
Unraveling Phylogenetic Relationships via A. carbonarius and A. tubingensis Sequence Analyses †
1
Phytopathology and Detection Methodology Unit (PMDV) Mycomol Laboratory, INRAE Versailles Route de Saint Cyr, 78026 Versailles, France
2
Center of Biotechnology of Borj Cerdria-Tunisia, Laboratory of Molecular Plant Physiology, B.P. 901, Hammam-Lif 2050, Tunisia
3
Clinical Biochemistry & Toxicology Laboratory, University of Benin (UNIBEN), Nigeria 300213, Benin
*
Author to whom correspondence should be addressed.
†
Presented at the 2nd International Electronic Conference on Microbiology, 1–15 December 2023; Available online: https://ecm2023.sciforum.net.
Biol. Life Sci. Forum 2024, 31(1), 31; https://doi.org/10.3390/ECM2023-16445
Published: 30 November 2023
(This article belongs to the Proceedings of The 2nd International Electronic Conference on Microbiology)
Abstract
:The aspergilli comprise a diverse group of filamentous fungi spanning over 200 million years of evolution. In this work, we report a clarification of the phylogenetic relationship between A. carbonarius and A. tubingensis via the screening of ITS-5.8RDNA, (niiA-niaD), ß-tubulin, and eEF-1 genes. The phylogenetic analysis of aspergilli ITS-5.8rDNA sequences divided A. carbonarius and A. tubingensis into two coherent clusters and showed a close intergeneric relationship, which is in keeping with the existing morphological and taxonomic classification. Herein, A. carbonarius exhibits a complete similarity (100%) for the three target genes: intergenic region of (niiA-niaD), ß-tubulin, and eEF-1. The tree resulting from a bootstrap analysis indicates branch points when the significance is 98% minimum for each gene (ß-tubulin and eEF-1). The phylogenetic analysis of the aspergilli genome structure provided a quantitative evaluation of the forces driving long-term eukaryotic genome evolution. It also led to an experimentally validated model of mating-type locus evolution, suggesting the potential for sexual reproduction in A. carbonarius and A. tubingensis. This is the first work describing the evolutionary relationship between both species. Nevertheless, this discovery might be viewed as typical during the process of new taxonomic evolution.
1. Introduction
The Aspergillus genus is widespread in nature [1,2,3,4], and many species of this genus have significant negative effects on agriculture, particularly the production of mycotoxins such as ochratoxin A [1,5,6]. The filamentous fungi A. carbonarius and A. tubingensis have been extensively employed in genetic research, and toxigenic potential studies can be carried out on them [6]. The concern about these filamentous fungi in the vineyard has traditionally been linked to spoilage of grapes due to fungal growth. Nevertheless, these two saprobic fungi can produce mycotoxins in grapes [7,8,9]. The phenotypic concept is the classic approach based on the morphological characteristics suggesting that both species are morphologically far [10,11,12].
Partly due to its economic importance, the genus Aspergillus has one of the better classifications among filamentous fungi [11,13,14]. Raper and Fennell, in 1965, described 18 species groups within this genus based mainly on cultural and morphological features [8]. These were treated as sections belonging to six subgenera by Gams et al., 1985 [15]. Phylogenetic studies of ribosomal RNA gene sequences led to the acceptance of 3 subgenera with a total of 15 sections [16]. In the present work, considerable attention was devoted to developing molecular tools to give a general overview of the taxonomic relationships between A. carbonarius and A. tubingensis based mainly on our recent studies.
The techniques applied included PCR-based methods and phylogenetic analysis of ITS-5.8rDNA (niiA-niaD), ß-tubulin, and eEF-1 gene sequences in comparison with morphological features.
2. Methods
2.1. Organisms, Growth Conditions, and DNA Isolation
It is crucial for practical purposes that fungi are isolated and correctly identified. By appropriately adjusting the temperature and medium composition, it is possible to preserve the two morphological stages of fungal species in a lab setting; mycelia cells developed in vitro at a temperature of 25 °C in the dark. Strains from Tunisian grapes were employed in this investigation. A. carbonarius CBS 120167 and A. tubingensis strain CBS 11732 were used and collected from the American Type Culture Collection (ATCC) and employed as references. For subsequent uses, mycelia were collected by filtration via filter paper or Miracloth (Calcbiochem, Darmstadt, Germany), frozen in liquid nitrogen, crushed, and kept at −80 °C. Using the Fungal DNA miniprep kit (QIAGEN-France), fungal DNA was extracted from frozen mycelial powder in accordance with the manufacturer’s instructions. The DNA levels were measured using a NanoDrop® ND-1000 spectrophotometer (Nanodrop Technologies, Wilmington, NC, USA) at 260 nm and adjusted to approximately 25 ng/µL.
2.2. PCR Amplification, Cloning and Sequencing
The PCR amplification of ITS-5.8rDNA, the intergenic region of (niiA-niaD), ß-tub, and eEF1 genes were run in a “GeneAmp PCR 2700” thermal-cycler (AB-Applied Biosystem, CA, USA). PCR reactions were performed using the Hot Gold Star Mix (Eurogentec, Belgium). Fungal genomic DNA was used as a template to target the ITS-5.8rDNA, the intergenic region of (niiA-niaD), ß-tub, and eF1 genes in a 50 µL reaction mixture containing 25 µL Hot Gold Star Mix, 1.2 µL of each primer (PN1-PN34, Nit 462F-Nit 873R, ßtub-ßtubR), and eEF1-1526F-eEF1-1567R. The reactions were performed with the following program: initial denaturation at 95 °C for 1 min; 35 cycles, and each cycle consisted of three steps of 95 °C for 1 min; an annealing temperature (X) °C (Table 1) for ITS- 5.8rDNA, (niiA-niaD), eEF1, and ß-tub; and a last step at 72 °C for 1 min followed by a final incubation at 72 °C for 10 min. The products amplified were separated via horizontal electrophoresis on 2% agarose gel using Tris–Boron–EDTA buffer (TBE) 0.1X for 1 h at 100 V. Fragment size was estimated using a suitable molecular marker (Smart DNA Ladder, Eurogentec, Belgium), and the PCR products were purified using PEG (Polyethylene Glycol) and purification spin-columns.
The clones amplified by PCR were subcloned into the pGEMT vector (Promega) and, after heat shock, cloned in chemically competent Top10 one shot ® cells of Escherichia coli (Invitrogen, Karlsruhe, Germany). The primers PU-PR were used for cycle sequencing in a programmed GeneAmp PCR 2700 thermal-cycler (AB-Applied Biosystem, CA, USA). (Table 1) as insert-specific primers in the vector “pGEMT” with the following temperature profile: 20 s at 96 °C, 20 s at 50 °C, 4 min at 60 °C, for 30 cycles. The CEQTM 8000 Genetic Analysis System was used to perform the sequencing process.
2.3. Sequence Alignment and Phylogenetic Analyses
Nucleotide sequences were determined using sequence data, and predictions of the protein structure were analyzed using the Clustal W method with the MegAlign program of DNASTAR software (Lasergene, WI, USA).
3. Results
3.1. Screening of ITS-5.8 S rDNA in A. carbonarius and A. tubingensis
Interspecific variability of Aspergillus species assigned to section Nigri was also examined using phenotypic features and sequences of the intergenic transcribed spacer regions (ITS region) and the 5.8 S rDNA gene (Figure 1). The phylogenetic analysis of sequence data indicated that A. carbonarius and A. tubingensis are presented in two distinct groups. This result was also supported by phenotypic data.
The uniseriate species A. carbonarius and A. ibericus are clustered together and defined in a second clade, while A. awamori, A. foetidus, A. tubingensis, A. niger, and A. brasiliensis are on a separate branch with respect to other black Aspergillus species, according to the phylogenetic analysis of ITS sequence data. At least several species belong to this section. The last Aspergillus strain, A. flavus, was used as a control (Figure 1).
3.2. Screening and Phylogenetic Analysis of (niiA-niaD) Gene in A. carbonarius and A. tubingensis
The intergenic portion of the (niiA-niaD) gene was isolated within A. carbonarius and A. tubingensis. The nucleotide sequence suggests that A. carbonarius has two distinct DNA fragments; one of these (400 pb) was also amplified in A. tubingensis (Figure 2).
The alignment and the comparison of the two sequences suggest a full homology between both Aspergillus species. Following this result, a hypothesis was expressed about the existence of a phylogenetic relationship between both species. To confirm this hypothesis, two genes were targeted: ß-tubulin and eEF-1.
3.3. Screening and Phylogenetic Analysis of ß-Tubulin Sequences
The ß-tubulin gene was used to probe the genomic DNA of A. carbonarius and A. tubingensis. The alignment of the ß-tubulin genes isolated from A. carbonarius, A. tubingensis, A. carbonarius (CBS 120167), and A. tubingensis (CBS 11732) were used as a reference strain and gave evidence for a high degree of similarity (100%). In addition, the amino acid sequence deduced from the nucleotide sequence of ß-tubulin was used to develop a phylogenetic tree which indicates that A. carbonarius and A. tubingensis were clustered together in the same branch with amino acid sequences of A. tubingensis (DQ902579) and A. carbonarius (AY585532).
Isolates of A. carbonarius (AY585532), A. tubingensis (DQ902579), A. tubingensis 1–2, A. tubingensis (CBS 11732), A. carbonarius (CBS120167), and A. carbonarius 1–2 are clustered in group A. A. foetidus (DQ768454) and A. costaricaensis (AY820014) are clustered in group B. Group C is composed of two branches with A. brasiliensis (DQ900611), A. niger (AY80001), (AY585535), and A. lacticoffeatus (AM419748). The last group consists of A. ibericus (AM419748) and A. sclerotioniger (AY819996).
The tree results from a bootstrap analysis that indicates branch points when the significance is 98% minimum for A. tubingensis and A. carbonarius species (Figure 3). Our results suggest that the phylogenetic analysis of the ß-tubulin DNA sequence strongly supported the hypothesis.
3.4. Screening and Phylogenetic Analysis of eEF-1 Sequences
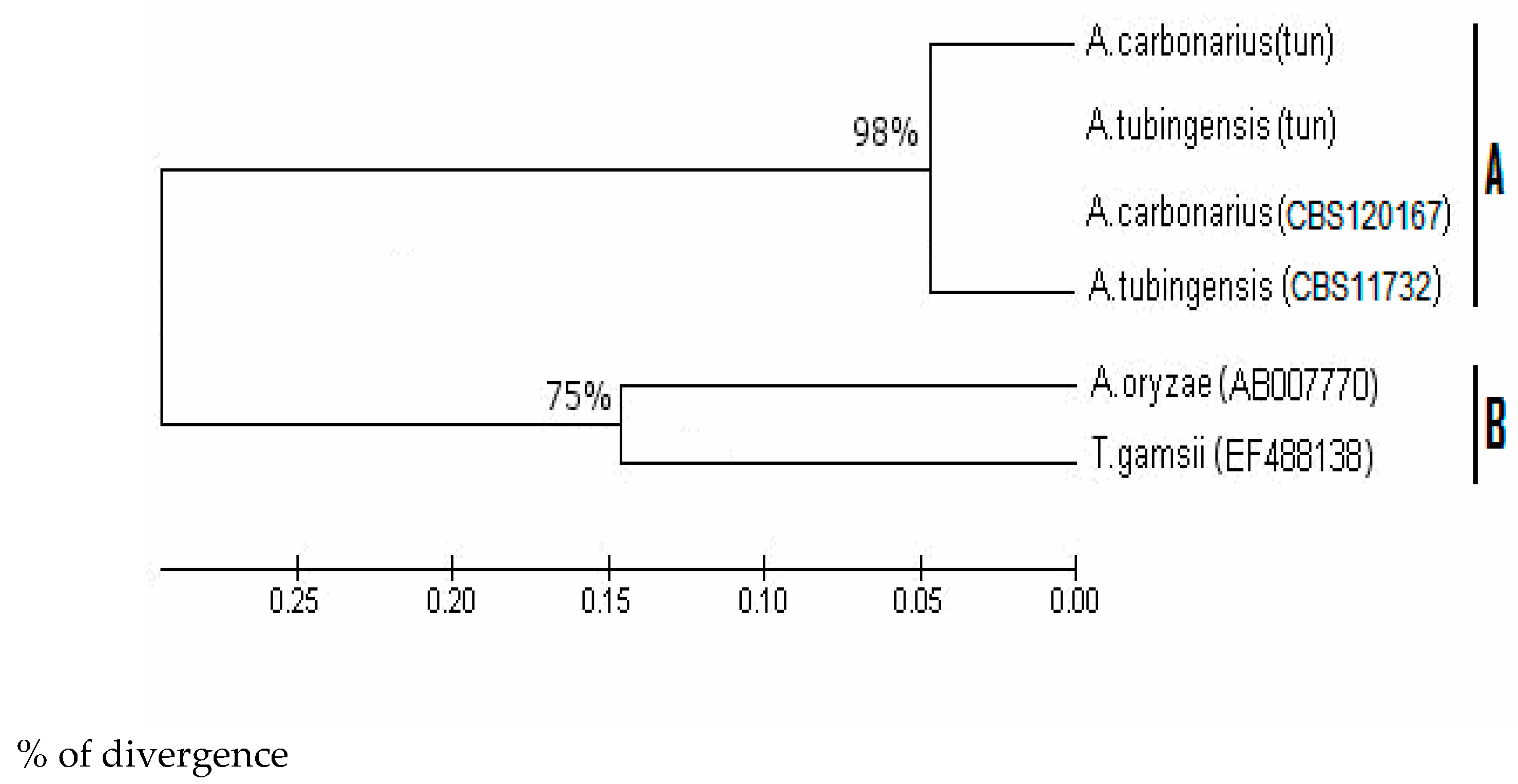
The alignment of eEF-1 sequences showed a full similarity between Aspergillus species (A. carbonarius, A. carbonarius (CBS 120167), A. tubigensis, and A. tubigensis (CBS 11732).
Two distinct groups within the eEF-1 sequences were obtained for the four isolates of Ascomycota (Aspergillus carbonarius, tubigensis, oryzae, and Trichoderma gamsii). Isolates within group A were separated from those in group B by approximately 15 base mutations.
The phylogenetic tree results from a bootstrap analysis that indicates branch points when the significance is 98% minimum within group A compared to 75% across isolates within group B (Figure 4).
The amino acid sequences were evident from both the visual inspection of the aligned sequences and the dendrogram produced via an analysis of the sequence data. The identification of an evolutionary link between the two species was highly confirmed by DNA sequences from ß-tubulin and eEF-1, supporting the hypothesis.
4. Discussion
Modern molecular tools used to identify phylogenetic relationships between fungi have opened new possibilities for expanding insight into and knowledge of this area. In recent years, the use of multiple gene analyses for phylogenetic studies has become more popular, and several studies based on protein-coding genes have been published, including fungal taxa [17,18,19]. De Jong et al. (2001) showed that the analysis of multiple loci can provide more resolution and therefore more reliable phylogenetic hypotheses in fungal phylogeny. The phylogenetic analysis of sequences of the ribosomal ITS-5.8rDNA gene cluster and the ß-tubulin gene were found to be very useful for clarifying taxonomic relationships among toxigenic aspergilla [2].
In this report, ITS-5.8rDNA was used for A. carbonarius and A. tubingensis, and analysis led to two far distinguished species. In fact, previous work based on large subunit rDNA data has led to a widely accepted phylogeny of the aspergilli in which A. tubigensis and A. niger are more related to one another than A. carbonarius [4,20]. However, single-gene phylogenies can contradict organismal phylogenies. In principle, whole-genome data provide greater resolving power by allowing trees to be constructed based on concatenated sets of genes. Using this approach to study the relationship of both aspergilli (A. tubingensis and A. carbonarius), we find support for an alternative phylogeny. In the current study, we reveal that 400 bp of the target gene (niiA-niaD) are subjected to a conserved sequence that separates A. tubingensis from A. carbonarius. PCR assay for (niiA- niaD) was conducted to a hypothesis of an evolution relationship. In disagreement with our previous work, the intergenic region of (niiA-niaD) was used to distinguish between A. niger and A. tubigensis, and two primer sets were designed for each species [6,13]. Here, our results based on sequence alignment and the phylogenetic analysis of the eEF-1 and β-tubulin strongly supported the recognition of this process of evolution within A. carbonarius and A. tubingensis.
According to many studies, the genes for tubulins, especially for β-tubulin, are receiving increasing attention in the investigation of evolutionary relationships at all levels: (i) in kingdom-level phylogenetic analyses [17,21] and (ii) in studies of complex species groups within protists, animals, fungi, and plants [13,22,23,24].
In agreement with several studies, β-tubulin was usually used to detect evolutions in fungal species [18,21].
Our hypothetical dataset is also used to demonstrate how genealogical relationships and population parameters can be inferred using PCR methods.
The basic principles of these approaches are graphically presented by phylogenetic trees, along with useful references such as A. tubingensis CBS 1732, A. carbonarius CBS 120167 and comments on key assumptions implicit in methods currently available after targeting β- tubulin and elongation factor 1 (eEF1). In agreement with previous studies, improved phylogenetic discrimination at the species and intraspecific level can be achieved via the analysis of genomic regions other than the ITS [16,25].
In this respect, O’Donnell and co-workers described a new strategy using β-tubulin and ITS marks to identify new strains in the Fusarium genus and explain that there are several factors relating to genetic, geographic, and ecological barriers which influence the chances of evolution genome in fungi, with useful references such as A. tubingensis CBS 1732, A. carbonarius CBS 120167, and comments on key assumptions implicit in methods currently available after targeting β- tubulin and elongation factor 1 (eEF1). In agreement with previous studies, improved phylogenetic discrimination at the species and intraspecific level can be achieved via the analysis of genomic regions other than the ITS [16,25]. We focus on the gene encoding elongation factor 1 (eEF-1) in order to investigate the genetic diversity amongst Aspergillus isolates. Our results suggested a full homology sequence between A. carbonarius and A. tubingensis, which confirms our hypothesis. Moreover, the eEF-1 gene usually exhibits greater variability than β-tubulin gene sequences that are often used in taxonomic phylogenetics. Moreover, analysis of the eEF-1 sequence data has previously permitted the identification of intraspecific variation [26,27,28].
To date, the number of ongoing filamentous fungal genome sequencing projects is almost tenfold fewer than those of bacterial and archaeal genome projects. The fungi chosen for sequencing represent narrow kingdom diversity; most are pathogens or models. We advocate for an ambitious, forward-looking phylogenetic-based genome sequencing program designed to capture metabolic diversity within the fungal kingdom, thereby enhancing research into alternative bioenergy sources, bioremediation, and fungal–environment interactions.
5. Conclusions
The study aimed to unravel the phylogenetic relationships between two important fungal species, A. carbonarius, and A. tubingensis, via sequence analyses. After an in-depth analysis of genetic data and comparison with related species, several key conclusions can be drawn from this research:
Phylogenetic Placement: The sequence analyses provided valuable insights into the evolutionary relationships of A. carbonarius and A. tubingensis within the Aspergillus genus. These two species were found to be closely related to each other, forming a distinct clade within the phylogenetic tree.
Species Distinctiveness: Despite their close phylogenetic relationship, this study also highlighted the genetic differences that distinguish A. carbonarius and A. tubingensis as distinct species. These differences may include unique genetic markers or variations in key genes, contributing to their ecological and functional differences.
Evolutionary History: This research shed light on the evolutionary history of these species, suggesting a common ancestor and subsequent divergence. Understanding their evolutionary trajectory can help explain their ecological niches, adaptation strategies, and potential differences in mycotoxin production, which are crucial for agricultural and food safety considerations.
Taxonomic Clarification: This study contributes to the taxonomic clarification of these fungal species. Accurate species identification and classification are fundamental in various fields, including agriculture, mycology, and food safety, where A. carbonarius and A. tubingensis play significant roles.
Applied Implications: Knowledge of the phylogenetic relationships between these fungi can have practical applications in agriculture, industry, and biotechnology. It might help with the creation of plans for bioremediation, managing mycotoxin levels, and creating valuable metabolites.
In conclusion, this research provides valuable insights into the phylogenetic relationships, genetic distinctiveness, and evolutionary history of A. carbonarius and A. tubingensis. These findings not only contribute to our understanding of fungal evolution and taxonomy but also have practical implications for various fields, emphasizing the importance of continued research in fungal genomics and phylogenetics. Further investigations may delve deeper into the functional implications of these genetic differences and their ecological relevance.
Author Contributions
Conceptualization: S.B.F.M. and A.M.; Methodology: S.B.F.M.; Software: S.B.F.M.; Validation: S.B.F.M. and A.M.; Formal analysis: S.B.F.M.; Investigation: S.B.F.M.; Resources: S.B.F.M.; Data curation: S.B.F.M.; writing—original draft preparation: S.B.F.M.; writing—review and editing: S.B.F.M., E.S.O. and A.M.; Visualization: E.S.O.; Supervision: A.M.; Project administration: A.M.; Funding acquisition: S.B.F.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the Unesco-l’Oréal for Women in Sciences granted for Mrs. Sabah BEN FREDJ MELKI in collaboration with the ministry of research in Tunisia.
Institutional Review Board Statement
This statement acknowledges the absence of human subjects in the research and emphasizes the organization’s commitment to ethical conduct across all research activities. It also encourages researchers to consider ethical considerations and seek guidance as needed, even in the absence of IRB oversight.
Informed Consent Statement
This statement ensures that participants are fully informed about the study before consenting to participate, thereby upholding ethical standards and protecting the rights and welfare of the participants.
Data Availability Statement
Data availability statements are available in section “MDPI Research Data Policies” at https://www.mdpi.com/ethics.
Acknowledgments
All individuals included in this section have consented to the acknowledgement. All the authors gratefully acknowledge the support of the PMDV Group—INRAE Versailles—France and the assistance of Professor Daniele Daffonchio (Department of Food, Nutrition and Environmental Sciences-University of Milan).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Abarca, M.L.; Bragulat, M.R.; Castella, G.; Cabanes, F.J. Ochratoxin A production by strains of Aspergillus niger var. niger. Appl. Environ. Microbiol. 1994, 60, 2650–2652. [Google Scholar] [CrossRef] [PubMed]
- De Jong, S.N.; Levesque, C.A.; Verkley, G.J.M.; Abeln, E.C.A.; Rahe, J.E.; Braun, P.G. Phylogenetic relationships among Neofabraea species causing tree cankers and bull’s-eye rot of apple based on DNA sequencing of ITS nuclear rDNA, mitochondrial rDNA, and the beta- tubulin gene. Mycol. Res. 2001, 105, 658–669. [Google Scholar] [CrossRef]
- Melki Ben Fredj, S.; Chebil, S.; Mliki, A. Isolation and characterization of ochratoxin A and aflatoxin B1 producing fungi infecting grapevines. Afr. J. Microbiol. Res. 2009, 3, 523–527. [Google Scholar]
- Schutze, J.; Krasko, A.; Custodio, M.R.; Efremova, S.M.; Muller, I.M.; Muller, W.E.G. Evolutionary relationships of Metazoa within the eukaryotes based on molecular data from Porifera. Proc. R. Soc. Lond. 1999, 266, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Melki Ben Fredj, S.; Gautier, A.; Brygoo, Y.; Mliki, A. Molecular strategy to discriminate between two ochratoxin A producing fungal species in Aspergillus niger aggregate group isolated from fresh and dried grapes. Ann. Microbiol. 2009, 59, 635–641. [Google Scholar] [CrossRef]
- Myburg, H.; Gryzenhout, M.; Heath, R.; Roux, J.; Wingfield, B.; Wingfield, M. Cryphonectria canker on Tibouchina in South Africa. Mycol. Res. 2002, 106, 1299–1306. [Google Scholar] [CrossRef]
- Keeling, P.J.; Luker, M.A.; Palmer, J.D. Evidence from beta-tubulin phylogeny that microsporidia evolved from within the fungi. Mol. Biol. Evol. 2000, 17, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Raper, K.B.; Fennell, D.I. The Genus Aspergillus; Williams & Wilkins: Baltimore, MD, USA, 1965. [Google Scholar] [CrossRef]
- Teren, J.; Varga, J.; Hamari, Z.; Rinyu, E.; Kevei, F. Immunochemical detection of ochratoxin A in black Aspergillus strains. Mycopathologia 1996, 134, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Gams, W.; Christensen, M.; Onions, A.H.S.; Pitt, J.I.; Samson, R.A. Infrageneric taxa of Aspergillus. In Advances in Penicillium and Aspergillus Systematics; Samson, R.A., Pitt, J.I., Eds.; Plenum Press: New York, NY, USA, 1985; pp. 55–61. [Google Scholar] [CrossRef]
- Hawksworth, D.L. Fungi and biodiversity: International incentives. Microbiologia 1997, 13, 221–226. [Google Scholar] [PubMed]
- Inui, T.; Takeda, Y.; Iizuka, H. Taxonomical studies on genus Rhizopus. J. Gen. Appl. Microbiol. 1965, 11, 1–121. [Google Scholar] [CrossRef]
- Edgcomb, V.P.; Roger, A.J.; Simpson, A.G.B.; Kysela, D.T.; Sogin, M.L. Evolutionary relationships among “jakobid” flagellates as indicated by alpha- and beta-tubulin phylogenies. Mol. Biol. Evol. 2001, 18, 514–522. [Google Scholar] [CrossRef] [PubMed]
- Keller, N.P. Fungal secondary metabolism: Regulation, function and drug discovery. Nat. Rev. Microbiol. 2019, 17, 167–180. [Google Scholar] [CrossRef] [PubMed]
- Hawksworth, D.L. The fungal dimension of biodiversity: Magnitude, significance, and conservation. Mycol. Res. 1991, 95, 641–655. [Google Scholar] [CrossRef]
- Peterson, S.W. Phylogenetic relationships in Aspergillus based on rDNA sequence analysis. In Integration of Modern Taxonomic Methods for Penicillium and Aspergillus Classification; Samson, R.A., Pitt, J.I., Eds.; Harwood Academic Publishers: Amsterdam, The Netherlands, 2000; pp. 323–355. [Google Scholar] [CrossRef]
- Baldauf, S.L.; Roger, A.J.; Wenk-Siefert, I.; Doolittle, W.F. A kingdom-level phylogeny of eukaryotes based on combined protein data. Science 2000, 290, 972–977. [Google Scholar] [CrossRef] [PubMed]
- Geiser, D.M.; Frisvad, J.; Taylor, J. Evolutionary relationships in Aspergillus section Fumigati inferred from partial ß-tubulin and hydrophobin DNA sequences. Mycologia 1998, 90, 831–845. [Google Scholar] [CrossRef]
- O’Donnell, K.; Cigelnik, E.; Casper, H.H. Molecular phylogenetic, morphological and mycotoxin data support reidentification of the Quorn mycoprotein fungus as Fusarium venenatum. Fungal Genet. Biol. 1998, 23, 57–67. [Google Scholar] [CrossRef]
- Eichler, E.E.; Sankoff, D. Structural dynamics of eukaryotic chromosome evolution. Science 2003, 301, 793–797. [Google Scholar] [CrossRef] [PubMed]
- Keeling, P.J.; Doolittle, W.F. Alphα-tubulin from early-diverging eukaryotic lineages and the evolution of the tubulin family. Mol. Biol. Evol. 1996, 13, 1297–1305. [Google Scholar] [CrossRef] [PubMed]
- Ayliffe, M.A.; Dodds, P.N.; Lawrence, G.J. Characterisation of a beta-tubulin gene from Melampsora lini and comparison of fungal beta-tubulin genes. Mycol. Res. 2001, 105, 818–826. [Google Scholar] [CrossRef]
- Mages, W.; Cresnar, B.; Harper, J.F.; Brüderlein, M.; Schmitt, R. Volvox carteri alpha-2- tubulin-encoding and beta-2-tubulin-encoding genes: Regulatory signals and transcription. Gene 1995, 160, 47–54. [Google Scholar] [CrossRef]
- Serra, R.; Lourenço, A.; Aliipio, P.; Venâncio, A. Influence of the region of origin on the mycobiota of grapes with emphasis on Aspergillus and Penicillium species. Mycol. Res. 2006, 110, 971–978. [Google Scholar] [CrossRef]
- Aoki, T.; O’Donnell, K. Morphological and molecular characterization of Fusarium pseudograminearum sp. nov., formerly recognised as the Group 1 population of F. graminearum. Mycologia 1999, 91, 597–609. [Google Scholar] [CrossRef]
- Hawksworth, D.L. The magnitude of fungal diversity: The 1.5 million species estimate revisited. Mycol. Res. 2001, 105, 1422–1432. [Google Scholar] [CrossRef]
- Maddison, W.P.; Maddison, D.R. MacClade: Analysis of Phylogeny and Character Evolution; Version 3; Sinauer Associates: Sunderland, MA, USA, 1992. [Google Scholar] [CrossRef]
- Melki Ben Fredj, S.; Chebil, S.; Lebrihi, A.; Lasram, S.; Ghorbel, A.; Mliki, A. Occurrence of pathogenic fungal species in Tunisian vineyards. Int. J. Food Microbiol. 2007, 113, 245–250. [Google Scholar] [CrossRef]
Figure 1.
Neighbor-joining tree of ITS sequences of species assigned to Aspergillus section Nigri, including A. niger, A. tubingensis, A. awamori, A. foetidus, and A. brasiliensis clustered in the same branch. Positive control A. flavus was employed, and A. carbonarius and A. ibericus were grouped together.
Figure 1.
Neighbor-joining tree of ITS sequences of species assigned to Aspergillus section Nigri, including A. niger, A. tubingensis, A. awamori, A. foetidus, and A. brasiliensis clustered in the same branch. Positive control A. flavus was employed, and A. carbonarius and A. ibericus were grouped together.
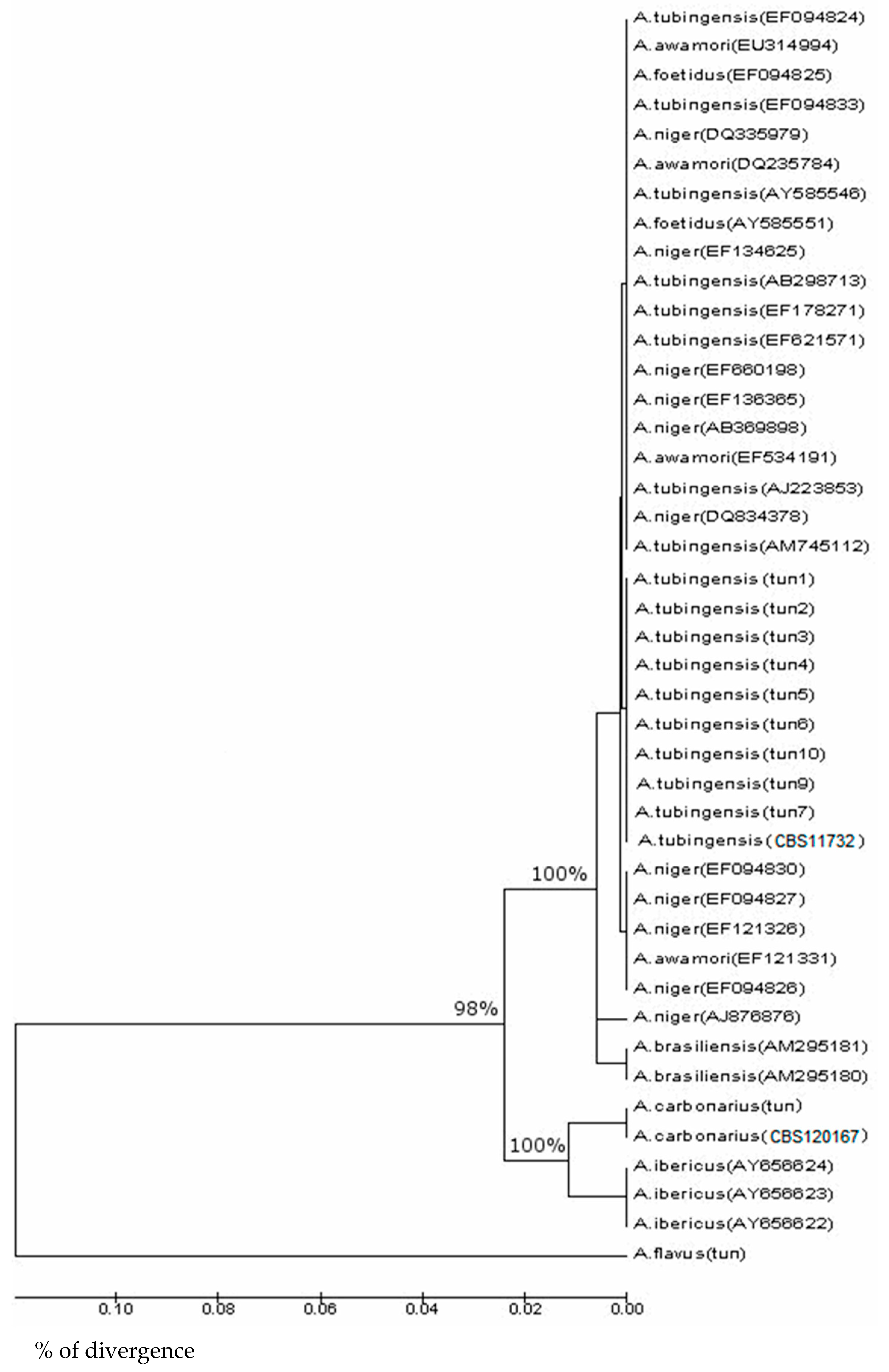
Figure 2.
Results of the PCR used to target 400 pb of the intergenic portion of (niiA-niaD). From the left to the right (AT1-AT3): A. tubingenis isolates from grapes; AC1-AC3: A. carbonarius isolates; T+: positive control; ATR: A. tubingenis CBS11732 (reference strain); ACR: A. carbonarius CBS 120167 (reference strain) and M: Size DNA markers.
Figure 2.
Results of the PCR used to target 400 pb of the intergenic portion of (niiA-niaD). From the left to the right (AT1-AT3): A. tubingenis isolates from grapes; AC1-AC3: A. carbonarius isolates; T+: positive control; ATR: A. tubingenis CBS11732 (reference strain); ACR: A. carbonarius CBS 120167 (reference strain) and M: Size DNA markers.
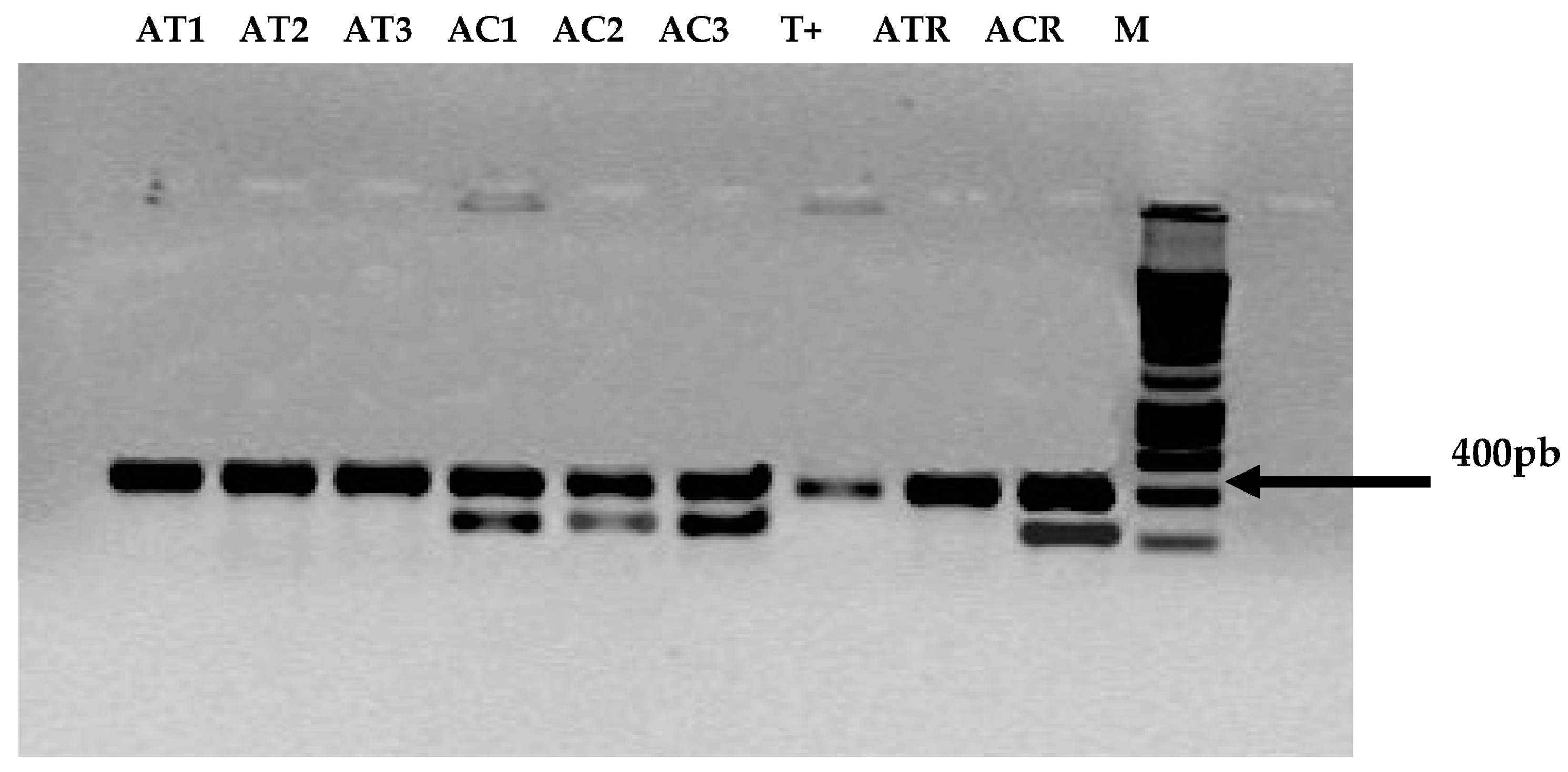
Figure 3.
Phylogenetic tree based on amino acid sequences of ß-tubulin within Ascomycetes: A. carbonarius (AY585532), A. tubingenis (DQ902579), A. tubingenis 1–2, A. tubingensis (CBS 11732), A. carbonarius (CBS 120167), and A. carbonarius 1–2 are clustered in group (A); A. foetidus (DQ768454) and A. costaricaensis (AY820014) are clustered in group (B) and related to group (A). The third group, (C), is composed of two branches containing A. brasiliensis (DQ900611), A. niger (AY80001 and AY585535), and A. lacticoffeatus (AM419748), and the group (D) consists of A. ibericus (AM419748) and A. sclerotioniger (AY819996). Bootstrap percentages were calculated and ranged from 85% to 100%.
Figure 3.
Phylogenetic tree based on amino acid sequences of ß-tubulin within Ascomycetes: A. carbonarius (AY585532), A. tubingenis (DQ902579), A. tubingenis 1–2, A. tubingensis (CBS 11732), A. carbonarius (CBS 120167), and A. carbonarius 1–2 are clustered in group (A); A. foetidus (DQ768454) and A. costaricaensis (AY820014) are clustered in group (B) and related to group (A). The third group, (C), is composed of two branches containing A. brasiliensis (DQ900611), A. niger (AY80001 and AY585535), and A. lacticoffeatus (AM419748), and the group (D) consists of A. ibericus (AM419748) and A. sclerotioniger (AY819996). Bootstrap percentages were calculated and ranged from 85% to 100%.

Figure 4.
Phylogenetic tree using eEF-1 amino acid sequences of Ascomycetes (A. carbonarius, A. tubingensis, A. oryzae (AB007770), and Trichoderma gamsii (EF488138). Bootstrap percentages were calculated and are indicated at nodes 98% for group (A) (A. carbonarius and A. tubingensis isolated from Tunisian grapes, A. carbonarius (CBS 120167), and A. tubingensis (CBS 11732)) and 75% for group (B) (A. oryzae and T. gamsii).
Figure 4.
Phylogenetic tree using eEF-1 amino acid sequences of Ascomycetes (A. carbonarius, A. tubingensis, A. oryzae (AB007770), and Trichoderma gamsii (EF488138). Bootstrap percentages were calculated and are indicated at nodes 98% for group (A) (A. carbonarius and A. tubingensis isolated from Tunisian grapes, A. carbonarius (CBS 120167), and A. tubingensis (CBS 11732)) and 75% for group (B) (A. oryzae and T. gamsii).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Primers used in the study.
| Primer Code | Primer Sequence (5′ to 3′) | Annealing Temperature (X) °C |
|---|---|---|
| PN1 PN34 | AGTAAAAGTCGTAACAAGG TTGCCGCTTCACTCGCCGTT | 55 |
| PU PR | CGTTGTAAAACGACGGCCAGT GTACCAGTATCGACAAAGGAG | 52 |
| Nit-462F Nit-873R | GTTACGGAAGCAAAGAGGTA GTTATTAGGGGCTATGGCAC | 54 |
| ß-tubF ß-tubR | CTCGAGCGTAGTAACGTCTAC AAACCCTGGAGGCAGTCGC | 55 |
| eEF1-526F eEF1-1567R | GTCGTYGTYATYGGHCAYGT ACHGT RCC RATACCACC RAT CTT | 58 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Melki, S.B.F.; Omoregie, E.S.; Mliki, A. Unraveling Phylogenetic Relationships via A. carbonarius and A. tubingensis Sequence Analyses. Biol. Life Sci. Forum 2024, 31, 31. https://doi.org/10.3390/ECM2023-16445
AMA Style
Melki SBF, Omoregie ES, Mliki A. Unraveling Phylogenetic Relationships via A. carbonarius and A. tubingensis Sequence Analyses. Biology and Life Sciences Forum. 2024; 31(1):31. https://doi.org/10.3390/ECM2023-16445
Chicago/Turabian StyleMelki, Sabah Ben Fredj, Ehi Sheena Omoregie, and Ahmed Mliki. 2024. "Unraveling Phylogenetic Relationships via A. carbonarius and A. tubingensis Sequence Analyses" Biology and Life Sciences Forum 31, no. 1: 31. https://doi.org/10.3390/ECM2023-16445