

Antifungals and Drug Resistance

 ,
,  , , and
, , and 
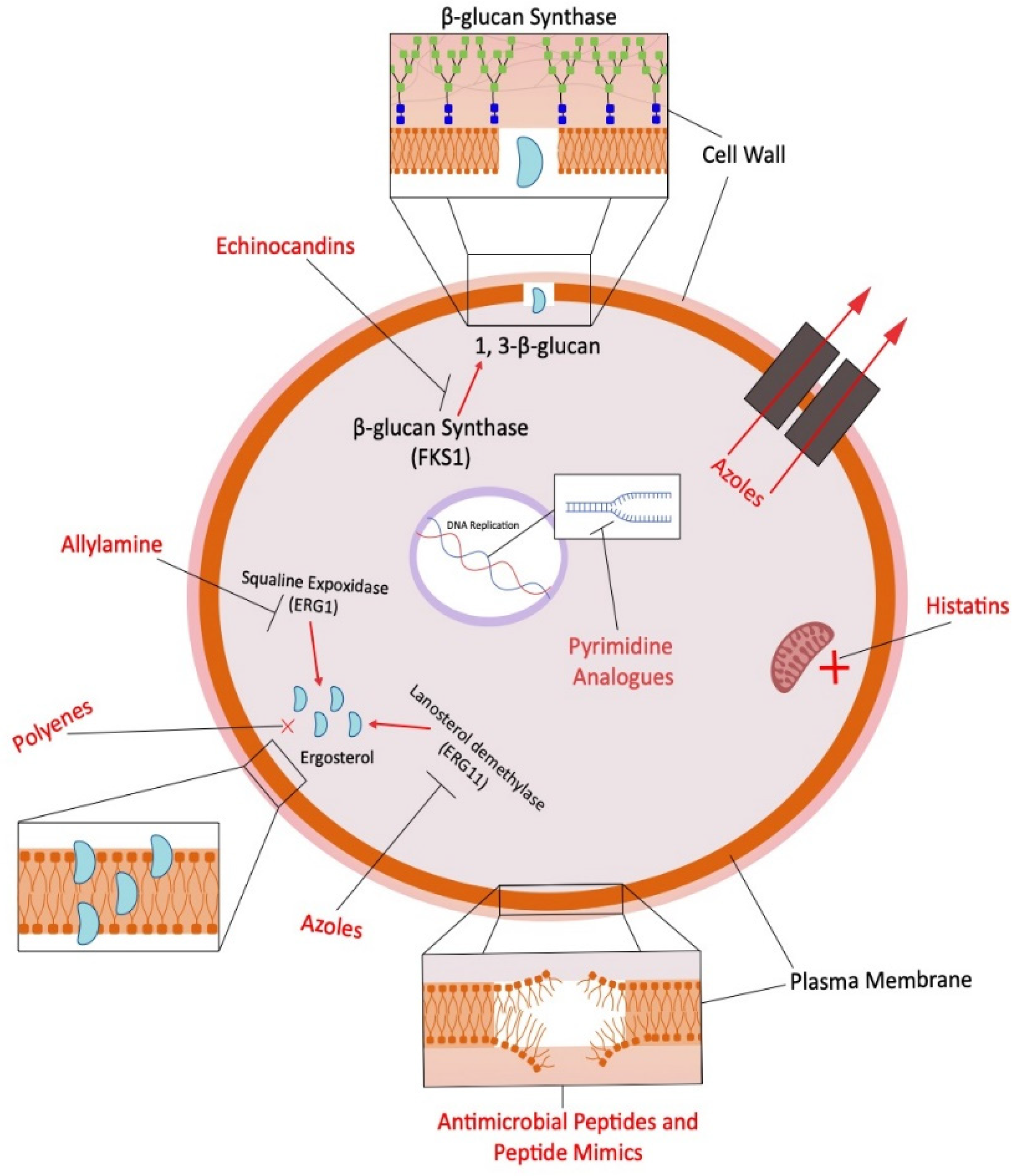{kind=link}
Definition
1. Introduction to Fungal Infection
1.1. Classification of Fungal Infections (Mycosis)
- Superficial infections
- Subcutaneous infections
- Systemic infections
1.1.1. Superficial Infections
1.1.2. Subcutaneous Fungal Infections
1.1.3. Systemic Fungal Infections
2. Overview of Antifungal Drugs and Their Mechanism of Action
2.1. Azoles
2.2. Polyenes
2.3. Pyrimidine Analogs
2.4. Allylamine, Thiocarbamates and Morpholines
2.5. Echinocandins
3. Pharmacological and Toxicological Study of Antifungal Drugs
4. Current Antifungal Therapies
4.1. Polyenes
4.2. Azoles
4.3. Echinocandins
4.4. Use of Drug Combinations
4.5. Novel Antifungal Agents
5. Antifungal Resistance
5.1. Overview of Causes of Antifungal Resistance and How to Tackle the Problem
- Antifungal medication misuse: When doses are skipped, therapy is stopped too soon, or the dose prescribed is too low, a fungus becomes more adept at fending off the effects of the antifungal drug [71].
- Fungicide use: Fungicidal drugs are often used in agriculture, as a method to protect crops from rotting [72]. As fungi have been exposed to more fungicides under these conditions, people working closely with fungicide-treated crops may become more susceptible to antifungal-resistant fungus diseases.
- Spontaneous resistance: This occurs when a fungus ceases to respond to a previously effective medication for no apparent cause.
- Transmitted Resistance: One can transmit a contagious drug-resistant fungal illness. Even though the recipient has had never used an antifungal drug, that individual now has an illness that will not react to that particular antifungal drug [73].
5.2. Types of Resistant Fungi
- Targeted protein overexpression (azoles);
- Mutations induced in targeted proteins (azoles and echinocandins);
- Enhancing the production of efflux pumps and/or increasing their insertion into cell membranes (observed in azoles); and
- Limited access to the target, as in the case of ergosterol sequestration (observed in polyenes).
- Overexpression of ERG11
- Mutations in ERG11
- Expression of efflux pumps.
6. Resistance Mechanisms of Antifungal Drugs
6.1. Assessing Host and Environmental Factors Influencing Antifungal Resistance
6.2. Molecular Mechanisms of Antifungal Resistance
Regulation of Drug Resistance Genes
6.3. Resistance to Azoles
6.3.1. Over-Expression of Membrane Transporters
- ABC-transporters
- MFS-transporters
6.3.2. Erg11 (Cyp51A) Substitutions: Ergosterol Biosynthetic Enzymes Alterations
6.3.3. Alterations in Ergosterol Biosynthetic Enzymes
6.3.4. Diffusion of Drugs
6.4. Resistance to Echinocandins
7. Multidrug Resistant (MDR) Fungi
8. The Need for Development of New Antifungal Drugs
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Gnat, S.; Łagowski, D.; Nowakiewicz, A.; Dyląg, M. A global view on fungal infections in humans and animals: Infections caused by dimorphic fungi and dermatophytoses. J. Appl. Microbiol. 2021, 131, 2688–2704. [Google Scholar] [CrossRef] [PubMed]
- Kainz, K.; Bauer, M.A.; Madeo, F.; Carmona-Gutierrez, D. Fungal infections in humans: The silent crisis. Microb. Cell 2020, 7, 143–145. [Google Scholar] [CrossRef] [PubMed]
- Nnadi, N.E.; Carter, D.A. Climate change and the emergence of fungal pathogens. PLoS Pathog. 2021, 17, e1009503–e1009509. [Google Scholar] [CrossRef]
- Chen, S.C.A.; Lewis, R.E.; Kontoyiannis, D.P. Direct effects of non-antifungal agents used in cancer chemotherapy and organ transplantation on the development and virulence of Candida and Aspergillus species. Virulence 2011, 2, 280–295. [Google Scholar] [CrossRef] [PubMed]
- Rautemaa-Richardson, R.; Richardson, M.D. Systemic fungal infections. Medicine 2017, 45, 757–762. [Google Scholar] [CrossRef]
- Ruiz-Camps, I.; Cuenca-Estrella, M. Antifungals for systemic use. Enferm. Infecc. Microbiol. Clínica 2009, 27, 353–362. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira Santos, G.C.; Vasconcelos, C.C.; Lopes, A.J.; de Sousa Cartágenes, M.D.S.; Filho, A.K.; do Nascimento, F.R.; Ramos, R.M.; Pires, E.R.; de Andrade, M.S.; Rocha, F.M.; et al. Candida infections and therapeutic strategies: Mechanisms of action for traditional and alternative agents. Front. Microbiol. 2018, 9, 1351–1374. [Google Scholar] [CrossRef]
- Ahmad, S.; Joseph, L.; Parker, J.E.; Asadzadeh, M.; Kelly, S.L.; Meis, J.F.; Khan, Z. ERG6 and ERG2 are major targets conferring reduced susceptibility to amphotericin B in clinical Candida glabrata isolates in Kuwait. Antimicrob. Agents Chemother. 2019, 63, e01900–e01918. [Google Scholar] [CrossRef]
- Cuenca-Estrella, M. Antifungal drug resistance mechanisms in pathogenic fungi: From bench to bedside. Clin. Microbiol. Infect. 2014, 20, 54–59. [Google Scholar] [CrossRef]
- Carolus, H.; Pierson, S.; Lagrou, K.; Van Dijck, P. Amphotericin B and other polyenes—Discovery, clinical use, mode of action and drug resistance. J. Fungi 2020, 6, 321. [Google Scholar] [CrossRef]
- Sharma, B.; Nonzom, S. Superficial mycoses, a matter of concern: Global and Indian scenario-an updated analysis. Mycoses 2021, 64, 890–908. [Google Scholar] [CrossRef] [PubMed]
- Havlickova, B.; Czaika, V.A.; Friedrich, M. Epidemiological trends in skin mycoses worldwide. Mycoses 2008, 51, 2–15. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.K.; Ryder, J.E.; Nicol, K.; Cooper, E.A. Superficial fungal infections: An update on pityriasis versicolor, seborrheic dermatitis, tinea capitis, and onychomycosis. Clin. Dermatol. 2003, 21, 417–425. [Google Scholar] [CrossRef] [PubMed]
- Bonifaz, A.; Vázquez-González, D.; Perusquía-Ortiz, A.M. Subcutaneous mycoses: Chromoblastomycosis, sporotrichosis and mycetoma. JDDG J. Dtsch. Dermatol. Ges. 2010, 8, 619–628. [Google Scholar] [CrossRef]
- McGinnis, M.R. Chromoblastomycosis and phaeohyphomycosis: New concepts, diagnosis, and mycology. J. Am. Acad. Dermatol. 1983, 8, 1–16. [Google Scholar] [CrossRef]
- Walsh, T.J.; Dixon, D.M. Spectrum of mycoses. In Medical Microbiology, 4th ed.; Baron, S., Ed.; University of Texas at Galvelston: Galveston, TX, USA, 1996; pp. 919–925. [Google Scholar]
- Patterson, J.W. Mycosis and algal infections. In Weedon’s Skin Pathology, 4th ed.; Elsevier Health Sciences: Amsterdam, The Netherlands, 2014; pp. 683–716. [Google Scholar]
- Georgiev, V.S. Membrane transporters and antifungal drug resistance. Curr. Drug Targets 2000, 1, 261–284. [Google Scholar] [CrossRef] [PubMed]
- Castelli, M.V.; Butassi, E.; Monteiro, M.C.; Svetaz, L.A.; Vicente, F.; Zacchino, S.A. Novel antifungal agents: A patent review (2011–present). Exp. Opin. Ther. Pat. 2014, 24, 323–338. [Google Scholar] [CrossRef]
- Sanglard, D.; Coste, A.T. Activity of isavuconazole and other azoles against Candida clinical isolates and yeast model systems with known azole resistance mechanisms. Antimicrob. Agents Chemother. 2016, 60, 229–238. [Google Scholar] [CrossRef]
- Cowen, L.E.; Sanglard, D.; Howard, S.J.; Rogers, P.D.; Perlin, D.S. Mechanisms of antifungal drug resistance. Cold Spring Harb. Perspect. Med. 2015, 5, a019752–a019785. [Google Scholar] [CrossRef]
- Vandeputte, P.; Ferrari, S.; Coste, A.T. Antifungal resistance and new strategies to control fungal infections. Int. J. Microbiol. 2012, 2012, 713687–713713. [Google Scholar] [CrossRef]
- Sanglard, D.; Coste, A.; Ferrari, S. Antifungal drug resistance mechanisms in fungal pathogens from the perspective of transcriptional gene regulation. FEMS Yeast Res. 2009, 9, 1029–1050. [Google Scholar] [CrossRef] [PubMed]
- Lemke, A.; Kiderlen, A.F.; Kayser, O. Amphotericin, B. Appl. Microbiol. Biotechnol. 2005, 68, 151–162. [Google Scholar] [CrossRef] [PubMed]
- Polak, A. Mode of action studies. In Chemotherapy of Fungal Diseases; Ryley, J.F., Ed.; Springer: Berlin/Heidelberg, Germany, 1990; pp. 153–182. [Google Scholar]
- Mercer, E.I. Morpholine antifungals and their mode of action. Biochem. Soc. Transact. 1991, 19, 788–793. [Google Scholar] [CrossRef] [PubMed]
- Haria, M.; Bryson, H.M. Amorolfine. Drugs 1995, 49, 103–120. [Google Scholar] [CrossRef]
- Shapiro, R.S.; Robbins, N.; Cowen, L.E. Regulatory circuitry governing fungal development, drug resistance, and disease. Microbiol. Mol. Biol. Rev. 2011, 75, 213–267. [Google Scholar] [CrossRef] [PubMed]
- Ghannoum, M.A.; Rice, L.B. Antifungal agents: Mode of action, mechanisms of resistance, and correlation of these mechanisms with bacterial resistance. Clin. Microbiol. Rev. 1999, 12, 501–517. [Google Scholar] [CrossRef]
- Perlin, D.S. Current perspectives on echinocandin class drugs. Future Microbiol. 2011, 6, 441–457. [Google Scholar] [CrossRef]
- Cavassin, F.B.; Baú-Carneiro, J.L.; Vilas-Boas, R.R.; Queiroz-Telles, F. Sixty years of Amphotericin B: An overview of the main antifungal agent used to treat invasive fungal infections. Infect. Dis. Ther. 2021, 10, 115–147. [Google Scholar] [CrossRef]
- Stone, N.R.H.; Bicanic, T.; Salim, R.; Hope, W. Liposomal amphotericin B (AmBisome®): A review of the pharmakinetics, pharmacodynamics, clinical experience and future directions. Drugs 2016, 76, 485–500. [Google Scholar] [CrossRef]
- Sawant, B.; Khan, T. Recent advances in delivery of antifungal agents for therapeutic management of candidiasis. Biomed. Pharmacother. 2017, 96, 1478–1490. [Google Scholar] [CrossRef]
- Kinsky, S.C. Polyene antibiotics. In Antibiotics; Springer: New York, NY, USA, 1967; pp. 122–141. [Google Scholar]
- Delattin, N.; Cammue, B.P.; Thevissen, K. Reactive oxygen species-inducing antifungal agents and their activity against fungal biofilms. Future Med. Chem. 2014, 6, 77–90. [Google Scholar] [CrossRef] [PubMed]
- Ellis, D. Amphotericin B: Spectrum and resistance. J. Antimicrob. Chemother. 2002, 49, 7–10. [Google Scholar] [CrossRef] [PubMed]
- Parente-Rocha, J.A.; Bailão, A.M.; Amaral, A.C.; Taborda, C.P.; Paccez, J.D.; Borges, C.L.; Pereira, M. Antifungal resistance, metabolic routes as drug targets, and new antifungal agents: An overview about endemic dimorphic fungi. Mediat. Inflamm. 2017, 2017, 9870679–9870695. [Google Scholar] [CrossRef]
- Fujita, K.I.; Tatsumi, M.; Ogita, A.; Kubo, I.; Tanaka, T. Anethole induces apoptotic cell death accompanied by reactive oxygen species production and DNA fragmentation in Aspergillus fumigatus and Saccharomyces cerevisiae. FEBS J. 2014, 281, 1304–1313. [Google Scholar] [CrossRef]
- Grudzinski, W.; Sagan, J.; Welc, R.; Luchowski, R.; Gruszecki, W.I. Molecular organization, localization and orientation of antifungal antibiotic amphotericin B in a single lipid bilayer. Sci. Rep. 2016, 6, 32780. [Google Scholar] [CrossRef] [PubMed]
- Almeida, B.; Silva, A.; Mesquita, A.; Sampaio-Marques, B.; Rodrigues, F.; Ludovico, P. Drug-induced apoptosis in yeast. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2008, 1783, 1436–1448. [Google Scholar] [CrossRef] [PubMed]
- Perfect, J.R. The antifungal pipeline: A reality check. Nat. Rev. Drug Dis. 2017, 16, 603–616. [Google Scholar] [CrossRef] [PubMed]
- Larkin, E.L.; Ali, A.A.L.; Swindell, K. History of antifungals. In Antifungal Therapy, 2nd ed.; Ghannoum, M.A., Perfect, J.R., Eds.; CRC Press: Boca Raton, FL, USA, 2019; pp. 1–10. [Google Scholar]
- Campoy, S.; Adrio, J.L. Antifungals. Biochem. Pharmacol. 2017, 133, 86–96. [Google Scholar] [CrossRef]
- Shukla, P.K.; Singh, P.; Yadav, R.K.; Pandey, S.; Bhunia, S.S. Past, present, and future of antifungal drug development. Commun. Dis. Dev. World 2016, 29, 125–167. [Google Scholar]
- Perlin, D.S. Mechanisms of echinocandin antifungal drug resistance. Ann. N. Y. Acad. Sci. 2015, 1354, 1–11. [Google Scholar] [CrossRef]
- Spitzer, M.; Robbins, N.; Wright, G.D. Combinatorial strategies for combating invasive fungal infections. Virulence 2017, 8, 169–185. [Google Scholar] [CrossRef] [PubMed]
- Garbati, M.A.; Alasmari, F.A.; Al-Tannir, M.A.; Tleyjeh, I.M. The role of combination antifungal therapy in the treatment of invasive aspergillosis: A systematic review. Int. J. Infect. Dis. 2012, 16, e76–e81. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kim, J.H.; Cheng, L.W.; Chan, K.L.; Tam, C.C.; Mahoney, N.; Friedman, M.; Shilman, M.M.; Land, K.M. Antifungal drug repurposing. Antibiotics 2020, 9, 812. [Google Scholar] [CrossRef] [PubMed]
- Das, R.; Kotra, K.; Singh, P.; Loh, B.; Leptihn, S.; Bajpai, U. Alternative treatment strategies for secondary bacterial and fungal infections associated with COVID-19. Infect. Dis. Ther. 2021, 11, 53–78. [Google Scholar] [CrossRef]
- Fiołka, M.J.; Czaplewska, P.; Wójck-Mieszawska, S.; Lewańdowska, A.; Lewtak, K.; Sofinska-Chmiel, W.; Buchwald, T. Metabolic, structural, and proteomic changes in Candida albicans cells induced by the protein-carbohydrate fraction of Dendrobaena veneta coelomic fluid. Sci. Rep. 2021, 11, 16711–16729. [Google Scholar] [CrossRef]
- Hwang, J.H.; Hwang, I.S.; Liu, Q.H.; Woo, E.R.; Lee, D.G. (+)-Medioresinol leads to intracellular RPS accumulation and mitochondria-mediated apoptotic cell death in Candida albicans. Biochemie 2012, 94, 1784–1793. [Google Scholar] [CrossRef]
- Liu, X.; Ma, Z.; Zhang, J.; Yang, L. Antifungal compounds against Candida infection from tradition Chinese medicine. BioMed Res. Int. 2017, 2017, 4614183–4614195. [Google Scholar] [CrossRef]
- Lemar, K.M.; Passa, O.; Aon, M.A.; Cortassa, S.; Müller, C.T.; Plummer, S.; O’Rourke, B.; Lloyd, D. Allyl alcohol and garlic (Allium sativum) extract produce oxidative stress in Candida albicans. Microbiology 2005, 151, 3257–3265. [Google Scholar] [CrossRef]
- Yang, S.; Fu, Y.; Wu, X.; Zhou, Z.; Xu, J.; Zeng, X.; Kuang, N.; Zeng, Y. Baicalin prevents Candida albicans infections via increasing its apoptosis rate. Biochem. Biophys. Res. Comm. 2014, 451, 36–41. [Google Scholar] [CrossRef]
- Sharma, M.; Manoharlal, M.R.; Negi, A.S.; Prasad, R. Synergistic anticandidal activity of pure polyphenol curcumin I in combination with azoles and polyenes generates reactive oxygen species leading to apoptosis. FEMS Yeast Res. 2010, 10, 570–578. [Google Scholar] [CrossRef]
- Sharma, M.; Manoharlal, M.R.; Puri, N.; Prasad, R. Antifungal curcumin induces reactive oxygen species and triggers an early apoptosis but prevents hyphae development by targeting the global repressor RUP in Candida albicans. Biosci. Rep. 2010, 30, 91–404. [Google Scholar] [CrossRef] [PubMed]
- Miao, H.; Zhao, L.; Li, C.; Shang, Q.; Lu, H.; Fu, Z.; Wang, L.; Jiang, Y.; Cao, Y. Inhibitory effect of shikonin on Candida albicans growth. Biol. Pharm. Bull. 2012, 35, 1956–1963. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Tan, F.; Miao, H.; Wang, H.; Cao, Y. Effect of shikonin on Candida albicans biofilms. Front. Microbiol. 2019, 10, 1085–1096. [Google Scholar] [CrossRef] [PubMed]
- Struyfs, C.; Cammue, B.P.; Thevissen, K. Membrane-interacting antifungal peptides. Front. Cell Dev. Biol. 2021, 9, 649875–649892. [Google Scholar] [CrossRef]
- Li, T.; Li, L.; Du, F.; Sun, L.; Shi, J.; Long, M.; Chen, Z. Activity and mechanism of action of antifungal peptides from microorganisms: A review. Molecules 2021, 26, 3438. [Google Scholar] [CrossRef]
- Puri, S.; Edgerton, M. How does it kill: Understanding the candidacidal mechanism of salivary histatin 5. Eukaryot. Cell 2014, 13, 958–964. [Google Scholar] [CrossRef]
- Vriens, K.; Cools, T.L.; Harvey, P.J.; Craik, D.J.; Braem, A.; Vleugels, J.; De Coninck, B.; Cammue, B.P.; Thevissen, K. The radish defensins RsAFP1 and RsAFP2 act synergistically with caspofungin against Candida albicans biofilms. Peptides 2016, 75, 71–79. [Google Scholar] [CrossRef]
- Cools, T.L.; Struyfs, C.; Cammue, B.P.; Thevissen, K. Antifungal plant defensins: Increased insight in their mode of action as a basis for their use to combat fungal infections. Future Microbiol. 2017, 12, 441–454. [Google Scholar] [CrossRef]
- Chowdhury, M.H.; Ryan, L.K.; Cherabuddi, K.; Freeman, K.B.; Weaver, D.G.; Pelletier, J.C.; Scott, R.W.; Diamond, G. Antifungal potential of host defense peptide mimetics in a mouse model of disseminated candidiasis. J. Fungi 2018, 4, 30. [Google Scholar] [CrossRef]
- Menzel, L.P.; Chowdhury, H.M.; Masso-Silva, J.A.; Ruddick, W.; Falkovsky, K.; Vorona, R.; Malsbary, A.; Cherabuddi, K.; Ryan, L.K.; DiFranco, K.M.; et al. Potent in vitro and in vivo antifungal activity of a small molecule host defense peptide mimic through a membrane-active mechanism. Sci. Rep. 2017, 7, 4353. [Google Scholar] [CrossRef]
- Ryan, L.K.; Freeman, K.B.; Masso-Silva, J.A.; Falkovsky, K.; Aloyouny, A.; Markowitz, K.; Hise, A.G.; Fatahzadeh, M.; Scott, R.W.; Diamond, G. Activity of potent and selective host defense peptide mimetics in mouse models of oral candidiasis. Antimicrob. Agents Chemother. 2014, 58, 3820–3827. [Google Scholar] [CrossRef]
- Fulda, S.; Debatin, K.M. Extrinsic versus intrinsic apoptosis pathways in anticancer chemotherapy. Oncogene 2006, 25, 4798–4811. [Google Scholar] [CrossRef]
- Fisher, M.C.; Alastruey-Izquierdo, A.; Berman, J.; Bicanic, T.; Bignell, E.M.; Bowyer, P.; Bromley, M.; Brüggemann, R.; Garber, G.; Cornely, O.A.; et al. Tackling the emerging threat of antifungal resistance to human health. Nat. Rev. Microbiol. 2022, 20, 557–571. [Google Scholar] [CrossRef] [PubMed]
- Ferris, D.G.; Nyirjesy, P.; Sobel, J.D.; Soper, D.; Pavletic, A.; Litaker, M.S. Over-the-counter antifungal drug misuse associated with patient-diagnosed vulvovaginal candidiasis. Obstet. Gynecol. 2002, 99, 419–425. [Google Scholar]
- Pathakumari, B.; Liang, G.; Liu, W. Immune defence to invasive fungal infections: A comprehensive review. Biomed. Pharm. 2020, 130, 110550–110567. [Google Scholar] [CrossRef]
- Eliopoulos, G.M.; Perea, S.; Patterson, T.F. Antifungal resistance in pathogenic fungi. Clin. Infect. Dis. 2002, 35, 1073–1080. [Google Scholar]
- Brauer, V.S.; Rezende, C.P.; Pessoni, A.M.; De Paula, R.G.; Rangappa, K.S.; Nayaka, S.C.; Gupta, V.K.; Almeida, F. Antifungal agents in agriculture: Friends and foes of public health. Biomolecules 2019, 9, 521. [Google Scholar] [CrossRef] [PubMed]
- Mayr, A.; Lass-Flörl, C. Epidemiology and antifungal resistance in invasive aspergillosis according to primary disease-review of the literature. Eur. J. Med. Res. 2011, 16, 153–157. [Google Scholar] [CrossRef]
- Robbins, N.; Caplan, T.; Cowan, L.E. Molecular evolution of antifungal drug resistance. Ann. Rev. Microbiol. 2017, 71, 753–775. [Google Scholar] [CrossRef]
- Tharappel, A.M.; Li, Z.; Zhu, Y.C.; Wu, X.; Chaturvedi, S.; Zhang, Q.Y.; Li, H. Calcimycin inhibits Cryptococcus neoformans in vitro and in vivo by targeting the Prp8 intein splicing. ACS Infect. Dis. 2022, 8, 1851–1868. [Google Scholar] [CrossRef]
- Pfaller, M.A. Antifungal drug resistance: Mechanisms, epidemiology, and consequences for treatment. Am. J. Med. 2012, 125, S3–S13. [Google Scholar] [CrossRef]
- Forsberg, K.; Woodworth, K.; Walters, M.; Berkow, E.L.; Jackson, B.; Chiller, T.; Vallabhaneni, S. Candida auris: The recent emergence of a multidrug-resistant fungal pathogen. Med. Mycol. 2019, 57, 1–12. [Google Scholar] [CrossRef]
- Lee, Y.; Puumala, E.; Robbins, N.; Cowen, L.E. Antifungal drug resistance: Molecular mechanisms in Candida albicans and beyond. Chem. Rev. 2020, 121, 3390–3411. [Google Scholar] [CrossRef] [PubMed]
- Lockhart, S.R.; Iqbal, N.; Cleveland, A.A.; Farley, M.M.; Harrison, L.H.; Bolden, C.B.; Baughman, W.; Stein, B.; Hollick, R.; Park, B.J.; et al. Species identification and antifungal susceptibility testing of Candida bloodstream isolates from population-based surveillance studies in two US cities from 2008 to 2011. J. Clin. Microbiol. 2012, 50, 3435–3442. [Google Scholar] [CrossRef] [PubMed]
- Franz, R.; Kelly, S.L.; Lamb, D.C.; Kelly, D.E.; Ruhnke, M.; Morschhäuser, J. Multiple molecular mechanisms contribute to a stepwise development of fluconazole resistance in clinical Candida albicans strains. Antimicrob. Agents Chemother. 1998, 42, 3065–3072. [Google Scholar] [CrossRef] [PubMed]
- Feng, W.; Yang, J.; Xi, Z.; Qiao, Z.; Lv, Y.; Wang, Y.; Ma, Y.; Wang, Y.; Cen, W. Mutations and/or overexpressions of ERG4 and ERG11 genes in clinical azoles-resistant isolates of Candida albicans. Microb. Drug Resist. 2017, 23, 563–570. [Google Scholar] [CrossRef] [PubMed]
- Mane, A.; Vidhate, P.; Kusro, C.; Waman, V.; Saxena, V.; Kulkarni-Kale, U.; Risbud, A. Molecular mechanisms associated with fluconazole resistance in clinical Candida albicans isolates from India. Mycoses 2016, 59, 93–100. [Google Scholar] [CrossRef]
- Hargrove, T.Y.; Friggeri, L.; Wawrzak, Z.; Qi, A.; Hoekstra, W.J.; Schotzinger, R.J.; York, J.D.; Guengerich, F.P.; Lepesheva, G.I. Structural analyses of Candida albicans sterol 14α-demethylase complexed with azole drugs address the molecular basis of azole-mediated inhibition of fungal sterol biosynthesis. J. Biol. Chem. 2017, 292, 6728–6743. [Google Scholar] [CrossRef] [PubMed]
- Nigam, P.K. Antifungal drugs and resistance: Current concepts. Our Dermatol. Online 2015, 6, 212. [Google Scholar] [CrossRef]
- Perlin, D.S.; Shor, E.; Zhao, Y. Update on antifungal drug resistance. Curr. Clin. Microbiol. Rep. 2015, 2, 84–95. [Google Scholar] [CrossRef] [PubMed]
- De Béthune, M.P. Non-nucleoside reverse transcriptase inhibitors (NNRTIs), their discovery, development, and use in the treatment of HIV-1 infection: A review of the last 20 years (1989–2009). Antivir. Res. 2010, 85, 75–90. [Google Scholar] [CrossRef] [PubMed]
- Healey, K.R.; Perlin, D.S. Fungal resistance to echinocandins and the MDR phenomenon in Candida glabrata. J. Fungi 2018, 4, 105. [Google Scholar] [CrossRef] [PubMed]
- Indiveri, C.; Galluccio, M.; Scalise, M.; Pochini, L. Strategies of bacterial over expression of membrane transporters relevant in human health: The successful case of the three members of OCTN subfamily. Mol. Biotechnol. 2013, 54, 724–736. [Google Scholar] [CrossRef] [PubMed]
- Khandelwal, N.K.; Kaemmer, P.; Förster, T.M.; Singh, A.; Coste, A.T.; Andes, D.R.; Hube, B.; Sanglard, D.; Chauhan, N.; Kaur, R.; et al. Pleiotropic effects of the vacuolar ABC transporter MLT1 of Candida albicans on cell function and virulence. Biochem. J. 2016, 473, 1537–1552. [Google Scholar] [CrossRef] [PubMed]
- Alcazar-Fuoli, L.; Mellado, E. Ergosterol biosynthesis in Aspergillus fumigatus: Its relevance as an antifungal target and role in antifungal drug resistance. Front. Microbiol. 2013, 3, 439–445. [Google Scholar] [CrossRef] [PubMed]
- Mellado, E.; Garcia-Effron, G.; Alcazar-Fuoli, L.; Melchers, W.J.G.; Verweij, P.E.; Cuenca-Estrella, M.; Rodriguez-Tudela, J.L. A new Aspergillus fumigatus resistance mechanism conferring in vitro cross-resistance to azole antifungals involves a combination of cyp51A alterations. Antimicrob. Agents Chemother. 2007, 51, 1897–1904. [Google Scholar] [CrossRef] [PubMed]
- Tuohy, M.J.; Reja, V.; Park, S.; Perlin, D.S.; Wnek, M.; Procop, G.W.; Yen-Lieberman, B. Use of a high-resolution melt assay to characterize codon 54 of the cyp51A gene of Aspergillus fumigatus on a Rotor-Gene 6000 instrument. Antimicrob. Agents Chemother. 2010, 54, 2248–2251. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Mavridou, E.; Brüggemann, R.J.; Melchers, W.J.; Mouton, J.W.; Verweij, P.E. Efficacy of posaconazole against three clinical Aspergillus fumigatus isolates with mutations in the cyp51A gene. Antimicrob. Agents Chemother. 2010, 54, 860–865. [Google Scholar] [CrossRef] [PubMed]
- Morschhäuser, J. The development of fluconazole resistance in Candida albicans—An example of microevolution of a fungal pathogen. J. Microbiol. 2016, 54, 192–201. [Google Scholar] [CrossRef] [PubMed]
- Lv, Q.Z.; Yan, L.; Jiang, Y.Y. The synthesis, regulation, and functions of sterols in Candida albicans: Well-known but still lots to learn. Virulence 2016, 7, 649–659. [Google Scholar] [CrossRef]
- Marie, C.; White, T.C. Genetic basis of antifungal drug resistance. Curr. Fungal Infect. Rep. 2009, 3, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Chowdhary, A.; Kathuria, S.; Xu, J.; Meis, J.F. Correction: Emergence of azole-resistant Aspergillus fumigatus strains due to agricultural azole use creates an increasing threat to human health. PLoS Pathog. 2013, 9, e1003633–e1003638. [Google Scholar] [CrossRef]
- Garcia, E.O. Infection Structure-Specificity of β-1,3-Glucan Synthase is Essential for Pathogenicity of Colletotrichum graminicola and Evasion of Glucan-Triggered Immunity. Ph.D. Thesis, Universitäts-und Landesbibliothek Sachsen-Anhalt, Halle (Saale), Germany, 2013. [Google Scholar]
- LaFayette, S.L.; Collins, C.; Zaas, A.K.; Schell, W.A.; Betancourt-Quiroz, M.; Gunatilaka, A.L.; Perfect, J.R.; Cowen, L.E. PKC signaling regulates drug resistance of the fungal pathogen Candida albicans via circuitry comprised of Mkc1, calcineurin, and Hsp90. PLoS Pathog. 2010, 6, e1001069–e1001092. [Google Scholar]
- Denning, D.W. Antifungal drug resistance: An update. Eur. J. Hosp. Pharm. 2022, 29, 109–112. [Google Scholar] [CrossRef]
- Arendrup, M.C.; Patterson, T.F. Multidrug-resistant Candida: Epidemiology, molecular mechanisms, and treatment. J. Infect. Dis. 2017, 216, S445–S451. [Google Scholar] [CrossRef] [PubMed]
- Pfaller, M.A.; Diekema, D. Epidemiology of invasive candidiasis: A persistent public health problem. Clin. Microbiol. Rev. 2007, 20, 133–163. [Google Scholar] [CrossRef] [PubMed]
- Rapp, R.P. Changing strategies for the management of invasive fungal infections. Pharmacother. J. Hum. Pharmacol. Drug Ther. 2004, 24, 4S–28S. [Google Scholar]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hossain, C.M.; Ryan, L.K.; Gera, M.; Choudhuri, S.; Lyle, N.; Ali, K.A.; Diamond, G. Antifungals and Drug Resistance. Encyclopedia 2022, 2, 1722-1737. https://doi.org/10.3390/encyclopedia2040118
Hossain CM, Ryan LK, Gera M, Choudhuri S, Lyle N, Ali KA, Diamond G. Antifungals and Drug Resistance. Encyclopedia. 2022; 2(4):1722-1737. https://doi.org/10.3390/encyclopedia2040118
Chicago/Turabian StyleHossain, Chowdhury Mobaswar, Lisa Kathleen Ryan, Meeta Gera, Sabyasachi Choudhuri, Nazmun Lyle, Kazi Asraf Ali, and Gill Diamond. 2022. "Antifungals and Drug Resistance" Encyclopedia 2, no. 4: 1722-1737. https://doi.org/10.3390/encyclopedia2040118
APA StyleHossain, C. M., Ryan, L. K., Gera, M., Choudhuri, S., Lyle, N., Ali, K. A., & Diamond, G. (2022). Antifungals and Drug Resistance. Encyclopedia, 2(4), 1722-1737. https://doi.org/10.3390/encyclopedia2040118