
Lichen as Multipartner Symbiotic Relationships




Definition
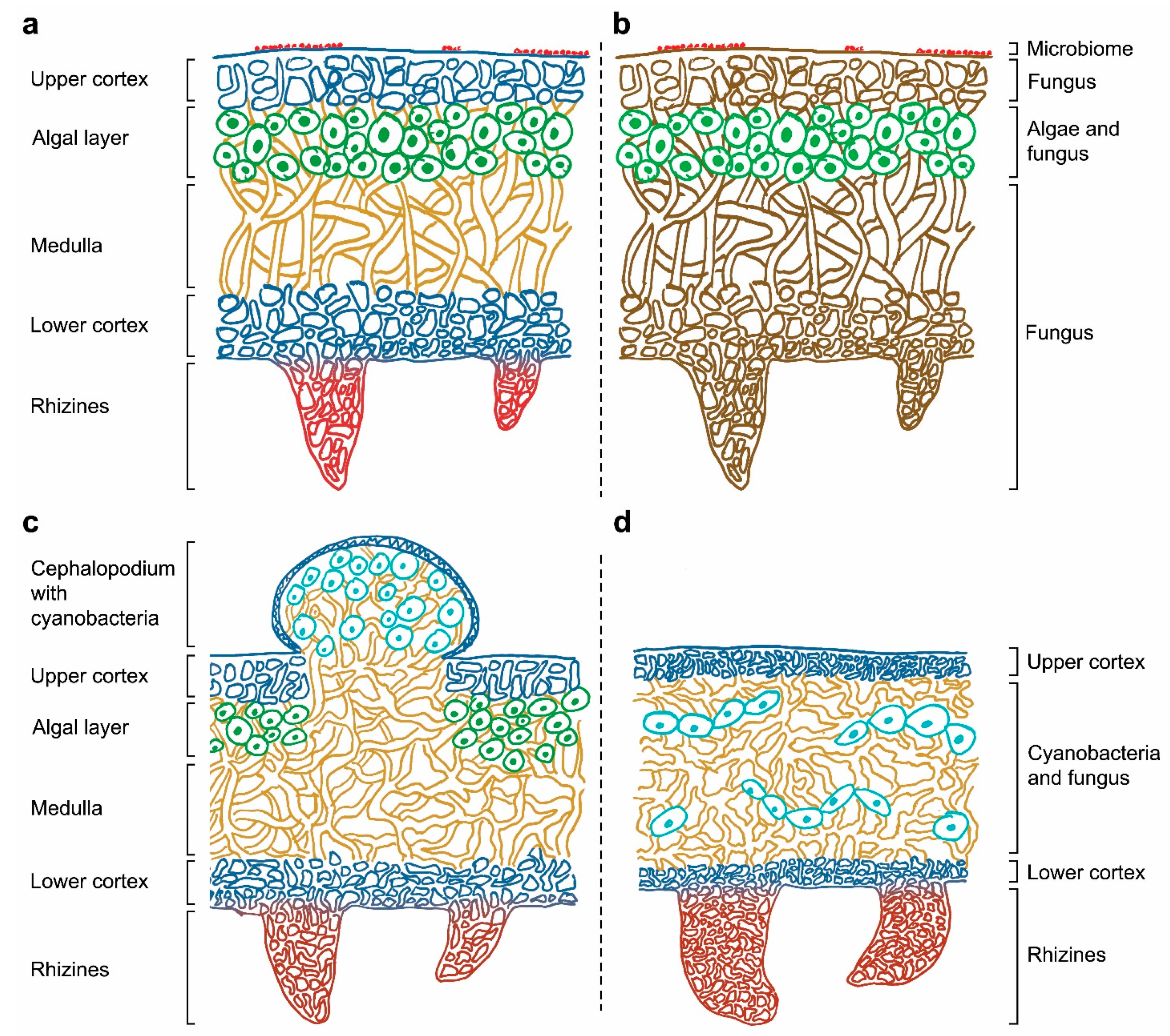
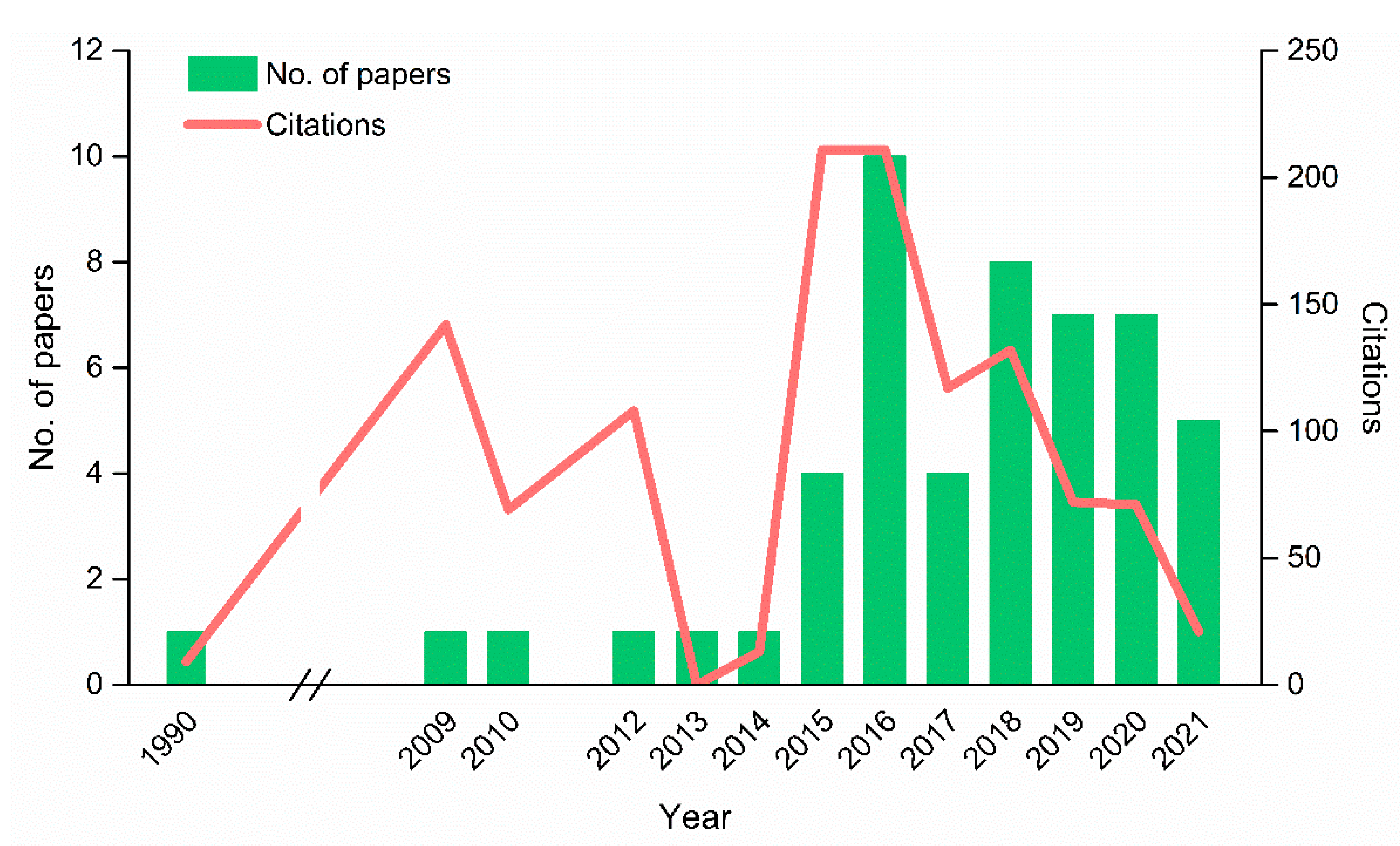
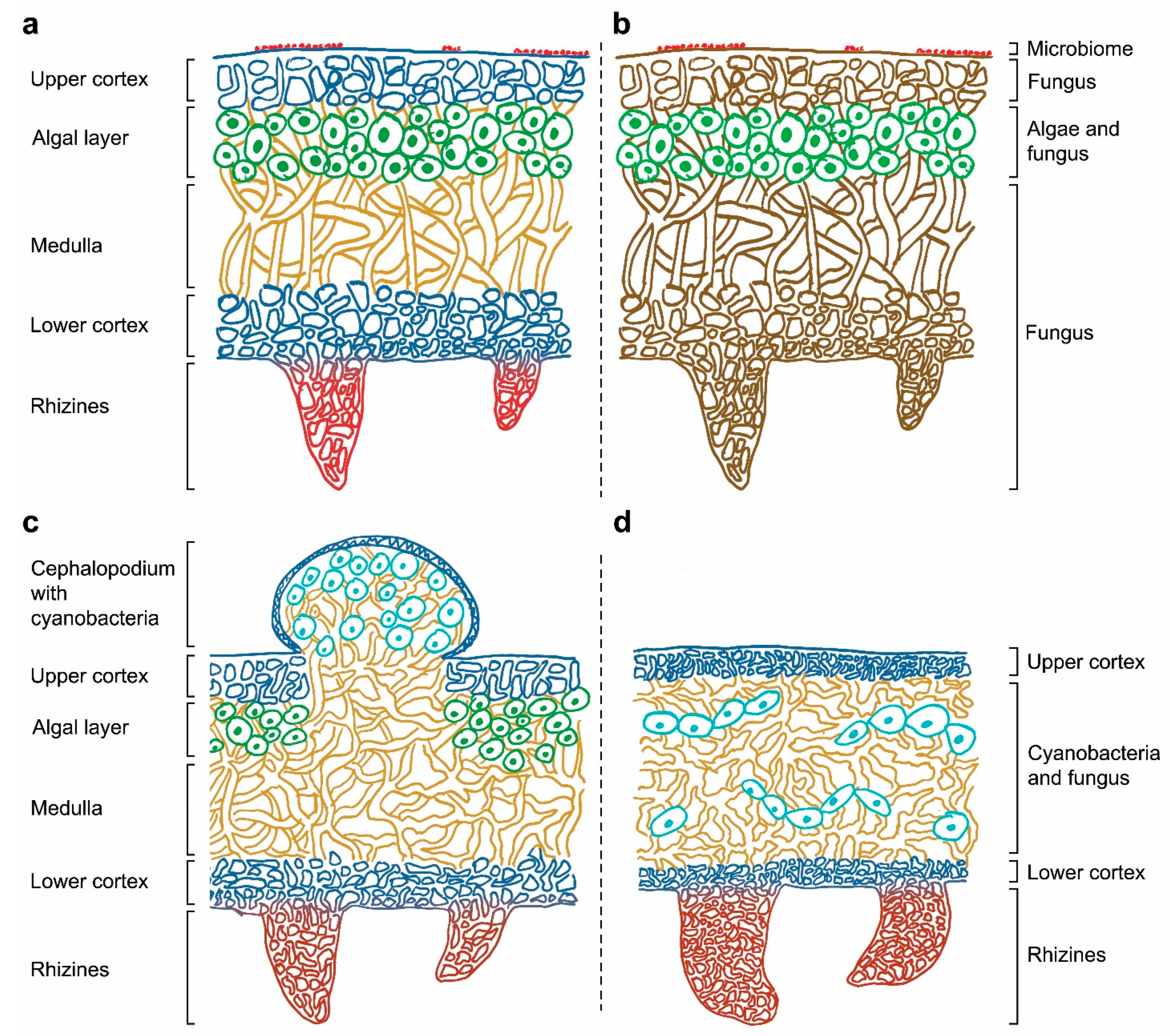
:1. General Context of Lichens
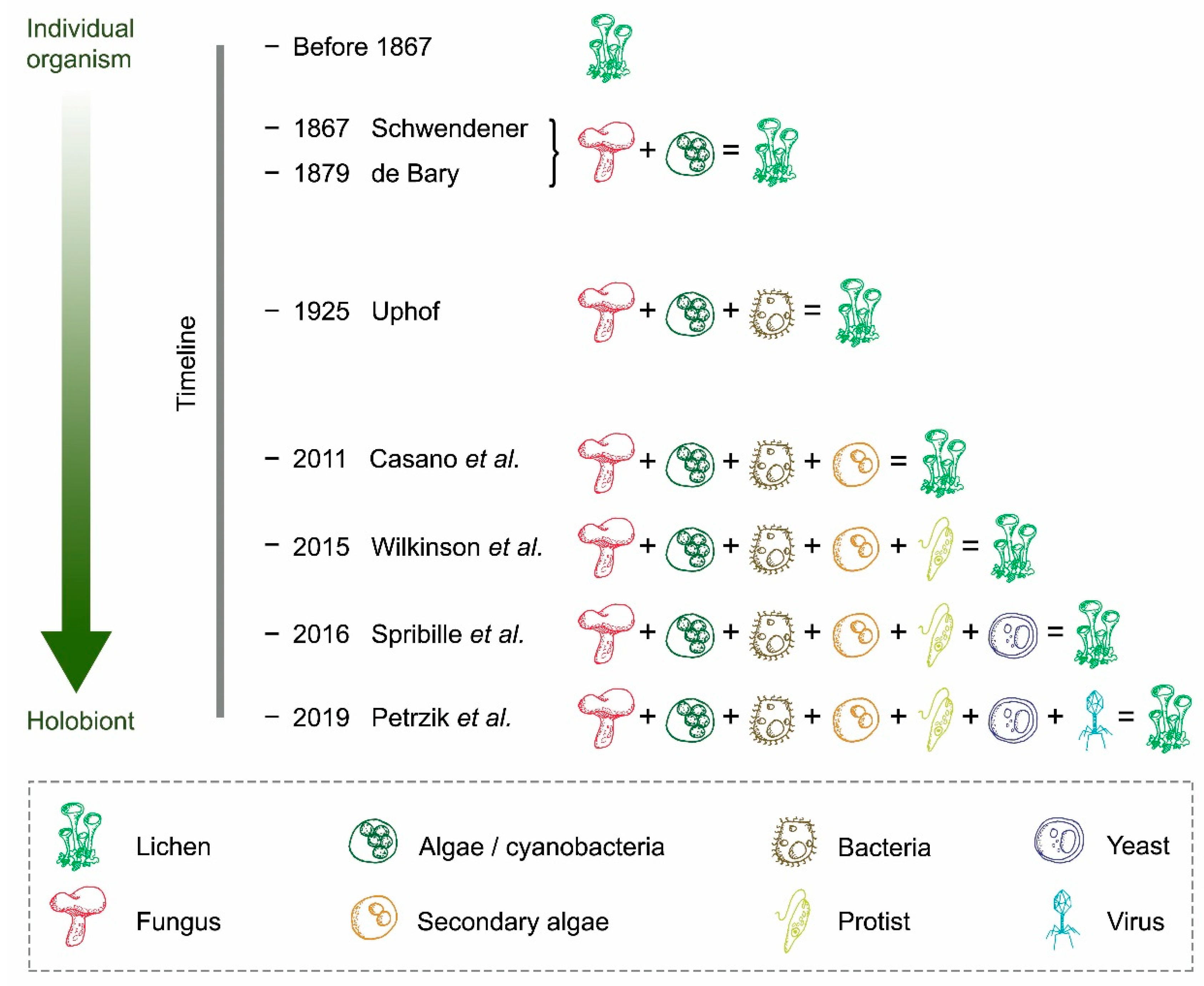
2. From a Dual Partnership to a Multi-Species Symbiotic Relationship
3. Potential Roles of Recently Discovered Partners

{kind=link}
{kind=link}
{kind=link}
4. The Increasing Complexity Surrounding the Concept of Lichen
5. Mutualism or Parasitism?
6. Conclusions and Prospects
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Agerer, R. Characterization of Ectomycorrhiza. Methods Microbiol. 1991, 23, 25–73. [Google Scholar]
- Rasmussen, H.N. Recent developments in the study of orchid mycorrhiza. Plant Soil 2002, 244, 149–163. [Google Scholar] [CrossRef]
- Hawksworth, D.L. Observations on three algicolous microfungi. Notes R. Bot. Gard. Edinburgh 1987, 44, 549–560.ill. [Google Scholar]
- Hawksworth, D.L. The variety of fungal-algal symbioses, their evolutionary significance, and the nature of lichens. Bot. J. Linn. Soc. 1988, 96, 3–20. [Google Scholar] [CrossRef]
- Kohlmeyer, J.; Demoulin, D. Parasitic and Symbiotic Fungi on Marine Algae. Bot. Mar. 1981, 24, 9–18. [Google Scholar] [CrossRef]
- Sønstebø, J.H.; Rohrlack, T. Possible implications of Chytrid parasitism for population subdivision in freshwater cyanobacteria of the genus Planktothrix. Appl. Environ. Microbiol. 2011, 77, 1344–1351. [Google Scholar] [CrossRef]
- Kohlmeyer, J.; Kohlmeyer, E. Is Ascophyllum nodosum Lichenized? Bot. Mar. 1972, 15, 109–112. [Google Scholar] [CrossRef]
- Nash, T.H.I. Lichen Biology, 2nd ed.; Cambridge University Press: Cambridge, UK, 2008; ISBN 9780521871624. [Google Scholar]
- Schwendener, S. Die Algentypen der Flechtengonidien; Universitætsbuchdruckerei von C. Schultze: Basel, Switzerland, 1869. [Google Scholar]
- Oulhen, N.; Schulz, B.J.; Carrier, T.J. English translation of Heinrich Anton de Bary’s 1878 speech, ‘Die Erscheinung der Symbiose’ (‘De la symbiose’). Symbiosis 2016, 69, 131–139. [Google Scholar] [CrossRef]
- Rikkinen, J. Molecular studies on cyanobacterial diversity in lichen symbioses. MycoKeys 2013, 6, 3–32. [Google Scholar] [CrossRef]
- Margulis, L.; Fester, R. Symbiosis as a Source of Evolutionary Innovation: Speciation and Morphogenesis; Margulis, L., Fester, R., Eds.; MIT Press: Cambridge, MA, USA, 1991; ISBN 0262132699. [Google Scholar]
- Douglas, A.E.; Werren, J.H. Holes in the hologenome: Why host-microbe symbioses are not holobionts. mBio 2016, 7, e02099-15. [Google Scholar] [CrossRef]
- Rafferty, N.E.; Caradonna, P.J.; Bronstein, J.L. Phenological shifts and the fate of mutualisms. Oikos 2015, 124, 14–21. [Google Scholar] [CrossRef]
- Chomicki, G.; Renner, S.S. Partner abundance controls mutualism stability and the pace of morphological change over geologic time. Proc. Natl. Acad. Sci. USA 2017, 114, 3951–3956. [Google Scholar] [CrossRef]
- Büdel, B.; Scheidegger, C. Thallus morphology and anatomy. In Lichen Biology; Nash, T.H.I., Ed.; Cambridge University Press: Cambridge, UK, 2008; pp. 40–68. ISBN 9780521871624. [Google Scholar]
- Lücking, R.; Dal-Forno, M.; Sikaroodi, M.; Gillevet, P.M.; Bungartz, F.; Moncada, B.; Yánez-Ayabaca, A.; Chaves, J.L.; Coca, L.F.; Lawrey, J.D. A single macrolichen constitutes hundreds of unrecognized species. Proc. Natl. Acad. Sci. USA 2014, 111, 11091–11096. [Google Scholar] [CrossRef] [PubMed]
- Honegger, R. Lichen-Forming Fungi and Their Photobionts. In Plant Relationships; Deising, H.B., Ed.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 307–333. ISBN 978-3-540-87407-2. [Google Scholar]
- Henskens, F.L.; Green, T.G.A.; Wilkins, A. Cyanolichens can have both cyanobacteria and green algae in a common layer as major contributors to photosynthesis. Ann. Bot. 2012, 110, 555–563. [Google Scholar] [CrossRef]
- Piercey-Normore, M.D.; DePriest, P.T. Algal switching among lichen symbioses. Am. J. Bot. 2001, 88, 1490–1498. [Google Scholar] [CrossRef]
- Magain, N.; Miadlikowska, J.; Goffinet, B.; Serusiaux, E.; Lutzoni, F. Macroevolution of specificity in cyanolichens of the genus Peltigera section Polydactylon (Lecanoromycetes, Ascomycota). Syst. Biol. 2017, 66, 74–99. [Google Scholar] [CrossRef]
- Wirtz, N.; Lumbsch, H.T.; Green, T.G.A.; Türk, R.; Pintado, A.; Sancho, L.; Schroeter, B. Lichen fungi have low cyanobiont selectivity in maritime Antarctica. New Phytol. 2003, 160, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Guzow-Krzemińska, B. Photobiont flexibility in the lichen Protoparmeliopsis muralis as revealed by ITS rDNA analyses. Lichenol. 2006, 38, 469–476. [Google Scholar] [CrossRef]
- Muggia, L.; Vancurova, L.; Škaloud, P.; Peksa, O.; Wedin, M.; Grube, M. The symbiotic playground of lichen thalli—A highly flexible photobiont association in rock-inhabiting lichens. FEMS Microbiol. Ecol. 2013, 85, 313–323. [Google Scholar] [CrossRef]
- Blaha, J.; Baloch, E.; Grube, M. High photobiont diversity associated with the euryoecious lichen-forming ascomycete Lecanora rupicola (Lecanoraceae, Ascomycota). Biol. J. Linn. Soc. 2006, 88, 283–293. [Google Scholar] [CrossRef]
- Yahr, R.; Vilgalys, R.; DePriest, P.T. Geographic variation in algal partners of Cladonia subtenuis (Cladoniaceae) highlights the dynamic nature of a lichen symbiosis. New Phytol. 2006, 171, 847–860. [Google Scholar] [CrossRef]
- Fernández-Mendoza, F.; Domaschke, S.; García, M.A.; Jordan, P.; Martín, M.P.; Printzen, C. Population structure of mycobionts and photobionts of the widespread lichen Cetraria aculeata. Mol. Ecol. 2011, 20, 1208–1232. [Google Scholar] [CrossRef]
- Vargas Castillo, R.; Beck, A. Photobiont selectivity and specificity in Caloplaca species in a fog-induced community in the Atacama Desert, northern Chile. Fungal Biol. 2012, 116, 665–676. [Google Scholar] [CrossRef]
- Scheidegger, C. Systematische Studien zur Krustenflechte Anzina carneonivea (Trapeliaceae, Lecanorales). Nov. Hedwigia 1985, 41, 191–218. [Google Scholar]
- Hoz, C.J.P.-D.L.; Magain, N.; Lutzoni, F.; Goward, T.; Restrepo, S.; Miadlikowska, J. Contrasting Symbiotic Patterns in Two Closely Related Lineages of Trimembered Lichens of the Genus Peltigera. Front. Microbiol. 2018, 9, 2770. [Google Scholar] [CrossRef]
- del Campo, E.M.; Casano, L.M.; Gasulla, F.; Barreno, E. Suitability of chloroplast LSU rDNA and its diverse group I introns for species recognition and phylogenetic analyses of lichen-forming Trebouxia algae. Mol. Phylogenet. Evol. 2010, 54, 437–444. [Google Scholar] [CrossRef] [PubMed]
- Moya, P.; Molins, A.; Chiva, S.; Bastida, J.; Barreno, E. Symbiotic microalgal diversity within lichenicolous lichens and crustose hosts on Iberian Peninsula gypsum biocrusts. Sci. Rep. 2020, 10, 14060. [Google Scholar] [CrossRef]
- Bačkor, M.; Peksa, O.; Škaloud, P.; Bačkorová, M. Photobiont diversity in lichens from metal-rich substrata based on ITS rDNA sequences. Ecotoxicol. Environ. Saf. 2010, 73, 603–612. [Google Scholar] [CrossRef]
- Casano, L.M.; Del Campo, E.M.; García-Breijo, F.J.; Reig-Armiñana, J.; Gasulla, F.; Del Hoyo, A.; Guéra, A.; Barreno, E. Two Trebouxia algae with different physiological performances are ever-present in lichen thalli of Ramalina farinacea. Coexistence versus Competition? Environ. Microbiol. 2011, 13, 806–818. [Google Scholar] [CrossRef]
- del Campo, E.M.; Catalá, S.; Gimeno, J.; del Hoyo, A.; Martínez-Alberola, F.; Casano, L.M.; Grube, M.; Barreno, E. The genetic structure of the cosmopolitan three-partner lichen Ramalina farinacea evidences the concerted diversification of symbionts. FEMS Microbiol. Ecol. 2013, 83, 310–323. [Google Scholar] [CrossRef]
- Uphof, J.C.T. Purple bacteria as symbionts of a lichen. Science 1925, 61, 67. [Google Scholar] [CrossRef] [PubMed]
- Henkel, P.A.; Yuzhakova, L.A. Nitrogen-fixing bacteria in lichens. Izv. Biol. Inst. Permsk. Gos. Univ. 1936, 10, 9–10. [Google Scholar]
- Iskina, R.Y. On nitrogen fixing bacteria in lichens. Isv. Biol. Inst. Permsk. 1938, 11, 133–139. [Google Scholar]
- González, I.; Ayuso-Sacido, A.; Anderson, A.; Genilloud, O. Actinomycetes isolated from lichens: Evaluation of their diversity and detection of biosynthetic gene sequences. FEMS Microbiol. Ecol. 2005, 54, 401–415. [Google Scholar] [CrossRef]
- Amann, R.I.; Ludwig, W.; Schleifer, K.-H. Phylogenetic identification and in situ detection of individual microbial cells without cultivation. Microbiol. Rev. 1995, 59, 143–169. [Google Scholar] [CrossRef]
- Rappé, M.S.; Giovannoni, S.J. The Uncultured Microbial Majority. Annu. Rev. Microbiol. 2003, 57, 369–394. [Google Scholar] [CrossRef]
- Almendras, K.; García, J.; Carú, M.; Orlando, J. Nitrogen-fixing bacteria associated with Peltigera cyanolichens and Cladonia chlorolichens. Molecules 2018, 23, 3077. [Google Scholar] [CrossRef]
- Zúñiga, C.; Leiva, D.; Carú, M.; Orlando, J. Substrates of Peltigera Lichens as a Potential Source of Cyanobionts. Microb. Ecol. 2017, 74, 561–569. [Google Scholar] [CrossRef]
- Graham, L.E.; Trest, M.T.; Will-Wolf, S.; Miicke, N.S.; Atonio, L.M.; Piotrowski, M.J.; Knack, J.J. Microscopic and metagenomic analyses of peltigera ponojensis (Peltigerales, ascomycota). Int. J. Plant Sci. 2018, 179, 241–255. [Google Scholar] [CrossRef]
- Grube, M.; Cardinale, M.; De Castro, J.V., Jr.; Müller, H.; Berg, G. Species-specific structural and functional diversity of bacterial communities in lichen symbioses. ISME J. 2009, 3, 1105–1115. [Google Scholar] [CrossRef] [PubMed]
- Hodkinson, B.P.; Lutzoni, F. A microbiotic survey of lichen-associated bacteria reveals a new lineage from the Rhizobiales. Symbiosis 2009, 49, 163–180. [Google Scholar] [CrossRef]
- Bates, S.T.; Cropsey, G.W.G.; Caporaso, J.G.; Knight, R.; Fierer, N. Bacterial communities associated with the lichen symbiosis. Appl. Environ. Microbiol. 2011, 77, 1309–1314. [Google Scholar] [CrossRef] [PubMed]
- Krassowski, M.; Das, V.; Sahu, S.K.; Misra, B.B. State of the Field in Multi-Omics Research: From Computational Needs to Data Mining and Sharing. Front. Genet. 2020, 11, 610798. [Google Scholar] [CrossRef] [PubMed]
- Grube, M.; Cernava, T.; Soh, J.; Fuchs, S.; Aschenbrenner, I.; Lassek, C.; Wegner, U.; Becher, D.; Riedel, K.; Sensen, C.W.; et al. Exploring functional contexts of symbiotic sustain within lichen-associated bacteria by comparative omics. ISME J. 2015, 9, 412–424. [Google Scholar] [CrossRef] [PubMed]
- Cernava, T.; Erlacher, A.; Aschenbrenner, I.A.; Krug, L.; Lassek, C.; Riedel, K.; Grube, M.; Berg, G. Deciphering functional diversification within the lichen microbiota by meta-omics. Microbiome 2017, 5, 82. [Google Scholar] [CrossRef]
- Lendemer, J.C.; Keepers, K.G.; Tripp, E.A.; Pogoda, C.S.; McCain, C.M.; Kane, N.C. A taxonomically broad metagenomic survey of 339 species spanning 57 families suggests cystobasidiomycete yeasts are not ubiquitous across all lichens. Am. J. Bot. 2019, 106, 1090–1095. [Google Scholar] [CrossRef]
- Kono, M.; Kon, Y.; Ohmura, Y.; Satta, Y.; Terai, Y. In vitro resynthesis of lichenization reveals the genetic background of symbiosis-specific fungal-algal interaction in Usnea hakonensis. BMC Genom. 2020, 21, 671. [Google Scholar] [CrossRef]
- Tuovinen, V.; Ekman, S.; Thor, G.; Vanderpool, D.; Spribille, T.; Johannesson, H. Two Basidiomycete Fungi in the Cortex of Wolf Lichens. Curr. Biol. 2019, 29, 476–483.e5. [Google Scholar] [CrossRef]
- Noh, H.-J.; Park, Y.; Hong, S.G.; Lee, Y.M. Diversity and Physiological Characteristics of Antarctic Lichens-Associated Bacteria. Microorganisms 2021, 9, 607. [Google Scholar] [CrossRef] [PubMed]
- Cardinale, M.; Vieira De Castro Jr., J.; Müller, H.; Berg, G.; Grube, M. In situ analysis of the bacterial community associated with the reindeer lichen Cladonia arbuscula reveals predominance of Alphaproteobacteria. FEMS Microbiol. Ecol. 2008, 66, 63–71. [Google Scholar] [CrossRef]
- Aschenbrenner, I.A.; Cernava, T.; Berg, G.; Grube, M. Understanding microbial multi-species symbioses. Front. Microbiol. 2016, 7, 180. [Google Scholar] [CrossRef]
- Spribille, T.; Tagirdzhanova, G.; Goyette, S.; Tuovinen, V.; Case, R.; Zandberg, W.F. 3D biofilms: In search of the polysaccharides holding together lichen symbioses. FEMS Microbiol. Lett. 2020, 367, fnaa023. [Google Scholar] [CrossRef]
- Spribille, T.; Tuovinen, V.; Resl, P.; Vanderpool, D.; Wolinski, H.; Aime, M.C.; Schneider, K.; Stabentheiner, E.; Toome-Heller, M.; Thor, G.; et al. Basidiomycete yeasts in the cortex of ascomycete macrolichens. Science 2016, 353, 488–492. [Google Scholar] [CrossRef]
- Černajová, I.; Škaloud, P. The first survey of Cystobasidiomycete yeasts in the lichen genus Cladonia; with the description of Lichenozyma pisutiana gen. nov., sp. nov. Fungal Biol. 2019, 123, 625–637. [Google Scholar] [CrossRef]
- Millanes, A.M.; Diederich, P.; Wedin, M. Cyphobasidium gen. nov., a new lichen-inhabiting lineage in the Cystobasidiomycetes (Pucciniomycotina, Basidiomycota, Fungi). Fungal Biol. 2016, 120, 1468–1477. [Google Scholar] [CrossRef]
- Oberwinkler, F. Yeasts in Pucciniomycotina. Mycol. Prog. 2017, 16, 831–856. [Google Scholar] [CrossRef]
- Wilkinson, D.M.; Creevy, A.L.; Kalu, C.L.; Schwartzman, D.W. Are heterotrophic and silica-rich eukaryotic microbes an important part of the lichen symbiosis? Mycology 2015, 6, 4–7. [Google Scholar] [CrossRef]
- Petrzik, K.; Koloniuk, I.; Sehadová, H.; Sarkisova, T. Chrysoviruses inhabited symbiotic fungi of lichens. Viruses 2019, 11, 1120. [Google Scholar] [CrossRef]
- Simon, J.-C.; Marchesi, J.R.; Mougel, C.; Selosse, M.-A. Host-microbiota interactions: From holobiont theory to analysis. Microbiome 2019, 7, 5. [Google Scholar] [CrossRef] [PubMed]
- Hawksworth, D.L.; Grube, M. Lichens redefined as complex ecosystems. New Phytol. 2020, 227, 1281–1283. [Google Scholar] [CrossRef] [PubMed]
- Honegger, R. The symbiotic phenotype of lichen-forming ascomycetes and their endo- and epibionts. In Fungal Associations, 2nd ed.; Springer Berlin Heidelberg; Institute of Plant Biology, University of Zürich: Zürich, Switzerland, 2012; Volume 9, pp. 287–339. ISBN 9783642308260. [Google Scholar]
- Grimm, M.; Grube, M.; Schiefelbein, U.; Zühlke, D.; Bernhardt, J.; Riedel, K. The Lichens’ Microbiota, Still a Mystery? Front. Microbiol. 2021, 12, 714. [Google Scholar] [CrossRef] [PubMed]
- Ahmadjian, V.; Jacobs, J.B. Relationship between fungus and alga in the lichen Cladonia cristatella Tuck. Nature 1981, 289, 169–172. [Google Scholar] [CrossRef]
- Athukorala, S.N.P.; Huebner, E.; Piercey-Normore, M.D. Identification and comparison of the 3 early stages of resynthesis for the lichen Cladonia rangiferina. Can. J. Microbiol. 2014, 60, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Honegger, R. Morphogenesis. In Lichen Biology; Nash, T.H., III, Ed.; Cambridge University Press: Cambridge, UK, 2008; pp. 69–93. ISBN 9780521871624. [Google Scholar]
- Meeßen, J.; Ott, S. Recognition mechanisms during the pre-contact state of lichens: I. Mycobiont-photobiont interactions of the mycobiont of Fulgensia bracteata. Symbiosis 2013, 59, 121–130. [Google Scholar] [CrossRef]
- Joneson, S.; Armaleo, D.; Lutzoni, F. Fungal and algal gene expression in early developmental stages of lichen-symbiosis. Mycologia 2011, 103, 291–306. [Google Scholar] [CrossRef] [PubMed]
- Ahmadjian, V.; Jacobs, J.B.; Russell, L.A. Scanning Electron Microscope Study of Early Lichen Synthesis. Science 1978, 200, 1062–1064. [Google Scholar] [CrossRef] [PubMed]
- Galun, M. Lichenization. In CRC Handbook of Lichenology; Galun, M., Ed.; CRC Press: Boca Raton, FL, USA, 1988; Volume II, pp. 153–169. [Google Scholar]
- Armaleo, D. Experimental Microbiology of Lichens—Mycelia Fragmentation, a Novel Growth Chamber, and the Origins of Thallus Differentiation. Symbiosis 1991, 11, 163–177. [Google Scholar]
- Joneson, S.; Lutzoni, F. Compatibility and thigmotropism in the lichen symbiosis: A reappraisal. Symbiosis 2009, 47, 109–115. [Google Scholar] [CrossRef]
- Yoshimura, I.; Kurokawa, T.; Yamamoto, Y.; Kinoshita, Y. Development of Lichen Thalli in Vitro. Bryologist 1993, 96, 412–421. [Google Scholar] [CrossRef]
- Kon, Y.; Kashiwadani, H.; Masada, M.; Tamura, G. Artificial Syntheses of Mycobionts of Usnea confusa ssp. kitamiensis and Usnea orientalis with Their Natural and Non-Natural Phycobiont. J. Jpn. Bot. 1993, 68, 129–137. [Google Scholar]
- Stocker-Wörgötter, E. Experimental studies of the lichen symbiosis: DNA-analyses, differentiation and secondary chemistry of selected mycobionts, artificial resynthesis of two- and tripartite symbioses. Symbiosis 2001, 30, 207–227. [Google Scholar]
- Trembley, M.L.; Ringli, C.; Honegger, R. Morphological and molecular analysis of early stages in the resynthesis of the lichen Baeomyces rufus. Mycol. Res. 2002, 106, 768–776. [Google Scholar] [CrossRef]
- Guzow-Krzemińska, B.; Stocker-Wörgötter, E. In vitro culturing and resynthesis of the mycobiont Protoparmeliopsis muralis with algal bionts. Lichenologist 2013, 45, 65–76. [Google Scholar] [CrossRef]
- Ahmadjian, V.; Russell, L.A.; Hildreth, K.C. Artificial Reestablishment of Lichens. I. Morphological Interactions Between the Phycobionts of Different Lichens and the Mycobionts Cladonia Cristatella and Lecanora Chrysoleuca. Mycologia 1980, 72, 73–89. [Google Scholar] [CrossRef]
- Bubrick, P.; Galun, M. Spore to spore resynthesis of Xanthoria Parietina. Lichenologist 1986, 18, 47–49. [Google Scholar] [CrossRef]
- Culberson, C.F.; Culberson, W.L.; Johnson, A. Genetic and environmental effects of growth and production of secondary compounds in Cladonia cristatella. Biochem. Syst. Ecol. 1983, 11, 77–84. [Google Scholar] [CrossRef]
- Stocker-Wörgötter, E.; Türk, R. Artificial cultures of the cyanobacterial lichenPeltigera didactyla (Peltigeraceae) in the natural environment. Plant Syst. Evol. 1989, 165, 39–48. [Google Scholar] [CrossRef]
- Stocker-Wörgötter, E.; Türk, R. The resynthesis of thalli of Dermatocarpon miniatum under laboratory conditions. Symbiosis 1989, 7, 37–50. [Google Scholar]
- Athukorala, S.N.P.; Piercey-Normore, M.D. Recognition-and defense-related gene expression at 3 resynthesis stages in lichen symbionts. Can. J. Microbiol. 2014, 61, 1–12. [Google Scholar] [CrossRef]
- Insarova, I.D.; Blagoveshchenskaya, E.Y. Lichen symbiosis: Search and recognition of partners. Biol. Bull. 2016, 43, 408–418. [Google Scholar] [CrossRef]
- Gaßmann, A.; Ott, S. Growth strategy and the gradual symbiotic interactions of the lichen Ochrolechia frigida. Plant Biol. 2000, 2, 368–378. [Google Scholar] [CrossRef]
- Meeßen, J.; Eppenstein, S.; Ott, S. Recognition mechanisms during the pre-contact state of lichens: II. Influence of algal exudates and ribitol on the response of the mycobiont of Fulgensia bracteata. Symbiosis 2013, 59, 131–143. [Google Scholar] [CrossRef]
- Díaz, E.M.; Vicente-Manzanares, M.; Sacristan, M.; Vicente, C.; Legaz, M.-E. Fungal lectin of Peltigera canina induces chemotropism of compatible Nostoc cells by constriction-relaxation pulses of cyanobiont cytoskeleton. Plant Signal. Behav. 2011, 6, 1525–1536. [Google Scholar] [CrossRef] [PubMed]
- Munzi, S.; Gouveia, C.; Cruz, C.; Branquinho, C.; Coelho, A.V. Proteomic analysis contributes to unveil the mechanisms of nitrogen tolerance in the lichen Xanthoria parietina. Not. Della Soc. Lichenol. Ital. 2018, 31, 23. [Google Scholar]
- Crosino, A.; Moscato, E.; Blangetti, M.; Carotenuto, G.; Spina, F.; Bordignon, S.; Puech-Pagès, V.; Anfossi, L.; Volpe, V.; Prandi, C.; et al. Extraction of short chain chitooligosaccharides from fungal biomass and their use as promoters of arbuscular mycorrhizal symbiosis. Sci. Rep. 2021, 11, 3798. [Google Scholar] [CrossRef]
- Volpe, V.; Carotenuto, G.; Berzero, C.; Cagnina, L.; Puech-Pagès, V.; Genre, A. Short chain chito-oligosaccharides promote arbuscular mycorrhizal colonization in Medicago truncatula. Carbohydr. Polym. 2020, 229, 115505. [Google Scholar] [CrossRef]
- Feng, F.; Sun, J.; Radhakrishnan, G.V.; Lee, T.; Bozsóki, Z.; Fort, S.; Gavrin, A.; Gysel, K.; Thygesen, M.B.; Andersen, K.R.; et al. A combination of chitooligosaccharide and lipochitooligosaccharide recognition promotes arbuscular mycorrhizal associations in Medicago truncatula. Nat. Commun. 2019, 10, 5047. [Google Scholar] [CrossRef]
- Diédhiou, I.; Diouf, D. Transcription factors network in root endosymbiosis establishment and development. World J. Microbiol. Biotechnol. 2018, 34, 37. [Google Scholar] [CrossRef]
- Omoarelojie, L.O.; Van Staden, J. Plant-endophytic fungi interactions: A strigolactone perspective. S. Afr. J. Bot. 2020, 134, 280–284. [Google Scholar] [CrossRef]
- Peláez-Vico, M.A.; Bernabéu-Roda, L.; Kohlen, W.; Soto, M.J.; López-Ráez, J.A. Strigolactones in the Rhizobium-legume symbiosis: Stimulatory effect on bacterial surface motility and down-regulation of their levels in nodulated plants. Plant Sci. 2016, 245, 119–127. [Google Scholar] [CrossRef]
- Maclean, A.M.; Bravo, A.; Harrison, M.J. Plant signaling and metabolic pathways enabling arbuscular mycorrhizal symbiosis. Plant Cell 2017, 29, 2319–2335. [Google Scholar] [CrossRef] [PubMed]
- Kuga, Y.; Schläppi, K.; Reinhardt, D. From Imaging to Functional Traits in Interactions between Roots and Microbes. In Methods in Rhizosphere Biology Research; Reinhardt, D., Sharma, A.K., Eds.; Springer: Singapore, 2019; pp. 227–239. ISBN 978-981-13-5767-1. [Google Scholar]
- Liu, C.-W.; Breakspear, A.; Stacey, N.; Findlay, K.; Nakashima, J.; Ramakrishnan, K.; Liu, M.; Xie, F.; Endre, G.; de Carvalho-Niebel, F.; et al. A protein complex required for polar growth of rhizobial infection threads. Nat. Commun. 2019, 10, 2848. [Google Scholar] [CrossRef] [PubMed]
- Radhakrishnan, G.V.; Keller, J.; Rich, M.K.; Vernié, T.; Mbadinga Mbadinga, D.L.; Vigneron, N.; Cottret, L.; Clemente, H.S.; Libourel, C.; Cheema, J.; et al. An ancestral signalling pathway is conserved in intracellular symbioses-forming plant lineages. Nat. Plants 2020, 6, 280–289. [Google Scholar] [CrossRef] [PubMed]
- Boschiero, C.; Dai, X.; Lundquist, P.K.; Roy, S.; de Bang, T.C.; Zhang, S.; Zhuang, Z.; Torres-Jerez, I.; Udvardi, M.K.; Scheible, W.-R.; et al. MtSSPDB: The Medicago truncatula Small Secreted Peptide Database. Plant Physiol. 2020, 183, 399–413. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morillas, L.; Roales, J.; Cruz, C.; Munzi, S. Lichen as Multipartner Symbiotic Relationships. Encyclopedia 2022, 2, 1421-1431. https://doi.org/10.3390/encyclopedia2030096
Morillas L, Roales J, Cruz C, Munzi S. Lichen as Multipartner Symbiotic Relationships. Encyclopedia. 2022; 2(3):1421-1431. https://doi.org/10.3390/encyclopedia2030096
Chicago/Turabian StyleMorillas, Lourdes, Javier Roales, Cristina Cruz, and Silvana Munzi. 2022. "Lichen as Multipartner Symbiotic Relationships" Encyclopedia 2, no. 3: 1421-1431. https://doi.org/10.3390/encyclopedia2030096
APA StyleMorillas, L., Roales, J., Cruz, C., & Munzi, S. (2022). Lichen as Multipartner Symbiotic Relationships. Encyclopedia, 2(3), 1421-1431. https://doi.org/10.3390/encyclopedia2030096