

Probiotics as Antibiotic Alternatives for Human and Animal Applications


Abstract
:1. Introduction
2. Antibiotics: Antimicrobial Resistance Causes and Potential Alternatives
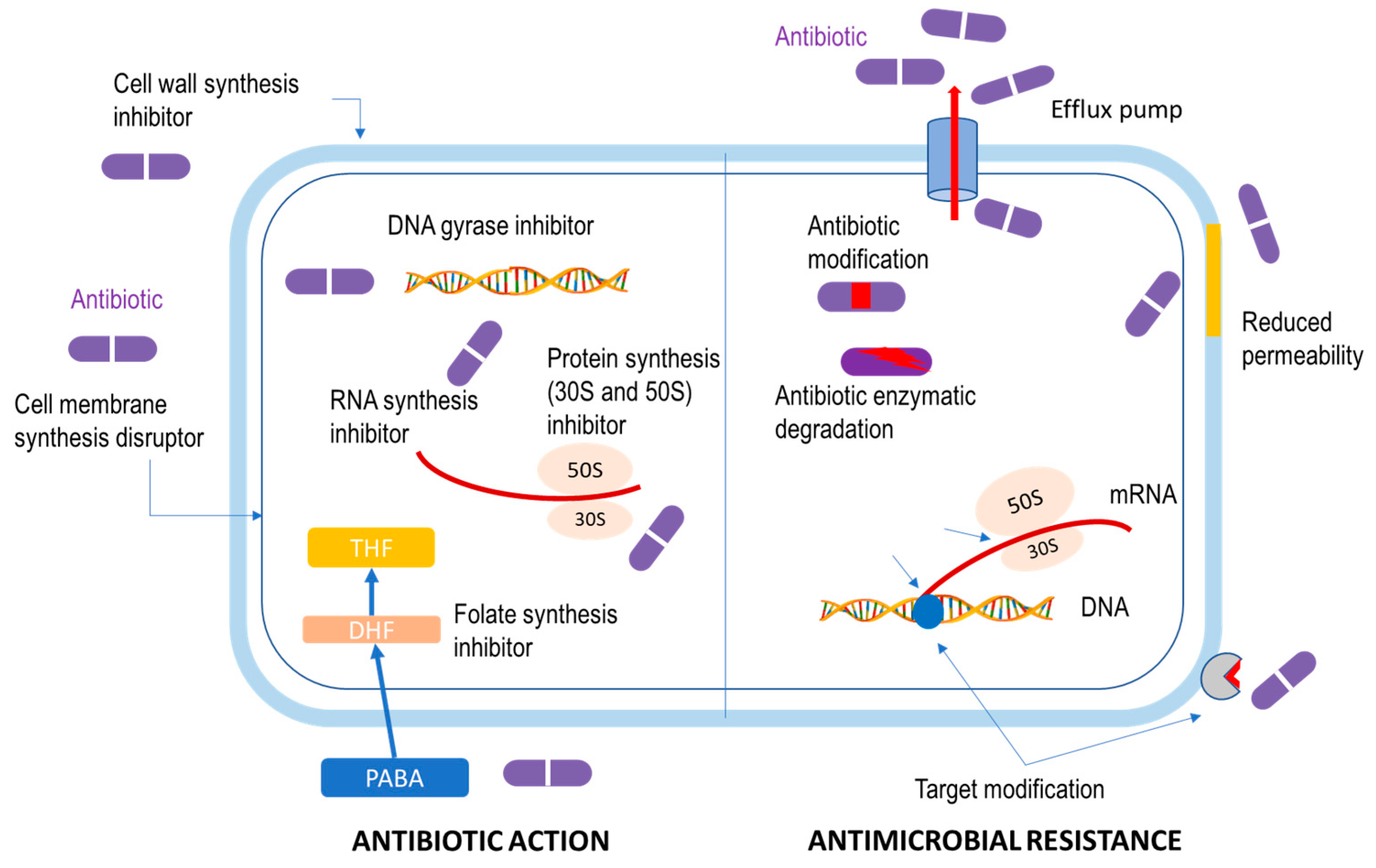
2.1. Basic Concept of Antibiotics
2.2. Antimicrobial Resistance (AMR) Issues
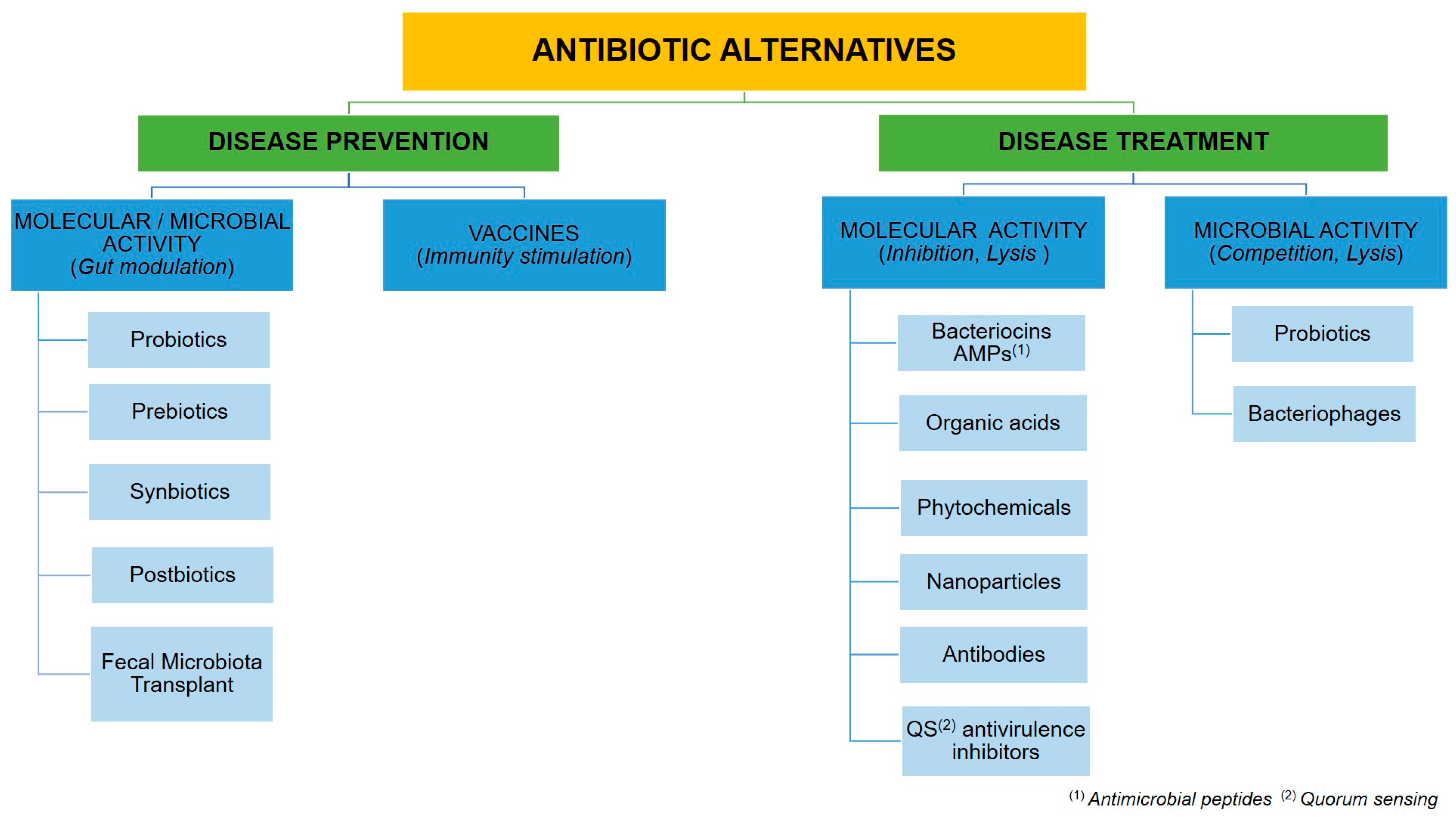
2.3. Alternatives to Antibiotics
3. Probiotics as Potential Alternative to Antibiotics
3.1. Competitive Exclusion of Pathogens
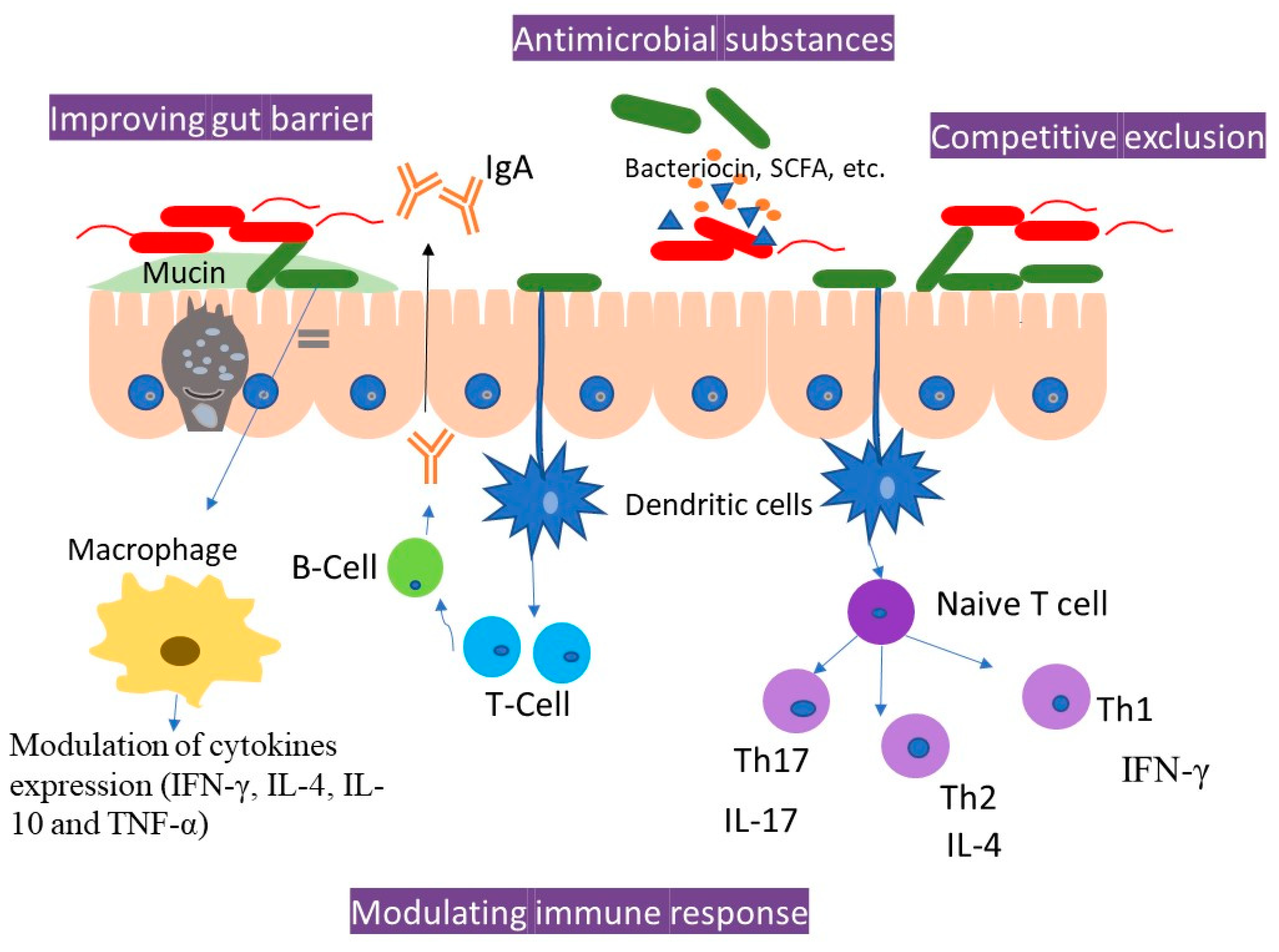
3.2. Improvement of Intestinal Barriers
3.3. Secretion of Antimicrobial Peptides (AMPs)
3.4. Modulation of Host Immune System
3.4.1. Stimulation of Phagocytic Activity
3.4.2. Balancing of Pro- and Anti-Inflammatory Cytokines
3.4.3. Enhancing Immunoglobulin A (IgA) Production
4. Advantages and Disadvantages of Probiotics as Antibiotic Alternatives
5. Human Applications
6. Animal Applications
7. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kourkouta, L.; Koukourikos, K.; Iliadis, C.; Plati, P.; Dimitriadou, A. History of Antibiotics. Sumer. J. Med. Healthc. 2018, 1, 51–55. [Google Scholar]
- World Health Organization. Antimicrobial Resistance: Global Report on Surveillance; World Health Organization: Geneva, Switzerland, 2014; ISBN 92-4-156474-1. [Google Scholar]
- O’Neill, J. Tackling Drug-Resistant Infections Globally: Final Report and Recommendations. 2016. Available online: https://amr-review.org/sites/default/files/160525_Final%20paper_with%20cover.pdf (accessed on 24 March 2023).
- Tegegne, B.A.; Kebede, B. Probiotics, their prophylactic and therapeutic applications in human health development: A review of the literature. Heliyon 2022, 8, e09725. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Sood, U.; Gupta, V.; Singh, M.; Scaria, J.; Lal, R. Recent Advancements in the Development of Modern Probiotics for Restoring Human Gut Microbiome Dysbiosis. Indian J. Microbiol. 2020, 60, 12–25. [Google Scholar] [CrossRef] [PubMed]
- Allen, H.K.; Trachsel, J.; Looft, T.; Casey, T.A. Finding alternatives to antibiotics. Ann. N. Y. Acad. Sci. 2014, 1323, 91–100. [Google Scholar] [CrossRef]
- Reid, G. Probiotics to Prevent the Need for, and Augment the Use of, Antibiotics. Can. J. Infect. Dis. Med Microbiol. 2006, 17, 291–295. [Google Scholar] [CrossRef]
- Petersen, C.; Round, J.L. Defining dysbiosis and its influence on host immunity and disease. Cell. Microbiol. 2014, 16, 1024–1033. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef]
- Vieco-Saiz, N.; Belguesmia, Y.; Raspoet, R.; Auclair, E.; Gancel, F.; Kempf, I.; Drider, D. Benefits and Inputs from Lactic Acid Bacteria and Their Bacteriocins as Alternatives to Antibiotic Growth Promoters during Food-Animal Production. Front. Microbiol. 2019, 10, 57. [Google Scholar] [CrossRef]
- Aghebati-Maleki, L.; Hasannezhad, P.; Abbasi, A.; Khani, N. Antibacterial, Antiviral, Antioxidant, and Anticancer Activities of Postbiotics: A Review of Mechanisms and Therapeutic Perspectives. Biointerface Res. Appl. Chem. 2021, 12, 2629–2645. [Google Scholar]
- Knipe, H.; Temperton, B.; Lange, A.; Bass, D.; Tyler, C.R. Probiotics and competitive exclusion of pathogens in shrimp aquaculture. Rev. Aquac. 2021, 13, 324–352. [Google Scholar] [CrossRef]
- Azad, M.; Kalam, A.; Sarker, M.; Wan, D. Immunomodulatory Effects of Probiotics on Cytokine Profiles. BioMed Res. Int. 2018, 2018, 8063647. [Google Scholar] [CrossRef] [PubMed]
- Yaqoob, M.U.; Wang, G.; Wang, M. An updated review on probiotics as an alternative of antibiotics in poultry—A review. Anim. Biosci. 2022, 35, 1109–1120. [Google Scholar] [CrossRef] [PubMed]
- Saettone, V.; Biasato, I.; Radice, E.; Schiavone, A.; Bergero, D.; Meineri, G. State-of-the-Art of the Nutritional Alternatives to the Use of Antibiotics in Humans and Monogastric Animals. Animals 2020, 10, 2199. [Google Scholar] [CrossRef] [PubMed]
- Bentley, R.; Bennett, J.W. What Is an Antibiotic? Revisited. Adv. Appl. Microbiol. 2003, 52, 303–332. [Google Scholar] [CrossRef] [PubMed]
- Etebu, E.; Arikekpar, I. Antibiotics: Classification and Mechanisms of Action with Emphasis on Molecular Perspectives. Int. J. Appl. Microbiol. Biotechnol. Res. 2016, 4, 90–101. [Google Scholar]
- Smith, A.D.; Datta, S.P.; Smith, G.H.; Campbell, P.N.; Bentley, R.; McKenzie, H.A.; Jakoby, W.B. Oxford Dictionary of Biochemistry and Molecular Biology. Trends Biochem. Sci. 1998, 23, 228. [Google Scholar]
- Hutchings, M.I.; Truman, A.W.; Wilkinson, B. Antibiotics: Past, Present and Future. Curr. Opin. Microbiol. 2019, 51, 72–80. [Google Scholar] [CrossRef]
- Pissowotzki, K.; Mansouri, K.; Piepersberg, W. Genetics of streptomycin production in Streptomyces griseus: Molecular structure and putative function of genes strELMB2N. Mol. Gen. Genet. MGG 1991, 231, 113–123. [Google Scholar] [CrossRef]
- Laich, F.; Fierro, F.; Martín, J.F. Production of Penicillin by Fungi Growing on Food Products: Identification of a Complete Penicillin Gene Cluster in Penicillium griseofulvum and a Truncated Cluster in Penicillium verrucosum. Appl. Environ. Microbiol. 2002, 68, 1211–1219. [Google Scholar] [CrossRef]
- Jung, H.-M.; Kim, S.-Y.; Moon, H.-J.; Oh, D.-K.; Lee, J.-K. Optimization of culture conditions and scale-up to pilot and plant scales for vancomycin production by Amycolatopsis orientalis. Appl. Microbiol. Biotechnol. 2007, 77, 789–795. [Google Scholar] [CrossRef]
- Miao, V.; Coëffet-LeGal, M.-F.; Brian, P.; Brost, R.; Penn, J.; Whiting, A.; Martin, S.; Ford, R.; Parr, I.; Bouchard, M.; et al. Daptomycin biosynthesis in Streptomyces roseosporus: Cloning and analysis of the gene cluster and revision of peptide stereochemistry. Microbiology 2005, 151, 1507–1523. [Google Scholar] [CrossRef] [PubMed]
- Weber, J.M.; Wierman, C.K.; Hutchinson, C.R. Genetic analysis of erythromycin production in Streptomyces erythreus. J. Bacteriol. 1985, 164, 425–433. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Martínez, L.T.; Borsetto, C.; Gomez-Escribano, J.P.; Bibb, M.J.; Al-Bassam, M.M.; Chandra, G.; Bibb, M.J. New Insights into Chloramphenicol Biosynthesis in Streptomyces venezuelae ATCC 10712. Antimicrob. Agents Chemother. 2014, 58, 7441–7450. [Google Scholar] [CrossRef] [PubMed]
- Fang, A.; Pierson, D.L.; Mishra, S.K.; Koenig, D.W.; Demain, A.L. Gramicidin S Production by Bacillus brevis in Simulated Microgravity. Curr. Microbiol. 1997, 34, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Naghmouchi, K.; Hammami, R.; Fliss, I.; Teather, R.; Baah, J.; Drider, D. Colistin A and colistin B among inhibitory substances of Paenibacillus polymyxa JB05-01-1. Arch. Microbiol. 2012, 194, 363–370. [Google Scholar] [CrossRef]
- Petković, H.; Lukežič, T.; Šušković, J. Biosynthesis of Oxytetracycline by Streptomyces rimosus: Past, Present and Future Directions in the Development of Tetracycline Antibiotics. Food Technol. Biotechnol. 2017, 55, 3–13. [Google Scholar] [CrossRef]
- Dowling, A.; O’dwyer, J.; Adley, C. Antibiotics: Mode of Action and Mechanisms of Resistance. Antimicrob. Res. Nov. Bioknowledge Educ. Programs 2017, 1, 536–545. [Google Scholar]
- Abushaheen, M.A.; Fatani, A.J.; Alosaimi, M.; Mansy, W.; George, M.; Acharya, S.; Rathod, S.; Divakar, D.D.; Jhugroo, C.; Vellappally, S. Antimicrobial Resistance, Mechanisms and Its Clinical Significance. Dis. Mon. 2020, 66, 100971. [Google Scholar] [CrossRef]
- Aghamohammad, S.; Rohani, M. Antibiotic resistance and the alternatives to conventional antibiotics: The role of probiotics and microbiota in combating antimicrobial resistance. Microbiol. Res. 2022, 267, 127275. [Google Scholar] [CrossRef]
- Bai, H.; He, L.-Y.; Wu, D.-L.; Gao, F.-Z.; Zhang, M.; Zou, H.-Y.; Yao, M.-S.; Ying, G.-G. Spread of airborne antibiotic resistance from animal farms to the environment: Dispersal pattern and exposure risk. Environ. Int. 2021, 158, 106927. [Google Scholar] [CrossRef]
- Ma, F.; Xu, S.; Tang, Z.; Li, Z.; Zhang, L. Use of antimicrobials in food animals and impact of transmission of antimicrobial resistance on humans. Biosaf. Health 2021, 3, 32–38. [Google Scholar] [CrossRef]
- Papadopoulos, P.; Angelidis, A.S.; Papadopoulos, T.; Kotzamanidis, C.; Zdragas, A.; Papa, A.; Filioussis, G.; Sergelidis, D. Staphylococcus aureus and methicillin-resistant S. aureus (MRSA) in bulk tank milk, livestock and dairy-farm personnel in north-central and north-eastern Greece: Prevalence, characterization and genetic relatedness. Food Microbiol. 2019, 84, 103249. [Google Scholar] [CrossRef] [PubMed]
- Modi, S.R.; Collins, J.J.; Relman, D.A. Antibiotics and the Gut Microbiota. J. Clin. Investig. 2014, 124, 4212–4218. [Google Scholar] [CrossRef]
- Smillie, C.S.; Smith, M.B.; Friedman, J.; Cordero, O.X.; David, L.A.; Alm, E.J. Ecology drives a global network of gene exchange connecting the human microbiome. Nature 2011, 480, 241–244. [Google Scholar] [CrossRef] [PubMed]
- Brandl, K.; Plitas, G.; Mihu, C.N.; Ubeda, C.; Jia, T.; Fleisher, M.; Schnabl, B.; DeMatteo, R.P.; Pamer, E.G. Vancomycin-resistant enterococci exploit antibiotic-induced innate immune deficits. Nature 2008, 455, 804–807. [Google Scholar] [CrossRef]
- Fukuda, S.; Toh, H.; Hase, K.; Oshima, K.; Nakanishi, Y.; Yoshimura, K.; Tobe, T.; Clarke, J.M.; Topping, D.L.; Suzuki, T.; et al. Bifidobacteria can protect from enteropathogenic infection through production of acetate. Nature 2011, 469, 543–547. [Google Scholar] [CrossRef]
- Diep, B.A.; Gill, S.R.; Chang, R.F.; Phan, T.H.; Chen, J.H.; Davidson, M.G.; Lin, F.; Lin, J.; Carleton, H.A.; Mongodin, E.F.; et al. Complete genome sequence of USA300, an epidemic clone of community-acquired meticillin-resistant Staphylococcus aureus. Lancet 2006, 367, 731–739. [Google Scholar] [CrossRef]
- Darby, E.M.; Trampari, E.; Siasat, P.; Gaya, M.S.; Alav, I.; Webber, M.A.; Blair, J.M.A. Molecular mechanisms of antibiotic resistance revisited. Nat. Rev. Microbiol. 2022, 21, 280–295. [Google Scholar] [CrossRef]
- Singh, S.P.; Qureshi, A.; Hassan, W. Mechanisms of action by antimicrobial agents: A review. McGill J. Med. 2021, 19, 1–10. [Google Scholar] [CrossRef]
- Uddin, T.M.; Chakraborty, A.J.; Khusro, A.; Zidan, B.R.M.; Mitra, S.; Bin Emran, T.; Dhama, K.; Ripon, K.H.; Gajdács, M.; Sahibzada, M.U.K.; et al. Antibiotic resistance in microbes: History, mechanisms, therapeutic strategies and future prospects. J. Infect. Public Health 2021, 14, 1750–1766. [Google Scholar] [CrossRef]
- Krause, K.M.; Serio, A.W.; Kane, T.R.; Connolly, L.E. Aminoglycosides: An Overview. Cold Spring Harb. Perspect. Med. 2016, 6, a027029. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, R.; Amador, P.; Prudêncio, C. β-Lactams: Chemical Structure, Mode of Action and Mechanisms of Resistance. Rev. Res. Med. Microbiol. 2013, 24, 7–17. [Google Scholar] [CrossRef]
- Zeng, D.; Debabov, D.; Hartsell, T.L.; Cano, R.J.; Adams, S.; Schuyler, J.A.; McMillan, R.; Pace, J.L. Approved Glycopeptide Antibacterial Drugs: Mechanism of Action and Resistance. Cold Spring Harb. Perspect. Med. 2016, 6, a026989. [Google Scholar] [CrossRef] [PubMed]
- Redgrave, L.S.; Sutton, S.B.; Webber, M.A.; Piddock, L.J. Fluoroquinolone resistance: Mechanisms, impact on bacteria, and role in evolutionary success. Trends Microbiol. 2014, 22, 438–445. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.-W.; Thawng, C.N.; Lee, K.; Wellington, E.M.; Cha, C.-J. A novel sulfonamide resistance mechanism by two-component flavin-dependent monooxygenase system in sulfonamide-degrading actinobacteria. Environ. Int. 2019, 127, 206–215. [Google Scholar] [CrossRef]
- Grossman, T.H. Tetracycline Antibiotics and Resistance. Cold Spring Harb. Perspect. Med. 2016, 6, a025387. [Google Scholar] [CrossRef]
- Schwarz, S.; Shen, J.; Kadlec, K.; Wang, Y.; Michael, G.B.; Feßler, A.T.; Vester, B. Lincosamides, Streptogramins, Phenicols, and Pleuromutilins: Mode of Action and Mechanisms of Resistance. Cold Spring Harb. Perspect. Med. 2016, 6, a027037. [Google Scholar] [CrossRef]
- Helmy, Y.A.; Taha-Abdelaziz, K.; Hawwas, H.A.E.-H.; Ghosh, S.; AlKafaas, S.S.; Moawad, M.M.M.; Saied, E.M.; Kassem, I.I.; Mawad, A.M.M. Antimicrobial Resistance and Recent Alternatives to Antibiotics for the Control of Bacterial Pathogens with an Emphasis on Foodborne Pathogens. Antibiotics 2023, 12, 274. [Google Scholar] [CrossRef]
- Silva, D.R.; Sardi, J.D.C.O.; de Souza Pitangui, N.; Roque, S.M.; da Silva, A.C.B.; Rosalen, P.L. Probiotics as an alternative antimicrobial therapy: Current reality and future directions. J. Funct. Foods 2020, 73, 104080. [Google Scholar] [CrossRef]
- Zamojska, D.; Nowak, A.; Nowak, I.; Macierzyńska-Piotrowska, E. Probiotics and Postbiotics as Substitutes of Antibiotics in Farm Animals: A Review. Animals 2021, 11, 3431. [Google Scholar] [CrossRef]
- Kunyeit, L.; K A, A.-A.; Rao, R.P. Application of Probiotic Yeasts on Candida Species Associated Infection. J. Fungi 2020, 6, 189. [Google Scholar] [CrossRef] [PubMed]
- El-Sharkawy, H.; Tahoun, A.; Rizk, A.M.; Suzuki, T.; Elmonir, W.; Nassef, E.; Shukry, M.; Germoush, M.O.; Farrag, F.; Bin-Jumah, M.; et al. Evaluation of Bifidobacteria and Lactobacillus Probiotics as Alternative Therapy for Salmonella typhimurium Infection in Broiler Chickens. Animals 2020, 10, 1023. [Google Scholar] [CrossRef]
- Raheem, A.; Liang, L.; Zhang, G.; Cui, S. Modulatory Effects of Probiotics During Pathogenic Infections with Emphasis on Immune Regulation. Front. Immunol. 2021, 12, 616713. [Google Scholar] [CrossRef]
- Schluter, J.; Nadell, C.D.; Bassler, B.L.; Foster, K.R. Adhesion as a weapon in microbial competition. ISME J. 2015, 9, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Siedler, S.; Rau, M.H.; Bidstrup, S.; Vento, J.M.; Aunsbjerg, S.D.; Bosma, E.F.; McNair, L.M.; Beisel, C.L.; Neves, A.R. Competitive exclusion is a major bioprotective mechanism of lactobacilli against fungal spoilage in fermented milk products. Appl. Environ. Microbiol. 2020, 86, e02312-19. [Google Scholar] [CrossRef] [PubMed]
- Zuo, F.; Appaswamy, A.; Gebremariam, H.G.; Jonsson, A.-B. Role of Sortase A in Lactobacillus gasseri Kx110A1 Adhesion to Gastric Epithelial Cells and Competitive Exclusion of Helicobacter pylori. Front. Microbiol. 2019, 10, 2770. [Google Scholar] [CrossRef] [PubMed]
- Lau, L.Y.J.; Chye, F.Y. Antagonistic effects of Lactobacillus plantarum 0612 on the adhesion of selected foodborne enteropathogens in various colonic environments. Food Control 2018, 91, 237–247. [Google Scholar] [CrossRef]
- Dhanani, A.; Bagchi, T. The expression of adhesin EF-Tu in response to mucin and its role in Lactobacillus adhesion and competitive inhibition of enteropathogens to mucin. J. Appl. Microbiol. 2013, 115, 546–554. [Google Scholar] [CrossRef]
- Vancamelbeke, M.; Vermeire, S. The intestinal barrier: A fundamental role in health and disease. Expert Rev. Gastroenterol. Hepatol. 2017, 11, 821–834. [Google Scholar] [CrossRef]
- Gou, H.-Z.; Zhang, Y.-L.; Ren, L.-F.; Li, Z.-J.; Zhang, L. How Do Intestinal Probiotics Restore the Intestinal Barrier? Front. Microbiol. 2022, 13, 929346. [Google Scholar] [CrossRef]
- Al-Sadi, R.; Nighot, P.; Nighot, M.; Haque, M.; Rawat, M.; Ma, T.Y. Lactobacillus acidophilus Induces a Strain-specific and Toll-Like Receptor 2–Dependent Enhancement of Intestinal Epithelial Tight Junction Barrier and Protection against Intestinal Inflammation. Am. J. Pathol. 2021, 191, 872–884. [Google Scholar] [CrossRef] [PubMed]
- La Fata, G.; Weber, P.; Mohajeri, M.H. Probiotics and the Gut Immune System: Indirect Regulation. Probiotics Antimicrob. Proteins 2018, 10, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.H.; Jeong, C.H.; Cheng, W.N.; Choi, Y.; Shin, D.M.; Lee, S.; Han, S.G. Quality characteristics of yogurts fermented with short-chain fatty acid-producing probiotics and their effects on mucin production and probiotic adhesion onto human colon epithelial cells. J. Dairy Sci. 2021, 104, 7415–7425. [Google Scholar] [CrossRef] [PubMed]
- Szczerbiec, D.; Piechocka, J.; Głowacki, R.; Torzewska, A. Organic Acids Secreted by Lactobacillus spp. Isolated from Urine and Their Antimicrobial Activity against Uropathogenic Proteus mirabilis. Molecules 2022, 27, 5557. [Google Scholar] [CrossRef]
- Huang, C.B.; Alimova, Y.; Myers, T.M.; Ebersole, J.L. Short- and medium-chain fatty acids exhibit antimicrobial activity for oral microorganisms. Arch. Oral Biol. 2011, 56, 650–654. [Google Scholar] [CrossRef]
- Kim, Y.; Yoon, S.; Shin, H.; Jo, M.; Lee, S.; Kim, S.-H. Isolation of Lactococcus lactis ssp. cremoris LRCC5306 and Optimization of Diacetyl Production Conditions for Manufacturing Sour Cream. Food Sci. Anim. Resour. 2021, 41, 373–385. [Google Scholar] [CrossRef]
- Tomás, M.S.J.; Otero, M.C.; Ocaña, V.; Nader-Macías, M.E. Production of Antimicrobial Substances by Lactic Acid Bacteria I: Determination of Hydrogen Peroxide. Public Health Microbiol. Methods Protoc. 2004, 268, 337–346. [Google Scholar] [CrossRef]
- Mejía-Pitta, A.; Broset, E.; de la Fuente-Nunez, C. Probiotic engineering strategies for the heterologous production of antimicrobial peptides. Adv. Drug Deliv. Rev. 2021, 176, 113863. [Google Scholar] [CrossRef]
- Hassan, M.; Kjos, M.; Nes, I.F.; Diep, D.B.; Lotfipour, F. Natural antimicrobial peptides from bacteria: Characteristics and potential applications to fight against antibiotic resistance. J. Appl. Microbiol. 2012, 113, 723–736. [Google Scholar] [CrossRef]
- Negash, A.W.; Tsehai, B.A. Current Applications of Bacteriocin. Int. J. Microbiol. 2020, 2020, 4374891. [Google Scholar] [CrossRef]
- Bharti, V.; Mehta, A.; Singh, S.; Jain, N.; Ahirwal, L.; Mehta, S. Bacteriocin: A Novel Approach for Preservation of Food. Int. J. Pharm. Pharm. Sci. 2015, 7, 20–29. [Google Scholar]
- Darvishi, N.; Fard, N.A.; Sadrnia, M. Genomic and proteomic comparisons of bacteriocins in probiotic species Lactobacillus and Bifidobacterium and inhibitory ability of Escherichia coli MG 1655. Biotechnol. Rep. 2021, 31, e00654. [Google Scholar] [CrossRef] [PubMed]
- Brötz, H.; Josten, M.; Wiedemann, I.; Schneider, U.; Götz, F.; Bierbaum, G.; Sahl, H.-G. Role of Lipid-bound Peptidoglycan Precursors in the Formation of Pores by Nisin, Epidermin and Other Lantibiotics. Mol. Microbiol. 1998, 30, 317–327. [Google Scholar] [CrossRef] [PubMed]
- Panina, I.; Taldaev, A.; Efremov, R.; Chugunov, A. Molecular Dynamics Insight into the Lipid II Recognition by Type a Lantibiotics: Nisin, Epidermin, and Gallidermin. Micromachines 2021, 12, 1169. [Google Scholar] [CrossRef]
- Wang, X.; Gu, Q.; Breukink, E. Non-lipid II targeting lantibiotics. Biochim. Biophys. Acta (BBA)-Biomembr. 2020, 1862, 183244. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, B.; Sukumar, G.; Ghosh, A.R. Purification and characterization of pediocin from probiotic Pediococcus pentosaceus GS4, MTCC 12683. Folia Microbiol. 2019, 64, 765–778. [Google Scholar] [CrossRef]
- Gaspar, C.; Donders, G.G.; Palmeira-De-Oliveira, R.; Queiroz, J.A.; Tomaz, C.; Martinez-De-Oliveira, J. Bacteriocin production of the probiotic Lactobacillus acidophilus KS400. AMB Express 2018, 8, 153. [Google Scholar] [CrossRef]
- Lauková, A.; Styková, E.; Kubašová, I.; Gancarčíková, S.; Plachá, I.; Mudroňová, D.; Kandričáková, A.; Miltko, R.; Belzecki, G.; Valocký, I.; et al. Enterocin M and its Beneficial Effects in Horses—A Pilot Experiment. Probiotics Antimicrob. Proteins 2018, 10, 420–426. [Google Scholar] [CrossRef]
- Lei, W.; Hao, L.; You, S.; Yao, H.; Liu, C.; Zhou, H. Partial purification and application of a bacteriocin produced by probiotic Lactococcus lactis C15 isolated from raw milk. LWT 2022, 169, 113917. [Google Scholar] [CrossRef]
- De Giani, A.; Bovio, F.; Forcella, M.; Fusi, P.; Sello, G.; Di Gennaro, P. Identification of a bacteriocin-like compound from Lactobacillus plantarum with antimicrobial activity and effects on normal and cancerogenic human intestinal cells. AMB Express 2019, 9, 88. [Google Scholar] [CrossRef]
- Wei, Z.; Shan, C.; Zhang, L.; Wang, Y.; Xia, X.; Liu, X.; Zhou, J. A novel subtilin-like lantibiotics subtilin JS-4 produced by Bacillus subtilis JS-4, and its antibacterial mechanism against Listeria monocytogenes. LWT 2021, 142, 110993. [Google Scholar] [CrossRef]
- Zambori, C.; Cumpănăşoiu, C.; Moţ, D.; Huţu, I.; Gurban, C.; Tîrziu, E. The Antimicrobial Role of Probiotics in the Oral Cavity in Humans and Dogs. Anim. Sci. Biotechnol. 2014, 47, 126–130. [Google Scholar]
- Mazziotta, C.; Tognon, M.; Martini, F.; Torreggiani, E.; Rotondo, J.C. Probiotics Mechanism of Action on Immune Cells and Beneficial Effects on Human Health. Cells 2023, 12, 184. [Google Scholar] [CrossRef] [PubMed]
- Jha, A.; Mishra, V.K.; Mohammad, G. Immunomodulation and anticancer potentials of yogurt probiotic. 2008. EXCLI J. 2008, 7, 177–184. [Google Scholar]
- Noh, H.-J.; Park, J.M.; Kwon, Y.J.; Kim, K.; Park, S.Y.; Kim, I.; Lim, J.H.; Kim, B.K.; Kim, B.-Y. Immunostimulatory Effect of Heat-Killed Probiotics on RAW264. 7 Macrophages. J. Microbiol. Biotechnol. 2022, 32, 638–644. [Google Scholar] [CrossRef]
- Duarte, J.; Vinderola, G.; Ritz, B.; Perdigón, G.; Matar, C. Immunomodulating capacity of commercial fish protein hydrolysate for diet supplementation. Immunobiology 2006, 211, 341–350. [Google Scholar] [CrossRef]
- Lebeer, S.; Vanderleyden, J.; De Keersmaecker, S.C. Host interactions of probiotic bacterial surface molecules: Comparison with commensals and pathogens. Nat. Rev. Microbiol. 2010, 8, 171–184. [Google Scholar] [CrossRef]
- Rocha-Ramírez, L.M.; Pérez-Solano, R.A.; Castañón-Alonso, S.L.; Moreno Guerrero, S.S.; Ramírez Pacheco, A.; García Garibay, M.; Eslava, C. Probiotic Lactobacillus Strains Stimulate the Inflammatory Response and Activate Human Macrophages. J. Immunol. Res. 2017, 2017, 4607491. [Google Scholar] [CrossRef]
- Donnet-Hughes, A.; Rochat, F.; Serrant, P.; Aeschlimann, J.M.; Schiffrin, E.J. Modulation of Nonspecific Mechanisms of Defense by Lactic Acid Bacteria: Effective Dose. J. Dairy Sci. 1999, 82, 863–869. [Google Scholar] [CrossRef]
- Kany, S.; Vollrath, J.T.; Relja, B. Cytokines in Inflammatory Disease. Int. J. Mol. Sci. 2019, 20, 6008. [Google Scholar] [CrossRef]
- Cavaillon, J.M. Pro- versus anti-inflammatory cytokines: Myth or reality. Cell. Mol. Biol.-Paris-Wegmann 2001, 47, 695–702. [Google Scholar]
- Dargahi, N.; Matsoukas, J.; Apostolopoulos, V. Streptococcus thermophilus ST285 Alters Pro-Inflammatory to Anti-Inflammatory Cytokine Secretion against Multiple Sclerosis Peptide in Mice. Brain Sci. 2020, 10, 126. [Google Scholar] [CrossRef] [PubMed]
- Dargahi, N.; Johnson, J.C.; Apostolopoulos, V. Immune Modulatory Effects of Probiotic Streptococcus thermophilus on Human Monocytes. Biologics 2021, 1, 396–415. [Google Scholar] [CrossRef]
- López-Varela, S.; Gonzalez-Gross, M.; Marcos, A. Functional foods and the immune system: A review. Eur. J. Clin. Nutr. 2002, 56, S29–S33. [Google Scholar] [CrossRef]
- Curciarello, R.; Canziani, K.E.; Salto, I.; Romero, E.B.; Rocca, A.; Doldan, I.; Peton, E.; Brayer, S.; Sambuelli, A.M.; Goncalves, S.; et al. Probiotic Lactobacilli Isolated from Kefir Promote Down-Regulation of Inflammatory Lamina Propria T Cells from Patients with Active IBD. Front. Pharmacol. 2021, 12, 658026. [Google Scholar] [CrossRef]
- Pietrzak, B.; Tomela, K.; Olejnik-Schmidt, A.; Mackiewicz, A.; Schmidt, M. Secretory IgA in Intestinal Mucosal Secretions as an Adaptive Barrier against Microbial Cells. Int. J. Mol. Sci. 2020, 21, 9254. [Google Scholar] [CrossRef]
- Kawashima, T.; Ikari, N.; Kouchi, T.; Kowatari, Y.; Kubota, Y.; Shimojo, N.; Tsuji, N.M. The molecular mechanism for activating IgA production by Pediococcus acidilactici K15 and the clinical impact in a randomized trial. Sci. Rep. 2018, 8, 5065. [Google Scholar] [CrossRef]
- Sakai, F.; Hosoya, T.; Ono-Ohmachi, A.; Ukibe, K.; Ogawa, A.; Moriya, T.; Kadooka, Y.; Shiozaki, T.; Nakagawa, H.; Nakayama, Y.; et al. Lactobacillus gasseri SBT2055 Induces TGF-β Expression in Dendritic Cells and Activates TLR2 Signal to Produce IgA in the Small Intestine. PLoS ONE 2014, 9, e105370. [Google Scholar] [CrossRef]
- Thoreux, K.; Owen, R.; Schmucker, D.L. Functional Foods, Mucosal Immunity and Aging: Effect of Probiotics on Intestinal Immunity in Young and Old Rats. Commun. Curr. Res. Educ. Top. Trends Appl. Microbiol. 2007, 1, 458–465. [Google Scholar]
- Perdigon, G.; Alvarez, S.; Rachid, M.; Agüero, G.; Gobbato, N. Immune System Stimulation by Probiotics. J. Dairy Sci. 1995, 78, 1597–1606. [Google Scholar] [CrossRef]
- Lin, W.-Y.; Kuo, Y.-W.; Chen, C.-W.; Huang, Y.-F.; Hsu, C.-H.; Lin, J.-H.; Liu, C.-R.; Chen, J.-F.; Hsia, K.-C.; Ho, H.-H. Viable and Heat-Killed Probiotic Strains Improve Oral Immunity by Elevating the IgA Concentration in the Oral Mucosa. Curr. Microbiol. 2021, 78, 3541–3549. [Google Scholar] [CrossRef] [PubMed]
- Kaila, M.; Isolauri, E.; Soppi, E.S.A.; Virtanen, E.; Laine, S.; Arvilommi, H. Enhancement of the Circulating Antibody Secreting Cell Response in Human Diarrhea by a Human Lactobacillus Strain. Pediatr. Res. 1992, 32, 141–144. [Google Scholar] [CrossRef] [PubMed]
- Finamore, A.; Roselli, M.; Donini, L.M.; Brasili, E.; Rami, R.; Carnevali, P.; Mistura, L.; Pinto, A.; Giusti, A.; Mengheri, E. Supplementation with Bifidobacterium longum Bar33 and Lactobacillus helveticus Bar13 mixture improves immunity in elderly humans (over 75 years) and aged mice. Nutrition 2019, 63–64, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Pahumunto, N.; Sophatha, B.; Piwat, S.; Teanpaisan, R. Increasing salivary IgA and reducing Streptococcus mutans by probiotic Lactobacillus paracasei SD1: A double-blind, randomized, controlled study. J. Dent. Sci. 2019, 14, 178–184. [Google Scholar] [CrossRef]
- Wang, M.; Wu, H.; Lu, L.; Jiang, L.; Yu, Q. Lactobacillus reuteri Promotes Intestinal Development and Regulates Mucosal Immune Function in Newborn Piglets. Front. Veter Sci. 2020, 7, 42. [Google Scholar] [CrossRef]
- Garcia-Castillo, V.; Komatsu, R.; Clua, P.; Indo, Y.; Takagi, M.; Salva, S.; Islam, A.; Alvarez, S.; Takahashi, H.; Garcia-Cancino, A.; et al. Evaluation of the Immunomodulatory Activities of the Probiotic Strain Lactobacillus fermentum UCO-979C. Front. Immunol. 2019, 10, 1376. [Google Scholar] [CrossRef]
- Foysal, J.; Fotedar, R.; Siddik, M.A.B.; Tay, A. Lactobacillus acidophilus and L. plantarum improve health status, modulate gut microbiota and innate immune response of marron (Cherax cainii). Sci. Rep. 2020, 10, 1–13. [Google Scholar] [CrossRef]
- Wu, Z.; Yang, K.; Zhang, A.; Chang, W.; Zheng, A.; Chen, Z.; Cai, H.; Liu, G. Effects of Lactobacillus acidophilus on the growth performance, immune response, and intestinal barrier function of broiler chickens challenged with Escherichia coli O157. Poult. Sci. 2021, 100, 101323. [Google Scholar] [CrossRef]
- Kosgey, J.C.; Jia, L.; Fang, Y.; Yang, J.; Gao, L.; Wang, J.; Nyamao, R.; Cheteu, M.; Tong, D.; Wekesa, V.; et al. Probiotics as antifungal agents: Experimental confirmation and future prospects. J. Microbiol. Methods 2019, 162, 28–37. [Google Scholar] [CrossRef]
- Kesika, P.; Sivamaruthi, B.S.; Thangaleela, S.; Chaiyasut, C. The Antiviral Potential of Probiotics—A Review on Scientific Outcomes. Appl. Sci. 2021, 11, 8687. [Google Scholar] [CrossRef]
- Rezaee, P.; Kermanshahi, R.K.; Falsafi, T. Antibacterial activity of lactobacilli probiotics on clinical strains of Helicobacter pylori. Iran. J. Basic Med. Sci. 2019, 22, 1118–1124. [Google Scholar] [CrossRef] [PubMed]
- I Moré, M.; Vandenplas, Y. Saccharomyces boulardii CNCM I-745 Improves Intestinal Enzyme Function: A Trophic Effects Review. Clin. Med. Insights: Gastroenterol. 2018, 11, 1179552217752679. [Google Scholar] [CrossRef] [PubMed]
- Dahiya, D.; Nigam, P.S. Antibiotic-Therapy-Induced Gut Dysbiosis Affecting Gut Microbiota—Brain Axis and Cognition: Restoration by Intake of Probiotics and Synbiotics. Int. J. Mol. Sci. 2023, 24, 3074. [Google Scholar] [CrossRef] [PubMed]
- Matsubara, V.H.; Fakhruddin, K.S.; Ngo, H.; Samaranayake, L.P. Probiotic Bifidobacteria in Managing Periodontal Disease: A Systematic Review. Int. Dent. J. 2022, 73, 11–20. [Google Scholar] [CrossRef]
- Knackstedt, R.; Knackstedt, T.; Gatherwright, J. The role of topical probiotics in skin conditions: A systematic review of animal and human studies and implications for future therapies. Exp. Dermatol. 2020, 29, 15–21. [Google Scholar] [CrossRef]
- Pourmasoumi, M.; Najafgholizadeh, A.; Hadi, A.; Mansour-Ghanaei, F.; Joukar, F. The effect of synbiotics in improving Helicobacter pylori eradication: A systematic review and meta-analysis. Complement. Ther. Med. 2019, 43, 36–43. [Google Scholar] [CrossRef]
- Tidbury, F.D.; Langhart, A.; Weidlinger, S.; Stute, P. Non-antibiotic treatment of bacterial vaginosis—A systematic review. Arch. Gynecol. Obstet. 2021, 303, 37–45. [Google Scholar] [CrossRef]
- Huang, R.; Xing, H.-Y.; Liu, H.-J.; Chen, Z.-F.; Tang, B.-B. Efficacy of probiotics in the treatment of acute diarrhea in children: A systematic review and meta-analysis of clinical trials. Transl. Pediatr. 2021, 10, 3248–3260. [Google Scholar] [CrossRef]
- Ma, Y.; Yang, J.Y.; Peng, X.; Xiao, K.Y.; Xu, Q.; Wang, C. Which probiotic has the best effect on preventing Clostridium difficile-associated diarrhea? A systematic review and network meta-analysis. J. Dig. Dis. 2020, 21, 69–80. [Google Scholar] [CrossRef]
- Piewngam, P.; Khongthong, S.; Roekngam, N.; Theapparat, Y.; Sunpaweravong, S.; Faroongsarng, D.; Otto, M. Probiotic for pathogen-specific Staphylococcus aureus decolonisation in Thailand: A phase 2, double-blind, randomised, placebo-controlled trial. Lancet Microbe 2023, 4, e75–e83. [Google Scholar] [CrossRef]
- Liao, H.; Liu, S.; Wang, H.; Su, H.; Liu, Z. Enhanced antifungal activity of bovine lactoferrin-producing probiotic Lactobacillus casei in the murine model of vulvovaginal candidiasis. BMC Microbiol. 2019, 19, 7. [Google Scholar] [CrossRef] [PubMed]
- Khmaladze, I.; Butler, É.; Fabre, S.; Gillbro, J.M. Lactobacillus reuteri DSM 17938—A comparative study on the effect of probiotics and lysates on human skin. Exp. Dermatol. 2019, 28, 822–828. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.-H.; Kim, B.S.; Kang, S.-S. Bacteriocin of Pediococcus acidilactici HW01 Inhibits Biofilm Formation and Virulence Factor Production by Pseudomonas aeruginosa. Probiotics Antimicrob. Proteins 2020, 12, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Bin Lee, H.; Kim, K.H.; Kang, G.A.; Lee, K.-G.; Kang, S.-S. Antibiofilm, AntiAdhesive and Anti-Invasive Activities of Bacterial Lysates Extracted from Pediococcus acidilactici against Listeria monocytogenes. Foods 2022, 11, 2948. [Google Scholar] [CrossRef]
- Invernici, M.M.; Salvador, S.L.; Silva, P.H.; Soares, M.S.; Casarin, R.; Palioto, D.B.; Souza, S.L.; Taba, M., Jr.; Novaes, A.B., Jr.; Furlaneto, F.A. Effects of Bifidobacterium Probiotic on the Treatment of Chronic Periodontitis: A Randomized Clinical Trial. J. Clin. Periodontol. 2018, 45, 1198–1210. [Google Scholar] [CrossRef]
- Michelotti, A.; Cestone, E.; De Ponti, I.; Giardina, S.; Pisati, M.; Spartà, E.; Tursi, F. Efficacy of a probiotic supplement in patients with atopic dermatitis: A randomized, double-blind, placebo-controlled clinical trial. Eur. J. Dermatol. 2021, 31, 225–232. [Google Scholar] [CrossRef]
- Rahmayani, T.; Putra, I.B.; Jusuf, N.K. The Effect of Oral Probiotic on the Interleukin-10 Serum Levels of Acne Vulgaris. Open Access Maced. J. Med Sci. 2019, 7, 3249–3252. [Google Scholar] [CrossRef]
- Zaharuddin, L.; Mokhtar, N.M.; Nawawi, K.N.M.; Ali, R.A.R. A randomized double-blind placebo-controlled trial of probiotics in post-surgical colorectal cancer. BMC Gastroenterol. 2019, 19, 131. [Google Scholar] [CrossRef]
- Tsilika, M.; Thoma, G.; Aidoni, Z.; Tsaousi, G.; Fotiadis, K.; Stavrou, G.; Malliou, P.; Chorti, A.; Massa, H.; Antypa, E.; et al. A four-probiotic preparation for ventilator-associated pneumonia in multi-trauma patients: Results of a randomized clinical trial. Int. J. Antimicrob. Agents 2022, 59, 106471. [Google Scholar] [CrossRef]
- Mageswary, M.U.; Ang, X.-Y.; Lee, B.-K.; Chung, Y.-L.F.; Azhar, S.N.A.; Hamid, I.J.A.; Abu Bakar, H.; Roslan, N.S.; Liu, X.; Kang, X.; et al. Probiotic Bifidobacterium lactis Probio-M8 treated and prevented acute RTI, reduced antibiotic use and hospital stay in hospitalized young children: A randomized, double-blind, placebo-controlled study. Eur. J. Nutr. 2022, 61, 1679–1691. [Google Scholar] [CrossRef]
- Ahrén, I.L.; Hillman, M.; Nordström, E.A.; Larsson, N.; Niskanen, T.M. Fewer Fewer community-acquired colds with daily consumption of Lactiplantibacillus plantarum HEAL9 and Lacticaseibacillus paracasei 8700: 2. A randomized, placebo-controlled clinical trial. J. Nutr. 2021, 151, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Liang, B.; Yuan, Y.; Peng, X.-J.; Liu, X.-L.; Hu, X.-K.; Xing, D.-M. Current and future perspectives for Helicobacter pylori treatment and management: From antibiotics to probiotics. Front. Cell. Infect. Microbiol. 2022, 12, 1740. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.H.; Jang, Y.S.; Kang, D.; Chang, D.K.; Min, Y.W. Efficacy and Safety of New Lactobacilli Probiotics for Unconstipated Irritable Bowel Syndrome: A Randomized, Double-Blind, Placebo-Controlled Trial. Nutrients 2019, 11, 2887. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Feng, D. Therapeutic effect of Saccharomyces boulardii combined with Bifidobacterium and on cellular immune function in children with acute diarrhea. Exp. Ther. Med. 2019, 18, 2653–2659. [Google Scholar] [CrossRef]
- Cohen, C.R.; Wierzbicki, M.R.; French, A.L.; Morris, S.; Newmann, S.; Reno, H.; Green, L.; Miller, S.; Powell, J.; Parks, T.; et al. Randomized Trial of Lactin-V to Prevent Recurrence of Bacterial Vaginosis. N. Engl. J. Med. 2020, 382, 1906–1915. [Google Scholar] [CrossRef]
- Roth, N.; Käsbohrer, A.; Mayrhofer, S.; Zitz, U.; Hofacre, C.; Domig, K.J. The application of antibiotics in broiler production and the resulting antibiotic resistance in Escherichia coli: A global overview. Poult. Sci. 2019, 98, 1791–1804. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, R.; Jia, H.; Zhu, Z.; Li, H.; Ma, Y. Supplementation of probiotics in water beneficial growth performance, carcass traits, immune function, and antioxidant capacity in broiler chickens. Open Life Sci. 2021, 16, 311–322. [Google Scholar] [CrossRef]
- Lokapirnasari, W.P.; Pribadi, T.B.; Al Arif, A.; Soeharsono, S.; Hidanah, S.; Harijani, N.; Najwan, R.; Huda, K.; Wardhani, H.C.P.; Rahman, N.F.N.; et al. Potency of probiotics Bifidobacterium spp. and Lactobacillus casei to improve growth performance and business analysis in organic laying hens. Veter World 2019, 12, 860–867. [Google Scholar] [CrossRef]
- Wang, L.; Li, L.; Lv, Y.; Chen, Q.; Feng, J.; Zhao, X. Lactobacillus plantarum Restores Intestinal Permeability Disrupted by Salmonella Infection in Newly-hatched Chicks. Sci. Rep. 2018, 8, 2229. [Google Scholar] [CrossRef]
- Fesseha, H.; Demlie, T.; Mathewos, M.; Eshetu, E. Effect of Lactobacillus Species Probiotics on Growth Performance of Dual-Purpose Chicken. Veter Med. Res. Rep. 2021, 12, 75–83. [Google Scholar] [CrossRef]
- Wang, H.; Ni, X.; Qing, X.; Liu, L.; Xin, J.; Luo, M.; Khalique, A.; Dan, Y.; Pan, K.; Jing, B.; et al. Probiotic Lactobacillus johnsonii BS15 Improves Blood Parameters Related to Immunity in Broilers Experimentally Infected with Subclinical Necrotic Enteritis. Front. Microbiol. 2018, 9, 49. [Google Scholar] [CrossRef] [PubMed]
- Kan, L.; Guo, F.; Liu, Y.; Pham, V.H.; Guo, Y.; Wang, Z. Probiotics Bacillus licheniformis Improves Intestinal Health of Subclinical Necrotic Enteritis-Challenged Broilers. Front. Microbiol. 2021, 12, 623739. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Peng, X.-Y.; Gao, B.; Wei, Q.-L.; Xiang, R.; Yuan, M.-G.; Xu, Z.-H. The Effect of Clostridium butyricum on Gut Microbiota, Immune Response and Intestinal Barrier Function during the Development of Necrotic Enteritis in Chickens. Front. Microbiol. 2019, 10, 2309. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Yoo, D.H.; Ao, X.; Kim, I.H. Effects of dietary probiotic, liquid feed and nutritional concentration on the growth performance, nutrient digestibility and fecal score of weaning piglets. Asian-Australas. J. Anim. Sci. 2020, 33, 1617–1623. [Google Scholar] [CrossRef]
- Pupa, P.; Apiwatsiri, P.; Sirichokchatchawan, W.; Pirarat, N.; Nedumpun, T.; Hampson, D.J.; Muangsin, N.; Prapasarakul, N. Microencapsulated probiotic Lactiplantibacillus plantarum and/or Pediococcus acidilactici strains ameliorate diarrhoea in piglets challenged with enterotoxigenic Escherichia coli. Sci. Rep. 2022, 12, 7210. [Google Scholar] [CrossRef]
- Yasmin, F.; Alam, M.J.; Kabir, M.E.; Al Maruf, A.; Islam, M.A.; Hossain, M.M. Influence of Probiotics supplementation on Growth and Haemato-biochemical Parameters in Growing Cattle. Int. J. Livest. Res. 2021, 11, 36–42. [Google Scholar] [CrossRef]
- Merati, Z.; Towhidi, A. Effect of a Multispecies Probiotics on Productive and Reproductive Performance of Holstein Cows. Iran. J. Appl. Anim. Sci. 2022, 12, 237–247. [Google Scholar]
- Genís, S.; Sánchez-Chardi, A.; Bach, À.; Fàbregas, F.; Arís, A. A combination of lactic acid bacteria regulates Escherichia coli infection and inflammation of the bovine endometrium. J. Dairy Sci. 2017, 100, 479–492. [Google Scholar] [CrossRef]
- Devyatkin, V.; Mishurov, A.; Kolodina, E. Probiotic effect of Bacillus subtilis B-2998D, B-3057D, and Bacillus licheniformis B-2999D complex on sheep and lambs. J. Adv. Veter Anim. Res. 2021, 8, 146–157. [Google Scholar] [CrossRef]
- Islam, S.M.; Rohani, F. Shahjahan Probiotic yeast enhances growth performance of Nile tilapia (Oreochromis niloticus) through morphological modifications of intestine. Aquac. Rep. 2021, 21, 100800. [Google Scholar] [CrossRef]
- Cavalcante, R.B.; Telli, G.S.; Tachibana, L.; Dias, D.D.C.; Oshiro, E.; Natori, M.M.; da Silva, W.F.; Ranzani-Paiva, M.J. Probiotics, Prebiotics and Synbiotics for Nile tilapia: Growth performance and protection against Aeromonas hydrophila infection. Aquac. Rep. 2020, 17, 100343. [Google Scholar] [CrossRef]
- Ahmadifar, E.; Sadegh, T.H.; Dawood, M.A.; Dadar, M.; Sheikhzadeh, N. The effects of dietary Pediococcus pentosaceus on growth performance, hemato-immunological parameters and digestive enzyme activities of common carp (Cyprinus carpio). Aquaculture 2020, 516, 734656. [Google Scholar] [CrossRef]
- Saravanan, K.; Sivaramakrishnan, T.; Praveenraj, J.; Kiruba-Sankar, R.; Haridas, H.; Kumar, S.; Varghese, B. Effects of single and multi-strain probiotics on the growth, hemato-immunological, enzymatic activity, gut morphology and disease resistance in Rohu, Labeo rohita. Aquaculture 2021, 540, 736749. [Google Scholar] [CrossRef]
- Won, S.; Hamidoghli, A.; Choi, W.; Bae, J.; Jang, W.J.; Lee, S.; Bai, S.C. Evaluation of Potential Probiotics Bacillus subtilis WB60, Pediococcus pentosaceus, and Lactococcus lactis on Growth Performance, Immune Response, Gut Histology and Immune-Related Genes in Whiteleg Shrimp, Litopenaeus vannamei. Microorganisms 2020, 8, 281. [Google Scholar] [CrossRef] [PubMed]
- Kewcharoen, W.; Srisapoome, P. Probiotic effects of Bacillus spp. from Pacific white shrimp (Litopenaeus vannamei) on water quality and shrimp growth, immune responses, and resistance to Vibrio parahaemolyticus (AHPND strains). Fish Shellfish. Immunol. 2019, 94, 175–189. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibiotics | Example | Source | Reference |
|---|---|---|---|
| Aminoglycosides | Streptomycin | Streptomyces griseus | [20] |
| Β-Lactams | Penicillin | Penicillium griseofulvum | [21] |
| Glycopeptides | Vancomycin | Amycolatopsis orientalis | [22] |
| Lipopetides | Daptomycin | Streptomyces roseosporus | [23] |
| Macrolides | Erythromycin | Streptomyces erythreus | [24] |
| Oxazolidinones | Linezolid | Chemical synthesis | [19] |
| Phenicols | Chloramphenicol | Streptomyces venezuelae | [25] |
| Polypeptides | Gramicidin | Bacillus brevis | [26] |
| Polymixin | Colistin | Paenibacillus polymyxa | [27] |
| Quinolones | Ciproxacin | Chemical synthesis | [19] |
| Sulfonamides | Mafenide | Chemical synthesis | [19] |
| Tetracyclines | Oxytetracyclines | Streptomyces rimosus | [28] |
| Class | Mode of Action | Resistance Mechanism | Reference |
|---|---|---|---|
| Aminoglycosides | Inhibition of protein synthesis (30S ribosomal subunit inhibitor) | Binding inhibition by phosphorylation, adenylation, and acetylation of aminoglycosides Aminoglycoside-modifying enzymes (e.g., acetyltransferases, phosphotransferases) 16S rRNA methylation Efflux-mediated resistance | [41,42,43] |
| β-Lactams | Inhibition of cell wall synthesis (peptidoglycan) | Production of β-Lactamases Permeability change (Efflux) | [41,42,44] |
| Glycopeptides | Inhibition of cell wall synthesis (peptidoglycans) | Intrinsic resistance in Gram-negative cells by impermeable outer membrane Presence of enzymes that modify and hydrolyze peptidoglycan precursors Low permeability | [41,42,45] |
| Fluoroquinolones | Inhibition of nucleic acid synthesis | Mutations in DNA gyrase or topoisomerase IV | [41,42,46] |
| Sulfonamides | Blockage of key metabolic pathways (folate synthesis inhibitors) | Mutations in folP gene encoding dihydropteroate synthase, sul1, sul2 genes, sulfonamide monooxygenase gene sulX | [41,42,47] |
| Tetracyclines | Inhibition of protein synthesis (30S-ribosomal subunit inhibitor) | Enzymatic inactivation Binding site mutation | [41,42,48] |
| Chloramphenicols | Inhibition of protein synthesis (50S-ribosomal subunit inhibitor) | Mutations within 23S rRNA of the 50S ribosomal subunit Enzymatic inactivation via acetyltransferases Active efflux | [41,42,49] |
| Bacteriocins | Probiotic | Target Microorganisms | Reference |
|---|---|---|---|
| Bacteriocin | L. acidophilus KS400 | Gardnerella vaginalis, Streptococcus agalactiae, P. aeruginosa | [79] |
| Enterocin M | Enterococcus faecium AL41 | Campylobacter spp. Clostridium spp. | [80] |
| Nisin-like bacteriocin | L. lactis C15 | E. coli | [81] |
| Pediocin | Ped. pentosaceus GS4 (MTCC 12683) | S. aureus (ATCC 25923), E. coli (ATCC 25922), P. aeruginosa (ATCC 25619), and L. monocytogenes (ATCC 15313) | [78] |
| Plantaricin P1053 | L. plantarum PBS067 | S. aureus and E. coli | [82] |
| Subtilin-like bacteriocin—Subtilin JS-4 | Bacillus subtilis JS-4 | L. monocytogenes | [83] |
| Probiotics | Studied Model | Effects on Immunity | Reference |
|---|---|---|---|
| Bifidobacterium longum Bar33 and L. helveticus Bar13 | Older adults (over 75 years) | Increase naive T cells Increase activated memory, regulatory T cells, B cells, and natural killer (NK) activity Decrease memory T cells | [105] |
| L. paracasei SD1 | Children | Decrease of Streptococcus mutans pathogens Increase of salivary IgA | [106] |
| Limosilactobacillus reuteri D8 | Piglets | Increase of goblet cells and antimicrobial peptides (AMPs), expressions of Muc2, Lyz1, and porcine β-defensins 1 (pBD1) Increase of CD3+ T cells, combined with increased expression of IL-4 and IFN-γ | [107] |
| Lactobacillus fermentum UCO-979C | Mice | Increase the production of intestinal IFN-γ, stimulate intestinal and peritoneal macrophages, increase the number of Peyer’s patches CD4+ T cells Increase intestinal IL-6, intestinal IgA, and the number of mature B cells | [108] |
| L. acidophilus and L. plantarum | Freshwater crayfish | Upregulation of cytokine gene families (IL1β, IL8, IL10, and IL17F), proPO, and cytMnSOD | [109] |
| L. acidophilus | Broilers challenged with E. coli | Reduce the mortality rate caused by E. coli challenge Decrease the serum C-reactive protein, diamine oxydase, and endotoxin lipopolysaccharide levels at 14 days and 21 days Upregulate the mRNA expression of occludin and zona occludens protein 1 (ZO-1) in the jejunum and ileum (tight junction) Downregulate the mRNA expression of inducible nitric oxide synthase (iNOS), IL-8, and IL-1β in the jejunum in E. coli challenged birds at 21 days | [110] |
| Antibiotic | Probiotic | |
|---|---|---|
| Characteristic features | Active substance Natural or synthetic One function Non-growth over time (static process) | Live microorganism Natural Multifunction Growth over time (dynamic process) |
| Action mechanisms | Cell membrane breakdown Cell wall synthesis inhibition Nucleic acid structure/function and protein synthesis inhibition Key metabolic pathway blockage | Gut barrier protection Nutrient/space competitive exclusion Antimicrobial substance secretion Immunomodulation |
| Strength | Specificity Short-time treatment | No side effects Antibacterial and antiviral properties Generally recognized as safe (GRAS) Natural and biodegradable |
| Weakness | Destroy beneficial microbes Antimicrobial resistance induction Not effective on viruses Low biodegradability for synthetic compounds | Cell viability maintaining challenge Long-term treatment Sensitivity under stress conditions Antimicrobial resistance risk if genes transfer |
| Probiotics | Pathogens | Observation | Reference |
|---|---|---|---|
| B. subtilis MB40 | S. aureus | Significant reduction of S. aureus colonization in body human without modification of microbiome | [122] |
| L. casei | C. albicans | Fungicidal effect in vulvovaginal candidiasis (VVC) murine model | [123] |
| L. reuteri DSM 17938 | S. aureus, S. pyogenes M1, Cutibacterium acnes AS12, P. aeruginosa | Antimicrobial action against pathogenic skin bacteria and reduction of proinflammatory IL-6 and IL-8 in reconstructed human epidermis and native skin models | [124] |
| Pediococcus acidilactici HW01 | P. aeruginosa | Inhibition of biofilm formation by bacteriocin and decrease of the production of virulence factors, such as pyocyanin, protease, and rhamnolipid | [125] |
| Ped. acidilactici HW01 | L. monocytogenes | Inhibition of biofilm formation, adhesion, and invasion of HT-29 cells (human-intestinal-epithelial cell line) by bacterial lysate | [126] |
| Disease | Probiotics | Outcome | Reference | |
|---|---|---|---|---|
| Oral and dental health | Chronic periodontitis | B. animalis subsp. lactis (B. lactis) HN019 | Decreasing significantly the periodontal pathogens of red and orange complexes; reducing proinflammatory cytokine levels; promoting clinical, microbiological, and immunological benefits in the treatment of chronic periodontitis | [127] |
| Skin | Atopic dermatitis | L. plantarum PBS067 L. reuteri PBS072 L. rhamnosus LRH020 | Improvement in skin smoothness, skin moisturization, self-perception, and decrease in scoring atopic dermatitis (SCORAD) index and levels of inflammatory markers | [128] |
| Acne vulgaris | B. lactis W51, B. lactis W52, L. acidophilus W55, L. casei W56, L. salivarius W57, L. lactis W58 combined with rice starch and maltodextrin | Increasing serum IL-10 levels after oral probiotic in acne vulgaris | [129] | |
| Surgical wound infection | L. acidophilus BCMC® 12130 L. lactis BCMC® 12451, L. casei subsp BCMC® 12313, B. longum BCMC® 02120, B. bifidum BCMC® 02290, and B. infantis BCMC® 02129 | Reduction of pro-inflammatory cytokines (except for IFN-gamma) in colorectal cancer patients after consumption for 4 weeks | [130] | |
| Respiratory tract | Ventilator-associated pneumonia (VAP) | L. acidophilus LA-5, L. plantarum UBLP-40, B. animalis subsp. lactis BB-12, and S. boulardii | Decreasing the incidence of VAP induced by Acinobacter baumannii and P. aeruginosa in patients subjected to prolonged mechanical ventilation for severe multiple trauma, including brain injury | [131] |
| Acute respiratory tract infection | B. lactis Probio-M8 | Reducing antibiotic prescription, preventing antibiotic new prescription in non-prescribed patients, decreasing oral cytokine levels of TNF-α, and increased IL-10 (over 4 weeks post-discharge) | [132] | |
| Virus associated respiratory tract infection | L. plantarum HEAL9 L. paracasei 8700 | No effect on symptom severity but significantly fewer colds | [133] | |
| Stomach | Helicobacter pylori infection, gastritis | L. reuteri DSM 17648 | Effectively reducing H. pylori load and improving gastrointestinal symptoms in adults and children | [134] |
| Intestines | Inflammatory bowel syndrome (IBS) | L. paracasei, L. salivarius, and L. plantarum | Effective global relief of IBS symptoms and abdominal pain without significant adverse events | [135] |
| C. difficile-associated diarrhea (CDAD) | L. casei | Reduction of the incidence rates of CDAD | [121] | |
| Acute diarrhea | S. boulardii combined with bifidobacterium | Shortening the duration of diarrhea and hospital stay, reducing the number of diarrhea, enhancing cellular immune function | [136] | |
| Female urogenital tract | Bacterial vaginosis | L. crispatus CTV-05 (Lactin-V) | Prevention of the recurrence of bacterial vaginosis | [137] |
| Animals | Probiotics | Form of Administration | Effects | Reference |
|---|---|---|---|---|
| Poultry | ||||
| Broilers | L. casei, L. acidophilus, and Bifidobacterium | Supplementing 1% of probiotics in water | Increasing growth performance, carcass traits, immune function, gut microbial population, and antioxidant capacity | [139] |
| Laying hens | Bifidobacterium spp. and L. casei | Feeding | Improving the growth performance, increase of egg weight, and feed efficiency | [140] |
| Newly hatched chicks | L. plantarum LTC-113 | Oral vaccination | Protection from Salmonella colonization by regulating expression of tight junction genes and inflammatory mediators | [141] |
| Chickens | L. paracasei ssp. paracasei and L. rhamnosus | Feeding | Improving growth performance | [142] |
| Broiler | L. johnsonii BS15 | Feeding | Preventing subclinical necrotic enteritis | [143] |
| Bacillus licheniformis | Feeding | Alleviating intestinal damage caused by SNE challenge, modulating intestinal microflora structure and barrier function, and regulating intestinal mucosal immune responses | [144] | |
| Chickens | C. butyricum | Feeding | Promoting anti-inflammatory expression and tight junction protein genes Inhibiting pro-inflammatory genes in C. perfringens-challenged chickens | [145] |
| Swine | ||||
| Weaning piglets | B. subtilis, E. faecium | Liquid feed | Improve growth performance | [146] |
| Piglets | L. plantarum (strains 22F and 25F) and Ped. acidilactici (strain 72N) | Feeding | Reducing the infection severity with enterotoxigenic E. coli (ETEC) in weaned pigs | [147] |
| Cattle | ||||
| Cattle | L. gallinarum JCM 2011(T), S. infantarius subsp. coli HDP90246 (T), S. salivarius subsp. thermophilus ATCC 19258(T), and
S. equinus ATCC 9812(T) Saccharomyces cerevisiae | Improving the growth and haemato-biochemical parameters of growing cattle | [148] | |
| Dairy cows | S. cerevisiae, B. subtilis, B. lichenformis, E. faecium, L. acidophilus, L. plantarum, B. tedium and calcium carbonate | Feeding | Improving reproductive performance Increasing milk yield and milk fat and protein percentage | [149] |
| L. rhamnosus, P. acidilactici, and L. reuteri | Ex vivo bovine endometrial explants | Reducing acute inflammation under E. coli infection, decreasing IL-8, IL-1β, and IL-6 | [150] | |
| Sheep | ||||
| Sheep, Lamb | Enzimsporin™ (B. subtilis B-2998D, B-3057D, and B. licheniformis B-2999D) | Feeding | Increasing body weight gain and improving intestinal microbiota | [151] |
| Fish | ||||
| Nile Tilapia (Oreochromis niloticus) | S. cerevisiae | Feeding | Increasing growth performance and feed utilization indices | [152] |
| Nile Tilapia (Oreochromis niloticus) | DBA® (B. sp., L. acidophilus and E. faecium) | Feeding | Protection against A. hydrophila infection without growth reduction | [153] |
| Common carp (Cyprinus carpio) | Ped. pentosaceus | Feeding | Improving growth performance, digestive enzyme activity, and haemato-immunological responses | [154] |
| Rohu fingerlings (Labeo rohita) | B. amyloliquefaciens BN06, B. subtilis WN07, and B. megaterium | Feeding | Improving growth and haemato-immunological parameters | [155] |
| Shrimp | ||||
| Whiteleg shrimp, (Litopenaeus vannamei) | B. subtilis, Ped. pentosaceus, and L. lactis | Feeding | Improving growth, immunity, histology, gene expression, digestive enzyme activity, and disease resistance | [156] |
| Pacific white shrimp (Litopenaeus vannamei) | B. subtilis AQAHBS001 | Feeding | Improving the growth performance, immune response, and resistance to Vibrio parahaemolyticus | [157] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rabetafika, H.N.; Razafindralambo, A.; Ebenso, B.; Razafindralambo, H.L. Probiotics as Antibiotic Alternatives for Human and Animal Applications. Encyclopedia 2023, 3, 561-581. https://doi.org/10.3390/encyclopedia3020040
Rabetafika HN, Razafindralambo A, Ebenso B, Razafindralambo HL. Probiotics as Antibiotic Alternatives for Human and Animal Applications. Encyclopedia. 2023; 3(2):561-581. https://doi.org/10.3390/encyclopedia3020040
Chicago/Turabian StyleRabetafika, Holy N., Aurélie Razafindralambo, Bassey Ebenso, and Hary L. Razafindralambo. 2023. "Probiotics as Antibiotic Alternatives for Human and Animal Applications" Encyclopedia 3, no. 2: 561-581. https://doi.org/10.3390/encyclopedia3020040
APA StyleRabetafika, H. N., Razafindralambo, A., Ebenso, B., & Razafindralambo, H. L. (2023). Probiotics as Antibiotic Alternatives for Human and Animal Applications. Encyclopedia, 3(2), 561-581. https://doi.org/10.3390/encyclopedia3020040