Expression Profiling of Non-Aflatoxigenic Aspergillus parasiticus Mutants Obtained by 5-Azacytosine Treatment or Serial Mycelial Transfer

Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strains
2.2. Culture Conditions

2.3. Aflatoxin Evaluation
2.4. Microarray Analyses
2.5. Quantitative RT-PCR (qRT-PCR)

{kind=link}
{kind=link}
| NCBI or TIGR Number | Sequence (5' to 3') | Product size (bp) | |
|---|---|---|---|
| aflJ | TGACTCTCCTTTTGCCGAATGT | 250 | |
| GGGAATGGCAACGGTGGGCG | |||
| XM_001824706 (TC9112) | TATGAGCCCCAACGCAACAGACAG | 268 | |
| CGGGTACGGCTCATTAGAAGGAC | |||
| XM_002372755(TC10739) | GCCCCAGCTTACCAAACGAGA | 336 | |
| AGGGATGAGGGGGAATGAAGTG | |||
| XM_002373878(TC10202) | TGCGGGAAATAATACGACAGGAA | 359 | |
| AAAGTAGCGACCCGGGCAGTAGTG | |||
| XM_002380627(TC11938) | GACCGCATCGTGGCTTTTCTAC | 334 | |
| AAGCATCATTCGTCATTCGTTCTG | |||
| XM_002374352(TC9364) | CCCACCGAAGACCTGACCTACAT | 322 | |
| CTGCCATCTGCCAAACTCCATTA | |||
3. Results
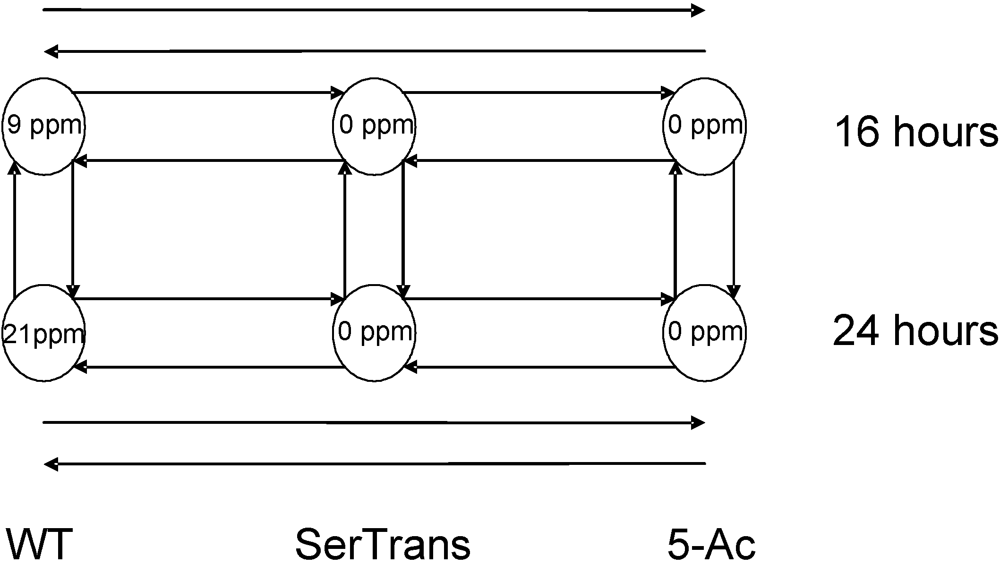
3.1. Loss of Aflatoxin Production after 5-Azacytosine Treatment and Serial Mycelial Transfer
3.2. Microarray Transcription Profiles in 5-Azacytosine-Treated and Serial Transfer Isolated Colonies Compared to Wild-Type
| Gene | Ratios | Probabilities b | ||
|---|---|---|---|---|
| Untreated vs. 5-AC-treated | Untreated vs. 5-AC-treated | Untreated vs. Serial transfer | Untreated vs. Serial transfer | |
| adhA | 1.22 | 1.10 | 0.4388 | 0.3307 |
| aflJ * | 1.76 | 1.59 | 0.0018 | 0.0015 |
| aflR | 1.29 | 1.12 | 0.3367 | 0.3465 |
| aflT | 1.32 | 1.11 | 0.3321 | 0.2423 |
| avfA | 1.31 | 1.17 | 0.2902 | 0.4271 |
| avnA | 1.25 | 1.18 | 0.3835 | 0.2162 |
| cypX | 1.33 | 1.15 | 0.1235 | 0.0835 |
| estA | 1.33 | 1.25 | 0.1929 | 0.0493 |
| hexA | 1.37 | 1.12 | 0.0768 | 0.1850 |
| hypB1 | 1.24 | 0.95 | 0.2552 | 0.8827 |
| hypB2 | 2.49 | 2.16 | 0.4722 | 0.5134 |
| moxY | 1.26 | 1.09 | 0.1828 | 0.1875 |
| nadA | 1.24 | 1.07 | 0.4882 | 0.4211 |
| nor-1 | 1.41 | 1.12 | 0.0570 | 0.0620 |
| norA | 1.33 | 1.07 | 0.0553 | 0.2726 |
| omtA | 1.35 | 1.28 | 0.0422 | 0.0741 |
| omtB | 1.37 | 1.08 | 0.0385 | 0.2716 |
| ordA | 1.34 | 1.25 | 0.2363 | 0.1889 |
| ordB | 1.35 | 1.31 | 0.1566 | 0.0602 |
| pksA | 1.27 | 1.08 | 0.2264 | 0.1826 |
| sugR | 1.32 | 1.10 | 0.3869 | 0.3722 |
| vbs | 1.26 | 1.05 | 0.1547 | 0.1843 |
| ver-1 | 1.23 | 1.10 | 0.7285 | 0.9483 |
| verA | 1.16 | 1.08 | 0.5312 | 0.8985 |
| verB | 1.24 | 1.15 | 0.1683 | 0.1181 |
| TC Number or EST b | Best Match GenBank c | Putative Protein | Ratios d | Probabilities d | ||
|---|---|---|---|---|---|---|
| Untreated vs. 5-AC-treated | Untreated vs. Serial transfer | Untreated vs. 5-AC-treated | Untreated vs. Serial transfer | |||
| TC8344 | XM_001824592 | HET&ankyrin domain protein | 10.02 | 8.53 | 0.0000 | 0.0000 |
| TC10739 * | XM_002372755 | Ankyrin repeat-containing protein | 5.69 | 4.52 | 0.0000 | 0.0000 |
| TC9364 * | XM_002374352 | FG-GAP (integrin) repeat protein | 4.79 | 3.97 | 0.0000 | 0.0000 |
| TC9256 | XM_002375859 | Nitrite reductase (NiiA) | 4.58 | 3.38 | 0.0000 | 0.0000 |
| NAGAO52TV | XM_002380681 | Polysaccharide synthase Cps1 | 4.54 | 4.72 | 0.0000 | 0.0000 |
| TC10370 | XM_002384463 | Opsin | 4.18 | 3.49 | 0.0000 | 0.0000 |
| TC8557 | XM_001827098 | alpha1,3-glucan synthase | 3.71 | 2.62 | 0.0000 | 0.0000 |
| TC10767 * | XM_003096248 | Integral membrane protein | 3.54 | 2.49 | 0.0000 | 0.0002 |
| TC10268 | XM_002383371 | 12 kda heat shock protein | 3.53 | 3.48 | 0.0000 | 0.0000 |
| TC9112 * | XM_001824706 | GPI-anchored protein | 3.45 | 3.00 | 0.0000 | 0.0000 |
| TC10523 | XM_002374369 | Integral membrane prot-exocytic transport from Golgi | 3.40 | 3.53 | 0.0000 | 0.0000 |
| NAGCY20TV | XM_002379925 | Gal4 transcription factor-AFLA_139560 | 3.28 | 3.20 | 0.0000 | 0.0000 |
| NAGAE41TV | XM_002382445 | Calcium binding protein Caleosin | 3.23 | 3.09 | 0.0000 | 0.0000 |
| TC10524 | XM_002374369 | Exocytic transport protein | 3.21 | 3.28 | 0.0000 | 0.0000 |
| TC10334 | XM_002379834 | GABA-permease | 3.11 | 2.69 | 0.0000 | 0.0000 |
| TC11140 | XM_002383723 | MFS-transporter-related | 3.02 | 2.18 | 0.0000 | 0.0007 |
| TC11938 * | XM_002380627 | EsdC | 2.98 | 2.71 | 0.0000 | 0.0000 |
| NAFCO41TV | XM_001819214 | Transcriptional regulatory protein pro1 (HLH protein) | 2.88 | 2.35 | 0.0000 | 0.0000 |
| TC11262 | XM_002384600 | Helix-loop-helix DNA-binding domain protein | 2.75 | 2.40 | 0.0000 | 0.0001 |
| TC10225 | XM_003189329 | SH3 domain kinase binding protein (predicted membrane protein) | 2.70 | 2.47 | 0.0000 | 0.0000 |
| TC8802 | XM_002378642 | Ser-thr rich protein AOS28(solid-state culture specific gene) | 2.66 | 2.63 | 0.0000 | 0.0000 |
| TC11294 | XM_002384659 | Arrestin domain protein | 2.54 | 1.92 | 0.0000 | 0.0007 |
| TC8781 | XM_002375350 | MAPEG superfamily protein pfam01124 | 2.45 | 1.81 | 0.0000 | 0.0003 |
| TC10523 | XM_002374369 | Hypothetical protein | 2.40 | 2.54 | 0.0000 | 0.0000 |
| TC10950 | XM_002377280 | RTA domain protein (RTA1 superfamily, pfam04479) | 2.15 | 1.82 | 0.0000 | 0.0000 |
| TC8911 | XM_002383317 | Aspartic endopeptidase; aspartyl proteinase | 2.13 | 1.82 | 0.0000 | 0.0002 |
| TC8680 | XM_002382580 | GPI-anchored glycoprotein (AFLA_004200) | 2.03 | 1.98 | 0.0000 | 0.0000 |
| TC9120 | XM_002381180 | bZIP transcription factor ATFB (atf21, AFLA_094010) | 2.02 | 1.96 | 0.0000 | 0.0000 |
| TC8511 | XM_002383753 | Hypothetical protein | 1.92 | 2.11 | 0.0000 | 0.0000 |
| NAFBR39TV | XM_002378094 | alpha/beta hydrolase domain protein | 1.90 | 1.66 | 0.0000 | 0.0002 |
| TC9105 | XM_001824752 | Hypothetical protein | 1.87 | 1.96 | 0.0000 | 0.0000 |
| TC10397 | XM_002379378 | PHD finger and SET domain protein | 1.67 | 1.75 | 0.0000 | 0.0000 |
| TC9269 | XM_002384586 | Metallophophodiesterase (AFLA_119200) | 1.66 | 1.55 | 0.0000 | 0.0001 |
| TC9718 | XM_002372994 | CVNH superfamily (cyanovirin-N family, pfam08881) | 0.53 | 0.61 | 0.0000 | 0.0001 |
| NAFEY64TV | XM_002377133 | Asparagine synthase (glutamine-hydrolyzing) | 0.49 | 0.52 | 0.0001 | 0.0003 |
| Gene | Ratios | Probabilities | Expressed Sequence Tag (EST) | GenBank Accession Number | AFLA # | |||
|---|---|---|---|---|---|---|---|---|
| Untreated vs. 5-AC-treated | Untreated vs. Serial transfer | Untreated vs. 5-AC-treated | Untreated vs. Serial transfer | |||||
| laeA | 0.75 | 0.77 | 0.8998 | 0.8874 | Unknown | XM_002374798 | 033290 | |
| laeA-1 1 | 0.41 | 0.40 | 0.0002 | 0.0002 | TC9243 | XM_002375064 | 035950 | |
| veA | 0.76 | 0.68 | 0.0307 | 0.0041 | TC11465 | XM_002380164 | 066460 | |
| nsdD | 1.83 | 0.99 | 0.0145 | 0.9512 | TC10891 | XM_002376000 | 020210A | |
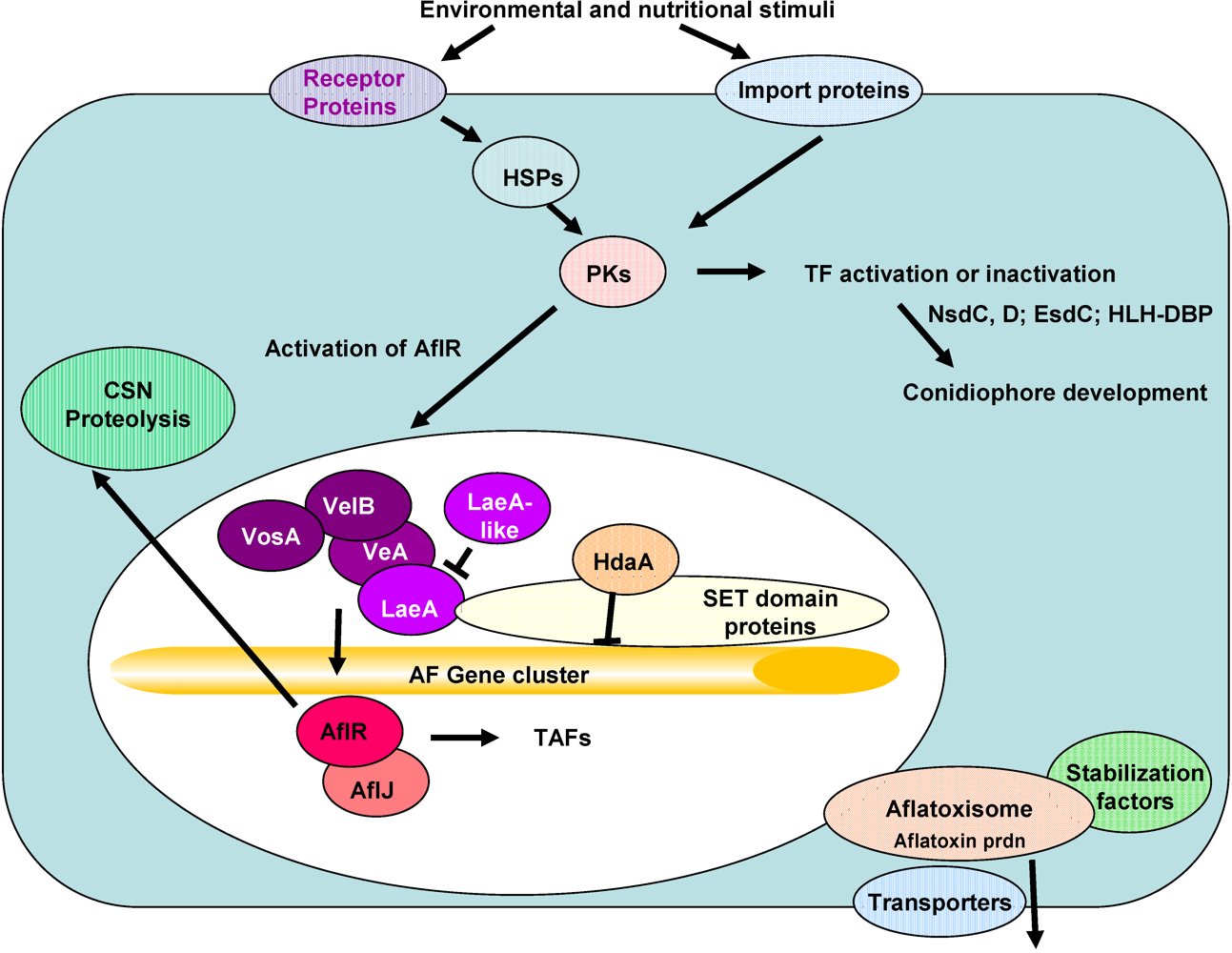
4. Discussion

5. Conclusions
Acknowledgements
References
- Etxebeste, O.; Ni, M.; Garzia, A.; Kwon, N.J.; Fischer, R.; Yu, J.H.; Espeso, E.A.; Ugalde, U. Basic-zipper-type transcription factor FlbB controls asexual development in Aspergillus nidulans. Eukaryot. Cell 2008, 7, 38–48. [Google Scholar] [PubMed]
- Souza-Cletus, A.D.; Lee, B.N.; Adams, T.H. Characterization of the role of the FluG protein in asexual development of Aspergillus nidulans. Genetics 2001, 158, 1027–1036. [Google Scholar] [PubMed]
- Shimizu, K.; Hicks, J.K.; Huang, T.P.; Keller, N.P. Pka, Ras and RGS protein interactions regulate activity of AflR, a Zn(II)2Cys6 transcription factor in Aspergillus nidulans. Genetics 2003, 165, 1095–1104. [Google Scholar] [PubMed]
- Bok, J.W.; Keller, N.P. LaeA, a regulator of secondary metabolism in Aspergillus spp. Eukaryot. Cell 2004, 3, 527–535. [Google Scholar] [CrossRef] [PubMed]
- Bok, J.W.; Noordermeer, D.; Kale, S.P.; Keller, N.P. Secondary metabolic gene cluster silencing in Aspergillus nidulans. Mol. Microbiol. 2006, 61, 1636–1645. [Google Scholar] [CrossRef] [PubMed]
- Duran, R.M.; Cary, J.W.; Calvo, A.M. The role of veA in Aspergillus flavus infection of peanut, corn and cotton. Open Mycol. 2009, 3, 27–36. [Google Scholar] [CrossRef]
- Kale, S.P.; Milde, L.; Trapp, M.K.; Frisvad, J.C.; Keller, N.P.; Bok, J.W. Requirement of LaeA for secondary metabolism and sclerotial production in Aspergillus flavus. Fungal Genet. Biol. 2008, 45, 1422–1429. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Chang, P.-K.; Cleveland, T.E.; Bennett, J.W. Aflatoxins. In Encyclopedia of Industrial Biotechnology: Bioprocess, Bioseparation, and Cell Technology; Flickinger, M.C., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2010; Volume 1, pp. 1–12. [Google Scholar]
- Bayram, O.; Krappmann, S.; Ni, M.; Bok, J.W.; Helmstaedt, K.; Valerius, O.; Braus-Stromeyer, S.; Kwon, N.J.; Keller, N.P.; Yu, J.H.; et al. VelB/VeA/LaeA complex coordinates light signal with fungal development and secondary metabolism. Science 2008, 320, 1504–1506. [Google Scholar]
- Brodhagen, M.; Keller, N.P. Signalling pathways connecting mycotoxin production and sporulation. Mol. Plant Pathol. 2006, 7, 285–301. [Google Scholar]
- Calvo, A.M.; Bok, J.; Brooks, W.; Keller, N.P. veA is required for toxin and sclerotial production in Aspergillus parasiticus. Appl. Environ. Microbiol. 2004, 70, 4733–4739. [Google Scholar] [CrossRef]
- Calvo, A.M.; Wilson, R.A.; Bok, J.-W.; Keller, N.P. Relationship between secondary metabolism and fungal development. Microbiol. Mol. Biol. Rev. 2002, 66, 447–459. [Google Scholar]
- Palmer, J.M.; Perrin, R.M.; Dagenais, T.R.T.; Keller, N.P. H3K9 methylation regulates growth and development in Aspergillus fumigatus. Eukaryot. Cell 2008, 7, 2052–2060. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.R.; Chae, K.S.; Han, K.H.; Han, D.M. The nsdC gene encoding a putative C2H2-type transcription factor is a key activator of sexual development in Aspergillus nidulans. Genetics 2009, 182, 771–783. [Google Scholar] [CrossRef] [PubMed]
- Han, K.H.; Han, K.Y.; Yu, J.H.; Chae, K.S.; Jahng, K.Y.; Han, D.M. The nsdD gene encodes a putative GATA-type transcription factor necessary for sexual development of Aspergillus nidulans. Mol. Microbiol. 2001, 41, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Chang, P.K.; Ehrlich, K.C.; Cary, J.W.; Bhatnagar, D.; Cleveland, T.E.; Payne, G.A.; Linz, J.E.; Woloshuk, C.P.; Bennett, J.W. Clustered pathway genes in aflatoxin biosynthesis. Appl. Environ. Microbiol. 2004, 70, 1253–1262. [Google Scholar]
- Du, W.; Obrian, G.R.; Payne, G.A. Function and regulation of aflJ in the accumulation of aflatoxin early pathway intermediate in Aspergillus flavus. Food Addit. Contam. 2007, 24, 1043–1050. [Google Scholar] [CrossRef] [PubMed]
- Chang, P.K. The Aspergillus parasiticus protein AFLJ interacts with the aflatoxin pathway-specific regulator AFLR. Mol. Genet. Genomics 2003, 268, 711–719. [Google Scholar] [PubMed]
- Meyers, D.M.; Obrian, G.; Du, W.L.; Bhatnagar, D.; Payne, G.A. Characterization of aflJ, a gene required for conversion of pathway intermediates to aflatoxin. Appl. Environ. Microbiol. 1998, 64, 3713–3717. [Google Scholar] [PubMed]
- Ehrlich, K.C.; Montalbano, B.G.; Cary, J.W. Binding of the C6-zinc cluster protein, AFLR, to the promoters of aflatoxin pathway biosynthesis genes in Aspergillus parasiticus. Gene 1999, 230, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Chang, P.-K.; Ehrlich, K.C.; Yu, J.; Bhatnagar, D.; Cleveland, T.E. Increased expression of Aspergillus parasiticus aflR, encoding a sequence-specific DNA-binding protein, relieves nitrate inhibition of aflatoxin biosynthesis. Appl. Environ. Microbiol. 1995, 61, 2372–2377. [Google Scholar] [PubMed]
- Ehrlich, K.C.; Montalbano, B.G.; Bhatnagar, D.; Cleveland, T.E. Alteration of different domains in AFLR affects aflatoxin pathway metabolism in Aspergillus parasiticus transformants. Fungal Genet. Biol. 1998, 23, 279–287. [Google Scholar]
- Cary, J.W.; Ehrlich, K.C.; Wright, M.; Chang, P.-K.; Bhatnagar, D. Generation of aflR disruption mutants of A. parasiticus. Appl. Microbiol. Biotechnol. 2000, 53, 680–684. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, K.C.; Mack, B.M.; Cary, J.W.; Bhatnagar, D.; Kale, S.P. A hypothesis to explain how LaeA specifically regulates certain secondary metabolite biosynthesis gene clusters. World Mycotoxin J. 2011, 4, 53–58. [Google Scholar]
- Chang, P.K. Lack of interaction between AFLR and AFLJ contributes to nonaflatoxigenicity of Aspergillus sojae. J. Biotechnol. 2004, 107, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Roze, L.V.; Chanda, A.; Linz, J.E. Compartmentalization and molecular traffic in secondary metabolism: A new understanding of established cellular processes. Fungal Genet. Biol. 2011, 48, 35–48. [Google Scholar]
- Chanda, A.; Roze, L.V.; Linz, J.E. A possible role for exocytosis in aflatoxin export in Aspergillus parasiticus. Eukaryot. Cell 2010, 9, 1724–1727. [Google Scholar] [CrossRef] [PubMed]
- Chanda, A.; Roze, L.V.; Pastor, A.; Frame, M.K.; Linz, J.E. Purification of a vesicle-vacuole fraction functionally linked to aflatoxin synthesis in Aspergillus parasiticus. J. Microbiol. Methods 2009, 78, 28–33. [Google Scholar] [CrossRef] [PubMed]
- Chanda, A.; Roze, L.V.; Kang, S.; Artymovich, K.A.; Hicks, G.R.; Raikhel, N.V.; Calvo, A.M.; Linz, J.E. A key role for vesicles in fungal secondary metabolism. Proc Natl. Acad. Sci. USA 2009, 106, 19533–19538. [Google Scholar]
- Kale, S.P.; Bhatnagar, D.; Bennett, J.W. Isolation and characterization of morphological variants of Aspergillus parasiticus deficient in secondary metabolism production. Mycol. Res. 1994, 98, 645–652. [Google Scholar] [CrossRef]
- Kale, S.P.; Cary, J.W.; Bhatnagar, D.; Bennett, J.W. Characterization of experimentally induced, nonaflatoxigenic variant strains of Aspergillus parasiticus. Appl. Environ. Microbiol. 1996, 62, 3399–3404. [Google Scholar] [PubMed]
- Kale, S.P.; Cary, J.W.; Hollis, N.; Wilkinson, J.R.; Bhatnagar, D.; Yu, J.; Cleveland, T.E.; Bennett, J.W. Analysis of aflatoxin regulatory factors in serial transfer-induced non-aflatoxigenic Aspergillus parasiticus. Food Addit. Contam. 2007, 24, 1061–1069. [Google Scholar] [CrossRef] [PubMed]
- Tamame, M.; Antequera, F.; Villanueva, J.R.; Santos, T. High-frequency conversion to a “fluffy” developmental phenotype in Aspergillus spp. by 5-azacytidine treatment: evidence for involvement of a single nuclear gene. Mol. Cell. Biol. 1983, 3, 2287–2297. [Google Scholar] [PubMed]
- Tamame, M.; Antequera, F.; Santos, E. Developmental characterization and chromosomal mapping of the 5- azacytidine-sensitive fluF locus of Aspergillus nidulans. Mol. Cell. Biol. 1988, 8, 3043–3050. [Google Scholar] [PubMed]
- Lee, D.W.; Freitag, M.; Selker, E.U.; Aramayo, R. A cytosine methyltransferase homologue is essential for sexual development in Aspergillus nidulans. PLoS One 2008, 3, e2531. [Google Scholar] [PubMed]
- Gowher, H.; Ehrlich, K.C.; Jeltsch, A. DNA from Aspergillus flavus contains 5-methylcytosine. FEMS Microbiol. Lett. 2001, 205, 151–155. [Google Scholar] [CrossRef] [PubMed]
- Doiron, K.M.; Lavigne-Nicolas, J.; Cupples, C.G. Effect of interaction between 5-azacytidine and DNA (cytosine-5) methyltransferase on C-to-G and C-to-T mutations in Escherichia coli. Mutat Res. 1999, 429, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Abdollahi, A.; Buchanan, B.L. Regulation of aflatoxin biosynthesis: Induction of aflatoxin production by various carbohydrates. J. Food Sci. 1981, 46, 633–635. [Google Scholar]
- Dutton, M.F. Enzymes and aflatoxin biosynthesis. Microbiol. Rev. 1988, 52, 274–295. [Google Scholar]
- Sobolev, V.S.; Dorner, J.W. Cleanup procedure for determination of aflatoxin in major agricultural commodities by liquid chromatography. J. AOAC Int. 2002, 85, 642–645. [Google Scholar] [PubMed]
- Price, M.S.; Conners, S.B.; Tachdjian, S.; Kelly, R.M.; Payne, G.A. Aflatoxin conducive and non-conducive growth conditions reveal new gene associations with aflatoxin production. Fungal Genet. Biol. 2005, 42, 506–518. [Google Scholar]
- Wilkinson, J.R.; Yu, J.; Abbas, H.K.; Scheffler, B.E.; Kim, H.S.; Nierman, W.C.; Bhatnagar, D.; Cleveland, T.E. Aflatoxin formation and gene expression in response to carbon source media shift in Aspergillus parasiticus. Food Addit Contam. 2007, 24, 1051–1060. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, J.R.; Yu, J.; Bland, J.M.; Nierman, W.C.; Bhatnagar, D.; Cleveland, T.E. Amino acid supplementation reveals differential regulation of aflatoxin biosynthesis in Aspergillus flavus NRRL 3357 and Aspergillus parasiticus SRRC 143. Appl. Microbiol. Biotechnol. 2007, 74, 1308–1319. [Google Scholar] [PubMed]
- Chang, P.K.; Wilkinson, J.R.; Horn, B.W.; Yu, J.; Bhatnagar, D.; Cleveland, T.E. Genes differentially expressed by Aspergillus flavus strains after loss of aflatoxin production by serial transfers. Appl. Microbiol. Biotechnol. 2007, 77, 917–925. [Google Scholar] [CrossRef] [PubMed]
- Brazma, A.; Hingamp, P.; Quackenbush, J.; Sherlock, G.; Spellman, P.; Stoeckert, C.; Aach, J.; Ansorge, W.; Ball, C.A.; Causton, H.C.; et al. Minimum information about a microarray experiment (MIAME)-toward standards for microarray data. Nat. Genet. 2001, 29, 365–371. [Google Scholar]
- Kale, S.; Bennett, J.W. Strain instability in filamentous fungi. In Handbook of Applied Mycology: Volume 5: Mycotoxins in Ecological Systems; Bjhatnagar, D., Lillehoj, E.B., Arora, D.K., Eds.; Marcel Dekker: New York, NY, USA, 1992; pp. 311–332. [Google Scholar]
- Chang, P.K.; Wilkinson, J.W.; Horn, B.W.; Yu, J.; Bhatnagar, D.; Cleveland, T.E. Genes differentially expressed by Aspergillus flavus strains after loss of aflatoxin production by serial transfers. Appl. Microbiol. Biotechnol. 2007, 77, 917–925. [Google Scholar] [CrossRef] [PubMed]
- Fedorova, N.D.; Badger, J.H.; Robson, G.D.; Wortman, J.R.; Nierman, W.C. Comparative analysis of programmed cell death pathways in filamentous fungi. BMC Genomics 2005, 6, 177–190. [Google Scholar]
- Mosavi, L.K.; Cammett, T.J.; Desrosiers, D.C.; Peng, Z.Y. The ankyrin repeat as molecular architecture for protein recognition. Protein Sci. 2004, 13, 1435–1448. [Google Scholar]
- Shichida, Y.; Matsuyama, T. Evolution of opsins and phototransduction. Philos. Trans. R. Soc. Lond. B 2009, 364, 2881–2895. [Google Scholar]
- Hynes, R.O. Integrins: Bidirectional, allosteric signaling machines. Cell 2002, 110, 673–687. [Google Scholar]
- Naested, H.; Frandsen, G.I.; Jauh, G.Y.; Hernandez-Pinzon, I.; Nielsen, H.B.; Murphy, D.J.; Rogers, J.C.; Mundy, J. Caleosins: Ca2+-binding proteins associated with lipid bodies. Plant Mol. Biol. 2000, 44, 463–476. [Google Scholar]
- Do, J.H.; Yamaguchi, R.; Miyano, S. Exploring temporal transcription regulation structure of Aspergillus fumigatus in heat shock by state space model. BMC Genomics 2009, 10, 306. [Google Scholar]
- Zhang, D.; Aravind, L. Identification of novel families and classification of the C2 domain superfamily elucidate the origin and evolution of membrane targeting activities in eukaryotes. Gene 2010, 469, 18–30. [Google Scholar]
- Varma, R.; Mayor, S. GPI-anchored proteins are organized in submicron domains at the cell surface. Nature 1998, 394, 798–801. [Google Scholar]
- Sakamoto, K.; Iwashita, K.; Yamada, O.; Kobayashi, K.; Mizuno, A.; Akita, O.; Mikami, S.; Shimoi, H.; Gomi, K. Aspergillus oryzae atfA controls conidial germination and stress tolerance. Fungal Genet. Biol. 2009, 46, 887–897. [Google Scholar] [CrossRef] [PubMed]
- Adhvaryu, K.K.; Morris, S.A.; Strahl, B.D.; Selker, E.U. Methylation of histone H3 lysine 36 is required for normal development in Neurospora crassa. Eukaryot. Cell 2005, 4, 1455–1464. [Google Scholar] [CrossRef] [PubMed]
- Freitag, M.; Hickey, P.C.; Khlafallah, T.K.; Read, N.D.; Selker, E.U. HP1 is essential for DNA methylation in Neurospora. Mol. Cell. 2004, 13, 427–434. [Google Scholar] [PubMed]
- Zhang, X.; Tamaru, H.; Khan, S.I.; Horton, J.R.; Keefe, L.J.; Selker, E.U.; Cheng, X. Structure of the Neurospora SET domain protein DIM-5, a histone H3 lysine methyltransferase. Cell 2002, 111, 117–127. [Google Scholar]
- Palmer, J.M.; Keller, N.P. Delving into the Mechanism of LaeA Regulated Secondary Metabolism. Fungal Genetics Reports, Volume 56- Supplement. In Proceedings of the 25th Fungal Genetics Conference at Asilomar, Asilomar, CA, USA, 17-22 March 2009.
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wilkinson, J.R.; Kale, S.P.; Bhatnagar, D.; Yu, J.; Ehrlich, K.C. Expression Profiling of Non-Aflatoxigenic Aspergillus parasiticus Mutants Obtained by 5-Azacytosine Treatment or Serial Mycelial Transfer. Toxins 2011, 3, 932-948. https://doi.org/10.3390/toxins3080932
Wilkinson JR, Kale SP, Bhatnagar D, Yu J, Ehrlich KC. Expression Profiling of Non-Aflatoxigenic Aspergillus parasiticus Mutants Obtained by 5-Azacytosine Treatment or Serial Mycelial Transfer. Toxins. 2011; 3(8):932-948. https://doi.org/10.3390/toxins3080932
Chicago/Turabian StyleWilkinson, Jeffrey R., Shubha P. Kale, Deepak Bhatnagar, Jiujiang Yu, and Kenneth C. Ehrlich. 2011. "Expression Profiling of Non-Aflatoxigenic Aspergillus parasiticus Mutants Obtained by 5-Azacytosine Treatment or Serial Mycelial Transfer" Toxins 3, no. 8: 932-948. https://doi.org/10.3390/toxins3080932
APA StyleWilkinson, J. R., Kale, S. P., Bhatnagar, D., Yu, J., & Ehrlich, K. C. (2011). Expression Profiling of Non-Aflatoxigenic Aspergillus parasiticus Mutants Obtained by 5-Azacytosine Treatment or Serial Mycelial Transfer. Toxins, 3(8), 932-948. https://doi.org/10.3390/toxins3080932