Abstract
This study investigated the adaptability of Dunaliella salina to different salinity levels, with an emphasis on growth, pigment concentration, and desalination potential. It was found that among the 21 salinity levels, Salinity 75 produced consistently favorable results in cell count (13.08 × 103 ± 1.41 × 103 cells/mL), dry biomass (2.46 ± 0.06 g/L), pigment content (chlorophyll a = 97,500,000 ± 100,000 pg/L, chlorophyll b = 123,600,000 ± 300,000 pg/L), and desalination (9.32 ± 0.47 reduction). Therefore, Salinity 75 was selected for the final trial (scale-up), which revealed unanticipatedly high cell counts (58.96 × 103 ± 535.22 cells/mL), with the dry biomass weight being statistically different (higher) than expected (4.21 ± 0.02 g/L) (p < 0.0001), most likely due to the high cell count and energy reserve storage for high-salinity adaption in the form of bio-compounds. Pigment growth continued (chlorophyll a = 95,400,000 ± 2,200,000 pg/L, chlorophyll b = 128,100,000 ± 5,100,000 pg/L), indicating pigment production under salt stress. Notably, desalination did not occur in this stage, possibly due to the necessity for a bigger initial inoculate, prolonged exposure or bioaccumulation becoming the prevailing mechanism over desalination. Nevertheless, the trial highlights D. salina’s strong adaptation to various salinity levels. This suggests a promising future in halophyte research, particularly in understanding the mechanisms that prevent salt accumulation in cells and how to overcome this barrier. Additionally, these results suggest that microalgae could be a viable resource in saline-rich environments unsuitable for conventional agriculture, promoting industrial adaptation to adverse conditions.
1. Introduction
Water plays a pivotal role in driving economic and social development [1]. In current times, the world faces a range of water-related challenges, including limited access to freshwater, declining water quality, reduced financial resources, and fragmented water management efforts [2]. Water scarcity not only hinders economic development but also poses significant threats to human health, environmental degradation, and political stability [3,4,5]. Nations from arid environments are already experiencing a water crisis and it is expected to worsen with population rates and the expansion of the industrial and agricultural sectors, resulting in a higher demand, raising the cost of water and intensifying competition for the resource across various sectors [6]. These nations may see desalinating saline water from rivers and seas as an unavoidable solution for meeting water supply needs [7,8,9,10].
To address the growing demand for freshwater, it has become increasingly important to explore alternative solutions. One such solution is saltwater desalination, which has emerged as a crucial method to ensure a sustainable future for generations worldwide. In recent years, there has been considerable attention focused on the potential and possibilities of desalination technology in addressing the global water scarcity crisis [10]. In its simplest form, this technology has the potential to significantly mitigate water scarcity by tapping into limitless reserves of seawater and abundant reserves of brackish groundwater as new sources of freshwater supply.
Using microalgae to treat water for diverse applications is a cost-efficient approach [11,12]. This proposed solution holds the potential to address the challenge of minimal-expense desalination [13,14], yet microalgae desalination attempts have been reported for well over a decade now [15]. Algae-based desalination systems with algal biomass reuse have low environmental impacts due to lower energy consumption, lower operational and maintenance requirements, and the potential for electricity generation [11]. These processes are in fact promising, but challenges such as biomass harvest, treatment efficiency, and large-scale industry implementation need to be addressed [16].
These microorganisms are incredibly diverse, ranging from tiny single-celled organisms to larger, more complex species that can thrive in a wide range of environments, from different aquatic environments to arid environments [17]. Most microalgae species exhibit a capacity to thrive in saline waters that are saturated with water-soluble substances [18]. Through this process, these living organisms absorb water salts along with other nutrients into their biomass, ultimately decreasing the salt content in the water due to this absorption [19,20]. Some organisms tend to absorb more water-soluble substances than they require for their nutritional needs, making them effective in reducing water salinity [21].
The desalination or reduction in water salinity through biological means involves the utilization of various macrophytes (macroalgae), microphytes (microalgae), other microorganisms, or their combinations, which has been reported to lower water salinity [22]. Unlike typical terrestrial plants, algae can complete their life cycle across a wide range of salinity levels. Microalgae, for instance, have been employed for many years in tertiary wastewater treatment to remove nitrogen and phosphorus compounds following a reduction in BOD (Biochemical Oxygen Demand) and COD (Chemical Oxygen Demand) levels [23]. Numerous microalgae, including species like Scenedesmus obliquus, are mixotrophic, meaning they can utilize both organic matter and minerals during their growth process. This can be achieved through photosynthesis (like pure autotrophic algae, converting CO2 and H2O in organic compounds) and through inorganic nutrients’ uptake (such as nitrates and ammonium, phosphates, silicates, and other trace elements) [24]. This characteristic enables them to serve as an alternative secondary treatment method, effectively reducing organic matter and nutrient content [25].
Overall, bibliography indicates that the use of algae can be advantageous for mitigating water salinity. It is worth noting that fluctuations in physical and chemical factors, such as salinity, have substantial impacts on the growth and biochemical composition of green microalgae [17,18,20].
Dunaliella salina, is a species of microalgae that belongs to the Phylum Chlorophyta, Class Chlorophyceae, Order Chlamydomonadales [26]. It has shown remarkable resilience to high salinity (surviving in salinities up to 27 [27]) and is commonly found in saline environments such as lakes and saltwater lagoons.
Ehrenfeld and Cousin observed that during an initial hypertonic shock, D. salina cells exhibit a sudden increase in Na+ content due to an influx of Na+ through the cell membranes [28]. Despite this ability to grow in environments with a wide range of salt concentrations, Na+ levels in D. salina should eventually lower and stabilize. The antiporter in D. salina plays a crucial role in regulating the intracellular Na+ concentration [29]. The antiporter responsible for salt regulation is a specialized membrane protein that manages the organism’s internal ion concentrations in response to external salt levels. When D. salina is exposed to high salt concentrations, the antiporter facilitates the exchange of Na+ and possibly other ions across the cell membrane. Specifically, it expels Na+ while absorbing other ions, such as H+, to maintain osmotic balance and prevent sodium buildup in the cell [30,31]. Reports of an increase in intracellular Na+, however, indicate that D. salina cells can accumulate Na+, which likely contributes to osmoregulation in the early stages of exposure to high salt concentrations. This process involves compartmentalizing the accumulated salt within the cell before stabilizing, reflecting the cells’ adjustment to maintain osmoregulation during the transition from high to hyper-saline conditions [32]. The findings suggest that the effectiveness of this algae in high salinity conditions can be harnessed to reduce the salinity of various water sources, including seawater, sewage, and industrial wastewater recycling.
Still, there are not many studies that show how the microalgae reacts to different salinities, from none to extremely salty. In this study, the proposed range of salt tolerance was from 9 to 165.
This study aims to fill in the gap in the literature regarding the D. salina response to high salinity in terms of culture growth, pigment concentration, and desalination, exploring the possibility of this microalgae being fit to use in the desalination of salty sludges or waters, and if this culturing environment could implement a biorefinery approach in terms of the productivity changes implemented by the salty conditions. This study will provide an insight to the mechanism of salt regulation in halophyte microalgae and provide a solid base for future studies on the subject.
2. Materials and Methods
2.1. Sample Preparation for the Initial Trial
For the initial trial, triplicate 250 mL conical flasks with 150 mL of ASN-III media [33] (3.5 g MgSO4 × 7H2O, 2 g MgCl2 × 6H2O, 0.5 g CaCl2 × 2H2O, 0.5 g KCl, 3 mg citric acid, 3 mg fe-Amm-Citrate, 0.5 mg EDTA, 1 mL A5 trace metals mix, 0.75 g NaNO3, 0.75 g K2HPO4 × 3H2O, 0.02 g Na2CO3, 1000 mL deionized water, and NaCl) were prepared with different salinities by adding NaCl and then used as bioreactors (Table 1). Each bioreactor was inoculated with 20 mL of D. salina (CCAP 19/20, from the culture collection of algae and protozoa) solution (25 × 104 cells per mL) previously cultivated in its normal salinity range. All bioreactors were placed in a seesaw rocker with a constant rocking motion, light intensity of 2300 Lux, and light/dark cycle of 14:10.

Table 1.
Bioreactors with ASN-III media prepared under different salinities, in triplicate.
2.1.1. Salinity Assessment
A refractometer (Autoutlet, Beijing, China) with a precision of ±1 in the practical salinity scale was used to measure salinity [34]. No-salt ASN-III media was used to calibrate the refractometer. A sample volume of 1.5 mL was collected and subsequently centrifuged at 3500 rpm for 5 min to ensure the microalgae cells would not interfere with the salinity measurement. The supernatant was collected, and salinity was assessed in the refractometer. The practical salinity scale is defined as a conductivity ratio without units. A seawater sample with a conductivity ratio of 1.0 at 15 °C, when compared to a potassium chloride (KCl) solution containing 32.4356 g of KCl per 1 kg of solution, corresponds to a salinity of 35.000 [35,36].
2.1.2. Cell Counting
Cell counting was performed by flow cytometry (MACSQuant, Teterow, Germany) as described by Skufca et al. (2022) and Dominique et al. (2005) [37,38]. The gating strategy was based on chlorophyll autofluorescence in live cells and emission detected in channels B3 (488 nm/655–730 nm) and R1 (635/655–730 nm).
2.1.3. Dry Weight Assessment
To measure the dry weight of microalgae per liter, 20 mL of the culture was placed in a 50 mL vial tube, pre-weighed empty. The solution was centrifuged at 3500 rpm for 10 min and the supernatant discarded. Following this, deionized water was added to the tube and the pellet was homogenized by vortexing, the tube was then once again centrifuged and the supernatant discarded. This cleaning step was repeated 3 times to remove the NaCL from interfering with the final weight result. The tube with the formed pellet was placed to dry overnight at 60 °C and the next day would be weighed. The resulting difference observed showed us the dry weight of cells per 20 mL of solution. By multiplying this number by 50, we would obtain the weight of dry biomass per liter of solution (g/L) [39]. We also ran a blank control with 20 mL of dH2O.
2.1.4. Chlorophyll Assessment
To quantify chlorophyll a and b, procedures described by Caspers, 1970, were used [40]. A 2 mL sample of the cell solution was collected and placed into a 15 mL tube already containing 5 mL of acetone/water (9:1). This mixture was vortexed and then centrifuged at 3000× g for 3 min. The resulting supernatant was analyzed using a spectrophotometer at wavelengths of 630 nm, 647 nm, and 664 nm. The following equations were employed to convert the absorbance values into concentration measurements (pg/L) and the dilution factor was taken into account [41,42]:
Chlorophyll a = [(11.85 × DO664) + (−1.54 × DO647) + (−0.08 × DO630)] × 100,000
Chlorophyll b = [(−5.47 × DO664) + (21.03 × DO647) + (−2.66 × DO630)] × 100,000
2.2. Upscaling
For the final trial, after checking the preliminary results, a chosen salinity was upscaled to 5 L bioreactors in triplicate. The bioreactors were prepared with constant magnetic stirring at 120 rpm, air flow into an airstone (1.8 L/min), and light intensity of 2300 Lux with a light/dark cycle of 14:10. Salinity, cell count, dry weight, and chlorophyll assessments were conducted all throughout the trial (initially every 30 min, then daily).
2.3. Statistical Analysis
Statistical analysis was performed using one-way analysis of variance (ANOVA) with a Tukey post hoc test to determine differences. The software used to perform statistical analysis was Graphpad PRISM v.8.0.2 for Windows. All data collected in this study were expressed as the mean ± standard deviation. Difference significances were attributed as follows: not significant (ns: p ≥ 0.05), significant (*: p = 0.01 to 0.05), very significant (**: p = 0.001 to 0.01), and extremely significant (***: p = 0.0001 to 0.001, and **** = p < 0.0001).
3. Results
3.1. Initial Trial
After 14 days, the cultures in the initial trial were analyzed, and the results for salt removal (a), cell count (b), dry biomass (c), chlorophyll a (d), and chlorophyll b (e) are presented in Figure 1.
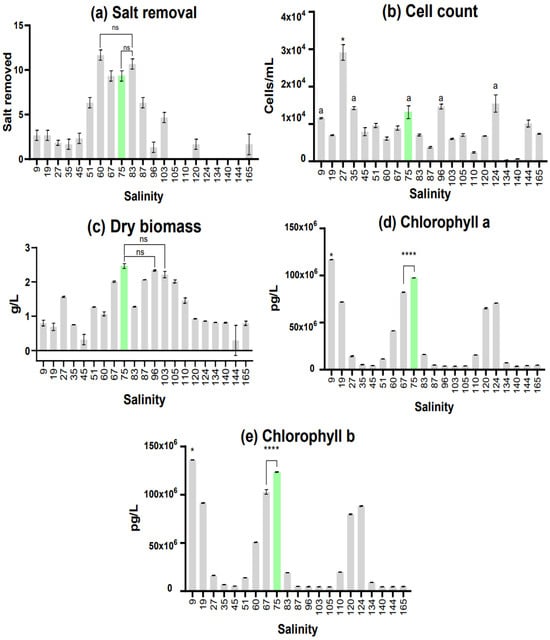
Figure 1.
All results obtained from the initial trial, n = 60, error bars represent standard deviation. Difference significances were attributed as follows: not significant (ns: p ≥ 0.05), significant (*: p = 0.01 to 0.05), and very significant (**** = p < 0.0001). (a) Result of salt percentage removed from media for each salinity after the initial trial culture; (b) Result of cell counting for each bioreactor after the initial trial, in cells per milliliter of culture, same letter shows no statistically significant difference; (c) Result of dry biomass assessment for each bioreactor after the initial trial, in grams of dry weight per liter of culture; (d,e) Result of pigment assessment for each bioreactor after the initial trial, in pg per liter of culture. Salinity 75 is highlighted in green as it was chosen for upscaling.
Desalination was measured in the percentage of salt removed. The best desalination was in Salinity 60 (11.67 ± 0.58), followed by 83 (10.67 ± 0.58) and 75/83 (9.33 ± 0.58). Desalination in the salinity levels of 105, 110, 124, 134, 140, and 144 did not occur. This was expected for such a high salt level, being that even the desalinations achieved in Salinities 120 (1.58 ± 0.47) and 165 (1.44 ± 0.94) were not significant.
When analyzing the cell count results, salinities close to 25 were expected to be among the highest due to it being the traditionally used salinity for D. salina media [33]. This was indeed observed for the Salinity 27 culture, obtaining a much higher cell count than the others, showing statistically significant differences from all other bioreactors in this regard. In addition to this, Salinities 9 (11.6 × 103 ± 140 cells/L), 35 (14.2 × 103 ± 320 cells/L), 75 (13 × 103 ± 1408 cells/L), 96 (14.6 × 103 ± 544 cells/L), 124 (15.3 × 103 ± 1882 cells/L), and 144 (10 × 103 ± 753 cells/L) all presented cell counts above 10 × 103 cells per milliliter. This proves these salinities can be used for culturing, since Salinities 9, 35, 75, 96, and 124 show no statistically significant differences.
As for dry biomass, the highest cell count culture (Salinity 27) obtained a value of 1.56 g of dry biomass per liter of culture. Salinities 67 (2.01 ± 0.016 g/L), 75 (2.46 ± 0.056 g/L), 87 (2.06 ± 0.003 g/L), 96 (2.33 ± 0.014 g/L), 103 (2.2 ± 0.077 g/L), and 105 (2.01 ± 0.037 g/L) all surpassed the 2 g/L mark. Salinity 75 expressed the maximum value of this trial with 2.4 g/L, presenting no significant differences from Salinities 96 and 103 (the second and third highest values) while being significantly statistically different from all other bioreactors.
Pigment assessment exhibited interesting results since there is a clear pattern with 3 peaks in pigment productivity; these are shown at Salinities 9 (chlorophyll a: 116,900,000 pg/L ± 140,000 pg/L, b: 136,200,000 pg/L ± 120,000 pg/L), 75 (chlorophyll a: 97,500,000 pg/L ± 120,000 pg/L, b: 123,600,000 pg/L ± 280,000 pg/L), and 124 (chlorophyll a: 70,700,000 pg/L ± 150,000 pg/L, b: 88,300,000 pg/L ± 380,000 pg/L). The highest peak observed was at Salinity 9, expressing significant differences from all other bioreactors. The second highest peak was shown in Salinity 75, also demonstrating statistically significant differences from all other bioreactors. When taking into consideration the amount of pigment per number of cells, the most productive salinities were 9 (10.08 pg/cell, of chlorophyll a; 11.75 pg/cell, of chlorophyll b), 19 (10.25 pg/cell, of chlorophyll a; 13.08 pg/cell, of chlorophyll b), 67 (9.27 pg/cell, of chlorophyll a; 11.59 pg/cell, of chlorophyll b), 75 (7.45 pg/cell, of chlorophyll a; 9.45 pg/cell, of chlorophyll b), 120 (9.64 pg/cell, of chlorophyll a; 11.79 pg/cell, of chlorophyll b), and 134 (24.33 pg/cell, of chlorophyll a; 29.95 pg/cell, of chlorophyll b).
3.2. Upscaling
Considering the results for these four parameters in the initial trial, we decided to use Salinity 75 as a model for upscaling. The results were divided in the first 5 h after inoculation (to check for quick responses to upscaling) and after the first day (with daily analysis whenever possible). The results obtained were expressed in graph form (Figure 2).
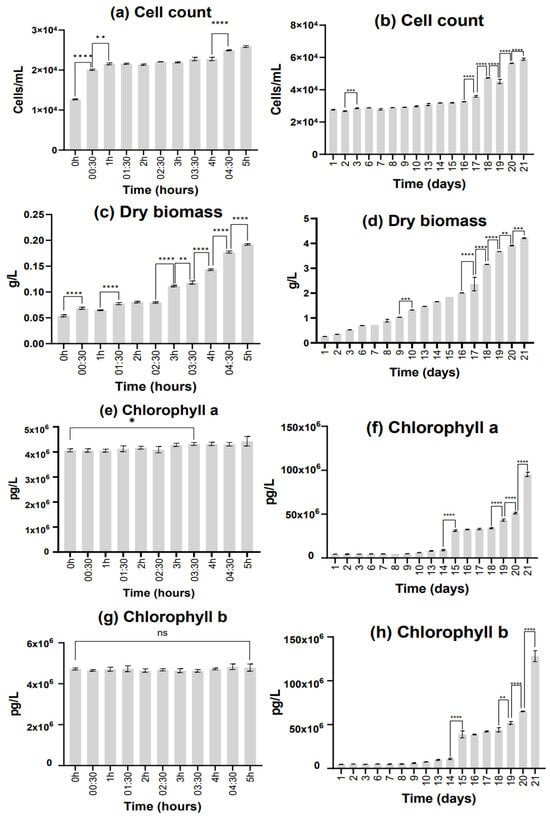
Figure 2.
Results obtained from the final trial, n = 3, bars represent standard deviation. Difference significances were attributed as follows: Not significant (ns: p ≥ 0.05), Significant (*: p = 0.01 to 0.05), Very significant (**: p = 0.001 to 0.01) and Extremely significant (***: p = 0.0001 to 0.001, and **** = p < 0.0001). The graphs are organized into two groups based on the time window of the data: The graphs on the left side show results for the first 5 h window. The graphs on the right side show results for daily samples up to Day 21. (a,b) Result of cell counting for each salinity, in cells per milliliter of culture; (c,d) Result of dry biomass assessment after 21 days of culture, in grams of dry weight per liter of culture; (e–h) Result of pigment assessment, in pg per liter of culture.
When analyzing the cell count results, there was noticeable growth. The cell count reached double the initial assessment after 5 h of the trial, and on the last day of the assessment (the 21st day), the cell count was 59.95 × 103 ± 535.22 cells/mL. When assessing continuous results for the first 5 h (from one 30 min sample to the next), statistically significant differences were observed between the first three samplings (from 0 h to 00:30, and 00:30 to 1 h), indicating that there was a good initial adaptation by D. salina to these new conditions. As for the remaining trial (21 days), although a significant difference can be observed from Day 2 to Day 3, it becomes apparent that the microalgae were going through an adaptation period until the 16th day. Every day after this one showed statistically significant differences from the next, demonstrating positive daily growth until the end of the trial (the 21st day).
As for dry biomass, the final value in the upscaling trial was higher than the initial trial. This may indicate that salt is increasing the cell weight, probably because of the necessity of storing energy reserves as a necessity for high-salinity adaptation in response to stress [43,44]. After 21 days, we obtained 4.2 g of dry biomass per liter of culture. In the first 5 h, there was significant growth with some adaptation periods (such as 00:30–1 h and 1:30–2:30). As for the remaining days, there was continuous growth with the first day-to-day samples presenting significant differences from Day 9 to Day 10. After this, the microalgae reached a point of continuous significant growth on Day 16 all the way to the end of the trial.
Pigment assessment exhibited that there was continuous pigment development throughout the whole study. For chlorophyll a, the first sample showing statistically significant growth from the sample at 0 h was the one collected at the 3 h and 30 min mark; this was 30 min after the significant ascending trend started in dry biomass. There were no significant differences in chlorophyll b content (from 0 h) for the first 5 h of the trial. As for the remaining 21 days, both pigments presented the same trends with a significant jump in production from Day 14 to Day 15, a three-day stabilization period, and statistically significant pigment production from Day 18 to the end of the trial, reaching a final maximum value of 95,400,000 ± 2,200,000 pg/L of chlorophyll a and 128,100,000 ± 5,100,000 pg/L of chlorophyll b. In the current trial, the maximum production in terms of pigment per cell was observed on the final day (Day 21–1.62 µg/cell, of chlorophyll a; 2.17 µg/cell, of chlorophyll b). This may indicate that the quantity of pigment in the cell was still increasing, and therefore the cell was still continuously producing pigment.
Desalination did not occur throughout the upscaling section of the study
4. Discussion
When analyzing desalination in the initial trial, the results were positive but still inferior to past studies which achieved a desalination of 20 on a 130 mS/cm solution in 2021, in a 5 L setup with a light intensity of 3500 Lux at 25 °C [45]. Desalination occurs due to an early influx of Na+ through the cell membranes. Yet this desalination action could not be replicated in the upscaling process. This might be attributed to high efficiency by the antiporter in the cell. As the initial ratio of the cell density to bioreactor volume in the upscale trial was inferior to the initial trial, the microalgae may have absorbed the salt soon after its inoculation without a substantial number of cells in the culture and therefore no measurable desalination. The antiporter mechanism was then triggered and created an osmotic equilibrium by expelling Na+ from the cell, facilitating the cell’s survival under high salinity. Following this trend, when the culture grew to a substantial cell density to potentially affect salinity, a majority of the cells had already reached osmotic equilibrium and therefore would not uptake salt.
Also, no desalination effect could be attributed to the adsorption of ions to the cells surface, previous to inoculation. The large surface and strong binding properties of the cell wall in D. salina increase this adsorption [46,47]. The continuous cell multiplication, dry biomass increase, and pigment production suggest that bioaccumulation became the prevailing mechanism [12,48,49] over desalination [50]. This indicates a problem to overcome with the microalgae desalination strategy, yet for future studies on a larger scale, controlling and monitoring culture conditions, with a focus on salt uptake by the cell, and different origins/quantities of inoculate could prove this desalination strategy as valuable. A bigger inoculate could trigger the salt uptake uniformly in all cells of the culture before the antiporter mechanism would be triggered, resulting in large-scale desalination.
The cell count was positive in both stages of the study, as there was an increase from the initial dilution of inoculate. For the upscaling, although there was a long period of adaptation, cells quickly multiplied on a daily basis from the 16th day (32.6 × 103 ± 0.095 × 103 cells/mL) to the 21st and final day of the trial (59.95 × 103 ± 0.535 × 103 cells/mL). These results are successful for cell culturing, and positive when compared to the past literature; for example, Sedjati et al. observed 1.231 ± 0.025 × 104 and 0.892 ± 0.005 × 104 cells/mL at Salinities 20 and 40, respectively [51]. These results prove highly interesting since a higher cell count could account for a higher productivity in every parameter, including desalination potential. It is worthy of note that the amount of inoculate used also affects the success of the culture and cell replication numbers [52], although this was not a variable in this study.
Dry biomass was also a highlight of this study, with various bioreactors in the initial trial surpassing the 2 g/L mark. The upscaling section of this study showed much higher results, with the last day (21st) presenting a dry biomass result of 4.21 ± 0.016 g/L. These results were highly positive when compared to the past literature, such as the study by Morowvat and Ghasemi where the maximum concentrations of D. salina in an optimized Johnson culture and basic culture medium were 0.997 and 0.571 g/L, respectively [53]. Previous studies have reported this boost in biomass production when the culture is subjected to high salinities [54]. The favorable biomass findings are particularly intriguing as microalgal biomass holds significant biotechnological promise for utilization in integrated multi-product biorefineries [55]. Salt concentration and user error when washing the cells of salt may have affected the results of dry biomass assessment.
The relation between cell count and dry biomass showed some variance. For the initial trial, Salinity 27 presented the highest cell count, yet the dry biomass for such salinity was significantly inferior to the highest group in this regard. This is the closest to the standard salinity for D. salina culture. The ratio of viable cells per total cells from this culture was therefore a lot higher then, for example, when compared to Salinity 103. In Salinity 103, the values for dry biomass had no statistically significant difference from the ones obtained by the highest dry biomass cultures (Salinity 75), yet the cell count is relatively low. Therefore, the number of viable cells per total cells in the culture is inferior. This is observed due to the presence of dead cells in the culture that clearly affect dry biomass assessments while not being viable [56].
As for the pigment assessment, an interesting pattern can be observed in Figure 1d,e, referring to the initial trial. There are three peaks that represent hotspots for pigment production. These are in Salinities 9, 75 and 124. These results could be interesting for the industry since we demonstrate three potential salinities to boost production. As for the upscaling section of this study, peak pigment production was reached on the final day of trial. This may indicate that the quantity of pigment in the cell was still increasing, and therefore the cell was still continuously producing pigment. For chlorophyll a, the first sample to show statistically significant growth from the sample at 0 h was the 3 h and 30 min sample; this was 30 min after the significant ascending trend started in dry biomass, implying that first the microalgae grew in mass, and then started developing its chlorophyll a content. The increase in cellular chlorophyll during photoacclimation is correlated with an increase in photosynthetic units, leading to improved efficiency and a reduced impact of low light on growth rate [57]. Colusse et al. in 2020 measured a chlorophyll content of 3,920,000 pg/L ± 430,000 pg/L using Conway medium [58]. The difference between medias can be explained by the fact that Conway medium is relatively simple compared to ASNIII medium, which contains a broader range of nutrients and more closely resembles natural seawater.
Although not considered in this study, another route could be using a carrier, such as an alginate bead encapsulating the microalgae, to introduce D. salina to the salty environment. Similar approaches have been successful for the removal of ammonia and phosphorus from aquaculture wastewater [59].
5. Conclusions
The findings from the upscaling phase indicate a lack of desalination activity; however, the trial reinforces the robust adaptation of D. salina to diverse salinity levels. This observation underscores an interesting area for future halophyte research, focusing on elucidating what influences mechanisms to reject salt accumulation within the cell and how to overcome this hurdle. Moreover, these results present the potential exploration of microalgae as a viable resource in saline-rich environments unsuitable for conventional agricultural practices, thereby facilitating industrial adaptation to adverse conditions. D. salina was utilized since it is one of the most investigated halophytes in terms of desalination, although other halophytes may be more suited or even more adapted for the end goal. Such halophytes include Chlorella vulgaris, which is also very well reported in the bibliography for its desalination properties [12,45].
The amount of inoculate undoubtedly plays a significant part in the effectiveness of the microalgae in its new media; a future study may introduce additional inoculates or directly add the NaCl to the culture, while ensuring that all other parameters are maintained.
In terms of the industry, the final results proved that Salinity 75 is an excellent salinity for cell count increase, higher dry biomass values, and viable pigment production. Reviewing these four parameters, one may conclude that the microalgae is still comfortable at Salinity 75, but as a desalination agent no activity was shown.
Temperature control was not considered. For a more realistic picture of the final application, temperature should be observed in the field and constantly monitored in the laboratory.
Author Contributions
Conceptualization, J.R.T. and G.W.F.; methodology, J.R.T. and G.W.F.; validation, J.R.T. and G.W.F.; formal analysis, J.R.T.; investigation, J.R.T. and G.W.F.; resources, Y.C., P.M. and R.P.; data curation, J.R.T.; writing—original draft preparation, J.R.T.; writing—review and editing, J.R.T., G.W.F., P.M., R.P. and Y.C.; visualization, J.R.T.; supervision, P.M., R.P. and Y.C.; project administration, J.R.T. and G.W.F.; funding acquisition, Y.C., P.M. and R.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Regional University Network—European Union.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in the study are included in the article, further inquiries can be directed to the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Chenoweth, J. Minimum water requirement for social and economic development. Desalination 2008, 229, 245–256. [Google Scholar] [CrossRef]
- Lall, U.; Heikkila, T.; Siegfried, T. Water in the 21st century: Defining the elements of global crises and potential solutions on jstor. J. Int. Aff. Editor. Board 2008, 61, 1–17. Available online: https://www.jstor.org/stable/24358108 (accessed on 13 December 2023).
- Bromley, D.W.; Taylor, D.C.; Parker, D.E. Water Reform and Economic Development: Institutional Aspects of Water Management in the Developing Countries. Econ. Dev. Cult. Chang. 1980, 28, 365–387. [Google Scholar] [CrossRef]
- Brown, C.; Lall, U. Water and economic development: The role of variability and a framework for resilience. Nat. Resour. Forum 2006, 30, 306–317. [Google Scholar] [CrossRef]
- Bhateria, R.; Jain, D. Water quality assessment of lake water: A review. Sustain. Water Resour. Manag. 2016, 2, 161–173. [Google Scholar] [CrossRef]
- Ayoub, J.; Alward, R. Water requirements and remote arid areas: The need for small-scale desalination. Desalination 1996, 107, 131–147. [Google Scholar] [CrossRef]
- McGinn, P.J.; Dickinson, K.E.; Park, K.C.; Whitney, C.G.; MacQuarrie, S.P.; Black, F.J.; Frigon, J.-C.; Guiot, S.R.; O’Leary, S.J. Assessment of the bioenergy and bioremediation potentials of the microalga Scenedesmus sp. AMDD cultivated in municipal wastewater effluent in batch and continuous mode. Algal Res. 2012, 1, 155–165. [Google Scholar] [CrossRef]
- Pittman, J.K.; Dean, A.P.; Osundeko, O. The potential of sustainable algal biofuel production using wastewater resources. Bioresour. Technol. 2011, 102, 17–25. [Google Scholar] [CrossRef]
- Abdel-Raouf, N.; Al-Homaidan, A.A.; Ibraheem, I.B.M. Microalgae and wastewater treatment. Saudi J. Biol. Sci. 2012, 19, 257–275. [Google Scholar] [CrossRef]
- Curto, D.; Franzitta, V.; Guercio, A. A Review of the Water Desalination Technologies. Appl. Sci. 2021, 11, 670. [Google Scholar] [CrossRef]
- Gao, L.; Zhang, J.; Liu, G. Life cycle assessment for algae-based desalination system. Desalination 2021, 512, 115148. [Google Scholar] [CrossRef]
- Ghobashy, M.O.I.; Bahattab, O.; Alatawi, A.; Aljohani, M.M.; Helal, M.M.I. A Novel Approach for the Biological Desalination of Major Anions in Seawater Using Three Microalgal Species: A Kinetic Study. Sustainability 2022, 14, 7018. [Google Scholar] [CrossRef]
- Mohsenpour, S.F.; Hennige, S.; Willoughby, N.; Adeloye, A.; Gutierrez, T. Integrating micro-algae into wastewater treatment: A review. Sci. Total. Environ. 2021, 752, 142168. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Zhang, X.; Fan, L.; Gray, S.; Li, M. Algae-Based Approach for Desalination: An Emerging Energy-Passive and Environmentally Friendly Desalination Technology. ACS Sustain. Chem. Eng. 2021, 9, 8663–8678. [Google Scholar] [CrossRef]
- El-Sayed, A.B.; El-Fouly, M.M.; El-Nour, E.-Z.A.E.-M.A.A. Immobilized Microalga Scenedesmus sp. for Biological Desalination of Red Sea Water: I. Effect on Growth. Nat. Sci. 2010, 8, 69–76. Available online: https://www.researchgate.net/publication/236983851_Immobilized_Microalga_Scenedesmus_sp_for_Biological_Desalination_of_Red_Sea_Water_I_Effect_on_Growth (accessed on 25 May 2024).
- Maru, M.; Sahle-Demissie, E.; Zewge, F. A review on biodesalination using halophytic microalgae: Opportunities and challenges. J. Water Supply Res. Technol. 2021, 70, jws2021068. [Google Scholar] [CrossRef]
- De-Bashan, L.E.; Bashan, Y. Immobilized microalgae for removing pollutants: Review of practical aspects. Bioresour. Technol. 2010, 101, 1611–1627. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, R.; Guieysse, B. Algal–bacterial processes for the treatment of hazardous contaminants: A review. Water Res. 2006, 40, 2799–2815. [Google Scholar] [CrossRef] [PubMed]
- Uma, L.; Selvaraj, K.; Subramanian, G.; Nagarkar, S.; Manjula, R. Biotechnological Potential of Marine Cyanobacteria in Wastewater Treatment—Disinfection of Raw Sewage by Oscillatoria willei BDU 130511. J. Microbiol. Biotechnol. 2002, 12, 699–701. Available online: https://www.jmb.or.kr/journal/view.html?spage=699&volume=12&number=4 (accessed on 13 December 2023).
- Mallick, N. Biotechnological potential of immobilized algae for wastewater N, P and metal removal: A review. BioMetals 2002, 15, 377–390. [Google Scholar] [CrossRef]
- Moayedi, A.; Yargholi, B.; Pazira, E.; Babazadeh, H. Investigated of Desalination of Saline Waters by Using Dunaliella salina Algae and Its Effect on Water Ions. Civ. Eng. J. 2019, 5, 2450–2460. [Google Scholar] [CrossRef]
- Sergany, E.; Hosseiny, E.; Nadi, E. The Optimum Algae Dose in Water Desalination by Algae Ponds. Int. Res. J. Adv. Eng. Sci. 2019, 4, 152–154. [Google Scholar]
- Laliberté, G.; Lessard, P.; de la Noüe, J.; Sylvestre, S. Effect of phosphorus addition on nutrient removal from wastewater with the cyanobacterium Phormidium bohneri. Bioresour. Technol. 1997, 59, 227–233. [Google Scholar] [CrossRef]
- Burkholder, J.M.; Glibert, P.M.; Skelton, H.M. Mixotrophy, a major mode of nutrition for harmful algal species in eutrophic waters. Harmful Algae 2008, 8, 77–93. [Google Scholar] [CrossRef]
- Martínez, M.E.; Sánchez, S.; Jiménez, J.M.; El Yousfi, F.; Muñoz, L. Nitrogen and phosphorus removal from urban wastewater by the microalga Scenedesmus obliquus. Bioresour. Technol. 2000, 73, 263–272. [Google Scholar] [CrossRef]
- Dunaliella salina (Dunal) Teodoresco: AlgaeBase. Available online: https://www.algaebase.org/search/species/detail/?species_id=27814 (accessed on 20 May 2024).
- Farhat, N.; Rabhi, M.; Falleh, H.; Jouini, J.; Abdelly, C.; Smaoui, A. Optimization of salt concentrations for a higher carotenoid production in Dunaliella salina (chlorophyceae)1. J. Phycol. 2011, 47, 1072–1077. [Google Scholar] [CrossRef]
- Ehrenfeld, J.; Cousin, J.-L. Ionic regulation of the unicellular green alga Dunaliella tertiolecta: Response to hypertonic shock. J. Membr. Biol. 1984, 77, 45–55. [Google Scholar] [CrossRef]
- Katz, A.; Bental, M.; Degani, H.; Avron, M. In vivo pH regulation by a Na+/H+ antiporter in the halotolerant alga Dunaliella salina. Plant Physiol. 1991, 96, 110–115. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Katz, A.; Kaback, H.; Avron, M. Na+/H+ antiport in isolated plasma membrane vesicles from the halotolerant alga Dunaliella salina. FEBS Lett. 1986, 202, 141–144. [Google Scholar] [CrossRef]
- Katz, A.; Kleyman, T.R.; Pick, U. Utilization of Amiloride Analogs for Characterization and Labeling of the Plasma Membrane Na+/H+ Antiporter from Dunaliella salina. Biochemistry 1994, 33, 2389–2393. [Google Scholar] [CrossRef]
- Mishra, A.; Mandoli, A.; Jha, B. Physiological characterization and stress-induced metabolic responses of Dunaliella salina isolated from salt pan. J. Ind. Microbiol. Biotechnol. 2008, 35, 1093–1101. [Google Scholar] [CrossRef]
- Rippka, R. Isolation and Purification of Cyanobacteria. Methods Enzymol. 1988, 167, 3–27. [Google Scholar] [CrossRef]
- Carr, C.J.; Scoville, J.; Ruble, J.; Condie, C.; Davis, G.; Floyd, C.L.; Kelly, L.; Monson, K.; Reichert, E.; Sarigul, B.; et al. An Audit and Comparison of pH, Measured Concentration, and Particulate Matter in Mannitol and Hypertonic Saline Solutions. Front. Neurol. 2021, 12, 667842. [Google Scholar] [CrossRef]
- Uchida, H.; Kayakawa, Y.; Maeda, Y. Ultra high-resolution seawater density sensor based on a refractive index measurement using the spectroscopic interference method. Nat. Sci. Rep. 2019, 9, 15482. [Google Scholar] [CrossRef]
- Lewis, E.L.; Perkin, R.G. Salinity: Its definition and calculation. J. Geophys. Res. Oceans 1978, 83, 466–478. [Google Scholar] [CrossRef]
- Škufca, D.; Božič, D.; Hočevar, M.; Jeran, M.; Zavec, A.B.; Kisovec, M.; Podobnik, M.; Podobnik, T.; Tomazin, R.; Iglič, A.; et al. Interaction between Microalgae P. tricornutum and Bacteria Thalassospira sp. for Removal of Bisphenols from Conditioned Media. Int. J. Mol. Sci. 2022, 23, 8447. [Google Scholar] [CrossRef] [PubMed]
- Marie, D.; Simon, N.; Vaulot, D. Algal Culturing Techniques; Academic Press: Cambridge, MA, USA, 2005. [Google Scholar]
- Moheimani, N.R.; Borowitzka, M.A.; Isdepsky, A.; Sing, S.F. Standard methods for measuring growth of algae and their composition. Algae Biofuels Energy 2013, 5, 265–284. [Google Scholar] [CrossRef]
- Strickland, J.D.H.; Parsons, T.R. A Practical Handbook of Seawater Analysis. Ottawa: Fisheries Research Board of Canada, Bulletin 167, 1968. 293 pp. Int. Rev. der Gesamten Hydrobiol. Hydrogr. 1970, 55, 167. [Google Scholar] [CrossRef]
- Jeffrey, S.W.; Humphrey, G.F. New spectrophotometric equations for determining chlorophylls a, b, c1 and c2 in higher plants, algae and natural phytoplankton. Biochem. Physiol. Pflanz. 1975, 167, 191–194. [Google Scholar] [CrossRef]
- ESS Method 150.1: Chlorophyll-Spectrophotometric. Environ. Sci. Sect. 1991. Available online: http://polk.wateratlas.usf.edu/upload/documents/methd150.pdf (accessed on 25 April 2024).
- Chen, L.; Li, D.; Song, L.; Hu, C.; Wang, G.; Liu, Y. Effects of Salt Stress on Carbohydrate Metabolism in Desert Soil Alga Microcoleus vaginatus Gom. J. Integr. Plant Biol. 2006, 48, 914–919. [Google Scholar] [CrossRef]
- El Arroussi, H.; Benhima, R.; Bennis, I.; El Mernissi, N.; Wahby, I. Improvement of the potential of Dunaliella tertiolecta as a source of biodiesel by auxin treatment coupled to salt stress. Renew. Energy 2015, 77, 15–19. [Google Scholar] [CrossRef]
- Moayedi, A.; Yargholi, B.; Pazira, E.; Babazadeh, H. Investigation of bio-desalination potential algae and their effect on water quality. Desalination Water Treat. 2021, 212, 78–86. [Google Scholar] [CrossRef]
- Priya, A.; Gnanasekaran, L.; Dutta, K.; Rajendran, S.; Balakrishnan, D.; Soto-Moscoso, M. Biosorption of heavy metals by microorganisms: Evaluation of different underlying mechanisms. Chemosphere 2022, 307, 135957. [Google Scholar] [CrossRef]
- Ramesh, B.; Saravanan, A.; Kumar, P.S.; Yaashikaa, P.; Thamarai, P.; Shaji, A.; Rangasamy, G. A review on algae biosorption for the removal of hazardous pollutants from wastewater: Limiting factors, prospects and recommendations. Environ. Pollut. 2023, 327, 121572. [Google Scholar] [CrossRef]
- Wei, J.; Gao, L.; Shen, G.; Yang, X.; Li, M. The role of adsorption in microalgae biological desalination: Salt removal from brackish water using Scenedesmus obliquus. Desalination 2020, 493, 114616. [Google Scholar] [CrossRef]
- Çelekli, A.; Bozkurt, H. Bio-sorption of cadmium and nickel ions using Spirulina platensis: Kinetic and equilibrium studies. Desalination 2011, 275, 141–147. [Google Scholar] [CrossRef]
- Mirzaei, M.; Jazini, M.; Aminiershad, G.; Refardt, D. Biodesalination of saline aquaculture wastewater with simultaneous nutrient removal and biomass production using the microalgae Arthrospira and Dunaliella in a circular economy approach. Desalination 2024, 581, 117564. [Google Scholar] [CrossRef]
- Sedjati, S.; Santosa, G.; Yudiati, E.; Supriyantini, E.; Ridlo, A.; Kimberly, F. Chlorophyll and Carotenoid Content of Dunaliella salina at Various Salinity Stress and Harvesting Time. IOP Conf. Ser. Earth Environ. Sci. 2019, 246, 012025. [Google Scholar] [CrossRef]
- Ma, X.; Zhou, W.; Fu, Z.; Cheng, Y.; Min, M.; Liu, Y.; Zhang, Y.; Chen, P.; Ruan, R. Effect of wastewater-borne bacteria on algal growth and nutrients removal in wastewater-based algae cultivation system. Bioresour. Technol. 2014, 167, 8–13. [Google Scholar] [CrossRef]
- Morowvat, M.H.; Ghasemi, Y. Culture medium optimization for enhanced β-carotene and biomass production by Dunaliella salina in mixotrophic culture. Biocatal. Agric. Biotechnol. 2016, 7, 217–223. [Google Scholar] [CrossRef]
- Djunaedi, A.; Suryono, C.A.; Sardjito, S. Kandungan Pigmen Polar Dan Biomassa Pada Mikroalga Dunaliella salina Dengan Salinitas Berbeda. J. Kelaut. Trop. 2017, 20, 1–6. [Google Scholar] [CrossRef]
- Félix, F.C.C.d.S.; Hidalgo, V.B.; de Carvalho, A.K.F.; Caetano, N.d.S.; Da Rós, P.C.M. Assessing the application of marine microalgae Dunaliella salina in a biorefinery context: Production of value-added biobased products. Biofuels Bioprod. Biorefining 2024, 18, 439–452. [Google Scholar] [CrossRef]
- Schagerl, M.; Siedler, R.; Konopáčová, E.; Ali, S.S. Estimating Biomass and Vitality of Microalgae for Monitoring Cultures: A Roadmap for Reliable Measurements. Cells 2022, 11, 2455. [Google Scholar] [CrossRef] [PubMed]
- Fisher, T.; Minnaard, J.; Dubinsky, Z. Photoacclimation in the marine alga Nannochloropsis sp. (Eustigmatophyte): A kinetic study. J. Plankton Res. 1996, 18, 1797–1818. [Google Scholar] [CrossRef]
- Colusse, G.A.; Mendes, C.R.B.; Duarte, M.E.R.; de Carvalho, J.C.; Noseda, M.D. Effects of different culture media on physiological features and laboratory scale production cost of Dunaliella salina. Biotechnol. Rep. 2020, 27, e00508. [Google Scholar] [CrossRef]
- Sarkheil, M.; Ameri, M.; Safari, O. Application of alginate-immobilized microalgae beads as biosorbent for removal of total ammonia and phosphorus from water of African cichlid (Labidochromis lividus) recirculating aquaculture system. Environ. Sci. Pollut. Res. 2022, 29, 11432–11444. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).