
Growth and Welfare of African Catfish (Clarias gariepinus Burchell, 1822) under Dietary Supplementation with Mixed-Layer Clay Mineral Montmorillonite-Illite/Muscovite in Commercial Aquaculture

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Production Systems and Maintenance
2.2. Experimental Feeds
2.3. Fish Stocking and Feeding
2.4. Sampling
2.5. Blood Parameters and External Injuries
2.6. Statistics
3. Results
3.1. Fish Growth Performance
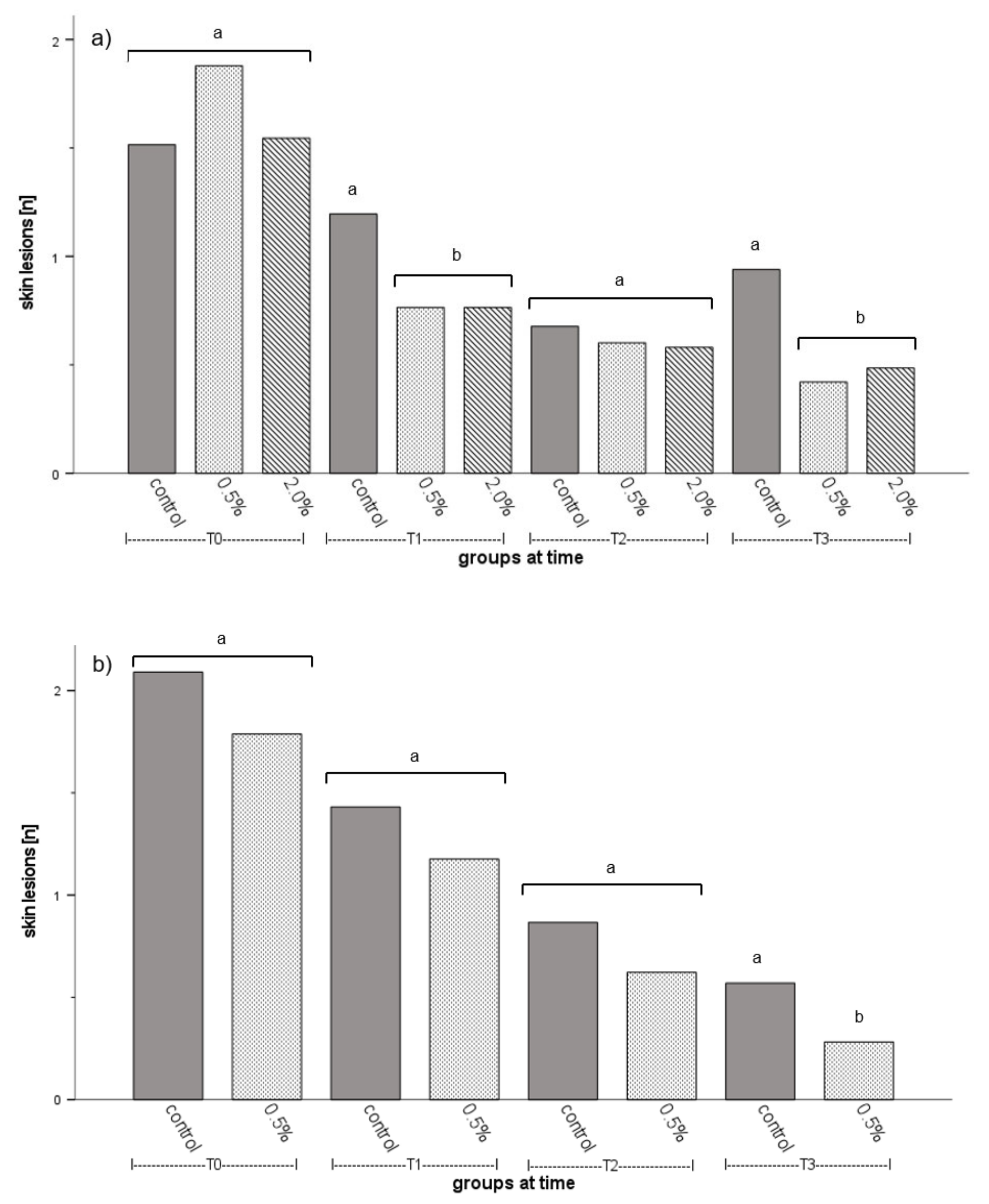
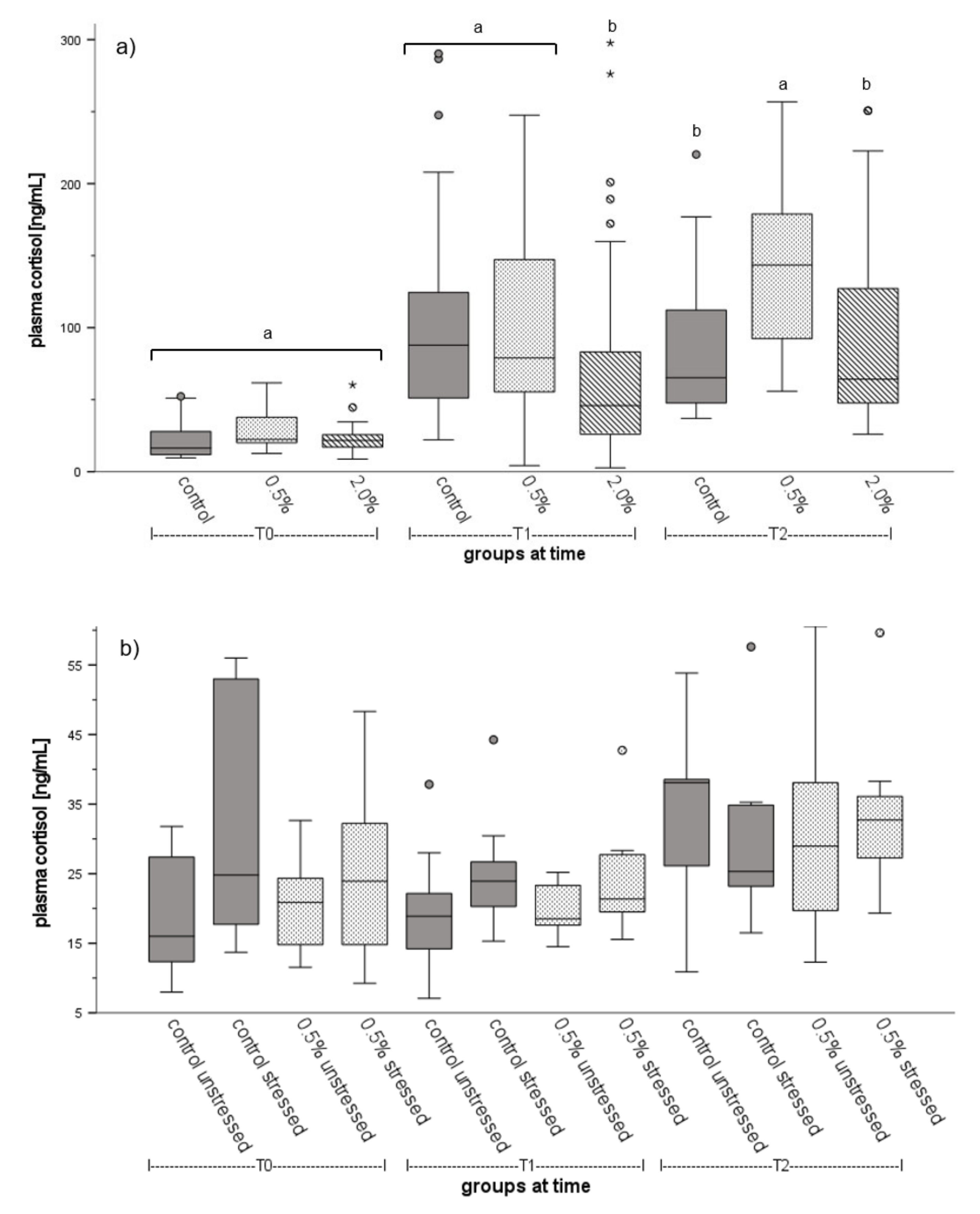
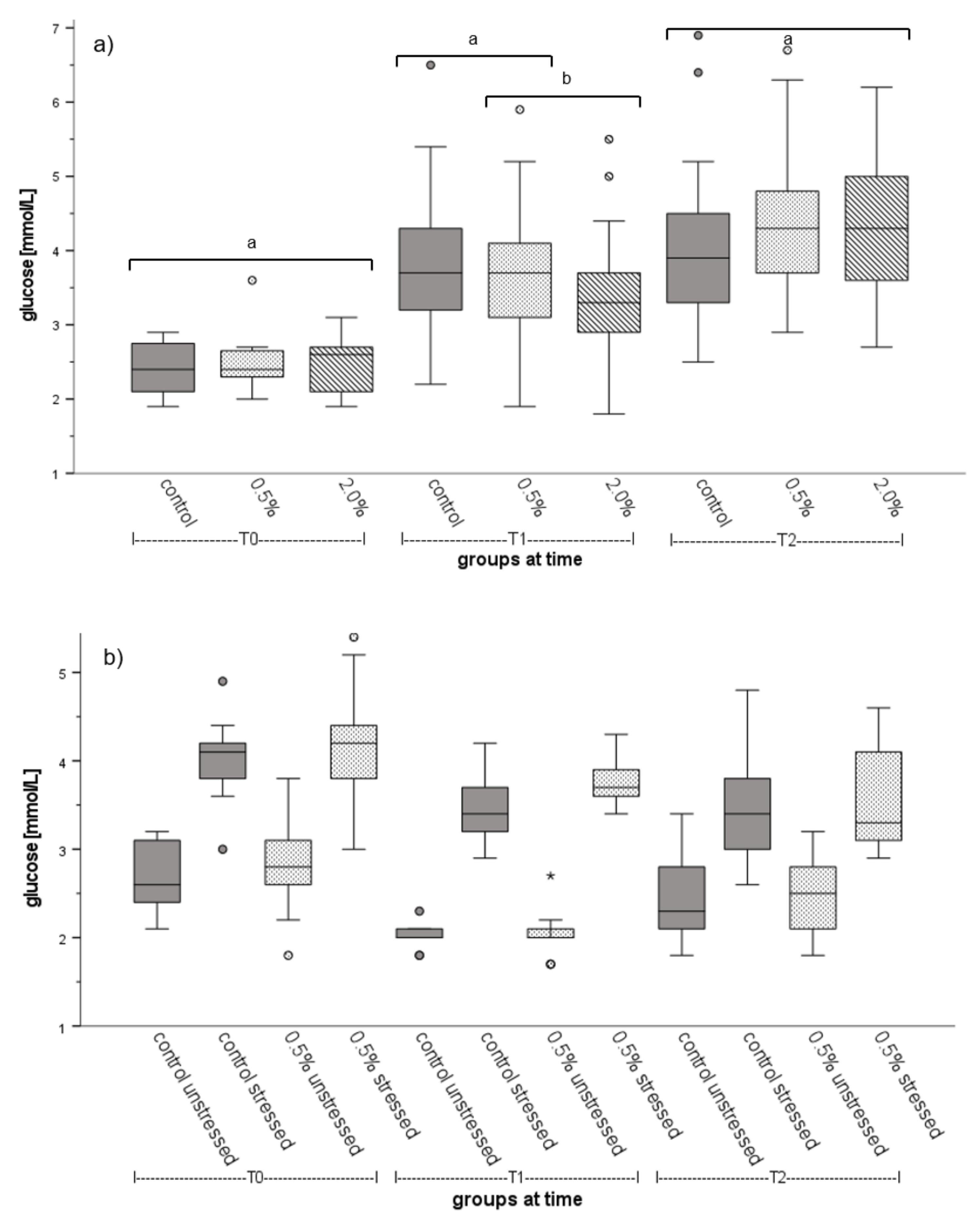
3.2. Fish Welfare
4. Discussion
4.1. Aquaculture Conditions
4.2. Fish Growth
4.3. Fish Welfare
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Destatis, Statistisches Bundesamt. Land und Forstwirtschaft, Fischerei. Erzeugung in Aquakulturbetrieben 2020. Available online: https://www.statistischebibliothek.de/mir/servlets/MCRFileNodeServlet/DEHeft_derivate_00061752/2030460207004.pdf (accessed on 14 February 2022).
- Destatis, Statistisches Bundesamt. Land und Forstwirtschaft, Fischerei. Erzeugung in Aquakulturbetrieben 2012. Available online: https://www.statistischebibliothek.de/mir/servlets/MCRFileNodeServlet/DEHeft_derivate_00009985/2030460127004.pdf (accessed on 14 February 2022).
- Belão, T.C.; Leite, C.A.C.; Florindo, L.H.; Kalinin, A.L.; Rantin, F.T. Cardiorespiratory responses to hypoxia in the African catfish, Clarias gariepinus (Burchell 1822), an air-breathing fish. J. Comp. Physiol. B 2011, 181, 905. [Google Scholar] [CrossRef] [PubMed]
- Schram, E.; Roques, J.A.; Abbink, W.; Yokohama, Y.; Spanings, T.; de Vries, P.; Bierman, S.; van de Vis, H.; Flik, G. The impact of elevated water nitrate concentration on physiology, growth and feed intake of African catfish Clarias gariepinus (Burchell 1822). Aquac. Res. 2014, 45, 1499–1511. [Google Scholar] [CrossRef]
- Roques, J.A.; Schram, E.; Spanings, T.; van Schaik, T.; Abbink, W.; Boerrigter, J.; de Vries, P.; van de Vis, H.; Flik, G. The impact of elevated water nitrite concentration on physiology, growth and feed intake of African catfish Clarias gariepinus (Burchell 1822). Aquac. Res. 2015, 46, 1384–1395. [Google Scholar] [CrossRef]
- Păpuc, T.; Petrescu-Mag, I.V.; Gavriloaie, C.; Botha, M.; Kovacs, E.; Coroian, C.O. Swimming in the mud—A short review of environmental parameter ranges tolerated by Clarias Gariepinus. Extrem. Life Biospeology Astrobiol. 2019, 11, 9–17. [Google Scholar]
- Van de Nieuwegiessen, P.G.; Olwo, J.; Khong, S.; Verreth, J.A.; Schrama, J.W. Effects of age and stocking density on the welfare of African catfish, Clarias gariepinus Burchell. Aquaculture 2009, 288, 69–75. [Google Scholar] [CrossRef]
- Martins, C.I.; Schrama, J.W.; Verreth, J.A. The effect of group composition on the welfare of African catfish (Clarias gariepinus). Appl. Anim. Behav. Sci. 2006, 97, 323–334. [Google Scholar] [CrossRef]
- Martins, C.I.; Schrama, J.W.; Verreth, J.A. The relationship between individual differences in feed efficiency and stress response in African catfish Clarias gariepinus. Aquaculture 2006, 256, 588–595. [Google Scholar] [CrossRef]
- Van de Nieuwegiessen, P.G.; Boerlage, A.S.; Verreth, J.A.; Schrama, J.W. Assessing the effects of a chronic stressor, stocking density, on welfare indicators of juvenile African catfish, Clarias gariepinus Burchell. Appl. Anim. Behav. Sci. 2008, 115, 233–243. [Google Scholar] [CrossRef]
- Baßmann, B.; Brenner, M.; Palm, H.W. Stress and Welfare of African Catfish (Clarias gariepinus Burchell, 1822) in a Coupled Aquaponic system. Water 2017, 9, 504. [Google Scholar] [CrossRef]
- Baßmann, B.; Harbach, H.; Weißbach, S.; Palm, H.W. Effect of plant density in coupled aquaponics on the welfare status of African catfish, Clarias gariepinus. J. World Aquac. Soc. 2020, 51, 183–199. [Google Scholar] [CrossRef]
- Eturki, S.; Ayari, F.; Jedidi, N.; Ben Dhia, H. Use of Clay Mineral to Reduce Ammonium from Wastewater. Effect of various parameters. Surf. Eng. Appl. Electrochem. 2012, 48, 276–283. [Google Scholar] [CrossRef]
- Seger, A.; Dorantes-Aranda, J.J.; Müller, M.N.; Body, A.; Peristyy, A.; Place, A.R.; Park, T.G.; Hallegraeff, G. Mitigating fish-killing Prymnesium parvum algal blooms in aquaculture ponds with clay: The importance of pH and clay type. J. Mar. Sci. Eng. 2015, 3, 154–174. [Google Scholar] [CrossRef]
- Edzwald, J.K.; Toensing, D.C.; Leung, M.C.Y. Phosphate Adsorption Reactions with Clay Minerals. Environ. Sci. Technol. 1976, 10, 485–490. [Google Scholar] [CrossRef]
- Heimann, R.B. Classic and Advanced Ceramics: From Fundamentals to Applications; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2010; p. 22. [Google Scholar]
- Attramadal, K.J.K.; Tøndel, B.; Salvesen, I.; Øie, G.; Vadstein, O.; Olsen, Y. Ceramic clay reduces the load of organic matter and bacteria in marine fish larval culture tanks. Aquac. Eng. 2012, 49, 23–34. [Google Scholar] [CrossRef]
- Seger, A.; Hallegraeff, G. Application of clay minerals to remove extracellular ichthyotoxins produced by the dinoflagellates Karlodinium veneficum and Karenia mikimotoi. Harmful Algae 2022, 111, 102151. [Google Scholar] [CrossRef]
- Desheng, Q.; Fan, L.; Yanhu, Y.; Niya, Z. Adsorption of aflatoxin B1 on montmorillonite. Poultry Sci. 2005, 84, 959–961. [Google Scholar] [CrossRef]
- Pasha, T.N.; Mahmood, A.; Khattak, F.M.; Jabbar, M.A.; Khan, A.D. The Effect of Feed Supplemented with Different Sodium Bentonite Treatments on Broiler Performance. Turk. J. Vet. Anim. Sci. 2008, 32, 245–248. [Google Scholar]
- Hassan, A.M.; Kenawy, A.M.; Abbas, W.T.; Abdel-Wahhab, M.A. Prevention of cytogenetic, histochemical and biochemical alterations in Oreochromis niloticus by dietary supplement of sorbent materials. Ecotox. Environ. Saf. 2010, 73, 1890–1895. [Google Scholar] [CrossRef]
- De Mil, T.; Devreese, M.; De Baere, S.; Van Ranst, E.; Eeckhout, M.; De Backer, P.; Croubels, S. Characterization of 27 mycotoxin binders and the relation with in vitro zearalenone adsorption at a single concentration. Toxins 2015, 7, 21–33. [Google Scholar] [CrossRef]
- Khoury, G.A.; Gehris, T.C.; Tribe, L.; Sánchez, R.M.T.; dos Santos Afonso, M. Glyphosate adsorption on montmorillonite: An experimental and theoretical study of surface complexes. Appl. Clay Sci. 2010, 50, 167–175. [Google Scholar] [CrossRef]
- Gill, J.P.K.; Sethi, N.; Mohan, A.; Datta, S.; Girdhar, M. Glyphosate toxicity for animals. Environ. Chem. Lett. 2018, 16, 401–426. [Google Scholar] [CrossRef]
- Marijani, E.; Kigadye, E.; Okoth, S. Occurrence of fungi and mycotoxins in fish feeds and their impact on fish health. Int. J. Microbiol. 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, M.; Vasconcelos, V. Occurrence of mycotoxins in fish feed and its effects: A review. Toxins 2020, 12, 160. [Google Scholar] [CrossRef] [PubMed]
- Koletsi, P.; Schrama, J.W.; Graat, E.A.; Wiegertjes, G.F.; Lyons, P.; Pietsch, C. The Occurrence of Mycotoxins in Raw Materials and Fish Feeds in Europe and the Potential Effects of Deoxynivalenol (DON) on the Health and Growth of Farmed Fish Species. Toxins 2021, 13, 403. [Google Scholar] [CrossRef]
- Deng, S.X.; Tian, L.X.; Liu, F.J.; Jin, S.J.; Liang, G.Y.; Yang, H.J.; Du, Z.Y.; Liu, Y.J. Toxic effects and residue of aflatoxin B1 in tilapia (Oreochromis niloticus × O. aureus) during long-term dietary exposure. Aquaculture 2010, 307, 233–240. [Google Scholar] [CrossRef]
- Anater, A.; Manyes, L.; Meca, G.; Ferrer, E.; Luciano, F.B.; Pimpao, C.T.; Font, G. Mycotoxins and their consequences in aquaculture: A review. Aquaculture 2016, 451, 1–10. [Google Scholar] [CrossRef]
- Palm, H.W.; Sörensen, H.; Knaus, U. Montmorillonite Clay Minerals with or without Microalgae as a Feed Additive in Larval White Leg Shrimp (Litopenaeus vannamei). Ann. Aquac. Res. 2015, 2, 1008. [Google Scholar]
- Palm, H.W.; Sörensen, H.; Knaus, U.; Dallwig, R. Use of Clay Minerals as a Food Additive and/or as Supplementary Food for Shrimps. European Patent EP 3 096 634 B1, 28 July 2021. [Google Scholar]
- Hu, C.H.; Xu, Y.; Xia, M.S.; Xiong, L.; Xu, Z.R. Effects of Cu2+-exchanged montmorillonite on growth performance, microbial ecology and intestinal morphology of Nile tilapia (Oreochromis niloticus). Aquaculture 2007, 270, 200–206. [Google Scholar] [CrossRef]
- Hu, C.H.; Xu, Y.; Xia, M.S.; Xiong, L.; Xu, Z.R. Effects of Cu2+-exchanged montmorillonite on intestinal microflora, digestibility and digestive enzyme activities of Nile tilapia. Aquac. Nutr. 2008, 14, 281–288. [Google Scholar] [CrossRef]
- Eya, J.C.; Parsons, A.; Haile, I.; Jagidi, P. Effects of Dietary Zeolites (Bentonite and Mordenite) on the Performance Juvenile Rainbow trout Onchorhynchus myskis [sic]. Aust. J. Basic Appl. Sci. 2008, 2, 961–967. [Google Scholar]
- Jawahar, S.; Nafar, A.; Paray, B.A.; Al-Sadoon, M.K.; Balasundaram, C.; Harikrishnan, R. Bentonite clay supplemented diet on immunity in stinging catfish, Heteropneustes fossilis against Aeromonas hydrophila. Fish Shellfish Immunol. 2018, 75, 27–31. [Google Scholar] [CrossRef]
- EFSA Panel on Additives and Products or Substances used in Animal Feed (FEEDAP). Scientific Opinion on the safety and efficacy of Friedland clay (montmorillonite-illite mixed layer clay) when used as technological additive for all animal species. EFSA J. 2014, 12, 3904. [Google Scholar] [CrossRef]
- European Commission. EU-Regulation (EU) 2016/1964, Official. J. Eur. U. 2016. Available online: https://eur-lex.europa.eu/legal-content/DE/TXT/PDF/?uri=CELEX:32016R1964&from=DE (accessed on 15 August 2019).
- Palm, H.W.; Knaus, U.; Wasenitz, B.; Bischoff, A.A.; Strauch, S.M. Proportional up scaling of African catfish (Clarias gariepinus Burchell, 1822) commercial recirculating aquaculture systems disproportionally affects nutrient dynamics. Aquaculture 2018, 491, 155–168. [Google Scholar] [CrossRef]
- Henning, K.H.; Kasbohm, J. Mineralbestand und Genese feinkörniger quartärer und präquartärer Sedimente in Nordostdeutschland unter besonderer Berücksichtigung des Friedländer Tones. In Berichte der Deutschen Ton- und Tonmineralgruppe (DTTG); Henning, K.H., Kasbohm, J., Eds.; 6, Greifswald, Germany; 1998; Available online: https://www.dttg.ethz.ch/cd_dttg1998/henning/henning.htm (accessed on 6 August 2019).
- FIM Biotech. Product Data Sheet 1g557-Montmorillonit-Illit; FIM Biotech GmbH: Berlin, Germany, 2017. [Google Scholar]
- Wendelaar Bonga, S.E. The stress response in fish. Physiol. Rev. 1997, 77, 591–625. [Google Scholar] [CrossRef]
- Deutscher Bundestag, Wissenschaftliche Dienste, WD 8-3000-087/20, WD 5-3000-131/20, WD 9-3000-105/20, Sachstand zu Tierversuchen und Tierversuchsfreien Alternativmethoden. 2020. Available online: https://www.bundestag.de/resource/blob/817022/7aacc71cdf8c7a2a69aabe20451699cd/WD-8-087-20_WD-5-131-20WD-9-105-20-pdf-data.pdf (accessed on 1 February 2022).
- Bhujel, R.C. Statistics for Aquaculture; Wiley-Blackwell: Ames, IA, USA, 2008. [Google Scholar]
- Masser, M.P.; Rakocy, J.; Losordo, T.M. Recirculating aquaculture tank production systems. Management of recirculating systems. SRAC Publ. 1999, 452. [Google Scholar]
- Ndubuisi, C.U.; Chimezie, J.A.; Chinedu, C.U.; Chikwem, C.I.; Alexander, U. Effect of pH on the growth performance and survival rate of Clarias gariepinus fry. Int. J. Res. Biosci. 2015, 4, 14–20. [Google Scholar]
- Lorsordo, T.M.; Masser, M.P.; Rakocy, J. Recirculating aquaculture tank production systems: An overview of critical considerations. SRAC Publ. 1998, 451. [Google Scholar]
- Dai, W.; Wang, X.; Guo, Y.; Wang, Q.; Ma, J. Growth performance, hematological and biochemical responses of African catfish (Clarias gariepinus) reared at different stocking densities. Afr. J. Agric. Res. 2011, 6, 6177–6182. [Google Scholar] [CrossRef]
- Manuel, R.; Boerrigter, J.; Roques, J.; van der Heul, J.; van den Bos, R.; Flik, G.; van de Vis, H. Stress in African catfish (Clarias gariepinus) following overland transportation. Fish Physiol. Biochem. 2014, 40, 33–44. [Google Scholar] [CrossRef]
- Ismaila, M.; Mulima, I.; Buba, U. Montmorillonite Clay as Mineral Supplement in the Diet of Catfish (Clarias gariepinus). J. Environ. Issues Agric. Dev. Ctries. 2011, 3, 98. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Water Parameters | E1 | E2 | ||||
|---|---|---|---|---|---|---|
| SI | SE | TF | SI | SE | TF | |
| T [°C] | 27.0 ± 0.2 | 27.1 ± 0.2 | 27.3 ± 0.2 | 28.7 ± 1.0 | 28.8 ± 1.0 | 28.8 ± 1.0 |
| O2 [mg L−1] | 6.5 ± 0.5 | 6.0 ± 0.7 | 7.5 ± 0.2 | 6.1 ± 0.7 | 5.0 ± 1.0 | 6.8 ± 0.5 |
| O2 [%] | 81.7 ± 5.9 | 75.3 ± 9.4 | 94.2 ± 2.7 | 78.8 ± 8.9 | 64.6 ± 11.9 | 87.7 ± 5.3 |
| pH | 6.6 ± 1.1 | 6.7 ± 1.1 | 6.8 ± 1.4 | 7.0 ± 0.7 | 7.0 ± 0.7 | 7.1 ± 0.8 |
| EC [µS cm−1] | 1254.5 ± 260.3 | 1256.5 ± 259.5 | 1263.1 ± 263.1 | 881.2 ± 80.5 | 881.4 ± 80.8 | 881.9 ± 81.0 |
| RedOx [mV] | 153.8 ± 42.6 | 157.8 ± 42.0 | 163.0 ± 47.4 | 160.4 ± 36.7 | 156.4 ± 34.0 | 155.8 ± 32.6 |
| NH4-N * [mg L−1] | 0.6 ± 0.9 | 0.6 ± 1.0 | 0.5 ± 0.9 | 0.7 ± 2.0 | 0.7 ± 2.0 | 0.7 ± 1.9 |
| NO2-N * [mg L−1] | 0.1 ± 0.1 | 0.1 ± 0.1 | 0.1 ± 0.1 | 0.3 ± 0.2 | 0.3 ± 0.3 | 0.2 ± 0.1 |
| NO3-N * [mg L−1] | 71.1 ± 36.4 | 71.8 ± 36.1 | 72.5 ± 36.1 | 196.4 ± 50.3 | 199.9 ± 52.9 | 195.5 ± 51.5 |
| PO4-P * [mg L−1] | 3.8 ± 1.8 | 3.9 ± 1.9 | 3.8 ± 1.9 | 17.2 ± 9.8 | 16.6 ± 9.8 | 16.6 ± 9.8 |
| Ingredient | Percentage |
|---|---|
| Wheat | 27.96 |
| Fish meal 70 M | 17.77 |
| Poultry meal | 11.22 |
| High protein (HP)-soya extract grist | 9.35 |
| Hemoglobin powder | 5.61 |
| Hydrolyzed feather meal | 5.61 |
| Pea protein | 5.61 |
| Monocalcium phosphate | 1.03 |
| Additional vitamins | [IU kg−1] |
| A; C; D; E | 12,000; 160; 1600; 160 |
| Nutrient | Coppens Special Pro EF 3–4.5 | Experimental Diets (from Beskow Feed Mill) | ||
|---|---|---|---|---|
| Adaptation Period | Control Group | 0.5% Group | 2.0% Group | |
| Crude protein [%] | 42.00 | 45.20 | 44.97 | 44.30 |
| Crude fat [%] | 13.00 | 15.00 | 15.00 | 15.00 |
| Carbohydrates [%] | Not specified | 19.60 | 19.50 | 19.21 |
| Crude ash [%] | 7.80 | 5.10 | 5.08 | 5.00 |
| Crude fiber [%] | 1.50 | 1.40 | 1.39 | 1.37 |
| Phosphorus [%] | 1.14 | 1.00 | 1.00 | 0.98 |
| Digestible energy [MJ kg−1] | 17.1 | 20.10 | 20.00 | 19.70 |
| 1g557 [%] | 0 | 0 | 0.5 | 2.0 |
| Sampling Time | n | Group | Weight [g] | Length [cm] | Δ [%] | CI [g/cm3] | |||
|---|---|---|---|---|---|---|---|---|---|
| E1 | |||||||||
| Before stocking and adaptation phase | 309 | C | 30.7 a,b | ±5.8 | 16.7 a,b | ±1.1 | 0 | 0.652 a | ±0.1 |
| 308 | 0.5% | 31.6 a | ±6.0 | 16.9 a | ±1.1 | +2.9 | 0.652 a | ±0.0 | |
| 309 | 2.0% | 30.3 b | ±6.1 | 16.6 b | ±1.1 | −1.3 | 0.661 b | ±0.0 | |
| T0 (start of experiment) | 33 | C | 107.8 a | ±17.2 | 24.8 a | ±1.5 | n.g. | 0.7 a | ±0.1 |
| 33 | 0.5% | 112.6 a | ±16.2 | 25.2 a | ±1.3 | n.g. | 0.7 a | ±0.0 | |
| 33 | 2.0% | 107.9 a | ±17.1 | 24.5 a | ±1.4 | n.g. | 0.7 a | ±0.0 | |
| T3 (after 70 d) | 166 | C | 480.5 a | ±89.2 | 39.2 a | ±3.0 | 0 | 0.8 a | ±0.1 |
| 164 | 0.5% | 484.2 a | ±86.7 | 39.4 a | ±2.8 | +0.8 | 0.8 a | ±0.1 | |
| 171 | 2.0% | 469.5 a | ±93.3 | 39.0 a | ±2.9 | −2.3 | 0.8 a | ±0.1 | |
| E2 | |||||||||
| Before stocking and adaptation phase | 309 | C | 29.8 a | ±3.6 | 17.0 a | ±0.8 | 0 | 0.6 a | ±0.1 |
| 309 | 0.5% | 29.7 a | ±3.3 | 17.0 a | ±0.8 | −0.3 | 0.6 a | ±0.0 | |
| T0 (start of experiment) | 33 | C | 115.6 a | ±20.8 | 25.3 a | ±1.6 | n.g. | 0.7 a | ±0.0 |
| 33 | 0.5% | 115.2 a | ±16.4 | 25.3 a | ±1.4 | n.g. | 0.7 a | ±0.1 | |
| T3 (after 70 d) | 172 | C | 409.2 a | ±73.1 | 37.5 a | ±2.4 | 0 | 0.8 a | ±0.2 |
| 163 | 0.5% | 422.2 a | ±76.1 | 37.9 a | ±2.5 | +3.2 | 0.8 a | ±0.2 | |
| Group | Stocking–T0 | T0–T1 | T1–T2 | T2–T3 | |
|---|---|---|---|---|---|
| E1 | C | 0.66 ± 0.01 a | 0.78 ± 0.02 a | 0.86 ± 0.03 a | 0.96 ± 0.08 a |
| 0.5% | 0.66 ± 0.00 a | 0.76 ± 0.02 a | 0.89 ± 0.02 a | 0.97 ± 0.05 a | |
| 2.0% | 0.65 ± 0.00 a | 0.77 ± 0.02 a | 0.92 ± 0.03 a | 0.94 ± 0.10 a | |
| E2 | C | 0.76 ± 0.01 a | 0.95 ± 0.04 a | 1.06 ± 0.04 a | 2.09 ± 0.52 a |
| 0.5% | 0.76 ± 0.01 a | 0.94 ± 0.02 a | 1.04 ± 0.03 a | 1.45 ± 0.19 a |
| Mortality | Groups at T3 | ||
|---|---|---|---|
| Control (E1/E2) | 0.5% (E1/E2) | 2.0% (E1) | |
| mean ± Sd total n | 3.67 ± 1.53 a/2.67 ± 2.52 a 11/8 | 5.00 ± 0.00 a/5.67 ± 2.89 a 15/17 | 3.00 ± 1.00 a 9 |
| % | 3.6/2.6 | 4.9/5.5 | 2.9 |
| Sampling Time | n | Group | AST (GOT) [U/L] | GLDH [U/I] | Urea [mmol/L] | Calcium [mmol/L] | Phosphate [mmol/L] | Total Protein [g/L] | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| E1 | ||||||||||||||
| T0 (start of experiment) | 9 | Base line | 333.7 | ±79.7 | 29.4 | ±3.7 | 1.1 | ±0.1 | 2.8 | ±0.1 | 3.9 | ±0.2 | 29.9 | ±1.4 |
| T3 (after 70 d) | 3 | C | 208.7 | ±38.2 | 28.2 | ±3.7 | 1.1 | ±0.1 | 3.3 | ±0.2 | 3.7 | ±0.6 | 39.3 | ±1.2 |
| 3 | 0.5% | 167.3 | ±12.2 | 28.1 | ±4.7 | 1.0 | ±0.1 | 3.3 | ±0.3 | 3.3 | ±0.1 | 38.0 | ±4.1 | |
| 3 | 2.0% | 179.0 | ±5.0 | 22.7 | ±2.1 | 1.0 | ±0.1 | 3.2 | ±0.4 | 3.2 | ±0.4 | 37.0 | ±2.2 | |
| E2 | ||||||||||||||
| T0 (start of experiment) | 12 | Base line | 150.8 | ±28.7 | 14.7 | ±2.5 | 1.3 | ±0.2 | 3.0 | ±0.1 | 3.0 | ±0.2 | 33.7 | ±1.2 |
| T3 (after 70 d) | 6 | C | 109.0 | ±18.8 | 18.4 | ±2.2 | 1.1 | ±0.1 | 3.4 | ±0.1 | 2.5 | ±0.2 | 39.5 | ±2.4 |
| 6 | 0.5% | 142.7 | ±48.8 | 26.5 | ±9.2 | 1.3 | ±0.1 | 3.5 | ±0.2 | 2.8 | ±0.2 | 41.1 | ±2.2 | |
| Sampling Time | Group | Leucocytes [G/L] | Erythrocytes [T/L] | Hematocrit [%] | Cholesterol [mmol/L] | Triglycerides [mmol/L] | Sodium [mmol/L] | Potassium [mmol/L] | Chloride [mmol/L] |
|---|---|---|---|---|---|---|---|---|---|
| E1 | |||||||||
| T0 | Base line | 3.3 ± 2.7 | 1.1 ± 0.6 | 21.5 ± 7.3 | 2.9 ± 0.2 | 1.8 ± 0.1 | 124.6 ± 2.6 | 13.0 ± 1.7 | 112.0 ± 2.0 |
| T3 | C | 0.9 ± 0.3 | 2.0 ± 0.5 | 26.2 ± 1.3 | 3.5 ± 0.1 | 2.0 ± 0.1 | 128.3 ± 1.2 | 9.3 ± 1.6 | 107.0 ± 1.4 |
| 0.5% | 1.0 ± 0.2 | 1.7 ± 0.3 | 25.8 ± 0.2 | 3.4 ± 0.3 | 2.0 ± 0.4 | 129.7 ± 0.9 | 8.4 ± 0.5 | 107.7 ± 0.5 | |
| 2.0% | 1.0 ± 0.0 | 1.3 ± 0.4 | 24.5 ± 0.7 | 3.1 ± 0.3 | 1.7 ± 0.2 | 128.7 ± 1.2 | 9.5 ± 2.3 | 110.7 ± 2.1 | |
| E2 | |||||||||
| T0 | Base line | 3.7 ± 0.8 | 1.9 ± 0.4 | 26.7 ± 2.0 | 2.8 ± 0.3 | 1.8 ± 0.1 | 130.0 ± 1.9 | 7.6 ± 1.3 | 111.5 ± 1.5 |
| T3 | C | 13.0 ± 7.5 | 2.1 ± 0.2 | 36.3 ± 3.0 | 3.4 ± 0.2 | 2.0 ± 0.1 | 133.5 ± 1.7 | 7.3 ± 0.6 | 110.5 ± 1.6 |
| 0.5% | 7.0 ± 3.3 | 1.7 ± 0.6 | 34.7 ± 7.1 | 3.4 ± 0.4 | 2.0 ± 0.1 | 133.7 ± 2.0 | 7.4 ± 0.3 | 111.0 ± 1.6 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palm, H.W.; Berchtold, E.; Gille, B.; Knaus, U.; Wenzel, L.C.; Baßmann, B. Growth and Welfare of African Catfish (Clarias gariepinus Burchell, 1822) under Dietary Supplementation with Mixed-Layer Clay Mineral Montmorillonite-Illite/Muscovite in Commercial Aquaculture. Aquac. J. 2022, 2, 227-245. https://doi.org/10.3390/aquacj2030013
Palm HW, Berchtold E, Gille B, Knaus U, Wenzel LC, Baßmann B. Growth and Welfare of African Catfish (Clarias gariepinus Burchell, 1822) under Dietary Supplementation with Mixed-Layer Clay Mineral Montmorillonite-Illite/Muscovite in Commercial Aquaculture. Aquaculture Journal. 2022; 2(3):227-245. https://doi.org/10.3390/aquacj2030013
Chicago/Turabian StylePalm, Harry Wilhelm, Erwin Berchtold, Benedikt Gille, Ulrich Knaus, Lisa Carolina Wenzel, and Björn Baßmann. 2022. "Growth and Welfare of African Catfish (Clarias gariepinus Burchell, 1822) under Dietary Supplementation with Mixed-Layer Clay Mineral Montmorillonite-Illite/Muscovite in Commercial Aquaculture" Aquaculture Journal 2, no. 3: 227-245. https://doi.org/10.3390/aquacj2030013
APA StylePalm, H. W., Berchtold, E., Gille, B., Knaus, U., Wenzel, L. C., & Baßmann, B. (2022). Growth and Welfare of African Catfish (Clarias gariepinus Burchell, 1822) under Dietary Supplementation with Mixed-Layer Clay Mineral Montmorillonite-Illite/Muscovite in Commercial Aquaculture. Aquaculture Journal, 2(3), 227-245. https://doi.org/10.3390/aquacj2030013