Abstract
Castor (Ricinus communis) is an oilseed usually cultivated in tropical and semi-arid conditions. The slow and uneven germination of castor seed is causing poor crop establishment and reduced seed yield. This series of studies investigated several factors influencing the time for castor seed germination and seedling emergence. Studies were made on the effect of seed coat permeability to water and the influences of temperature, morphology, and the hormone gibberellin. The best temperature for castor seed germination was near 31 °C, and the seed coat was permeable to water. The mechanical resistance of the seed coat was a mechanism controlling the germination of the castor seed. The time for germination was strongly associated with the seed coat relative weight, but it had little influence from other morphological characteristics. After the castor plants were subjected to three cycles of selection for fast germination, the seed weight was reduced, the caruncle became larger, and the seed coat became thinner. Gibberellin applied to the seed promoted a faster and higher germination rate. Pre-germination of the castor seed was not effective for promoting faster seedling emergence. Attempts to improve castor seed germination should integrate the many factors that influence this physiological process.
1. Introduction
Castor (Ricinus communis, Euphorbiaceae) is an industrial crop predominantly cultivated in tropical and semi-arid regions of the world [1]. Castor oil is valuable as a versatile raw material used as an ingredient in a long list of components such plastics, adhesives, lubricants, paints, waxes, and much more [2]. Castor crop is being increasingly adopted as an option for crop rotation in intensive cropping systems, such as in the high-input and fully mechanized soybean–corn–cotton system that predominates in Brazil [1].
The fast germination and emergence of castor seedlings is a requisite for adequate crop establishment and for increasing its seed yield. Nevertheless, the slow and uneven emergence that has been frequently observed is harming the seed yield [3,4]. The final seed yield is negatively impacted by uneven germination, because the seedlings that emerge a few days late are not able to compensate for this delayed initial growth, and they do not reach the full potential yield as the plants that emerge early. The occurrence of plants with varying ages is also a problem for weed management and for preparing the crop for mechanical harvest [5,6].
Another problem created by delayed germination is the persistence of castor seed in the following cropping seasons, which is a characteristic very common in weed species [7], but which should be absent in cultivated plants. Castor is frequently employed as an option for intercropping or rotation with food crops (soybean, corn, and beans). Many castor seeds fall in the ground because of natural dehiscence or in the harvest operation, and they may germinate in the subsequent season. If all volunteer castor plants emerged at the same time, they would be easily destroyed; however, their management becomes difficult, because they keep emerging in an extended period. It has been demonstrated that castor seeds may still be viable after two years in the soil, and they germinate after a soil disturbance [8].
Castor seed germination is a complex physiological process that is influenced by many factors, including its genetic traits, physiological condition, and the environmental conditions [4,6,9,10,11,12,13]. Many attempts have been made aiming to improve castor seed germination, such as caruncle removal, soaking in water, scarification, treatment with gibberellin, the selection of varieties with fast germination, harvesting by raceme order, and sorting the seeds according to size and density [3,4,14,15,16].
This article reports a series of studies evaluating factors that were previously observed to influence the seed germination of castor and other plant species. The objective was to test the following hypotheses: (1) that castor seed germination is delayed because the castor seed coat is impermeable to water [17,18,19], (2) that the time for germination is influenced by temperature [20,21,22], (3) that the mechanical resistance of the seed coat is a mechanism to delay germination [23,24,25,26,27,28,29,30], (4) that the germination ability is influenced by the seed weight, volume, density, and seed coat thickness [7,15,16], (5) that castor seed germination is influenced by the hormone gibberellin [4,29], (6) that selection for fast germination changes the morphological characteristics of the seed in its descendants [7,31,32], and (7) that the time for germination can be reduced with a pre-germination treatment [3,17].
2. Material and Methods
2.1. Imbibition and Germination Influenced by Temperature and a Puncture on the Seed Coat
The hypothesis tested in this experiment was that the seed coat is impermeable to water, and a puncture on the seed coat will allow for water uptake without resistance. Castor seeds were incubated to germinate at temperatures of 16, 19, 22, 25, 28, 31, and 34 °C. For each temperature, 70 castor seeds of the cultivar AKB 02 had the caruncle manually removed, and a small puncture was made in 35 seeds. The experiment was run in July/2021, with the castor seeds harvested nine months before and provided by Terasol. The puncture was approximately 1.5 mm wide and was made pressing the tip of a screw in the region of the seed coat where the caruncle was removed. Each seed was individually weighed (at a natural moisture content) and placed in plastic boxes (11 × 11 cm) on top of two sheets of germination paper (230 g/m2), in an assigned position that allowed for its successive weighing. The germinating paper had creases to avoid the seed moving from its position, and each box had only 10 seeds. The seeds with and without puncture were distributed randomly. The seed was always placed with the hilum (the region beneath the caruncle) facing down. Demineralized water was added to the germination paper in the proportion of 2.5 times its dry weight. The boxes were placed in germinating chambers at a constant temperature (according to the treatment) in continuous dark.
The individual seed weight was repeatedly measured after water was added. The first weighing was made between 8 and 18 h after initial hydration, and then at approximately 24 h intervals. The time of each weighing was registered. Before weighing, the seed was gently dried with a paper towel to remove the film of water that covering the seed coat. After weighing, the seed was returned to the same position in the plastic box. Additional water was added to the germination paper, always so that the seeds were not covered with a film of water. The seed was assumed as having germinated when the seed coat cracked and the radicle was visible. The seed was discarded after registering the time for germination and the radicle length.
The data on successive weighing were used to plot the curves of seed hydration. The weight measured on the day before the radicle protrusion occurred was not plotted in the curves, because the seeds were assumed to have entered Phase 3, in which intensive water absorption occurs before the radicle protrusion [29,33]. Vigor and malformed seeds were not registered as experimental variables. The experiment on each temperature was terminated when no seed germinated for 2 days or at 9 days after hydration.
The time for germination was corrected considering the radicle length when the observation was made, as previously described [4]. In short, the elongation rate of the radicle was estimated in a large number of seeds. Then, the precise time of the radicle protrusion was estimated considering the time elapsed after the radicle protrusion subtracted from the time of the day that the germination was observed.
Analyses of imbibition were made using the relative seed weight, which was calculated by dividing the weight of the hydrated seed by its own weight before imbibition. Aborted seeds were excluded from the analysis of imbibition, because they imbibe in a different pattern and skew the sample’s distribution. The seed was assumed as aborted when it was apparently normal at the external inspection, but it was in fact empty and lacking internal reserves. This condition could be confirmed when the seed subjected to hydration met the three following conditions: (i) it did not germinate, (ii) it reached a relative seed weight greater than the threshold of 1.8, and (iii) its weight continued to increase at and after 48 h of imbibition [34]. The data on the relative seed weight as a function of time after hydration were not analyzed by linear regression, because these data did not meet the assumption of constant variance. For that reason, the relative seed weight (Y) was just plotted against the time after hydration (X), and is presented as means and standard deviations for each temperature and separately for seeds with and without the puncture on their seed coat. When some comparisons were needed, the means were compared by t-test with a significance threshold of p < 0.05. The effects of temperature and puncture on the hydration in the first 24 h of imbibition were analyzed, with the interpolation of the weight of each single seed that was measured immediately previous to and after 24 h. The estimated values for each temperature were compared between seeds with and without puncture by t test (p < 0.05) and subjected to a linear regression analysis as a function of temperature (p < 0.05). When the data on seed weight did not meet the assumption of normal distribution by the test of Shapiro–Wilk, the comparison was made with the non-parametric Mann–Whitney Rank Sum test (p < 0.05). The analyses were made in the software SigmaPlot v. 12.3.
2.2. Visual Analysis of the Rupture on the Seed Coat
Pictures were taken on germinating castor seeds, from the same bag used for the experiment described in the Section 2.1, at different phases of germination, in order to observe how the seed coat broke before the radicle protrusion. Seeds with and without a puncture on the seed coat were incubated for 48 h at 30 °C. Seeds with radicle protrusion were observed under a microscope and pictures were taken for the illustration of relevant details on the puncture, seed coat cracking, and radicle protrusion. The pictures were taken using a Stemi 2000-C stereo microscope (Zeiss, Oberkochen, Germany) and an AxioCam ICc 5 camera (Zeiss, Oberkochen, Germany).
2.3. Morphological Characteristics Influencing the Time for Germination
A sample with 200 castor seeds of the cultivar BRS Energia was obtained from a commercial lot. The seed was harvested by Embrapa in October/2020, and the experiment was run in April/2021. The caruncle was manually removed, and the seeds were disinfected with sodium hypochlorite (NaClO) at 1% for 3 min, rinsed three times with deionized water, and allowed to dry for 24 h at room temperature. The weight, length, width, and height were measured in each seed, and they were incubated in a germination chamber in the dark at a constant temperature of 22 °C. The other procedures were as described in Section 2.1. The seeds were inspected daily for 8 days of incubation, and they were assumed as having germinated when the radicle was visible. The time for the germination of each individual seed was associated with its morphological characteristics. Just after the seed germinated or after the incubation (for ungerminated seeds), the seed coat was removed, oven dried (70 °C for 24 h), and weighed. Six seeds were discarded because of heavy fungal infection. The seed coat was not removed and weighed in 10 germinated seeds because they were used for another experiment [4].
The time for germination was corrected by the radicle length when the reading was made, as explained in Section 2.1. The area (A) and volume (V) of each seed was calculated with the equations:
where L is the length, W is the width, and H is the height. The formula for the area of ellipsoid was an approximation proposed by Klamkin [35]. The volumetric density was calculated as the ratio between the weight and volume, and it is expressed as g/cm−3. The seed coat thickness was calculated as the seed coat weight divided by the seed area (A), and it is expressed as g/cm−2. The relative weight of the seed coat was calculated as the weight of the seed coat divided by the seed weight, and it is expressed as a percentage.
A = 4π((L1.61 × W1.61+ L1.61 × H1.61+ W1.61 × H1.61)÷3)0.62, and
V = π × L × W × H ÷ 6
The influence of each morphological characteristic (x) on the time for germination (y) was analyzed with simple linear regression (p < 0.1). The influence of the five morphological characteristics combined on the time for germination was analyzed with multiple linear regression (p < 0.1). When the regression analysis included seed coat thickness or seed coat relative weight, the sample size was smaller because of the ten seeds that were used for another experiment, in which the seed coat was not removed and weighed. The results are presented in dispersion graphs, and the regression line was omitted when it was not significant. The mean of each morphological characteristic was compared between germinated and ungerminated seeds using a t test (p < 0.05). This analysis did not include the ungerminated seeds discarded due to fungal infection, aborted seeds, and the seeds selected for fast germination [4].
2.4. Selection for Fast Germination Modifying Castor Seed Morphology
The morphological characteristics were measured on castor seeds of the cultivar BRS Energia that were subjected to three cycles of single-seed selection for fast germination [4]. This selection consisted in the germination of 200 seeds on moisture paper, selecting the first 10 germinated seeds, which were transplanted to the field to grow and produce seeds. Samples were taken from the original seed (10 seeds S0) and in each generation of selection (100 seeds of S1 to S3), and data were taken on the length, width, height, and weight of the seed, caruncle, and seed coat. The values of the relative weight of the caruncle and seed coat, area, volume, seed coat density, and volumetric density were calculated using the same procedures described in Section 2.3. As the data failed to meet the assumptions of normal distribution and equal variance, they were subjected to a Kruskal–Wallis Analysis of Variance on Ranks and the medians were separated by Dunn’s test (p < 0.05). The analysis was made in the software SigmaPlot v. 12.3.
2.5. Gibberellin Influencing Castor Seed Germination
Castor seeds of the cultivar Mia were provided by Kaiima Seeds in May/2022, and the experiment was performed in October/2022. The seeds were incubated for germination in plastic boxes, as described in Section 2.1, except for the incubation temperature of 30 °C, duration of 9 days, and varying doses of gibberellin. Solutions were prepared using the commercial product ProGibb 400® (Sumitomo, Chiyoda, Japan) from 0 to 133 mg/L with increments of 13.3 (11 treatments). Fifty seeds were placed in each box, and the paper was moisturized with the solutions. Germinated seeds (radicle protrusion) were counted daily and then discarded. The data on the final germination rate were plotted as a function of the gibberellin doses and subjected to a regression analysis (p < 0.1) with the hyperbola model y = ax/(b + x) + c (p < 0.1). The progress of germination was plotted with three lines: control, low doses (from 13.3 to 66.7 mg/L), and high doses (from 80.0 to 133.0 mg/L).
2.6. Emergence of Pre-Germinated Castor Seed
Castor seeds of the cultivar AKB 05 were provided by Terasol in October/2021, and the experiment was performed in April/2022. The seeds were pre-germinated on trays, on the top of a germination paper moisturized with tap water, and incubated for 0, 12, 24, 36, 48, and 60 hours at a constant temperature of 30 °C, in the dark. Each incubation started at a different time in order to end the pre-germination treatments concomitantly. Just after the incubation, 50 pre-germinated seeds from each treatment were sowed 4 cm deep in plastic trays filled with a substrate of soil + manure (1:1 v:v), inside a greenhouse, under daily sprinkler irrigation. The 300 pre-germinated seeds were randomly distributed in 6 trays, and the position of each seed in the tray was registered in order to associate the emerged seedlings with its treatment. The trays were inspected every day to register the time for the emergence of each seedling. At 14 days after sowing, the shoot of the emerged plants was cut at the substrate level, placed on individual paper bags, oven dried for two days at 105 °C, and weighed.
The progress of emergence (%) was plotted as a function of the time after sowing. The data of the shoot dry weight from all the incubation times were pooled, plotted as a function of the time after emergence, and subjected to a regression analysis (p < 0.1) with the exponential model.
3. Results
3.1. Temperature and Seed Coat Puncture Influencing Seed Hydration
There was no evidence that the castor seed coat was impermeable to water at any temperature. After 24 h of hydration, the relative seed weight of the seeds with or without a puncture was equal at all temperatures (p > 0.05), except at 22 °C (Mann–Whitney test, p = 0.046) (Figure 1). However, water absorption occurred faster under a warmer temperature for the seeds that were both without puncture (y = 1.14 + 0.0032x, R2 = 0.131, p < 0.001) and with puncture (y = 1.13 + 0.0042x, R2 = 0.205, p < 0.001) (Figure 1).
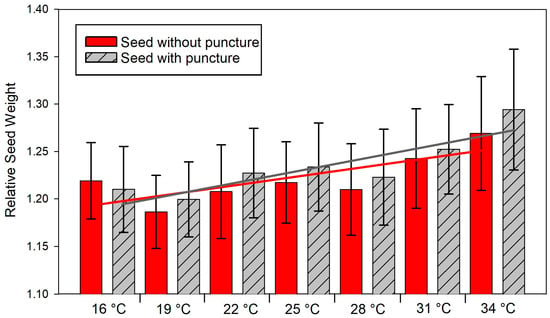
Figure 1.
Relative seed weight at 24 h after hydration of castor seed with an puncture (gray line and columns) and without a puncture (red line and columns) in the seed coat, imbibing at temperatures from 16 to 34 °C. The vertical bars are the standard deviation.
The progress of hydration followed the expected Phase 1 with fast water absorption and Phase 2, in which the weight was stable (Figure 2). The curves of seed hydration at 19, 22, and 25 °C apparently indicate that the relative seed weight of the punctured seeds was greater than that of intact seeds; however, that small difference was not significant, and it occurred because the punctured seeds were slightly ahead in the germination process (see the Section 3.2). The punctured seeds had a small weight increase because they were initiating Phase 3 of seed germination. In the transition from Phase 1 to Phase 2, which occurred between 40 and 47 h after hydration, the relative seed weight was statistically equal among the seeds germinating from 16 to 31 °C.
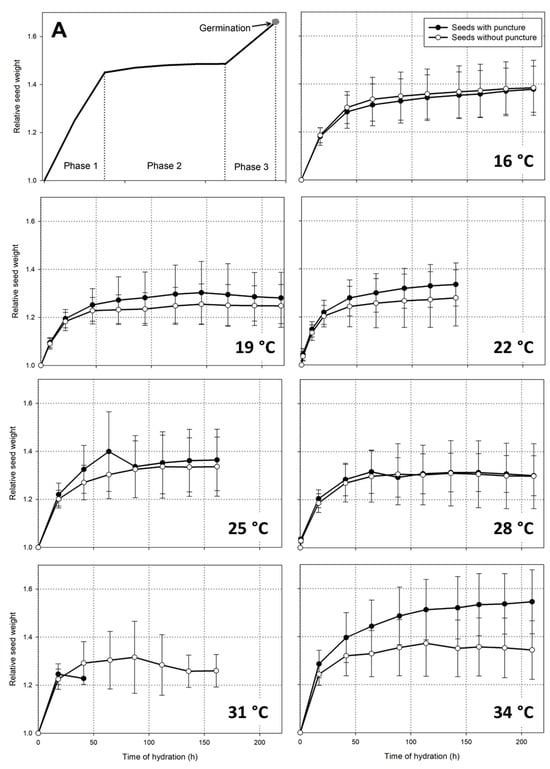
Figure 2.
(A) Phases of seed hydration [33]; hydration progress of castor seed of the cultivar AKB 02, with (closed circles) or without (open circles) a puncture on the seed coat, at temperatures varying from 16 to 34 °C.
Exceptionally, at 34 °C, the relative weight of the seeds with a puncture was significantly higher than that of intact seeds. In the measurement made at 210 h after hydration (Figure 2), the mean relative seed weight was 1.545 (n = 10) in the punctured seeds and 1.344 (n = 16) in the intact seeds (t = 3.966, p < 0.001). The physiological reasons for that effect at 34 °C were not further investigated. It was just observed that the progress of hydration in the punctured seeds was atypical, because many seeds did not enter Phase 2 (expected continuous weight), and remained absorbing water without progressing to radicle protrusion.
The typical Phase 3 of seed germination [29,33] was observed in the castor seed. On average, the relative seed weight was 1.295 ± 0.133 in the day prior to germination, and it increased to 1.412 ± 0.162 on the day that the radicle protrusion was observed. There were apparent events of reduction in the relative seed weight in the hydration curves of the seeds germinating at 25, 31, and 34 °C (Figure 2). This effect was not related to seed dehydration, but because the seeds with the highest moisture content germinated and were excluded from the sample, and consequently, the remaining seeds were lighter.
3.2. Temperature and Seed Coat Puncture Influencing the Time for Germination
The puncture on the seed coat promoted a faster and higher rate of castor seed germination at any temperature (Figure 3), except at 16 °C, at which no seed germinated after nine days of incubation. The fastest and highest germination rate was observed at 31 °C, in which 94% of the seeds with a puncture germinated at 76 h after hydration, while only 40% of the intact seeds germinated at the same time. It was confirmed that castor seeds can remain fully hydrated for many days (in Phase 2) without germinating.
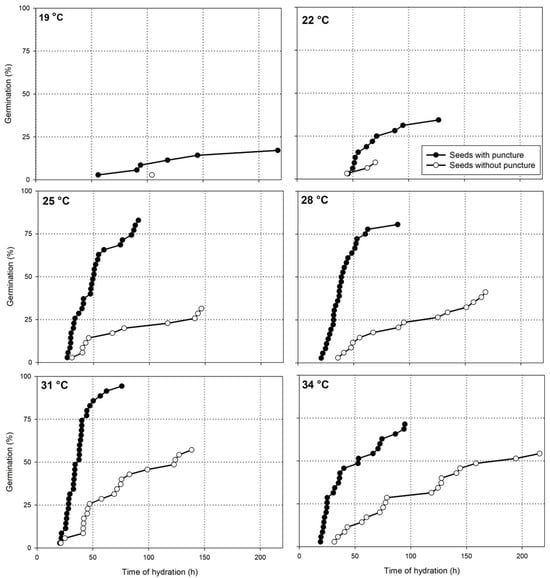
Figure 3.
Evolution of the castor seed germination, with (closed circles) or without (open circles) a puncture on the seed coat, incubated at temperatures varying from 19 to 34 °C.
3.3. Seed Coat Restricting Embryo Growth
It was observed in the germinating castor seeds that the rupture of the seed coat for the radicle protrusion of an intact castor seed always occurred in a specific region that was centered in the hilum and expanded, opening two lines on the dorsal side of the seed (Figure 4D) and one line on the ventral side (Figure 4F). However, the puncture on the seed coat altered this pattern, causing the crack to have irregular borders that expanded in random directions (Figure 4C,E). It is noteworthy that the crack in the seed coat always passed by the puncture. This pattern of cracking suggests that the puncture made on the seed coat created a fissure or a weak point that reduced the mechanical resistance to the embryo expansion, facilitated the radicle protrusion, and made the germination occur faster.
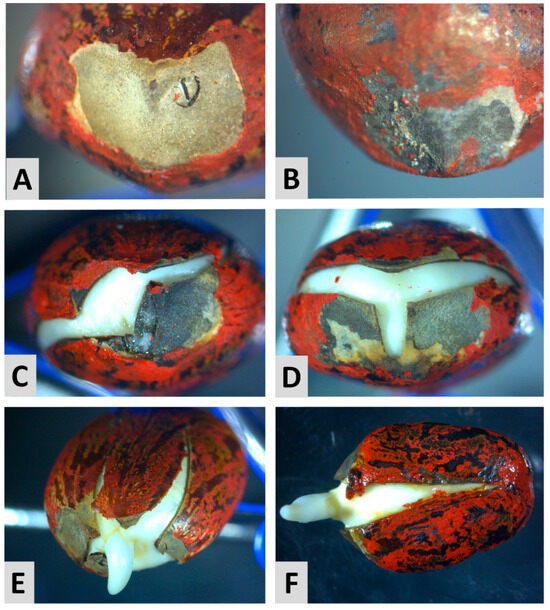
Figure 4.
Germinating castor seeds of the cultivar AKB 02 with (left) or without (right) a puncture on the seed coat. (A) A dry seed with the caruncle removed and a puncture on the hilum; (B) an intact dry castor seed with the caruncle removed; (C) prior to the radicle protrusion, the seed coat cracks with irregular shape with the fissure passing through the puncture; (D) without the puncture, the seed coat cracks with a regular shape always forming a single fissure on the ventral side and a double fissure on the dorsal side; (E) radicle protrusion through the crack on the seed coat with irregular shape; and (F) radicle protrusion and the linear fissure in the seed’s ventral side.
3.4. Time for Germination According to the Seed Morphology
The germination rate of the castor seed of the cultivar BRS Energia was 52% after 8 days of incubation at 22 °C, and the time for the germination of individual seeds varied from 64 to 184 h after hydration. The weight of the germinated seeds varied from 175 to 553 mg, but this characteristic was not significantly associated (p = 0.506) with the time for germination (Figure 5). The time for germination was significantly associated with the seed volume (p = 0.0191), seed volumetric density (p = 0.0014), seed coat thickness (p = 0.0656), and seed coat relative weight (p = 0.0002). Faster germination was observed in the castor seeds that had a smaller volume, higher volumetric density, and thinner seed coat.
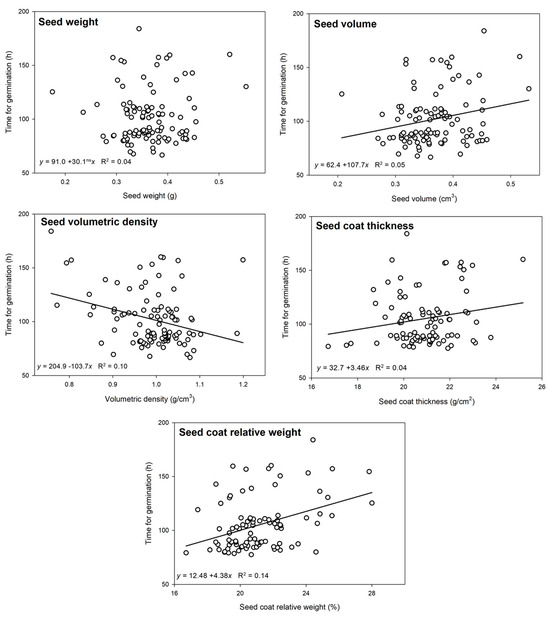
Figure 5.
Time for germination of castor seeds of the cultivar BRS Energia as a function of the seed weight, volume, volumetric density, seed coat thickness, and seed coat relative weight. The regression line was omitted when it was not significant at p < 0.1.
Comparing the seeds that succeeded and those that failed to germinate, it was observed that the germinated seeds were significantly heavier, had a higher volumetric density, had a thicker seed coat, and had a smaller relative seed coat weight (Table 1). Contrasting the time for germination (Figure 5) with the probability to germinate (Table 1), it was found that heavier castor seeds were more likely to germinate, but they took longer to germinate. The seed volume did not influence the chance to germinate, but a higher volume was associated with delayed germination. A higher volumetric density improved the chance and reduced the time to germinate. A thicker seed coat was associated with an increased germination rate but with delayed germination. A lower seed coat relative weight was associated with an increased germination rate and faster germination.

Table 1.
Comparison of morphological characteristics between germinated and ungerminated castor seeds 1.
When the five morphological characteristics were analyzed together, it was found that the relative seed coat weight was the only variable with a significant association (p = 0.0003) with the time for germination (Table 2). This is a valuable observation, because although each isolated morphological characteristic had some influence on the time for germination, the relative seed coat weight explained most of the variability, while the other traits had only indirect influence.
Table 2.
Multiple linear regression analysis 1 of the time for germination (y) in function of five morphological traits (x) of castor seed.
3.5. Selection for Fast Germination Influencing Morphological Characteristics
The selection for fast germination promoted significant changes in the morphology of the seeds. Compared to the original seed (S0), the castor seeds became lighter, the caruncle was heavier, and the weight of the seed coat increased in relation to the seed weight (Table 3). The seed produced by the castor plants selected for fast germination had a smaller area and volume, and a reduced seed coat density and volumetric density. Contrasting these results with the observations presented in Section 3.4, it was confirmed that the seed coat played a pivotal role regarding the time for germination; however, the seed coat relative weight increased in the three generations of selected plants, while the most relevant reduction was observed in the seed coat density. Looking into details and comparing the average of the original seed (S0) with the S1–S3 seeds, it happened that the reduction in the seed coat weight (13.5%) was smaller than the reduction in the whole seed weight (25.8%), resulting in an increased seed coat relative weight.
Table 3.
Medians of morphological traits in the original castor seed of the cultivar BRS Energia (S0) and on the three generation of single-seed selection for fast germination (S1 to S3) 1.
3.6. Gibberellin Influencing Castor Seed Germination
The hormone gibberellin influenced the rate and the time for germination of the castor seed (Figure 6). The germination rate increased until the dose of approximately 30 mg/L, and higher doses did not promote relevant improvements in the germination rate. The hormone also promoted faster germination in comparison to the untreated seeds. At 9 days of incubation, the germination rate of the seeds exposed to gibberellin was above 60%, while the germination of untreated seeds was just 32%.
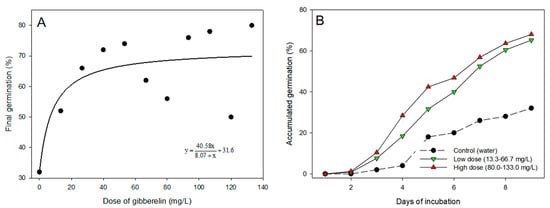
Figure 6.
(A) Germination of castor seed of the cultivar Mia after 9 days of incubation according to the dose of gibberellin; (B) progress of germination of seed in water (control), low doses (average of 13.33 to 66.7 mg/L, green triangles), and high doses (average of 80.0 to 133.0 mg/L, red triangles) of gibberellin.
3.7. Emergence of Pre-Germinated Castor Seed and Shoot Biomass Initial Growth
Pre-germination of the castor seed for 60 h favored early emergence of the seedlings. At 4 days after sowing, the emergence of the seeds incubated for 60 and 48 h reached 40% and 14%, respectively, while no seedling emerged from the control or shorter incubation times (Figure 7A). Nevertheless, at 6 days after sowing, the seedlings from the control treatment emerged at an equivalent or higher rate than the seeds from the pre-germination treatments. Overall, the pre-germination treatment caused delayed seedling emergence. At 14 days after sowing, the emergence rate of all seedlings reached the range of 72 to 86%.
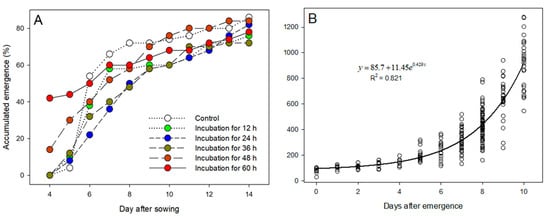
Figure 7.
(A) Progress of emergence of pre-germinated castor seedlings according to the incubation time; (B) evolution of the shoot dry weight of castor seedlings up to 10 days after emergence.
The shoot biomass grew exponentially in the 10 days after emergence (Figure 7B). On the day of emergence, the mean castor seedling shoot dry weight was 97 mg. If a seedling emerged late, the neighboring castor plants that emerged 10 days earlier would be far ahead in the biomass accumulation, with an estimated weight of 923 mg. The shoot dry weights from all the treatments were pooled, because the pre-germination treatments did not influence the initial growth pattern of the seedlings.
4. Discussion
4.1. Castor Seed Coat Is Permeable to Water
The hypothesis that castor seed coat is impermeable to water was investigated, because this condition is commonly observed in many species, and it was previously suggested as a reason for delayed castor seed germination [3,17]. It is estimated that about 15 families of the Clade Angiosperm have developed an impermeable seed coat (also called hard seeds) as a mechanism to regulate seed germination. For instance, it was found that some soybean seeds (Glycine max) are impermeable to water due to a strong reduction in the hydraulic conductivity of the seed coat [18,36]. The hydration of Uruchloa sp. seeds occurred faster and reached a higher level when the seed was punctured [26]. Impermeability to water was observed in seeds of Tilia miqueliana as a mechanism of physical seed dormancy [30]. However, the hypothesis that the germination of castor seed is delayed because of impermeability of the seed coat was refused. The puncture on the seed coat did not favor faster water absorption, as would occur if the seed coat was a strong barrier to water flux into the seed. There was some variability among the castor seeds in terms of water absorption, but no castor seed was found impermeable, as reported for soybean [18,36].
Temperature did not influence the time for hydration as well, and at any temperature, it followed the same pattern as previously reported for castor seed [13]. Imbibition is a physical process driven by the hydraulic conductance of the seed coat and the difference in water potential between the soil and the internal parts of the seed. As a physical process, imbibition is driven by the laws of diffusion and hydraulic flow, with a limited dependence on temperature, while it is more frequently influenced by characteristics such as seed size, density, and sphericity [23,33,37]. Only to a small extent, temperature can directly influence the permeability of the seed coat and the viscosity of the water [33]. This study confirmed that the hydration of castor seed occurred slightly faster under a warmer temperature (Figure 1); however, this effect was not related to the seed coat permeability, but because under a favorable temperature, more seeds reached Phase 3 of germination, in which the seed absorbed water for embryo growth. For instance, at 24 h after hydration, at least 74% of the punctured seeds exposed to 31 °C were in Phase 3 of germination compared with the temperatures of 16 or 19 °C, in which no seed entered that phase.
The relative seed weight was significantly higher only at the supra-optimal temperature (34 °C), because other physiological mechanisms were activated. A study with the hydration of castor seeds incubated at 35 °C with the seed coat removed also found increased hydration at the supra-optimal temperature, but not at 20 or 25 °C [11]. The same study also found that, among other amino acids and metabolites, the level of ascorbate increased by 30 times in seeds exposed to the supra-optimal temperature. Ascorbate is an antioxidant, and the increased hydration of castor seed may be caused by metabolites accumulating in the endosperm and embryo for osmotic adjustment in response to heat stress, aggravated in some way by the opening in the seed coat.
4.2. Temperature Plays a Pivotal Role in Castor Seed Germination
Temperature is a well-known factor controlling seed germination in many plant species [4,9,11,22,32], and it was confirmed in castor, which is a tropical plant adapted to temperatures in the warm range [1]. The optimal temperature for castor seed germination was found to be near 31 °C (Figure 3). It should be emphasized that most studies on castor seed germination follow the suggestion of standardized tests for quality purposes and incubate the seed at suboptimal temperatures—alternating 20/30 °C or in the range from 25 to 28 °C [3,8,13,14,15,17,38,39,40]. Although the procedures for quality assessment need to be standardized, experiments on castor seed germination searching for agronomical performance, vigor, or biological meaning could find better results if they adopt temperatures close to the optimal for this tropical plant species.
The uneven germination and emergence of castor seedlings may be related to the influence of soil temperature on seed physiology. Soil temperature oscillates in wide amplitude between night and day, and it is influenced by many factors such as sun radiation, soil reflectiveness, soil conductivity, soil moisture, and wind speed [41,42]. The continuous cover of the soil with crop residues is crucial for soil conservation, but this practice may reduce the soil temperature and impact the time for castor seedling emergence. Irrigation is frequently adopted on castor fields, and it can cool the soil to an extent that influences germination. As castor seed germination is very sensitive to temperature, the high variability in soil temperature under field conditions is potentially among the factors that make their emergence uneven.
There is genetic variability for germination under cold soil temperatures among genotypes of castor, as observed in other species such as rice (Oryza sativa), wheat (Triticum aestivum), and guar (Cyamopsis tetragonoloba) [4,9,43,44,45]. Just like that occurring in other crops, a strategy to be considered for improving the initial stand of castor fields is selecting varieties with the optimal germination and emergence under the soil temperatures of the target regions and planting time.
4.3. The Seed Coat Restricts the Germination of Castor Seed
The mechanical restriction to embryo growth is a common mechanism for delaying germination in many plant species. Embryo cells elongate inside the germinating seed before radicle protrusion and, together with the endosperm swelling, they make a pressure, forcing the seed coat to crack. In the opposite direction, the seed coat imposes a mechanical force to restrict embryo growth and block the radicle protrusion. As the embryo and endosperm increase the turgor, specific enzymes are produced in order to digest and weaken some specific seed coat cells to make it easier to crack [23,24,25,26,27,28,29,30].
The main hypothesis proposed to explain the fast germination of the punctured castor seeds was that the small hole on the seed coat created a fracture or a weak point that reduced the mechanical resistance that the seed coat may impose for delaying the radicle protrusion. The puncture on the castor seed coat was made on the region beneath the detached caruncle (Figure 4A). In an intact castor seed, that specific region becomes weaker during germination to allow for the opening for radicle expansion (Figure 4D). As it is also observed in other species, the layer where the seed coat cracks is a pre-determined breaking point, which is weakened by the digestion of the cell wall performed by enzymes [29,46]. In coffee seeds (Coffea arabica), the radicle protrusion was preceded by a build-up of internal pressure, increased activity of the enzymes cellulase and endo-β-mannanase, increased cell wall porosity of the restricting structure (endosperm in that case), and gradual reduction in the rupture force of the seed envelope [24]. In Arabidopsis thaliana and Lepidium sativum, the weakening of the covering layers was regulated by the balance between the hormones abscisic acid and gibberellin [27]. As the crack for radicle protrusion in the intact seeds consistently occurred in the same region of the seed (Figure 4D,F), it is likely that the digestion of some cells in the seed coat occurred in castor as well. It was observed in a previous study that removing the castor seed coat promoted the fastest germination compared to scarification, caruncle removal, and soaking in water [3]. Corroborating the observation that the weak point in castor seed coat is induced only when the seed begins the germination process, it was demonstrated that, when dry and non-germinating castor seeds were forced to break, the crack occurred in a completely different region of the seed coat, often on the side opposite to the hilum [47].
The role played by caruncle in castor seed germination is inconclusive [3,8,48], but the inconsistency among reports could be related to the mechanical resistance of the seed coat. The act of removing the caruncle for experimental purposes needs to be gentle to avoid creating invisible cracks that weaken the seed coat. Otherwise, germination would be influenced not by the absence or presence of the caruncle, but by the unintentional fracture hidden on the seed coat.
4.4. Morphological Traits Have Small Influence on the Time for Germination
This study confirmed the previous attempts reported in the literature that sorting castor seeds by weight, size, or density results in a higher germination rate but slower germination [15,16,49]. Nevertheless, sorting castor seeds according to their morphological characteristics has only a limited effect on the time for germination, and this practice alone would not be enough to obtain the high standard of emergence uniformity for adequate crop initial establishment.
Many studies on other plant species have demonstrated the weak association between seed size and the time for germination and seedling emergence [31,32]. Corn (Zea mays) producers blame the variability in seed size for uneven seedling emergence [50]; however, while the effect of seed size is occasionally observed in the laboratory, it is not confirmed in field conditions. Sometimes, the effect is observed in early development, but neither on the reproductive phase nor on the seed yield [50,51]. Associations between seed morphology and germination profile are sometimes found among different species, but not in comparisons of seeds from the same species [7].
Despite the significant associations that were observed in this study, the morphological characteristics of castor seed have a limited influence on the time for germination, because just a small part of the variability is explained by those measurable characteristics. For illustration, the difference in the time for germination between the first and the last castor seed to germinate in the study was 117 h, while the influence of the seed volume on the estimated time for germination was only 35 h and that of the seed density was 46 h (Figure 5). Additionally, the regression analyses had low coefficients of determination (R2 varying between 0.04 and 0.14), demonstrating that those variables explain just a small part of the variation in the time for germination.
The seed sample used in this study had a low number of very light seeds, because it was previously selected for commercial purposes, and the few remaining light seeds (2 out of 200) were assumed to be aborted and were discarded. However, it should be noted that very light seeds are clearly of low quality, and they can hardly germinate because they are aborted and have little or no reserves stored [34,38,52]. For that reason, the influence of the seed weight found in this study is only valid if aborted seeds are sorted out.
4.5. Selection for Fast Germination Altered Castor Seed Morphology
The relation between castor seed morphology and its germination traits is counterintuitive. While the seed morphology had a weak correlation with the time for germination when the seeds were just sampled from a bag (Figure 5), the selection for fast germination affected the seed morphology of the offspring (Table 3).
There is a well-documented trade-off between seed size and germination ability [32,53]. The resources available to grow seeds are limited, and the plant needs to choose between the strategies of producing few large seeds or producing many small seeds. On the one hand, a seedling that originates from a large seed has a higher chance to survive because of the larger amount of seed reserves. On the other hand, a higher quantity of seedlings increases the chance of more surviving individuals. In environments with abundant resources, plants tend to produce smaller seeds with fast germination [31]. This trade-off seems to operate in castor, in which the selection for fast germination triggered an evolutionary association with smaller seeds, because that configuration is likely to be better adapted for resource-abundant environments.
As discussed in Section 4.3, the caruncle does not play a direct role in castor seed germination; however, it was observed in this study that the selection of castor for fast germination also promoted a substantial increase in the caruncle weight in proportion to the whole seed (Table 3). The relation between caruncle size and fast germination may be explained by the evolutionary perspective. The function of the caruncle is related to seed dispersion. The caruncle attracts ants, which carry the seed to their nest for feeding on that nutritive structure [8]. Small seeds are easier to disperse [54]. Then, it seems plausible that the best strategy for fast germinating castor seed is favoring the dispersion by ants with small seeds with larger caruncles. As castor crop is intended for producing oil in the seed, the agronomical objective regarding the caruncle is opposite to the evolutionary drive for larger caruncles. This structure does not have an agronomical function, and a higher oil content can be achieved when the caruncle is reduced.
The selection for fast germination reduced the seed coat density, but at same time, it increased the relative seed coat weight (Table 3). These results corroborate the observation that the time for germination is driven by the characteristics of the seed coat; however, it is uncertain which characteristic has a stronger association. In the experiment with the morphological characteristics of individual seeds (Section 3.4), the most important characteristic was the seed coat relative weight. In the study of morphological changes after selection for fast germination (Section 3.5), the seed coat relative weight had the opposite effect (it increased in the progenies), while the change was more pronounced in the seed coat weight and in the seed coat density.
4.6. Pre-Germination Treatments and Initial Growth
The hormonal control of germination is well-documented, with abscisic acid inhibiting and gibberellins promoting this metabolic process [21,29], and castor plants are very sensitive to gibberellin applied to the seed and leaf [4,55]. It was confirmed that the balance of gibberellin and abscisic acid is one of the factors influencing the germination of castor seeds.
The pre-germination of castor seeds failed to promote the fast emergence of the seedlings. Some of the seeds that were pre-germinated for 48 or 60 hours emerged earlier, but just two days later, the untreated seeds outnumbered the pre-germinated seeds. Pre-germination for periods shorter than 48 hours delayed the emergence in relation to the control. It seems that castor seeds can sense fine cues from the environment [12] that inhibit the germination process, and just exposing them to water and temperature is not enough to promote radicle protrusion. When sowed in the soil environment, the right cues for germination were quickly perceived by untreated seeds, while the seeds that had been exposed to short pre-germination (from 12 to 36 hours) took longer to respond to the cues from the soil.
The exponential initial growth of the castor seedlings sheds light on the importance of fast emergence for a high seed yield. A few days of emergence delay creates a gap in the biomass accumulation that results in a reduced capacity to invest in height growth and leaf area (less root growth is presumed). Castor plants have a large plasticity to allocate the biomass among the roots, stems, and leaves according to the availability of water, nutrients, and light [55]. Although larger seeds result in larger seedlings [31], the early growth of castor seedlings may have an advantage because of early germination or because of larger reserves in the seed. This study did not measure how much the initial growth of a castor seedling was influenced by the seed weight, but it seems that, under favorable conditions that usually are found in cropping fields, the additional energy of a large seed is not likely to compensate for an early germination and emergence because the initial exponential growth in the shoot biomass (Figure 7B).
5. Conclusions
Many factors influence the germination of castor seed, and the seed coat plays a major role controlling this physiological process. Castor seed coat is highly permeable to water, and germination is not delayed because of restrictions to hydration. However, the castor seed coat exerts a mechanical restriction to the embryo expansion that acts as a mechanism of germination control. The best temperature for castor seed germination was 31 °C, and no seed germinated at 16 °C. Seed morphology has little influence on the time for germination, and the reduced seed coat relative weight was strongly associated with fast germination. The selection of castor plants for fast germination for three generations caused changes in the seed morphology, making them lighter with an increased caruncle and a reduced seed coat density. Gibberellin applied to the seed favored the germination to occur faster and at a higher rate. The incubation of castor seed as a pre-germination treatment was not effective to reduce the time for seedling emergence.
Funding
This research was partially funded by Terasol Oleos Vegetais (SEG 30.20.90.020.00.00) and Kaiima Brasil Sementes (SEG 30.22.90.036.00.00).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data obtained in these experiments are available upon request or in the links provided below: Seed morphology influencing the time for germination: https://doi.org/10.48432/IZDRRL. Hydration, temperature, puncture, and time for germination: https://doi.org/10.48432/6N3WQA. Seed morphology after selection for fast germination: https://doi.org/10.48432/JIXSGJ. Gibberellin influencing castor seed germination: https://doi.org/10.48432/4VGSL3. Pre-germination treatment and shoot biomass growth: https://doi.org/10.48432/HOPEYU.
Conflicts of Interest
The author declares that this study received funding from the companies Terasol Oleos Vegetais Ltda. And Kaiima Brasil Sementes. The funders were not involved in the study design, collection, analysis, interpretation of data, the writing of this article or on the decision to submit it for publication. The funders did not have access to the content of the article prior to submission for publication.
References
- Witt, T.W.; Flynn, K.C.; Zoz, T.; Lee, T.O.; Monteiro, J.E.B.A. A site suitability analysis for castor (Ricinus communis L.) production during Brazil’s second harvest incorporating disease prediction. Helyon 2023, 9, e18981. [Google Scholar] [CrossRef] [PubMed]
- Yeboah, A.; Ying, S.; Lu, J.; Xie, Y.; Amoanimaa-Dede, H.; Boateng, K.G.A.; Chen, M.; Yin, X. Castor oil (Ricinus communis): A review on the chemical composition and physicochemical properties. Food Sci. Technol. 2021, 41, 399–413. [Google Scholar] [CrossRef]
- Mendes, R.C.; Dias, D.C.F.S.; Pereira, M.D.; Berger, P.G. Pre-germinative treatments in castor bean (Ricinus communis L.) seeds. Rev. Bras. Sementes 2009, 31, 187–194. [Google Scholar] [CrossRef]
- Severino, L.S. Single-seed selection of fast-germinating genotypes of castor (Ricinus communis). Ind. Crops Prod. 2023, 194, 116307. [Google Scholar] [CrossRef]
- de Carvalho, M.L.M.; Alves, R.A.; Oliveira, L.M.D. Radiographic analysis in castor bean seeds (Ricinus communis L.). Rev. Bras. Sementes 2010, 32, 170–175. [Google Scholar] [CrossRef]
- Sun, Y.; Niu, G.; Osuna, P.; Ganjegunte, G.; Auld, D.; Zhao, L.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. Seedling emergence, growth, and leaf mineral nutrition of Ricinus communis L. cultivars irrigated with saline solution. Ind. Crops Prod. 2013, 49, 75–80. [Google Scholar] [CrossRef]
- Gardarin, A.; Colbach, N. How much of seed dormancy in weeds can be related to seed traits? Weed Res. 2015, 55, 14–25. [Google Scholar] [CrossRef]
- Martins, V.F.; Haddad, C.R.B.; Semir, J. Seed germination of Ricinus communis in predicted settings after autochorous and myrmecochorous dispersal. J. Torrey Bot. Soc. 2009, 136, 84–90. [Google Scholar] [CrossRef]
- Cheema, N.M.; Azim Malik, M.; Qadir, G.; Zubair Rafique, M.; Nawaz, N. Influence of temperature and osmotic stress on germination induction of different castor bean cultivars. Pak. J. Bot. 2010, 42, 4035–4041. [Google Scholar]
- Lago, A.A.; Zink, E.; Razera, L.F.; Banzatto, N.V.; Filho, A.S. Seed dormancy of three castor bean cultivars. Bragantia 1979, 38, 41–44. [Google Scholar] [CrossRef]
- Ribeiro, P.R.; Willems, L.A.; Mudde, E.; Fernandez, L.G.; de Castro, R.D.; Ligterink, W.; Hilhorst, H.W. Metabolite profiling of the oilseed crop Ricinus communis during early seed imbibition reveals a specific metabolic signature in response to temperature. Ind. Crops Prod. 2015, 67, 305–309. [Google Scholar] [CrossRef]
- Severino, L.S. Plants make smart decisions in complex environments. Plant Signal. Behav. 2021, 16, e1970448-2. [Google Scholar] [CrossRef]
- Souza, L.A.; Carvalho, M.L.M.; Kataoka, V.Y.; Oliveira, J.A. Eletrical conductivity test to evaluate physiological quality of castor bean seeds. Rev. Bras. Sementes 2009, 31, 60–67. [Google Scholar] [CrossRef]
- David, A.M.S.d.S.; Araújo, E.F.; Araújo, R.F.; de Resende, M.A.V.; Dias, D.C.F.d.S.; Nobre, D.A.C. Physiological quality of castor bean seeds originating from different racemes in the plant. J. Seed Sci. 2013, 35, 248–254. [Google Scholar] [CrossRef]
- Drumond, A.A.L.; Sales, J.d.F.; Zuchi, J.; Camelo, G.N.; Souza, M.M.V. Physiological quality of castor seeds (Ricinus communis L.) after processing. J. Seed Sci. 2019, 41, 224–232. [Google Scholar] [CrossRef]
- Zuchi, J.; Panozzo, L.E.; Heberle, E.; Dias, D.C.F.S. Physiological quality of castor bean seeds classified by size. Rev. Bras. Sementes 2010, 32, 177–183. [Google Scholar] [CrossRef]
- Nobre, D.A.C.; Damascena, J.G.; David, A.M.S.S.; Santos, M.P.; Pereira, A.R.; Pereira, C.G. Pre-germination treatments in castor seeds, cultivar IAC 226. Acta Biol. Colomb. 2013, 18, 473–478. [Google Scholar]
- Qutob, D.; Ma, F.; Peterson, C.A.; Bernards, M.A.; Gijzen, M. Structural and permeability properties of the soybean seed coat. Botany 2008, 86, 219–227. [Google Scholar] [CrossRef]
- Smýkal, P.; Vernoud, V.; Blair, M.W.; Soukup, A.; Thompson, R.D. The role of the testa during development and in establishment of dormancy of the legume seed. Front. Plant Sci. 2014, 5, 351. [Google Scholar]
- Coughlan, J.M.; Saha, A.; Donohue, K. Effects of pre- and post-dispersal temperature on primary and secondary dormancy dynamics in contrasting genotypes of Arabidopsis thaliana (Brassicaceae). Plant Species Biol. 2017, 32, 210–222. [Google Scholar] [CrossRef]
- Duclos, D.V.; Altobello, C.O.; Taylor, A.G. Investigating seed dormancy in switchgrass (Panicum virgatum L.): Elucidating the effect of temperature regimes and plant hormones on embryo dormancy. Ind. Crops Prod. 2014, 58, 148–159. [Google Scholar] [CrossRef]
- Lamichaney, A.; Basavaraja, T.; Katiyar, P.K.; Singh, N.P. Seed germination response of commonbean (Phaseolus vulgaris L.) genotypes to optimal and sub-optimal temperatures. Legume Res. 2021, 44, 419–424. [Google Scholar] [CrossRef]
- Bradbeer, J.W. Seed Dormancy and Germination; Blackie and Son Ltd.: London, UK, 1988; 146p. [Google Scholar]
- Da Silva, E.A.A.; Toorop, P.E.; Van Aelst, A.C.; Hilhorst, H.W.M. Abscisic acid controls embryo growth potential and endo-sperm cap weakening during coffee (Coffea arabica cv. Rubi) seed germination. Planta 2004, 220, 251–261. [Google Scholar] [CrossRef]
- Duclos, D.V.; Ray, D.T.; Johnson, D.J.; Taylor, A.G. Investigating seed dormancy in switchgrass (Panicum virgatum L.): Understanding the physiology and mechanisms of coat-imposed seed dormancy. Ind. Crops Prod. 2013, 45, 377–387. [Google Scholar] [CrossRef]
- Liu, J.; Wang, H.; Luo, F.; Wang, Y.; Xu, C.; Jiang, J. The influence of seed structures on dormancy in seeds of Urochloa hybrid cultivar ‘Mulato Ⅱ’. Trop. Grassl. Forrajes Trop. 2022, 10, 156–163. [Google Scholar] [CrossRef]
- Müller, K.; Tintelnot, S.; Leubner-Metzger, G. Endosperm-limited Brassicaceae seed germination: Abscisic acid inhibits embryo-induced endosperm weakening of Lepidium sativum (cress) and endosperm rupture of cress and Arabidopsis thaliana. Plant Cell Physiol. 2006, 47, 864–877. [Google Scholar] [CrossRef]
- Steinbrecher, T.; Leubner-Metzger, G. The biomechanics of seed germination. J. Exp. Bot. 2017, 68, 765–783. [Google Scholar] [CrossRef]
- Weitbrecht, K.; Müller, K.; Leubner-Metzger, G. First off the mark: Early seed germination. J. Exp. Bot. 2011, 62, 3289–3309. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Shen, Y. Dormancy in Tilia miqueliana is attributable to permeability barriers and mechanical constraints in the endosperm and seed coat. Braz. J. Bot. 2021, 44, 725–740. [Google Scholar] [CrossRef]
- Christie, K.; Pierson, N.R.; Lowry, D.B.; Holeski, L.M. Local adaptation of seed and seedling traits along a natural aridity gradient may both predict and constrain adaptive responses to climate change. Am. J. Bot. 2022, 109, 1529–1544. [Google Scholar] [CrossRef]
- Notarnicola, R.F.; Nicotra, A.B.; Kruuk, L.E.B.; Arnold, P.A. Effects of warming temperatures on germination responses and trade-offs between seed traits in an alpine plant. J. Ecol. 2022, 111, 62–76. [Google Scholar] [CrossRef]
- Vertucci, C.W. The kinetics of seed imbibition: Controlling factors and relevance to seedling vigor. Seed Moisture 2015, 14, 93–115. [Google Scholar]
- Severino, L.S.; Auld, D.L. Seed abortion and the individual weight of castor seed (Ricinus communis L.). Ind. Crops Prod. 2013, 49, 890–896. [Google Scholar] [CrossRef]
- Klamkin, M.S. Corrections to “Elementary approximations to the area of N-dimensional ellipsoids”. Am. Math. Mon. 1976, 83, 478. [Google Scholar] [CrossRef]
- Meyer, C.J.; Steudle, E.; Peterson, C.A. Patterns and kinetics of water uptake by soybean seeds. J. Exp. Bot. 2007, 58, 717–732. [Google Scholar] [CrossRef]
- Silva, A.R.; Leão-Araújo, É.F.; Rezende, B.R.; Santos, W.V.; Santana, H.A.; Silva, S.C.M.; Ferdandes, N.A.; Costa, D.S.; Mesquita, J.C.P. Modeling the three phases of the soaking kinetics of seeds. Agron. J. 2018, 110, 164–170. [Google Scholar] [CrossRef]
- Drumond, A.A.L.; Sales, J.d.F.; Zuchi, J.; Resende, O.; Camelo, G.N.; Souza, M.M.V. Physiological quality of castor bean seed genotypes stored at two temperatures. Acta Sci. Agron. 2019, 42, e43583. [Google Scholar] [CrossRef]
- Lara, B.Y.A.; Ramírez-Pimentel, J.G.; Mancilla, C.L.A.; Prieto, J.C.; Pérez, J.C.R. Breaking dormancy in castor bean (Ricinus communis L.) seeds. Interciencia 2018, 43, 858–863. [Google Scholar]
- Machado, C.G.; Martins, C.C.; Cruz, S.C.S.; Nakagawa, J.; Pereira, F.R.S. Quality of castor bean seeds (Ricinus communis L.) affected by raceme and fruit position during storage. Semin. Ciências Agrárias 2010, 31, 301–312. [Google Scholar] [CrossRef][Green Version]
- Luo, Y.; Loomis, R.S.; Hsiao, T.C. Simulation of soil temperature in crops. Agric. For. Meteorol. 1992, 61, 23–38. [Google Scholar] [CrossRef]
- Silva, V.R.; Reichert, J.M.; Reinert, D.J. Soil temperature variation in three different systems of soil management in blackbeans crop. Rev. Brasil. Ciênc. Solo 2006, 30, 391–399. [Google Scholar] [CrossRef]
- Singh, J.; Guzman, I.; Begna, S.; Trostle, C.; Angadi, S. Germination and early growth response of guar cultivars to low temperatures. Ind. Crops Prod. 2021, 159, 113082. [Google Scholar] [CrossRef]
- Streck, E.A.; Aguiar, G.A.; da Silva, P.U.; Fronza, R.T.L.; Junior, A.M.d.M. Genetic tolerance to low temperatures in irrigated rice. Rev. Cienc. Agron. 2020, 51, e20196938. [Google Scholar] [CrossRef]
- Tobeh, A.; Jamaati-e-Somarin, S. Low temperature stress effect on wheat cultivars germination. Afr. J. Microbiol. Res. 2012, 6, 1265–1269. [Google Scholar]
- Leubner-Metzger, G. Functions and regulation of β-1,3-glucanases during seed germination, dormancy release and after-ripening. Seed Sci. Res. 2003, 13, 17–34. [Google Scholar] [CrossRef]
- Yang, L.; Chen, H.; Xiao, J.; Fan, Y.; Song, S.; Zhang, Y.; Liu, X. Research on structural–mechanical properties during the castor episperm breaking process. Processes 2021, 9, 1777. [Google Scholar] [CrossRef]
- Severino, L.S.; Cordoba, O.J.; Zanotto, M.D.; Auld, D.L. The influence of the caruncle on the germination of castor seed under high salinity or low soil water content. Seed Sci. Technol. 2012, 40, 139–143. [Google Scholar] [CrossRef]
- Moshkin, V.A. Castor; Amerind: New Delhi, India, 1986; pp. 34–36. [Google Scholar]
- Kimmelshue, C.L.; Goggi, S.; Moore, K.J. Seed size, planting depth, and a perennial groundcover system effect on corn emergence and grain yield. Agronomy 2022, 12, 437. [Google Scholar] [CrossRef]
- Bockstaller, C.; Girardin, P. Effects of seed size on maize growth from emergence to silking. Maydica 1994, 39, 213–218. [Google Scholar]
- Gislum, R.; Nikneshan, P.; Shrestha, S.; Tadayyon, A.; Deleuran, L.C.; Boelt, B. Characterisation of castor (Ricinus communis L.) seed quality using Fourier transform near-infrared spectroscopy in combination with multivariate data analysis. Agriculture 2018, 8, 59. [Google Scholar] [CrossRef]
- Westoby, M.; Jurado, E.; Leishman, M. Comparative evolutionary ecology of seed size. Trends Ecol. Evol. 1992, 7, 368–372. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, O.; Jakobsson, A. Recruitment trade-offs and the evolution of dispersal mechanisms in plants. Evol. Ecol. 1999, 13, 411–423. [Google Scholar] [CrossRef]
- Severino, L.S.; Oliveira, E. Plant growth regulators influence the height and biomass partition of castor plants. Braz. J. Agric. Environ. Eng. 2024, 28, e276554. [Google Scholar]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).