodents from Central Coastal California and a review of T. Gondii Prevalence in Rodents. J. Parasitol. 2008, 94, 675–683. [Google Scholar] [CrossRef]
- Barros, M.; Cabezón, O.; Dubey, J.P.; Almería, S.; Ribas, M.P.; Escobar, L.E.; Ramos, B.; Medina-Vogel, G. Toxoplasma Gondii Infection in Wild Mustelids and Cats across an Urban-Rural Gradient. PLoS ONE 2018, 13, e0199085. [Google Scholar] [CrossRef]
- Dubey, J.P.; Murata, F.H.A.; Cerqueira-Cézar, C.K.; Kwok, O.C.H. Public health and economic importance of Toxoplasma Gondii Infections in Goats: The Last Decade. Res. Vet. Sci. 2020, 132, 292–307. [Google Scholar] [CrossRef]
- Dubey, J.P.; Murata, F.H.A.; Cerqueira-Cézar, C.K.; Kwok, O.C.H. Toxoplasma Gondii Infections in Horses, Donkeys, and Other Equids: The Last Decade. Res. Vet. Sci. 2020, 132, 492–499. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P.; Murata, F.H.A.; Cerqueira-Cézar, C.K.; Kwok, O.C.H.; Yang, Y.R. Public Health Significance of Toxoplasma Gondii Infections in Cattle: 2009–2020. J. Parasitol. 2020, 106, 772–788. [Google Scholar] [CrossRef] [PubMed]
- Montazeri, M.; Mikaeili Galeh, T.; Moosazadeh, M.; Sarvi, S.; Dodangeh, S.; Javidnia, J.; Sharif, M.; Daryani, A. The Global Serological Prevalence of Toxoplasma Gondii in Felids during the Last Five Decades (1967–2017): A Systematic Review and Meta-Analysis. Parasites Vectors 2020, 13, 82. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, K.; Bahia-Oliveira, L.; Dixon, B.; Dumètre, A.; de Wit, L.A.; VanWormer, E.; Villena, I. Environmental Transmission of Toxoplasma Gondii: Oocysts in Water, Soil and Food. Food Waterborne Parasitol. 2019, 15, e00049. [Google Scholar] [CrossRef] [PubMed]
- Maleki, B.; Ahmadi, N.; Olfatifar, M.; Gorgipour, M.; Taghipour, A.; Abdoli, A.; Khorshidi, A.; Foroutan, M.; Mirzapour, A. Toxoplasma Oocysts in the Soil of Public Places Worldwide: A Systematic Review and Meta-Analysis. Trans. R. Soc. Trop. Med. Hyg. 2021, 115, 471–481. [Google Scholar] [CrossRef] [PubMed]
- Moré, G.; Maksimov, P.; Pardini, L.; Herrmann, D.C.; Bacigalupe, D.; Maksimov, A.; Basso, W.; Conraths, F.J.; Schares, G.; Venturini, M.C. Toxoplasma Gondii Infection in Sentinel and Free-Range Chickens from Argentina. Vet. Parasitol. 2012, 184, 116–121. [Google Scholar] [CrossRef]
- Guo, M.; Dubey, J.P.; Hill, D.; Buchanan, R.L.; Ray Gamble, H.; Jones, J.L.; Pradhan, A.K. Prevalence and Risk Factors for Toxoplasma Gondii Infection in Meat Animals and Meat Products Destined for Human Consumption. J. Food Prot. 2015, 78, 457–476. [Google Scholar] [CrossRef]
- Dubey, J.P.; Frenkel, J.K. Immunity to Feline Toxoplasmosis: Modification by Administration of Corticosteroids. Vet. Pathol. 1974, 11, 350–379. [Google Scholar] [CrossRef]
- Kapperud, G.; Jenum, P.A.; Stray-Pedersen, B.; Melby, K.K.; Eskild, A.; Eng, J. Risk Factors for Toxoplasma Gondii Infection in Pregnancy: Results of a Prospective Case-Control Study in Norway. Am. J. Epidemiol. 1996, 144, 405–412. [Google Scholar] [CrossRef]
- Cook, A.J.; Gilbert, R.E.; Buffolano, W.; Zufferey, J.; Petersen, E.; Jenum, P.A.; Foulon, W.; Semprini, A.E.; Dunn, D.T. Sources of Toxoplasma Infection in Pregnant Women: European Multicentre Case-Control Study. European Research Network on Congenital Toxoplasmosis. BMJ 2000, 321, 142–147. [Google Scholar] [CrossRef]
- Berger, F.; Goulet, V.; Le Strat, Y.; Desenclos, J.C. Toxoplasmosis among Pregnant Women in France: Risk Factors and Change of Prevalence between 1995 and 2003. Rev. Epidemiol. Sante Publique 2009, 57, 241–248. [Google Scholar] [CrossRef]
- Jones, J.L.; Dargelas, V.; Roberts, J.; Press, C.; Remington, J.S.; Montoya, J.G. Risk Factors for Toxoplasma Gondii Infection in the United States. Clin. Infect. Dis. 2009, 49, 878–884. [Google Scholar] [CrossRef]
- Ramakrishnan, C.; Maier, S.; Walker, R.A.; Rehrauer, H.; Joekel, D.E.; Winiger, R.R.; Basso, W.U.; Grigg, M.E.; Hehl, A.B.; Deplazes, P.; et al. An Experimental Genetically Attenuated Live Vaccine to Prevent Transmission of Toxoplasma Gondii by Cats. Sci. Rep. 2019, 9, 1474. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P. Advances in the Life Cycle of Toxoplasma Gondii. Int. J. Parasitol. 1998, 28, 1019–1024. [Google Scholar] [CrossRef]
- Kean, B.H.; Kimball, A.C.; Christenson, W.N. An Epidemic of Acute Toxoplasmosis. JAMA 1969, 208, 1002–1004. [Google Scholar] [CrossRef] [PubMed]
- Desmonts, G.; Couvreur, J.; Alison, F.; Baudelot, J.; Gerbeaux, J.; Lelong, M. Étude Épidémiologique Sur La Toxoplasmose: De l’influence de La Cuisson Des Viandes de Boucherie Sur La Fréquence de l’infection Humaine. Rev. Fr. Études Clin. Biol. 1965, 10, 952–958. [Google Scholar]
- Hill, D.; Coss, C.; Dubey, J.P.; Wroblewski, K.; Sautter, M.; Hosten, T.; Muñoz-Zanzi, C.; Mui, E.; Withers, S.; Boyer, K.; et al. Identification of a Sporozoite-Specific Antigen from Toxoplasma Gondii. J. Parasitol. 2011, 97, 328–337. [Google Scholar] [CrossRef]
- Dubey, J.P. Toxoplasmosis in Sheep-The Last 20 Years. Vet. Parasitol. 2009, 163, 1–14. [Google Scholar] [CrossRef]
- Ryning, F.W.; McLeod, R.; Maddox, J.C.; Hunt, S.; Remington, J.S. Probable Transmission of Toxoplasma Gondii by Organ Transplantation. Ann. Intern. Med. 1979, 90, 47–49. [Google Scholar] [CrossRef] [PubMed]
- Sacks, J.J.; Roberto, R.R.; Brooks, N.F. Toxoplasmosis Infection Associated With Raw Goat’s Milk. JAMA J. Am. Med. Assoc. 1982, 248, 1728–1732. [Google Scholar] [CrossRef]
- Fernàndez-Sabé, N.; Cervera, C.; Fariñas, M.C.; Bodro, M.; Muñoz, P.; Gurguí, M.; Torre-Cisneros, J.; Martín-Dávila, P.; Noblejas, A.; Len, Ó.; et al. Risk Factors, Clinical Features, and Outcomes of Toxoplasmosis in Solid-Organ Transplant Recipients: A Matched Case-Control Study. Clin. Infect. Dis. 2012, 54, 355–361. [Google Scholar] [CrossRef]
- Dubey, J.P.; Verma, S.K.; Ferreira, L.R.; Oliveira, S.; Cassinelli, A.B.; Ying, Y.; Kwok, O.C.H.; Tuo, W.; Chiesa, O.A.; Jones, J.L. Detection and Survival of Toxoplasma Gondii in Milk and Cheese from Experimentally Infected Goats. J. Food Prot. 2014, 77, 1747–1753. [Google Scholar] [CrossRef]
- Milne, G.; Webster, J.P.; Walker, M. Toxoplasma Gondii: An Underestimated Threat? Trends Parasitol. 2020, 36, 959–969. [Google Scholar] [CrossRef]
- Montoya, J.G.; Liesenfeld, O. Toxoplasmosis. Lancet 2004, 363, 1965–1976. [Google Scholar] [CrossRef]
- Calero-Berna, R.; Gennari, S.M. Clinical Toxoplasmosis in Dogs and Cats: An Update. Front. Vet. Sci. 2019, 6, 54. [Google Scholar] [CrossRef] [PubMed]
- Dunay, I.R.; Gajurel, K.; Dhakal, R.; Liesenfeld, O.; Montoya, J.G. Treatment of Toxoplasmosis: Historical Perspective, Animal. Clin. Microbiol. Infect. 2018, 31, 1–33. [Google Scholar]
- McPhillie, M.J.; Zhou, Y.; Hickman, M.R.; Gordon, J.A.; Weber, C.R.; Li, Q.; Lee, P.J.; Amporndanai, K.; Johnson, R.M.; Darby, H.; et al. Potent Tetrahydroquinolone Eliminates Apicomplexan Parasites. Front. Cell. Infect. Microbiol. 2020, 10, 1–27. [Google Scholar] [CrossRef]
- Brandão, G.P.; Melo, M.N.; Caetano, B.C.; Carneiro, C.M.; Silva, L.A.; Vitor, R.W.A. Susceptibility to Re-Infection in C57BL/6 Mice with Recombinant Strains of Toxoplasma Gondii. Exp. Parasitol. 2011, 128, 433–437. [Google Scholar] [CrossRef]
- Lindsay, D.S.; Dubey, J.P. Toxoplasma Gondii: The Changing Paradigm of Congenital Toxoplasmosis. Parasitology 2011, 138, 1829–1831. [Google Scholar] [CrossRef]
- Zulpo, D.L.; Sammi, A.S.; dos Santos, J.R.; Sasse, J.P.; Martins, T.A.; Minutti, A.F.; Cardim, S.T.; de Barros, L.D.; Navarro, I.T.; Garcia, J.L. Toxoplasma Gondii: A Study of Oocyst Re-Shedding in Domestic Cats. Vet. Parasitol. 2018, 249, 17–20. [Google Scholar] [CrossRef]
- Blaga, R.; Aubert, D.; Thébault, A.; Perret, C.; Geers, R.; Thomas, M.; Alliot, A.; Djokic, V.; Ortis, N.; Halos, L.; et al. Toxoplasma Gondii in Beef Consumed in France: Regional Variation in Seroprevalence and Parasite Isolation. Parasite 2019, 26, 77. [Google Scholar] [CrossRef]
- Alves, B.F.; Oliveira, S.; Soares, H.S.; Pena, H.F.J.; Conte-Junior, C.A.; Gennari, S.M. Isolation of Viable Toxoplasma Gondii from Organs and Brazilian Commercial Meat Cuts of Experimentally Infected Pigs. Parasitol. Res. 2019, 118, 1331–1335. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P.; Cerqueira-Cézar, C.K.; Murata, F.H.A.; Kwok, O.C.H.; Hill, D.; Yang, Y.; Su, C. All about Toxoplasma Gondii Infections in Pigs: 2009–2020. Vet. Parasitol. 2020, 288, 109185. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P. Toxoplasma Gondii Infections in Chickens (Gallus Domesticus): Prevalence, Clinical Disease, Diagnosis and Public Health Significance. Zoonoses Public Health 2010, 57, 60–73. [Google Scholar] [CrossRef] [PubMed]
- Conrad, P.A.; Miller, M.A.; Kreuder, C.; James, E.R.; Mazet, J.; Dabritz, H.; Jessup, D.A.; Gulland, F.; Grigg, M.E. Transmission of Toxoplasma: Clues from the Study of Sea Otters as Sentinels of Toxoplasma Gondii Flow into the Marine Environment. Int. J. Parasitol. 2005, 35, 1155–1168. [Google Scholar] [CrossRef] [PubMed]
- Burgess, T.L.; Tinker, M.T.; Miller, M.A.; Bodkin, J.L.; Murray, M.J.; Saarinen, J.A.; Nichol, L.M.; Larson, S.; Conrad, P.A.; Johnson, C.K. Defining the Risk Landscape in the Context of Pathogen Pollution: Toxoplasma Gondii in Sea Otters along the Pacific Rim. R. Soc. Open Sci. 2018, 5, 171178. [Google Scholar] [CrossRef]
- Shapiro, K.; VanWormer, E.; Packham, A.; Dodd, E.; Conrad, P.A.; Miller, M. Type X Strains of Toxoplasma Gondii Are Virulent for Southern Sea Otters (Enhydra Lutris Nereis) and Present in Felids from Nearby Watersheds. Proc. R. Soc. B Biol. Sci. 2019, 286, 20191334. [Google Scholar] [CrossRef] [PubMed]
- Frénal, K.; Dubremetz, J.F.; Lebrun, M.; Soldati-Favre, D. Gliding Motility Powers Invasion and Egress in Apicomplexa. Nat. Rev. Microbiol. 2017, 15, 645–660. [Google Scholar] [CrossRef]
- McFadden, G.I.; Yeh, E. The Apicoplast: Now You See It, Now You Don’t. Int. J. Parasitol. 2017, 47, 137–144. [Google Scholar] [CrossRef]
- Walker, G.; Dorrell, R.G.; Schlacht, A.; Dacks, J.B. Eukaryotic Systematics: A User’s Guide for Cell Biologists and Parasitologists. Parasitology 2011, 138, 1638–1663. [Google Scholar] [CrossRef]
- Šlapeta, J.; Morin-Adeline, V. Apicomplexa Levine 1970, Sporozoa Leucart. 1879. Available online: http://tolweb.org/Apicomplexa/2446/2011.05.18 (accessed on 1 December 2021).
- Koreny, L.; Zeeshan, M.; Barylyuk, K.; Tromer, E.C.; van Hooff, J.J.E.; Brady, D.; Ke, H.; Chelaghma, S.; Ferguson, D.J.P.; Eme, L.; et al. Molecular Characterization of the Conoid Complex in Toxoplasma Reveals Its Conservation in All Apicomplexans, Including Plasmodium Species. PLoS Biol. 2021, 19, e3001081. [Google Scholar] [CrossRef]
- Cavalier-Smith, T. Kingdom Protozoa and Its 18 Phyla. Microbiol. Rev. 1993, 57, 953–994. [Google Scholar] [CrossRef]
- Cavalier-Smith, T. A Revised Six-Kingdom System of Life. Biol. Rev. Camb. Philos. Soc. 1998, 73, 203–266. [Google Scholar] [CrossRef] [PubMed]
- Taylor, M.A.; Coop, R.L.; Wall, R.L. Veterinary Parasitology; Wiley-Blackwell: Oxford, UK, 2016; ISBN 978-0-470-67162-7. [Google Scholar]
- Ruggiero, M.A.; Gordon, D.P.; Orrell, T.M.; Bailly, N.; Bourgoin, T.; Brusca, R.C.; Cavalier-Smith, T.; Guiry, M.D.; Kirk, P.M. A Higher Level Classification of All Living Organisms. PLoS ONE 2015, 10, e0119248. [Google Scholar] [CrossRef]
- Rees, T.; Vandepitte, L.; Vanhoorne, B.; Decock, W. All Genera of the World: An Overview and Estimates Based on the March 2020 Release of the Interim Register of Marine and Nonmarine Genera (IRMNG). Megataxa 2020, 1, 123–140. [Google Scholar] [CrossRef]
- Howe, D.K.; Sibley, L.D. Toxoplasma Gondii Comprises Three Clonal Lineages: Correlation of Parasite Genotype with Human Disease. J. Infect. Dis. 1995, 172, 1561–1566. [Google Scholar] [CrossRef]
- Su, C.; Evans, D.; Cole, R.H.; Kissinger, J.C.; Ajioka, J.W.; Sibley, L.D. Recent Expansion of Toxoplasma through Enhanced Oral Transmission. Science 2003, 299, 414–416. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Fux, B.; Su, C.; Dubey, J.P.; Darde, M.L.; Ajioka, J.W.; Rosenthal, B.M.; Sibley, L.D. Recent Transcontinental Sweep of Toxoplasma Gondii Driven by a Single Monomorphic Chromosome. Proc. Natl. Acad. Sci. USA 2007, 104, 14872–14877. [Google Scholar] [CrossRef]
- Pena, H.F.J.; Marvulo, M.F.V.; Horta, M.C.; Silva, M.A.; Silva, J.C.R.; Siqueira, D.B.; Lima, P.A.C.P.; Vitaliano, S.N.; Gennari, S.M. Isolation and Genetic Characterisation of Toxoplasma Gondii from a Red-Handed Howler Monkey (Alouatta Belzebul), a Jaguarundi (Puma Yagouaroundi), and a Black-Eared Opossum (Didelphis Aurita) from Brazil. Vet. Parasitol. 2011, 175, 377–381. [Google Scholar] [CrossRef]
- Fux, B.; Nawas, J.; Khan, A.; Gill, D.B.; Su, C.; Sibley, L.D. Toxoplasma Gondii Strains Defective in Oral Transmission Are Also Defective in Developmental Stage Differentiation. Infect. Immun. 2007, 75, 2580–2590. [Google Scholar] [CrossRef] [PubMed]
- Mercier, A.; Ajzenberg, D.; Devillard, S.; Demar, M.P.; de Thoisy, B.; Bonnabau, H.; Collinet, F.; Boukhari, R.; Blanchet, D.; Simon, S.; et al. Human Impact on Genetic Diversity of Toxoplasma Gondii: Example of the Anthropized Environment from French Guiana. Infect. Genet. Evol. 2011, 11, 1378–1387. [Google Scholar] [CrossRef]
- Khan, A.; Ajzenberg, D.; Mercier, A.; Demar, M.; Simon, S.; Dardé, M.L.; Wang, Q.; Verma, S.K.; Rosenthal, B.M.; Dubey, J.P.; et al. Geographic Separation of Domestic and Wild Strains of Toxoplasma Gondii in French Guiana Correlates with a Monomorphic Version of Chromosome1a. PLoS Negl. Trop. Dis. 2014, 8, e3182. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, D.J.P.; Dubremetz, J.-F. The Ultrastructure of Toxoplasma gondii. In Toxoplasma Gondii; Weiss, L.M., Kim, K., Eds.; Elsevier: Amsterdam, The Netherlands, 2014; pp. 19–59. ISBN 9780123964816. [Google Scholar]
- Harding, C.R.; Frischknecht, F. The Riveting Cellular Structures of Apicomplexan Parasites. Trends Parasitol. 2020, 36, 979–991. [Google Scholar] [CrossRef] [PubMed]
- Attias, M.; Teixeira, D.E.; Benchimol, M.; Vommaro, R.C.; Crepaldi, P.H.; De Souza, W. The Life-Cycle of Toxoplasma Gondii Reviewed Using Animations. Parasites Vectors 2020, 13, 588. [Google Scholar] [CrossRef]
- Melo, E.J.L.; Attias, M.; De Souza, W. The Single Mitochondrion of Tachyzoites of Toxoplasma Gondii. J. Struct. Biol. 2000, 130, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Striepen, B.; Crawford, M.J.; Shaw, M.K.; Tilney, L.G.; Seeber, F.; Roos, D.S. The Plastid of Toxoplasma Gondii Is Divided by Association with the Centrosomes. J. Cell Biol. 2000, 151, 1423–1434. [Google Scholar] [CrossRef]
- Chen, C.T.; Gubbels, M.J. The Toxoplasma Gondii Centrosome Is the Platform for Internal Daughter Budding as Revealed by a Nek1 Kinase Mutant. J. Cell Sci. 2013, 126, 3344–3355. [Google Scholar] [CrossRef]
- Brooks, C.F.; Johnsen, H.; van Dooren, G.G.; Muthalagi, M.; Lin, S.S.; Bohne, W.; Fischer, K.; Striepen, B. The Toxoplasma Apicoplast Phosphate Translocator Links Cytosolic and Apicoplast Metabolism and Is Essential for Parasite Survival. Cell Host Microbe 2010, 7, 62–73. [Google Scholar] [CrossRef]
- Janouškovec, J.; Horák, A.; Oborník, M.; Lukeš, J.; Keeling, P.J. A Common Red Algal Origin of the Apicomplexan, Dinoflagellate, and Heterokont Plastids. Proc. Natl. Acad. Sci. USA 2010, 107, 10949–10954. [Google Scholar] [CrossRef]
- Joiner, K.A.; Roos, D.S. Secretory Traffic in the Eukaryotic Parasite Toxoplasma Gondii: Less Is More. J. Cell Biol. 2002, 157, 557–563. [Google Scholar] [CrossRef]
- Lebrun, M.; Carruthers, V.B.; Cesbron-Delauw, M.-F. Toxoplasma Secretory Proteins and Their Roles in Cell Invasion and Intracellular Survival. In Toxoplasma Gondii; Elsevier: Amsterdam, The Netherlands, 2014; pp. 389–453. ISBN 9780123964816. [Google Scholar]
- Kentaro, K. How Does Toxoplama Gondii Invade Host Cells? J. Vet. Med. Sci. 2018, 80, 1702–1706. [Google Scholar] [CrossRef]
- Dubey, J.P.; Lindsay, D.S.; Speer, C.A. Structures of Toxoplasma Gondii Tachyzoites, Bradyzoites, and Sporozoites and Biology and Development of Tissue Cysts. Clin. Microbiol. Rev. 1998, 11, 267–299. [Google Scholar] [CrossRef] [PubMed]
- Russell, D.G.; Burns, R.G. The Polar Ring of Coccidian Sporozoites: A Unique Microtubule-Organizing Centre. J. Cell Sci. 1984, 65, 193–207. [Google Scholar] [CrossRef] [PubMed]
- Nichols, B.A.; Chiappino, M.L. Cytoskeleton of Toxoplasma Gondii 1. J. Protozool. 1987, 34, 217–226. [Google Scholar] [CrossRef]
- Morrissette, N.S.; Sibley, L.D. Cytoskeleton of Apicomplexan Parasites. Microbiol. Mol. Biol. Rev. 2002, 66, 21–38. [Google Scholar] [CrossRef]
- Hu, K.; Johnson, J.; Florens, L.; Fraunholz, M.; Suravajjala, S.; DiLullo, C.; Yates, J.; Roos, D.S.; Murray, J.M. Cytoskeletal Components of an Invasion Machine-The Apical Complex of Toxoplasma Gondii. PLoS Pathog. 2006, 2, e13. [Google Scholar] [CrossRef]
- Katris, N.J.; van Dooren, G.G.; McMillan, P.J.; Hanssen, E.; Tilley, L.; Waller, R.F. The Apical Complex Provides a Regulated Gateway for Secretion of Invasion Factors in Toxoplasma. PLoS Pathog. 2014, 10, e1004074. [Google Scholar] [CrossRef] [PubMed]
- Mann, T.; Beckers, C. Characterization of the Subpellicular Network, a Filamentous Membrane Skeletal Component in the Parasite Toxoplasma Gondii. Mol. Biochem. Parasitol. 2001, 115, 257–268. [Google Scholar] [CrossRef]
- Hu, K. Organizational Changes of the Daughter Basal Complex during the Parasite Replication of Toxoplasma Gondii. PLoS Pathog. 2008, 4, e10. [Google Scholar] [CrossRef]
- Delbac, F.; Sänger, A.; Neuhaus, E.M.; Stratmann, R.; Ajioka, J.W.; Toursel, C.; Herm-Götz, A.; Tomavo, S.; Soldati, T.; Soldati, D. Toxoplasma Gondii Myosins B/C: One Gene, Two Tails, Two Localizations, and a Role in Parasite Division. J. Cell Biol. 2001, 155, 613–623. [Google Scholar] [CrossRef] [PubMed]
- Gubbels, M.-J.; Vaishnava, S.; Boot, N.; Dubremetz, J.-F.; Striepen, B. A MORN-Repeat Protein Is a Dynamic Component of the Toxoplasma Gondii Cell Division Apparatus. J. Cell Sci. 2006, 119, 2236–2245. [Google Scholar] [CrossRef]
- Heaslip, A.T.; Dzierszinski, F.; Stein, B.; Hu, K. TgMORN1 Is a Key Organizer for the Basal Complex of Toxoplasma Gondii. PLoS Pathog. 2010, 6, e1000754. [Google Scholar] [CrossRef] [PubMed]
- Lorestani, A.; Ivey, F.D.; Thirugnanam, S.; Busby, M.A.; Marth, G.T.; Cheeseman, I.M.; Gubbels, M.-J. Targeted Proteomic Dissection of Toxoplasma Cytoskeleton Sub-Compartments Using MORN1. Cytoskeleton 2012, 69, 1069–1085. [Google Scholar] [CrossRef]
- Knoll, L.J.; Tomita, T.; Weiss, L.M. Bradyzoite Development. In Toxoplasma Gondii; Weiss, L.M., Kim, K., Eds.; Academic Press: Cambridge, MA, USA, 2014; pp. 521–549. ISBN 9780123964816. [Google Scholar]
- Jacobs, L.; Remington, J.S.; Melton, M.L. The Resistance of the Encysted Form of Toxoplasma Gondii. J. Parasitol. 1960, 46, 11. [Google Scholar] [CrossRef] [PubMed]
- Popiel, I.; Gold, M.C.; Booth, K.S. Quantification of Toxoplasma Gondii Bradyzoites. J. Parasitol. 1996, 82, 330–332. [Google Scholar] [CrossRef]
- Di Genova, B.M.; Wilson, S.K.; Dubey, J.P.; Knoll, L.J. Intestinal Delta-6-Desaturase Activity Determines Host Range for Toxoplasma Sexual Reproduction. PLoS Biol. 2019, 17, e3000364. [Google Scholar] [CrossRef]
- Freyre, A.; Dubey, J.P.; Smith, D.D.; Frenkel, J.K. Gondii Oocyst-Induced Infections. J. Parasitol. 1989, 75, 750–755. [Google Scholar] [CrossRef]
- Dubey, J.P. Oocyst Shedding by Cats Fed Isolated Bradyzoites and Comparison of Infectivity of Bradyzoites of the VEG Strain Toxoplasma Gondii to Cats and Mice. J. Parasitol. 2001, 87, 215–219. [Google Scholar] [CrossRef]
- Dubey, J.P. Comparative Infectivity of Oocysts and Bradyzoites of Toxoplasma Gondii for Intermediate (Mice) and Definitive (Cats) Hosts. Vet. Parasitol. 2006, 140, 69–75. [Google Scholar] [CrossRef]
- Tomasina, R.; Francia, M.E. The Structural and Molecular Underpinnings of Gametogenesis in Toxoplasma Gondii. Front. Cell. Infect. Microbiol. 2020, 10, 1–8. [Google Scholar] [CrossRef]
- Dubey, J.P.; Frenkel, J.K. Cyst-Induced Toxoplasmosis in Cats. J. Protozool. 1972, 19, 155–177. [Google Scholar] [CrossRef]
- Speer, C.A.; Dubey, J.P. Ultrastructural Differentiation of Toxoplasma Gondii Schizonts (Types B to E) and Gamonts in the Intestines of Cats Fed Bradyzoites. Int. J. Parasitol. 2005, 35, 193–206. [Google Scholar] [CrossRef] [PubMed]
- Francia, M.E.; Striepen, B. Cell Division in Apicomplexan Parasites. Nat. Rev. Microbiol. 2014, 12, 125–136. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P. Duration of Immunity to Shedding of Toxoplasma Gondii Oocysts by Cats. J. Parasitol. 1995, 81, 410–415. [Google Scholar] [CrossRef] [PubMed]
- Fritz, H.M.; Bowyer, P.W.; Bogyo, M.; Conrad, P.A.; Boothroyd, J.C. Proteomic Analysis of Fractionated Toxoplasma Oocysts Reveals Clues to Their Environmental Resistance. PLoS ONE 2012, 7, e29955. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, D.J.P. Toxoplasma Gondii and Sex: Essential or Optional Extra? Trends Parasitol. 2002, 18, 351–355. [Google Scholar] [CrossRef]
- Dubey, J.P. Pathogenicity and Infectivity of Toxoplasma Gondii Oocysts for Rats. J. Parasitol. 1996, 82, 951–956. [Google Scholar] [CrossRef]
- Dubey, R.; Harrison, B.; Dangoudoubiyam, S.; Bandini, G.; Cheng, K.; Kosber, A.; Agop-nersesian, C.; Howe, D.K.; Samuelson, J.; Ferguson, D.J.P.; et al. Differential Roles for Inner Membrane Complex Proteins across Toxoplasma Gondii and Sarcocystis Neurona Development. mSphere 2017, 2, 1–19. [Google Scholar] [CrossRef]
- Freppel, W.; Ferguson, D.J.P.; Shapiro, K.; Dubey, J.P.; Puech, P.H.; Dumètre, A. Structure, Composition, and Roles of the Toxoplasma Gondii Oocyst and Sporocyst Walls. Cell Surf. 2019, 5, 100016. [Google Scholar] [CrossRef]
- King, C.A. Cell Motility of Sporozoan Protozoa. Parasitol. Today 1988, 4, 315–319. [Google Scholar] [CrossRef]
- Håkansson, S.; Morisaki, H.; Heuser, J.; Sibley, L.D. Time-Lapse Video Microscopy of Gliding Motility in Toxoplasma Gondii Reveals a Novel, Biphasic Mechanism of Cell Locomotion. Mol. Biol. Cell 1999, 10, 3539–3547. [Google Scholar] [CrossRef] [PubMed]
- Dobrowolski, J.M.; Sibley, L.D. Toxoplasma Invasion of Mammalian Cells Is Powered by the Actin Cytoskeleton of the Parasite. Cell 1996, 84, 933–939. [Google Scholar] [CrossRef]
- Dobrowolski, J.M.; Carruthers, V.B.; Sibley, L.D. Participation of Myosin in Gliding Motility and Host Cell Invasion by Toxoplasma Gondii. Mol. Microbiol. 1997, 26, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Meissner, M.; Schlüter, D.; Soldati, D. Role of Toxoplasma Gondii Myosin a in Powering Parasite Gliding and Host Cell Invasion. Science 2002, 298, 837–840. [Google Scholar] [CrossRef]
- Opitz, C.; Soldati, D. “The Glideosome”: A Dynamic Complex Powering Gliding Motion and Host Cell Invasion by Toxoplasma Gondii. Mol. Microbiol. 2002, 45, 597–604. [Google Scholar] [CrossRef]
- Brossier, F.; Jewett, T.J.; Lovett, J.L.; Sibley, L.D. C-Terminal Processing of the Toxoplasma Protein MIC2 Is Essential for Invasion into Host Cells. J. Biol. Chem. 2003, 278, 6229–6234. [Google Scholar] [CrossRef]
- Gras, S.; Jimenez-Ruiz, E.; Klinger, C.M.; Schneider, K.; Klingl, A.; Lemgruber, L.; Meissner, M. An Endocytic-Secretory Cycle Participates in Toxoplasma Gondii in Motility. PLoS Biol. 2019, 17, e3000060. [Google Scholar] [CrossRef]
- Soldati, D.; Meissner, M. Toxoplasma as a Novel System for Motility. Curr. Opin. Cell Biol. 2004, 16, 32–40. [Google Scholar] [CrossRef]
- Mineo, J.R.; Kasper, L.H. Attachment of Toxoplasma Gondii to Host Cells Involves Major Surface Protein, SAG-1 (P-30). Exp. Parasitol. 1994, 79, 11–20. [Google Scholar] [CrossRef]
- Jacquet, A.; Coulon, L.; De Nève, J.; Daminet, V.; Haumont, M.; Garcia, L.; Bollen, A.; Jurado, M.; Biemans, R. The Surface Antigen SAG3 Mediates the Attachment of Toxoplasma Gondii to Cell-Surface Proteoglycans. Mol. Biochem. Parasitol. 2001, 116, 35–44. [Google Scholar] [CrossRef]
- Rabenau, K.E.; Sohrabi, A.; Tripathy, A.; Reitter, C.; Ajioka, J.W.; Tomley, F.M.; Carruthers, V.B. TgM2AP Participates in Toxoplasma Gondii Invasion of Host Cells and Is Tightly Associated with the Adhesive Protein TgMIC2. Mol. Microbiol. 2001, 41, 537–547. [Google Scholar] [CrossRef]
- Cérède, O.; Dubremetz, J.F.; Soête, M.; Deslée, D.; Vial, H.; Bout, D.; Lebrun, M. Synergistic Role of Micronemal Proteins in Toxoplasma Gondii Virulence. J. Exp. Med. 2005, 201, 453–463. [Google Scholar] [CrossRef]
- Whitelaw, J.A.; Latorre-Barragan, F.; Gras, S.; Pall, G.S.; Leung, J.M.; Heaslip, A.; Egarter, S.; Andenmatten, N.; Nelson, S.R.; Warshaw, D.M.; et al. Surface Attachment, Promoted by the Actomyosin System of Toxoplasma Gondii Is Important for Efficient Gliding Motility and Invasion. BMC Biol. 2017, 15, 1. [Google Scholar] [CrossRef]
- Alexander, D.L.; Mital, J.; Ward, G.E.; Bradley, P.; Boothroyd, J.C. Identification of the Moving Junction Complex of Toxoplasma Gondii: A Collaboration between Distinct Secretory Organelles. PLoS Pathog. 2005, 1, e17. [Google Scholar] [CrossRef]
- Carruthers, V.; Boothroyd, J.C. Pulling Together: An Integrated Model of Toxoplasma Cell Invasion. Curr. Opin. Microbiol. 2007, 10, 83–89. [Google Scholar] [CrossRef]
- Mordue, D.G.; Desai, N.; Dustin, M.; Sibley, L.D. Proteins on the Basis of Their Membrane Anchoring. J. Exp. Med 1999, 190, 1783–1792. [Google Scholar] [CrossRef] [PubMed]
- Charron, A.J.; Sibley, L.D. Molecular Partitioning during Host Cell Penetration by Toxoplasma Gondii. Traffic 2004, 5, 855–867. [Google Scholar] [CrossRef] [PubMed]
- Walker, M.E.; Hjort, E.E.; Smith, S.S.; Tripathi, A.; Hornick, J.E.; Hinchcliffe, E.H.; Archer, W.; Hager, K.M. Toxoplasma Gondii Actively Remodels the Microtubule Network in Host Cells. Microbes Infect. 2008, 10, 1440–1449. [Google Scholar] [CrossRef] [PubMed]
- Sweeney, K.R.; Morrissette, N.S.; Lachapelle, S.; Blader, I.J. Host Cell Invasion by Toxoplasma Gondii Is Temporally Regulated by the Host Microtubule Cytoskeleton. Eukaryot. Cell 2010, 9, 1680–1689. [Google Scholar] [CrossRef]
- Cardoso, R.; Nolasco, S.; Goncalves, J.; Cortes, H.C.; Leitao, A.; Soares, H. Besnoitia Besnoiti and Toxoplasma Gondii: Two Apicomplexan Strategies to Manipulate the Host Cell Centrosome and Golgi Apparatus. Parasitology 2014, 141, 1436–1454. [Google Scholar] [CrossRef] [PubMed]
- Sibley, L.D.; Niesman, I.R.; Parmley, S.F.; Cesbron-Delauw, M.F. Regulated Secretion of Multi-Lamellar Vesicles Leads to Formation of a Tubulovesicular Network in Host-Cell Vacuoles Occupied by Toxoplasma Gondii. J. Cell Sci. 1995, 108, 1669–1677. [Google Scholar] [CrossRef]
- Mercier, C.; Dubremetz, J.-F.; Rauscher, B.; Lecordier, L.; Sibley, L.D.; Cesbron-Delauw, M.-F. Biogenesis of Nanotubular Network in Toxoplasma Parasitophorous Vacuole Induced by Parasite Proteins. Mol. Biol. Cell 2002, 13, 2397–2409. [Google Scholar] [CrossRef] [PubMed]
- De Melo, E.J.T.; de Carvalho, T.U.; de Souza, W. Penetration of Toxoplasma Gondii into Host Cells Induces Changes in the Distribution of the Mitochondria and the Endoplasmic Reticulum. Cell Struct. Funct. 1992, 17, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Melo, E.J.T.; De Souza, W. Relationship between the Host Cell Endoplasmic Reticulum and the Parasitophorous Vacuole Containing Toxoplasma Gondii. Cell Struct. Funct. 1997, 22, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Sinai, A.P.; Webster, P.; Joiner, K.A. Association of Host Cell Endoplasmic Reticulum and Mitochondria with the Toxoplasma Gondii Parasitophorous Vacuole Membrane: A High Affinity Interaction. J. Cell Sci. 1997, 110, 2117–2128. [Google Scholar] [CrossRef]
- Melo, E.J.T.; Carvalho, T.M.U.; de Souza, W. Behaviour of Microtubules in Cells Infected with Toxoplasma Gondii. Biocell 2001, 25, 53–59. [Google Scholar]
- Sehgal, A.; Bettiol, S.; Pypaert, M.; Wenk, M.R.; Kaasch, A.; Blader, I.J.; Joiner, K.A.; Coppens, I. Peculiarities of Host Cholesterol Transport to the Unique Intracellular Vacuole Containing Toxoplasma. Traffic 2005, 6, 1125–1141. [Google Scholar] [CrossRef] [PubMed]
- Coppens, I.; Dunn, J.D.; Romano, J.D.; Pypaert, M.; Zhang, H.; Boothroyd, J.C.; Joiner, K.A. Toxoplasma Gondii Sequesters Lysosomes from Mammalian Hosts in the Vacuolar Space. Cell 2006, 125, 261–274. [Google Scholar] [CrossRef]
- Coppens, I.; Sinai, A.P.; Joiner, K.A. Toxoplasma Gondii Exploits Host Low-Density Lipoprotein Receptor- Mediated Endocytosis for Cholesterol Acquisition. J. Cell Biol. 2000, 149, 167–180. [Google Scholar] [CrossRef]
- Romano, J.D.; Sonda, S.; Bergbower, E.; Smith, M.E.; Coppens, I. Toxoplasma Gondii Salvages Sphingolipids from the Host Golgi through the Rerouting of Selected Rab Vesicles to the Parasitophorous Vacuole. Mol. Biol. Cell 2013, 24, 1974–1995. [Google Scholar] [CrossRef] [PubMed]
- Nolan, S.J.; Romano, J.D.; Coppens, I. Host Lipid Droplets: An Important Source of Lipids Salvaged by the Intracellular Parasite Toxoplasma Gondii. PLoS Pathog. 2017, 13, e1006362. [Google Scholar] [CrossRef] [PubMed]
- Romano, J.D.; Nolan, S.J.; Porter, C.; Ehrenman, K.; Hartman, E.J.; Hsia, R.-C.; Coppens, I. The Parasite Toxoplasma Sequesters Diverse Rab Host Vesicles within an Intravacuolar Network. J. Cell Biol. 2017, 216, 4235–4254. [Google Scholar] [CrossRef]
- Jones, T.C.; Hirsch, J.G. The Interaction between Toxoplasma Gondii and Mammalian Cells: II. The Absence of Lysosomal Fusion with Phagocytic Vacuoles Containing Living Parasites. J. Exp. Med. 1972, 136, 1173–1194. [Google Scholar] [CrossRef]
- Carvalho, C.S.; Melo, E.J.T. Acidification of the Parasitophorous Vacuole Containing Toxoplasma Gondii in the Presence of Hydroxyurea. An. Acad. Bras. Cienc. 2006, 78, 475–484. [Google Scholar] [CrossRef]
- Radke, J.R.; Striepen, B.; Guerini, M.N.; Jerome, M.E.; Roos, D.S.; White, M.W. Defining the Cell Cycle for the Tachyzoite Stage of Toxoplasma Gondii. Mol. Biochem. Parasitol. 2001, 115, 165–175. [Google Scholar] [CrossRef]
- Sheffield, H.G.; Melton, M.L. The Fine Structure and Reproduction of Toxoplasma Gondii. J. Parasitol. 1968, 54, 209–226. [Google Scholar] [CrossRef] [PubMed]
- Hu, K.; Mann, T.; Striepen, B.; Beckers, C.J.M.; Roos, D.S.; Murray, J.M. Daughter Cell Assembly in the Protozoan Parasite Toxoplasma Gondii. Mol. Biol. Cell 2002, 13, 593–606. [Google Scholar] [CrossRef]
- Hartmann, J.; Hu, K.; He, C.Y.; Pelletier, L.; Roos, D.S.; Warren, G. Golgi and Centrosome Cycles in Toxoplasma Gondii. Mol. Biochem. Parasitol. 2006, 145, 125–127. [Google Scholar] [CrossRef] [PubMed]
- Nishi, M.; Hu, K.; Murray, J.M.; Roos, D.S. Organellar Dynamics during the Cell Cycle of Toxoplasma Gondii. J. Cell Sci. 2008, 121, 1559–1568. [Google Scholar] [CrossRef]
- Anderson-White, B.; Beck, J.R.; Chen, C.-T.; Meissner, M.; Bradley, P.J.; Gubbels, M.-J. Cytoskeleton Assembly in Toxoplasma Gondii Cell Division. Int. Rev. Cell Mol. Biol. 2012, 298, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Gubbels, M.-J.; White, M.; Szatanek, T. The Cell Cycle and Toxoplasma Gondii Cell Division: Tightly Knit or Loosely Stitched? Int. J. Parasitol. 2008, 38, 1343–1358. [Google Scholar] [CrossRef]
- Francia, M.E.; Jordan, C.N.; Patel, J.D.; Sheiner, L.; Demerly, J.L.; Fellows, J.D.; de Leon, J.C.; Morrissette, N.S.; Dubremetz, J.F.; Striepen, B. Cell Division in Apicomplexan Parasites Is Organized by a Homolog of the Striated Rootlet Fiber of Algal Flagella. PLoS Biol. 2012, 10, e1001444. [Google Scholar] [CrossRef] [PubMed]
- Caldas, L.; de Souza, W. A Window to Toxoplasma Gondii Egress. Pathogens 2018, 7, 69. [Google Scholar] [CrossRef]
- Kafsack, B.F.C.; Pena, J.D.O.; Coppens, I.; Ravindran, S.; Boothroyd, J.C.; Carruthers, V.B. Rapid Membrane Disruption by a Perforin-like Protein Facilitates Parasite Exit from Host Cells. Science 2009, 323, 530–533. [Google Scholar] [CrossRef]
- Okada, T.; Marmansari, D.; Li, Z.M.; Adilbish, A.; Canko, S.; Ueno, A.; Shono, H.; Furuoka, H.; Igarashi, M. A Novel Dense Granule Protein, GRA22, Is Involved in Regulating Parasite Egress in Toxoplasma Gondii. Mol. Biochem. Parasitol. 2013, 189, 5–13. [Google Scholar] [CrossRef]
- Gras, S.; Jackson, A.; Woods, S.; Pall, G.; Whitelaw, J.; Leung, J.M.; Ward, G.E.; Roberts, C.W.; Meissner, M. Parasites Lacking the Micronemal Protein MIC2 Are Deficient in Surface Attachment and Host Cell Egress, but Remain Virulent In Vivo. Wellcome Open Res. 2017, 2, 1–26. [Google Scholar] [CrossRef]
- LaFavers, K.A.; Márquez-Nogueras, K.M.; Coppens, I.; Moreno, S.N.J.; Arrizabalaga, G. A Novel Dense Granule Protein, GRA41, Regulates Timing of Egress and Calcium Sensitivity in Toxoplasma Gondii. Cell. Microbiol. 2017, 19, e12749. [Google Scholar] [CrossRef]
- Schultz, A.J.; Carruthers, V.B. Toxoplasma Gondii LCAT Primarily Contributes to Tachyzoite Egress. mSphere 2018, 3, 1–10. [Google Scholar] [CrossRef]
- Frénal, K.; Marq, J.B.; Jacot, D.; Polonais, V.; Soldati-Favre, D. Plasticity between MyoC- and MyoA-Glideosomes: An Example of Functional Compensation in Toxoplasma Gondii Invasion. PLoS Pathog. 2014, 10, e1004504. [Google Scholar] [CrossRef] [PubMed]
- Graindorge, A.; Frénal, K.; Jacot, D.; Salamun, J.; Marq, J.B.; Soldati-Favre, D. The Conoid Associated Motor MyoH Is Indispensable for Toxoplasma Gondii Entry and Exit from Host Cells. PLoS Pathog. 2016, 12, e1005388. [Google Scholar] [CrossRef]
- Jacot, D.; Tosetti, N.; Pires, I.; Stock, J.; Graindorge, A.; Hung, Y.F.; Han, H.; Tewari, R.; Kursula, I.; Soldati-Favre, D. An Apicomplexan Actin-Binding Protein Serves as a Connector and Lipid Sensor to Coordinate Motility and Invasion. Cell Host Microbe 2016, 20, 731–743. [Google Scholar] [CrossRef] [PubMed]
- Bisio, H.; Soldati-Favre, D. Signaling Cascades Governing Entry into and Exit from Host Cells by Toxoplasma Gondii. Annu. Rev. Microbiol. 2019, 73, 579–599. [Google Scholar] [CrossRef]
- Endo, T.; Sethi, K.K.; Piekarski, G. Toxoplasma Gondii: Calcium Lonophore A23187-Mediated Exit of Trophozoites from Infected Murine Macrophages. Exp. Parasitol. 1982, 53, 179–188. [Google Scholar] [CrossRef]
- Lourido, S.; Tang, K.; David Sibley, L. Distinct Signalling Pathways Control Toxoplasma Egress and Host-Cell Invasion. EMBO J. 2012, 31, 4524–4534. [Google Scholar] [CrossRef]
- Radke, J.R.; Guerini, M.N.; Jerome, M.; White, M.W. A Change in the Premitotic Period of the Cell Cycle Is Associated with Bradyzoite Differentiation in Toxoplasma Gondii. Mol. Biochem. Parasitol. 2003, 131, 119–127. [Google Scholar] [CrossRef]
- Watts, E.; Zhao, Y.; Dhara, A.; Eller, B.; Patwardhan, A.; Sinai, P. Novel Approaches Reveal That Toxoplasma Gondii Bradyzoites within Tissue Cysts Are Dynamic and Replicating Entities In Vivo. MBio 2015, 6, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Weiss, L.M.; Kim, K. The Development and Biology of Bradyzoites of Toxoplasma Gondii. Front. Biosci. 2000, 5, 391–405. [Google Scholar] [CrossRef]
- Ferguson, D.J.P.; Hutchison, W.M. An Ultrastructural Study of the Early Development and Tissue Cyst Formation of Toxoplasma Gondii in the Brains of Mice. Parasitol. Res. 1987, 73, 483–491. [Google Scholar] [CrossRef]
- Boothroyd, J.C.; Black, M.; Bonnefoy, S.; Hehl, A.; Knoll, L.J.; Manger, I.D.; Ortega-Barria, E.; Tomavo, S. Genetic and Biochemical Analysis of Development in Toxoplasma Gondii. Philos. Trans. R. Soc. B Biol. Sci. 1997, 352, 1347–1354. [Google Scholar] [CrossRef]
- Paredes-Santos, T.C.; Tomita, T.; Yan Fen, M.; de Souza, W.; Attias, M.; Vommaro, R.C.; Weiss, L.M. Development of Dual Fluorescent Stage Specific Reporter Strain of Toxoplasma Gondii to Follow Tachyzoite and Bradyzoite Development In Vitro and In Vivo. Microbes Infect. 2016, 18, 39–47. [Google Scholar] [CrossRef]
- Paredes-Santos, T.C.; Martins-Duarte, E.S.; Vitor, R.W.A.; de Souza, W.; Attias, M.; Vommaro, R.C. Spontaneous Cystogenesis In Vitro of a Brazilian Strain of Toxoplasma Gondii. Parasitol. Int. 2013, 62, 181–188. [Google Scholar] [CrossRef]
- Lane, A.; Soete, M.; Dubremetz, J.F.; Smith, J.E. Toxoplasma Gondii: Appearance of Specific Markers during the Development of Tissue Cysts In Vitro. Parasitol. Res. 1996, 82, 340–346. [Google Scholar] [CrossRef] [PubMed]
- Soete, M.; Fortier, B.; Camus, D.; Dubremetz, J.F. Toxoplasma Gondii: Kinetics of Bradyzoite-Tachyzoite Interconversion In Vitro. Exp. Parasitol. 1993, 76, 259–264. [Google Scholar] [CrossRef] [PubMed]
- Soete, M.; Camus, D.; Dubrametz, J.F. Experimental Induction of Bradyzoite-Specific Antigen Expression and Cyst Formation by the RH Strain of Toxoplasma Gondii In Vitro. Exp. Parasitol. 1994, 78, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Weiss, L.M.; Laplace, D.; Takvorian, P.M.; Tanowitz, H.B.; Cali, A.; Wittner, M. A Cell Culture System for Study of the Development of Toxoplasma Gondii Bradyzoites. J. Eukaryot. Microbiol. 1995, 42, 150–157. [Google Scholar] [CrossRef]
- Bohne, W.; Heesemann, J.; Gross, U. Induction of Bradyzoite-Specific Toxoplasma Gondii Antigens in Gamma Interferon-Treated Mouse Macrophages. Infect. Immun. 1993, 61, 1141–1145. [Google Scholar] [CrossRef]
- Bohne, W.; Heesemann, J.; Gross, U. Reduced Replication of Toxoplasma Gondii Is Necessary for Induction of Bradyzoite-Specific Antigens: A Possible Role for Nitric Oxide in Triggering Stage Conversion. Infect. Immun. 1994, 62, 1761–1767. [Google Scholar] [CrossRef]
- Fox, B.A.; Gigley, J.P.; Bzik, D.J. Toxoplasma Gondii Lacks the Enzymes Required for de Novo Arginine Biosynthesis and Arginine Starvation Triggers Cyst Formation. Int. J. Parasitol. 2004, 34, 323–331. [Google Scholar] [CrossRef]
- Ihara, F.; Nishikawa, Y. Starvation of Low-Density Lipoprotein-Derived Cholesterol Induces Bradyzoite Conversion in Toxoplasma Gondii. Parasites Vectors 2014, 7, 248. [Google Scholar] [CrossRef]
- Tomavo, S.; Boothroyd, J.C. Interconnection between Organellar Functions, Development and Drug Resistance in the Protozoan Parasite, Toxoplasma Gondii. Int. J. Parasitol. 1995, 25, 1293–1299. [Google Scholar] [CrossRef]
- Kirkman, L.A.; Weiss, L.M.; Kim, K. Cyclic Nucleotide Signaling in Toxoplasma Gondii Bradyzoite Differentiation. Infect. Immun. 2001, 69, 148–153. [Google Scholar] [CrossRef] [PubMed]
- Eaton, M.S.; Weiss, L.M.; Kim, K. Cyclic Nucleotide Kinases and Tachyzoite—Bradyzoite Transition in Toxoplasma Gondii. Int. J. Parasitol. 2006, 36, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, W.J.; Narasimhan, J.; Bhatti, M.M.; Wek, R.C. Parasite-Specific EIF2 (Eukaryotic Initiation Factor-2) Kinase Required for Stress-Induced Translation Control. Biochem. J. 2004, 380, 523–531. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, W.J.; Hakimi, M.A. Histone Mediated Gene Activation in Toxoplasma Gondii. Mol. Biochem. Parasitol. 2006, 148, 109–116. [Google Scholar] [CrossRef]
- Bougdour, A.; Maubon, D.; Baldacci, P.; Ortet, P.; Bastien, O.; Bouillon, A.; Barale, J.C.; Pelloux, H.; Ménard, R.; Hakimi, M.A. Drug Inhibition of HDAC3 and Epigenetic Control of Differentiation in Apicomplexa Parasites. J. Exp. Med. 2009, 206, 953–966. [Google Scholar] [CrossRef]
- Radke, J.R.; Behnke, M.S.; Mackey, A.J.; Radke, J.B.; Roos, D.S.; White, M.W. The Transcriptome of Toxoplasma Gondii. BMC Biol. 2005, 3, 26. [Google Scholar] [CrossRef]
- Song, X.; Lin, M.; Li, M.; Yang, X.; Liu, J.; Liu, Q. Toxoplasma Gondii Metacaspase 2 Is an Important Factor That Influences Bradyzoite Formation in the Pru Strain. Parasitol. Res. 2020, 119, 2287–2298. [Google Scholar] [CrossRef]
- Huang, S.; Holmes, M.J.; Radke, J.B.; Hong, D.-P.; Liu, T.; White, M.W.; Sullivan, W.J. Toxoplasma Gondii AP2IX-4 Regulates Gene Expression during Bradyzoite Development. Host Microbe Biol. 2017, 2, 1–16. [Google Scholar] [CrossRef]
- Radke, J.B.; Worth, D.; Hong, D.; Huang, S.; Sullivan, W.J.; Wilson, E.H.; White, M.W. Transcriptional Repression by ApiAP2 Factors Is Central to Chronic Toxoplasmosis. PLoS Pathog. 2018, 14, e1007035. [Google Scholar] [CrossRef]
- Farhat, D.C.; Swale, C.; Dard, C.; Cannella, D.; Ortet, P.; Barakat, M.; Sindikubwabo, F.; Belmudes, L.; De Bock, P.J.; Couté, Y.; et al. A MORC-Driven Transcriptional Switch Controls Toxoplasma Developmental Trajectories and Sexual Commitment. Nat. Microbiol. 2020, 5, 570–583. [Google Scholar] [CrossRef] [PubMed]
- Waldman, B.S.; Schwarz, D.; Wadsworth, M.H.; Saeij, J.P.; Shalek, A.K.; Lourido, S. Identification of a Master Regulator of Differentiation in Toxoplasma. Cell 2020, 180, 359–372. [Google Scholar] [CrossRef] [PubMed]
- Hong, D.; Radke, J.B.; White, M.W. Opposing Transcriptional Mechanisms Regulate Toxoplasma Development. Mol. Biol. Physiol. 2017, 2, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Croken, M.M.; Qiu, W.; White, M.W.; Kim, K. Gene Set Enrichment Analysis (GSEA) of Toxoplasma Gondii Expression Datasets Links Cell Cycle Progression and the Bradyzoite Developmental Program. BMC Genom. 2014, 15, 515. [Google Scholar] [CrossRef]
- Behnke, M.S.; Wootton, J.C.; Lehmann, M.M.; Radke, J.B.; Lucas, O.; Nawas, J.; Sibley, L.D.; White, M.W. Coordinated Progression through Two Subtranscriptomes Underlies the Tachyzoite Cycle of Toxoplasma Gondii. PLoS ONE 2010, 5, e12354. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | Classical Classification 1 | Updated Classification 2 |
|---|---|---|
| Kingdom | Protozoa | Chromista |
| Subkingdom | Dictyozoa | Harosa |
| Infrakingdom | Neozoa | Halvaria |
| Superphylum | Alveolata | Alveolata |
| Phylum | Apicomplexa | Myozoa |
| Infraphylum | Sporozoa | Apicomplexa |
| Superclass | Coccidia | Sporozoa |
| Class | Eucoccidea | Coccidiomorphea |
| Order | Eucoccidiorida | Eimeriida |
| Family | Sarcocystidae | Sarcocystidae |
| Genus | Toxoplasma | Toxoplasma |
| Species | Toxoplasma gondii | Toxoplasma gondii |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Delgado, I.L.S.; Zúquete, S.; Santos, D.; Basto, A.P.; Leitão, A.; Nolasco, S. The Apicomplexan Parasite Toxoplasma gondii. Encyclopedia 2022, 2, 189-211. https://doi.org/10.3390/encyclopedia2010012
Delgado ILS, Zúquete S, Santos D, Basto AP, Leitão A, Nolasco S. The Apicomplexan Parasite Toxoplasma gondii. Encyclopedia. 2022; 2(1):189-211. https://doi.org/10.3390/encyclopedia2010012
Chicago/Turabian StyleDelgado, Inês L. S., Sara Zúquete, Dulce Santos, Afonso P. Basto, Alexandre Leitão, and Sofia Nolasco. 2022. "The Apicomplexan Parasite Toxoplasma gondii" Encyclopedia 2, no. 1: 189-211. https://doi.org/10.3390/encyclopedia2010012