1. Introduction
Diabetes mellitus (DM), a disorder that affects about 422 million people worldwide, is fast becoming a global concern [
1]. Type 2 diabetes (DM 2) is the most common variant of DM and usually starts in later life (over the age of 40) and mainly in obese individuals. In this type, the insulin-producing cells in the pancreas produce insulin, but the output is inadequate for the body’s need, or there is a defect in liberation or/and action of insulin, commonly referred to as ‘insulin resistance’. Despite the introduction of several oral non-insulin drugs to treat DM 2, patient compliance remains poor due to severe side effects. Hence, the search for new molecules with reduced side effects still continues.
The conventional strategies of treating DM 2 include stimulation of endogenous insulin secretion, enhancing insulin action at the target site, use of oral hypoglycaemic agents, and inhibition of dietary starch digestion by glycosidases and alpha-amylases. Partial inhibition of carbohydrate digestion is one of the modes of action to reduce the blood glucose level. Pancreatic alpha-amylase (E.C. 3.2.1.1) plays a key role in the digestion of food and catalyses the hydrolysis of starch to produce oligosaccharides. These oligosaccharides are further digested by other enzymes to produce glucose which later on enters the bloodstream leading to postprandial hyperglycaemia. The correlation between the postprandial hyperglycaemia and activity of human pancreatic alpha-amylase has been reported previously. Inhibition of alpha-amylase results in delayed starch digestion and further reduces the rate of absorption of glucose into the blood [
2].
Natural products are widely used for many human ailments due to lesser side effects on humans. Among these secondary metabolites, flavonoids, the common group of polyphenols, are consumed by humans regularly in their diet. Flavonoids are mainly divided into anthocyanins and anthoxanthins. Anthocyanins occur in colourful flowers and fruits, while anthoxanthins are colourless compounds which are further categorised into flavones, flavans, flavonols, flavanols, isoflavones, and their glycosides [
3]. Flavonoids act as plants’ chemical defence compounds [
4].
Recently, scientists have proven that polyphenolic compounds possess significant hypoglycaemic properties. They act via different modes of actions such as inhibition of carbohydrate digestion, insulin stimulation from pancreatic beta cells, activation of insulin receptors, intestinal glucose absorption, modulation of glucose release from the liver, and modulation of intracellular signalling pathways and gene expression [
5]. The phenols and flavonoids further inhibit the key enzymes in carbohydrate metabolism such as pancreatic alpha-amylase and intestinal alpha-glucosidase [
6]. HPLC analysis of methanolic extract of
Synsepalum dulcificum showed that it contained rutin, quercitrin, isoquercitrin, kaempferol, and few other secondary metabolites [
7]. The flavonoid-rich extract showed antidiabetic potential in type 2 diabetic rats [
8].
Tadera et al. studied the inhibitory effect of few flavonoids against yeast and rat small intestinal alpha-glucosidase and porcine pancreatic alpha-amylase. This study showed that the flavonoids myricetin, quercetin, and luteolin were potent porcine pancreatic alpha-amylase inhibitors, with IC
50 value less than 500 µM [
9].
In recent years, in silico methodologies have become an important aspect of the drug discovery process due to their time-saving and cost-effective nature. These computer-aided drug design (CADD) approaches are advantageous for reducing the use of animals for in vivo experiments. Many in silico studies have been proposed to prove the potency of natural products to inhibit the alpha-amylase and alpha-glucosidase enzymes.
Piparo et al. reported that the structural arrangement of the flavonoids is very important to inhibit the alpha-amylase enzyme. This computational ligand docking study suggested that the inhibitory potentiality of flavonoids depends on the conjugated π-system formation which stabilises the interaction with active pocket and hydrogen bonds present in between the catalytic residues of binding sites and hydroxyl groups of polyphenol ligand. They reported that the flavonols such as myricetin, kaempferol, rhamnetin, isorhamnetin, quercetin, quercetagetin, and fisetin showed higher docking scores, suggesting the strong binding and inhibition of salivary alpha-amylase. While for flavanols, flavanones, and isoflavones, docking scores were the lowest, suggesting their inability to inhibit salivary alpha-amylase [
10].
Argyreia nervosa (Burm.f.) Bojer is a well-known medicinal plant which has many therapeutic properties such as anti-inflammatory, antimicrobial, antiviral, antiulcer, hepatoprotective, analgesic, immunomodulatory, and antihyperglycemic [
11]. The silver nanoparticles, synthesised using the aqueous leaf extract of
A. nervosa, exhibited the alpha-amylase and alpha-glucosidase inhibitory activity with increasing concentration [
12]. The ethanolic root extract of
A. nervosa proved effective in controlling hyperglycaemia in normal and diabetic rats [
13]. Latha et al. demonstrated that the dose-dependent reduction in blood glucose level in normal and alloxan-induced diabetic rats was achieved when treated with alcoholic extract of
A. nervosa [
14]. Although this plant has been screened for its secondary metabolite content, very few studies have reported the hypoglycaemic activity and none have studied the mechanism of action of any of these metabolites.
The current study was carried out to identify and validate the alpha-amylase inhibitory flavonoids from Argyreia nervosa (Burm.f.) Bojer. The identification of the flavonoids from chloroform leaf extract was performed by using the LC-MS technique and the inhibitory activity was demonstrated using preliminary in vitro starch iodide assay. Further, the results were validated by the molecular docking studies, and highly promising compounds were shortlisted.
2. Materials and Methods
2.1. Collection of Plant Material
The fresh and healthy leaves of Argyreia nervosa (Burm.f.) Bojer were collected from Savitribai Phule Pune University campus (Latitude 18.549120° N and Longitude 73.828120° E). The authentication of plant material was conducted by the Botanical Survey of India, Western Regional Centre, Pune-7. (BSI/WRC/IDEN.CER./2020/94)
2.2. Preparation of Plant Extracts
The freshly collected leaves were washed under running tap water, dried, and then chopped into small pieces. Using a muslin cloth, this sample was wrapped. The hot extraction, i.e., the Soxhlet extraction method, was used. Sequential extraction was carried out using 6 solvents with increasing polarity. The extracts were concentrated using a rotary evaporator and stored at room temperature. In total, 50 g of fresh leaves were used for extraction. The yield of pet ether, chloroform, ethyl acetate, acetone, methanol, and water extracts was 0.19%, 0.33%, 3.5%, 3.24%, 5.74%, and 5.90%, respectively.
2.3. Preliminary Phytochemical Analysis
The extracts obtained were subjected to preliminary phytochemical analysis using standard procedures stated by Trease and Evans (2002) to determine the presence of different phytochemicals [
15].
2.4. Preliminary Starch-Iodide Assay
The method reported by Ponnusamy et al. (2011) was used to perform alpha-amylase inhibition assay [
16]. An alpha-amylase enzyme from the porcine pancreas (PPA) (Sigma-Aldrich, USA, Catalogue No. A6255) was used for the assay based on colour reaction produced by the starch iodine complex (starch–iodide assay). The assay mixture consisted of phosphate buffer (pH 6.9 containing 6.7 mM sodium chloride), 0.02 units of PPA enzyme, and 20 μL of plant extract in DMSO (1 mg/mL
w/
v). The assay mixture was incubated at 37 °C for 15 min. Then, soluble starch (1%,
w/
v) was added to each assay mixture and reincubated at 37 °C for 15 min. 20 μL of 0.2 M Tris solution was used to stop the enzymatic reaction after the incubation period and finally, 20 μL of iodine reagent was added. The colour change of the reaction mixture was observed. The positive drug control (SC) was a synthetic, known PPA inhibitor, acarbose (Glucobay tablets, Bayer). A dark-blue colour indicated the presence of starch, implicating inhibition of amylase enzyme by the plant extract. A yellow colour indicated the absence of starch, implicating fully active amylase enzyme that degraded the starch and hence the absence of amylase inhibitory activity in the plant extract. A brownish colour indicated partially degraded starch in the reaction mixture.
2.5. Determination of Total Flavonoid Content
For the determination of total flavonoid content, aluminium chloride colorimetric method was followed, with slight modifications [
17]. The stock solution of 5 mg/mL of chloroform extract was used. The sample (0–200 µL) was mixed with 1.25 mL distilled water and 75 µL 5% NaNO
2 solution prior to the 5 min incubation period. Then, 150 µL of 10% AlCl
3 was added, and again the test solution was incubated for 6 min. Then, 500 µL of 1 M NaOH and 275 µL distilled water was added to the test solution. The absorbance was read at 510 nm using a UV-VIS spectrophotometer (UV-1800, SHIMADZU, Kyoto, Japan). The total flavonoid content was calculated using a calibration curve. The results were expressed as quercetin (Sigma, St. Louis, MO, USA) equivalent in µg/mg of sample.
2.6. Column Chromatography
The column chromatography was performed to separate the compounds from the chloroform leaf extracts showing promising alpha-amylase inhibitory activity in the preliminary starch iodide assay. For this procedure, 5 mg of concentrated extract was dissolved in 1 mL of chloroform and passed through the 20 cm long silica gel (60–120 mesh size, SDFCL, Mumbai, India) column with 20 mL chloroform, methanol, and water each, successively. The fractions were collected and again checked for PPA inhibitory activity. The fraction showing positive results was carried forward for the LC-MS analysis.
2.7. LC-MS Analysis
The MS acquisitions were performed in both positive and negative electrospray ionisation mode. The capillary voltage, cone voltage, and fragmentor voltage were 4 kV, 45 V, and 170 V, respectively. The gas temperature was set at 325 °C. Data were acquired at a scan rate of 3 Hz in the mass range of 100–100 m/z. Data were further analysed with Mass Hunter qualitative software and the METLIN database.
The system used was Agilent 1260 binary LC System and 6540 Ultra-High-Definition Accurate Mass QTOF LC/MS system for liquid chromatography and mass spectrometry, respectively. A 1 µL aliquot of sample solution was injected into Agilent Zorbax Extend C18 RRHT column (50 × 2.1 mm, 1.8 µm) with a flow rate of 0.3 mL/min and run time of 30 min. The mobile phase consisted of H2O- 0.1% formic acid (A) and acetonitrile (B), and the gradient elution procedures were performed as follows: 0–5 min (95% A: 5% B), 5–18 min (5% A: 95% B), 18–27 min (5% A: 95% B), 27–27.10 min (95% A: 5% B) and 27.10–30 min (95% A: 5% B).
2.8. Molecular Docking Studies
2.8.1. Softwares and Tools Used
Protein Data Bank (PDB), PubChem, Open Babel, Discovery Studio Visualiser, AutoDock Version 4.2.6, MGL Tools, CastP.
2.8.2. Ligand Preparation
2.8.3. Protein Preparation
The crystal structure of PPA (PDB ID: 1OSE) was retrieved from Protein Data Bank (
https://www.rcsb.org/, accessed on 9 September 2020) in PDB format [
18]. This retrieved structure was complexed with water molecules, other atoms, and ligands which increased its resolution. The protein was prepared using Discovery Studio software where water molecules, other heteroatoms, and previously co-crystallised ligands were removed so as to free up the active sites on protein molecules. Using AutoDock 4.2.6, the polar hydrogen atoms and Kollman charges were added to the protein to achieve a well-protonated state. This prepared protein structure was saved as PDBQT format file prior to docking [
19].
2.8.4. Binding Pocket Identification
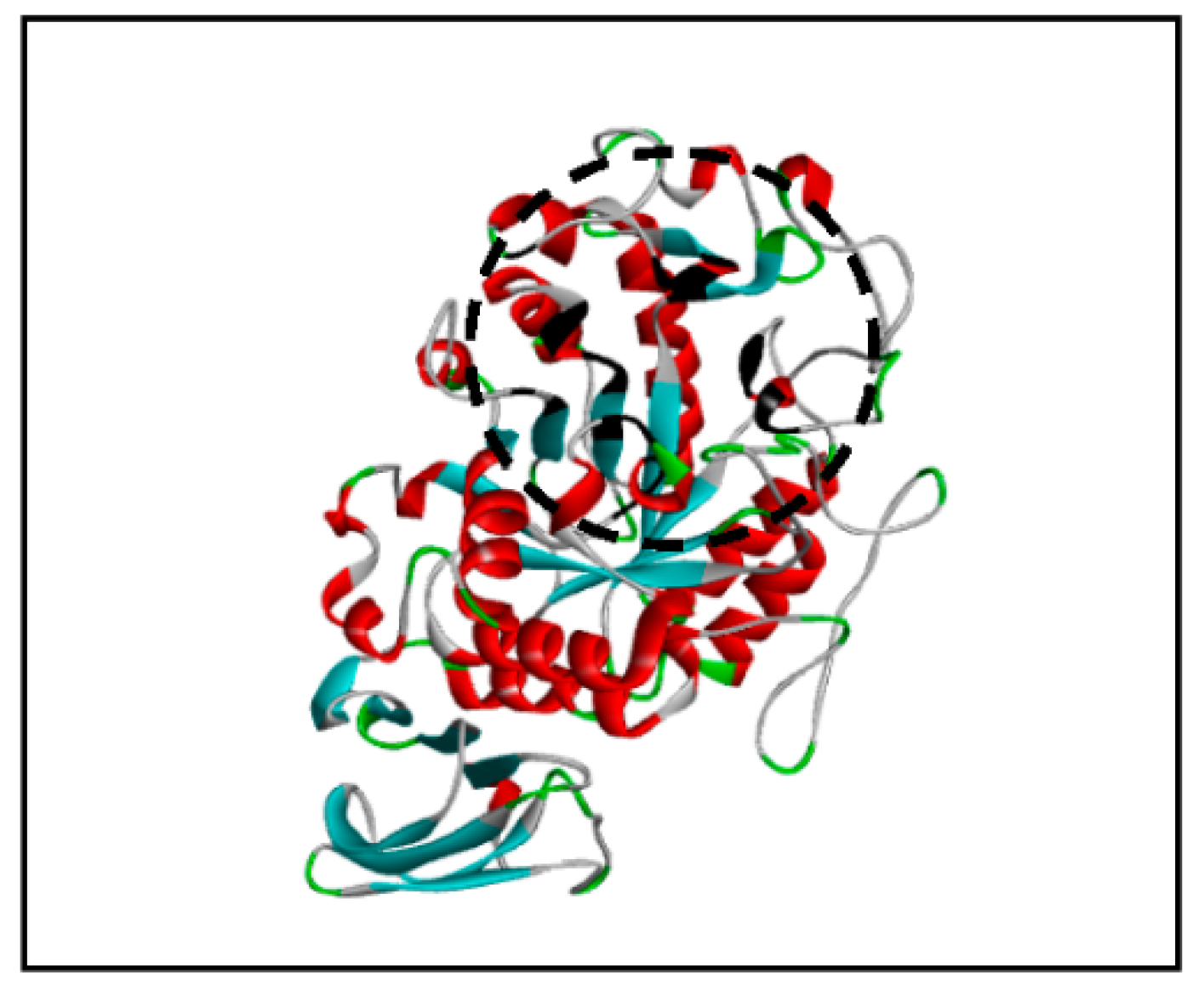
The binding pocket is a cavity on the surface or in the interior of a protein where the ligand molecule can fit while interacting with the protein. The online software CASTp (
http://sts.bioe.uic.edu/castp/, accessed on 13 July 2020) was used to identify the binding pocket of PPA. [
20].
2.8.5. Docking Studies Using AutoDock
The prepared ligands were docked on the receptor PPA protein (both in PDBQT file format) using AutoDock 4.2.6. A grid box with spacing 0.375 A° centred at (72 A° × 60 A° × 65 A°) was set using the knowledge of active sites present on the receptor protein. For docking, the receptor protein was set as a rigid molecule and the ligand was set as a flexible molecule.
2.8.6. Protein–Ligand Interaction
The software Discovery Studio Visualiser (
https://discover.3ds.com/discovery-studio-visualiser-download, accessed on 9 September 2020) was used to visualise the protein–ligand interactions [
19] in 2D and 3D formats (3D Data not shown). The PDB file containing the docked conformation was investigated for the presence of intermolecular interactions and their strengths which include hydrogen bonds and Van der Waal interactions.
2.8.7. Lipinski Rule for Ligands
The selected ligand molecules for docking studies were screened for Lipinski’s rule of five [
21]. This rule states that the drug molecule should not violate more than one among the five rules enlisted below:
(1) Molecular mass <500 Da, (2) hydrogen bond donors <5, (3) hydrogen bond acceptors <10, (4) high lipophilicity (Log p < 5), and (5) molar refractivity between 40 and 130.
The Supercomputing Facility for Bioinformatics and Computational Biology, IIT Delhi (
http://www.scfbio iitd.res.in/software/drugdesign/lipinski.jsp, accessed on 15 October 2020) was used to check the Lipinski’s rule for the selected metabolites where the PDB structures of the metabolites were uploaded to their online server [
22].
4. Discussion
Plants produce a variety of organic compounds as a result of their metabolic processes. These secondary metabolites play a key role in the plant’s own survival as they are involved in interactions with other plants, animals, and microbes. Flavonoids, a group of plant secondary metabolites, have been studied by researchers to a great extent as they possess useful properties such as anti-cholinesterase activity, anti-inflammatory activity, radical scavenging activity, etc. They also play a key role as phytoalexins, signal molecules, allopathic compounds, detoxifying agents, and antimicrobial defensive compounds. These flavonoids are also reported for their insecticidal and antimicrobial property as they interfere with nucleic acids and proteins. Therefore, they are important compounds in developing pesticides in the agriculture industry [
23].
In the present study, four compounds belonging to class flavonoid were studied for their porcine pancreatic alpha-amylase (PPA) inhibition potential. These were isolated from the PPA inhibitory chloroform extract of the leaf of A. nervosa. The phytochemical analysis of this extract showed the presence of phenols, flavonoids, terpenes, and quinones. Of those, flavonoids were present in abundance since the total flavonoid content was 378.83 ± 0.76 µg/mg sample.
Column chromatography of this chloroform extract was performed to dispose of the chlorophyll present and to separate metabolites. Among the fractions eluted, the fourth fraction eluted with chloroform exhibited significant PPA inhibitory activity. The LC-MS analysis of this fraction showed the presence of different secondary metabolites. Only a part of the data is presented here.
The four flavonoids, vitexin, rutin, myricetin, and isoquercetin were docked onto the PPA enzyme structure because they had shown better or comparable PPA inhibitory results to acarbose, the positive control used in our experiments. The mechanism of action of flavonoids suggests that the binding of these flavonoids depends on their structure and the bonding pattern with the receptor molecule. The conjugated π system formation is important for the stabilisation of the interaction with the binding pocket. In our study, it was observed that all four flavonoids formed π interactions with active site amino acid residues (
Figure 6). These π interactions were π–anion, π–alkyl, π–π T-shaped, π–π stacked, π–sigma, and π–donor hydrogen bond. When acarbose was studied for its π interactions, it was observed that no π interactions were formed by acarbose with a target protein. These results provided insight into the higher binding energy of flavonoids than acarbose. The π interactions formed between the flavonoid and target protein strengthened the binding and resulted in a significant docking score.
In molecular docking studies, it was observed that the amino acid residues ASP197, GLU233, and TRP59 were common for the four flavonoids from
A. nervosa, showing significant binding energies. In the literature, it is reported that the PPA binding pocket possesses these three amino acid residues as a part of the active site where the inhibitor or the ligand actually binds [
24]. Additionally, the other amino acid residues such as ASP300, HIS101, HIS299, and LYS200 are part of the active site of the receptor.
Similar to other glycosidases, the active site carboxylic acid is used to catalyse the hydrolytic reaction by alpha-amylase. The amino acid residues ASP197, GLU233, and ASP300 were described as the catalytic residues of the active site [
25]. It was suggested that the ASP197 acts as a nucleophile which attacks the sugar anomeric centre to form a covalently bound reaction intermediate. During this process, the substrate’s reducing end is cleaved off the sugar skeleton. Afterwards, to break the covalent bond formed between ASP197 and substrate, water molecules attack the anomeric centre. In both processes, GLU233 and ASP300 act as acid/base catalysts independently or cumulatively. The molecular docking results of the four studied flavonoids showed that the above-mentioned catalytic amino acid residues were involved in the hydrogen bond formation in between the ligand and the target protein which suggests that each of the ligands is inhibiting the protein efficiently.
When the results of four flavonoids were compared to the result of acarbose (
Table 1), they demonstrated that our ligand molecules formed strong interaction with catalytic residues ASP197 and GLU233 or ASP300 to inhibit the alpha-amylase enzyme. This also explained the strong PPA inhibition that we had observed in our biochemical assays. In the current study, all four flavonoids studied were interacting with these essential catalytic residues better than acarbose and these can be developed as potent alpha-amylase inhibitors. It will also be interesting to investigate whether these flavonoids show better PPA inhibition singly or synergistically.
The flavonoids were also checked for Lipinski’s rule of five. Myricetin and vitexin showed promising results as they violated just one rule of Lipinski. Though some parameters were not followed by the flavonoids studied, they can be considered as strong candidates in the drug designing process against hyperglycaemia.










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}