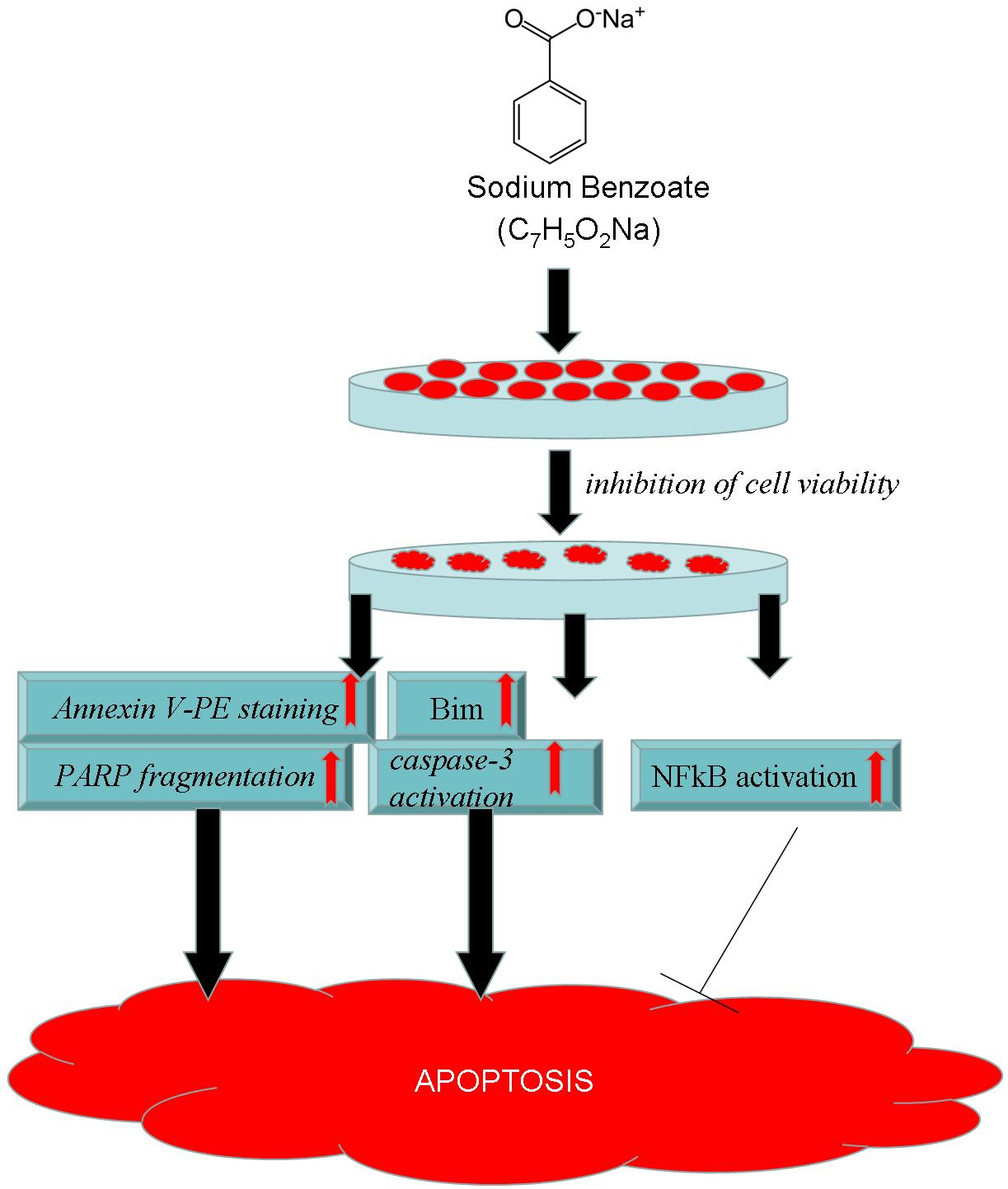
Food Additive Sodium Benzoate (NaB) Activates NF?B and Induces Apoptosis in HCT116 Cells

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. NaB Inhibited Proliferation of HCT116 Colon Cancer Cells at Concentrations Higher than 6 mM
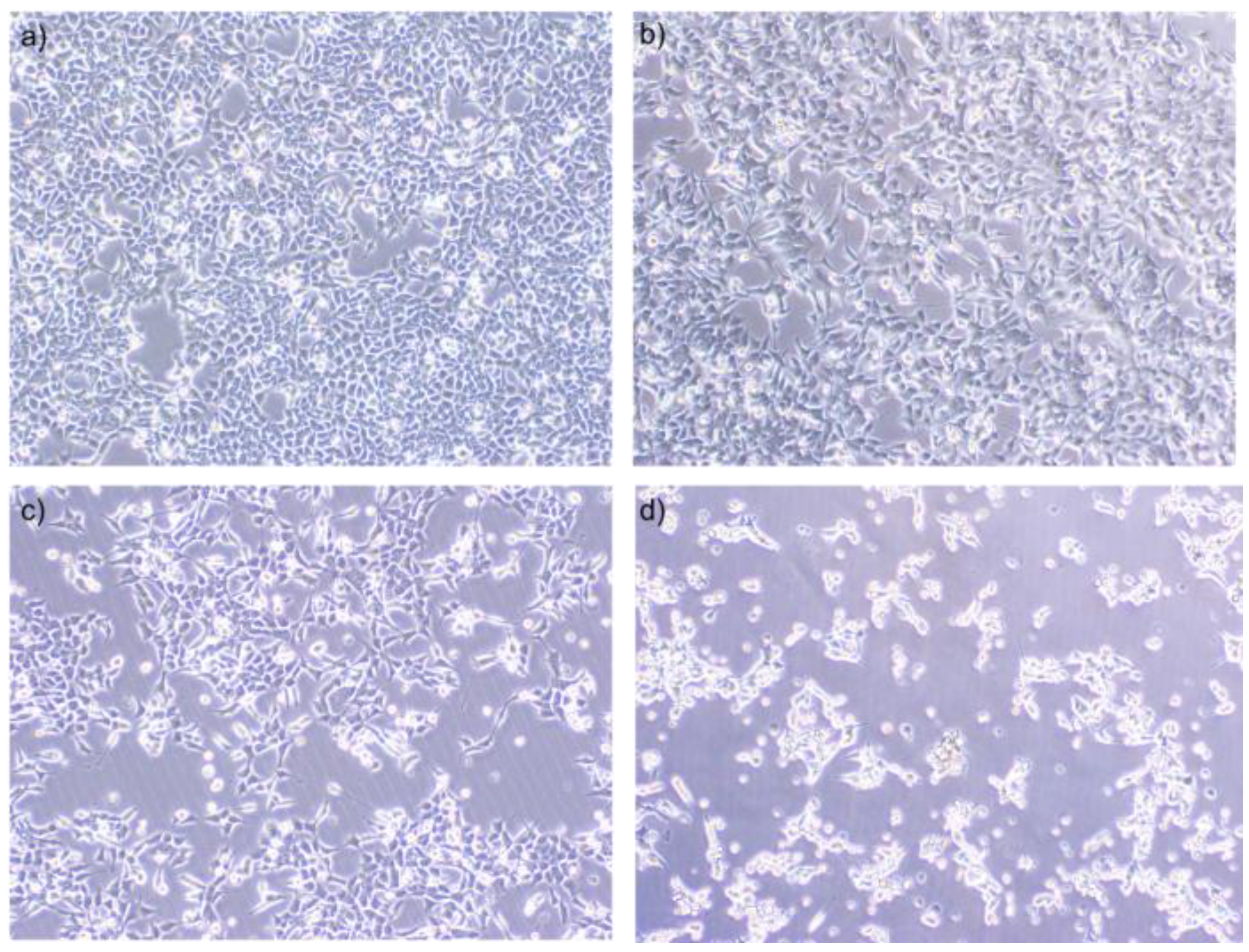
2.2. NaB Treatment Induced Morphological Changes in HCT116 Colon Cancer Cells
2.3. Effect of NaCl on the Viability of HCT116 Colon Cancer Cells
2.4. NaB Exhibited Less Cytotoxic Activity on L929 Fibroblast Cells Compared to HCT116 Cells
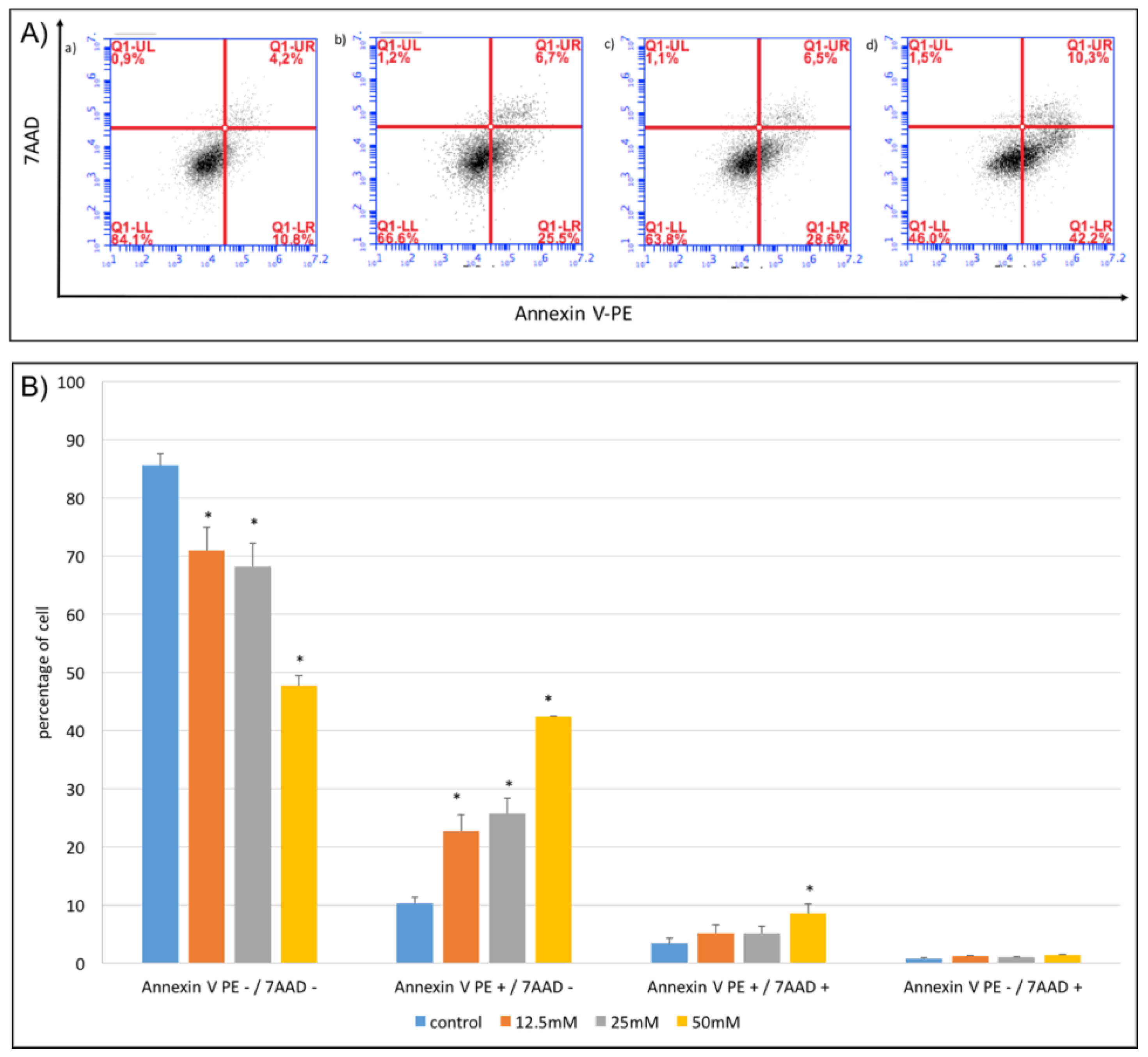
2.5. NaB Induced Loss in HCT116 Cell Viability Was Mediated by Apoptosis
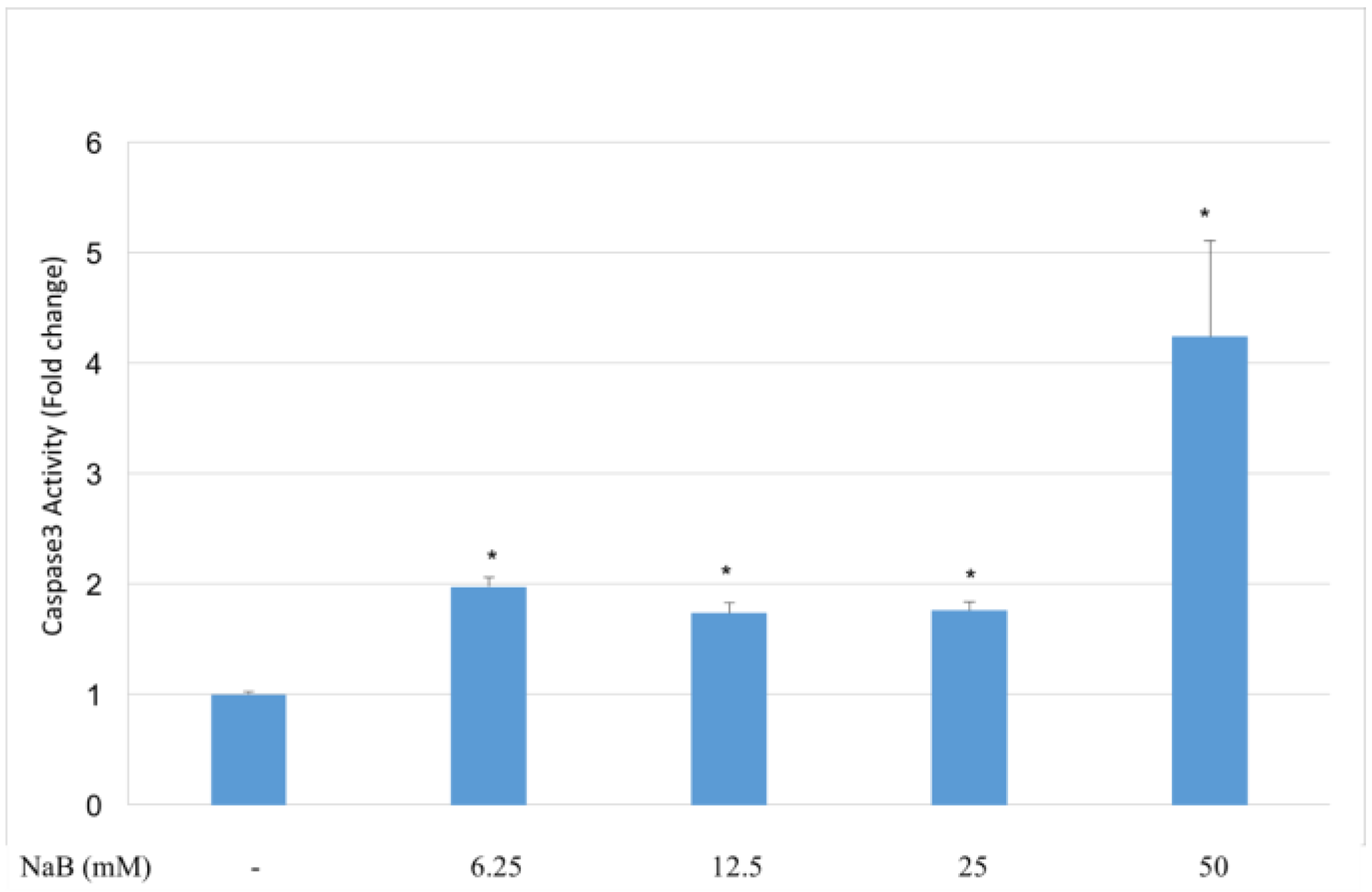
2.6. NaB Induced Caspase-3 Activation in HCT116 Cells at Its Apoptotic Concentrations
2.7. NFκB Activation Accompanies NaB Induced Loss of Cell Viability and Inhibition of NFκB Activation did not Induce a Massive Effect on Cell Viability, but Induced a More Apoptotic Profile Which May Involve Changed Bim but not Bcl-xl Expression
3. Discussion
4. Materials and Methods
4.1. Cell Line and Chemicals
4.2. Cell Culture
4.3. Cell Viability Experiments
4.4. Caspase-3 Activity Assay
4.5. Flow Cytometry for Detection of Apoptosis by Annexin V-PE and 7-AAD Staining
4.6. Western Blot for NFκB-p65, Bcl-xl, Bim and PARP
4.7. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Select Committee on GRAS Substances (SCOGS) Opinion: Benzoic Acid, Sodium Benzoate. Available online: https://www.fda.gov/Food/IngredientsPackagingLabeling/GRAS/SCOGS/ucm260036.htm (accessed on 15 February 2018).
- Husson, M.C.; Schiff, M.; Fouilhoux, A.; Cano, A.; Dobbelaere, D.; Brassier, A.; Mention, K.; Arnoux, J.B.; Feillet, F.; Chabrol, B.; et al. Efficacy and safety of i.v. NaB in urea cycle disorders: A multicentre retrospective study. Orphanet J. Rare Dis. 2016, 11, 127. [Google Scholar] [CrossRef] [PubMed]
- Pahan, K. Immunomodulation of experimental allergic encephalomyelitis by cinnamon metabolite sodium benzoate. Immunopharmacol. Immunotoxicol. 2011, 33, 586–593. [Google Scholar] [CrossRef] [PubMed]
- Brahmachari, S.; Pahan, K. NaB, a food additive and a metabolite of cinnamon, modifies T cells at multiple steps and inhibits adoptive transfer of experimental allergic encephalomyelitis. J. Immunol. 2007, 179, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Rezaei, N.; Amirghofran, Z.; Nikseresht, A.; Ashjazade, N.; Zoghi, S.; Tahvili, S.; Kamali-Sarvestani, E. In Vitro Effects of NaB on Th1/Th2 Deviation in Patients with Multiple Sclerosis. Immunol. Investig. 2016, 45, 679–691. [Google Scholar] [CrossRef] [PubMed]
- Maier, E.; Kurz, K.; Jenny, M.; Schennach, H.; Ueberall, F.; Fuchs, D. Food preservatives NaB and propionic acid and colorant curcumin suppress Th1-type immune response in vitro. Food Chem. Toxicol. 2010, 48, 1950–1956. [Google Scholar] [CrossRef] [PubMed]
- Ciardi, C.; Jenny, M.; Tschoner, A.; Ueberall, F.; Patsch, J.; Pedrini, M.; Ebenbichler, C.; Fuchs, D. Food additives such as sodium sulphite, NaB and curcumin inhibit leptin release in lipopolysaccharide-treated murine adipocytes in vitro. Br. J. Nutr. 2012, 107, 826–833. [Google Scholar] [CrossRef] [PubMed]
- Modi, K.K.; Jana, M.; Mondal, S.; Pahan, K. Sodium Benzoate, a Metabolite of Cinnamon and a Food Additive, Upregulates Ciliary Neurotrophic Factor in Astrocytes and Oligodendrocytes. Neurochem. Res. 2015, 40, 2333–2347. [Google Scholar] [CrossRef] [PubMed]
- Modi, K.K.; Roy, A.; Brahmachari, S.; Rangasamy, S.B.; Pahan, K. Cinnamon and Its Metabolite Sodium Benzoate Attenuate the Activation of p21rac and Protect Memory and Learning in an Animal Model of Alzheimer’s Disease. PLoS ONE 2015, 10, e0130398. [Google Scholar] [CrossRef] [PubMed]
- Pongsavee, M. Effect of sodium benzoate preservative on micronucleus induction, chromosome break, and Ala40Thr superoxide dismutase gene mutation in lymphocytes. Biomed. Res. Int. 2015, 2015, 103512. [Google Scholar] [CrossRef] [PubMed]
- Yadav, A.; Kumar, A.; Das, M.; Tripathi, A. Sodium benzoate, a food preservative, affects the functional and activation status of splenocytes at non cytotoxic dose. Food Chem. Toxicol. 2016, 88, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Tsay, H.J.; Wang, Y.H.; Chen, W.L.; Huang, M.Y.; Chen, Y.H. Treatment with sodium benzoate leads to malformation of zebrafish larvae. Neurotoxicol. Teratol. 2007, 29, 562–569. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; Tonouchi, H.; Sasayama, A.; Ashida, K. A Ketogenic Formula Prevents Tumor Progression and Cancer Cachexia by Attenuating Systemic Inflammation in Colon 26 Tumor-Bearing Mice. Nutrients 2018, 10, 206. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Kang, C.; Wang, X.; Zhou, M.; Chen, M.; Zhu, X.; Liu, K.; Wang, B.; Zhang, Q.; Zhu, J.; et al. Dietary Factors Modulate Colonic Tumorigenesis Through the Interaction of Gut Microbiota and Host Chloride Channels. Mol. Nutr. Food Res. 2018. [Google Scholar] [CrossRef] [PubMed]
- Grosso, G.; Bela, F.; Godos, J.; Sciacca, S.; Del Rio, D.; Ray, S.; Galvano, F.; Giovannucci, E.L. Possible role of diet in cancer: Systematic review and multiple meta-analyses of dietary patterns, lifestyle factors, and cancer risk. Nutr. Rev. 2017, 75, 405–419. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, A.M.; Burrington, C.M.; Gillaspie, E.A.; Lynch, D.T.; Horsman, M.J.; Grene, M.W. High-fat Western diet-induced obesity contributes to increased tumor growth in mouse models of human colon cancer. Nutr. Res. 2016, 36, 1325–1334. [Google Scholar] [CrossRef] [PubMed]
- World Cancer Report 2014. Available online: http://www.esmo.org/Oncology-News/World-Cancer-Report-2014 (accessed on 15 February 2018).
- Mariani, F.; Sena, P.; Roncucci, L. Inflammatory pathways in the early steps of colorectal cancer development. World J. Gastroenterol. 2014, 20, 9716–9731. [Google Scholar] [CrossRef] [PubMed]
- Zeligs, K.P.; Neuman, M.K.; Annunziata, C.M. Molecular Pathways: The Balance between Cancer and the Immune System Challenges the Therapeutic Specificity of Targeting Nuclear Factor-κB Signaling for Cancer Treatment. Clin. Cancer Res. 2016, 22, 4302–4308. [Google Scholar] [CrossRef] [PubMed]
- Nosrati, N.; Bakovic, M.; Paliyath, G. Molecular Mechanisms and Pathways as Targets for Cancer Prevention and progression with Dietary Compounds. Int. J. Mol. Sci. 2017, 18, 2050. [Google Scholar] [CrossRef] [PubMed]
- Brahmachari, S.; Jana, A.; Pahan, K. NaB, a metabolite of cinnamon and a food additive, reduces microglial and astroglial inflammatory responses. J. Immunol. 2009, 9, 5917–5927. [Google Scholar] [CrossRef] [PubMed]
- Raposa, B.; Pónusz, R.; Gerencsér, G.; Budán, F.; Gyöngyi, Z.; Tibold, A.; Hegyi, D.; Kiss, I.; Koller, Á.; Varjas, T. Food additives: NaB, potassium sorbate, azorubine, and tartrazine modify the expression of NFκB, GADD45α, and MAPK8 genes. Physiol. Int. 2016, 3, 334–343. [Google Scholar] [CrossRef] [PubMed]
- OECD. SIDS Initial Assessment Report for 13th SIDS Initial Assessment Meeting. Benzoates: Benzoic Acid, Sodioum Benzoate, Potassium Benzoate, and Benzyl Alcohol. Available online: https://hpvchemicals.oecd.org/UI/handler.axd?id=aa89d225-a2a7-4ed5-b8d6-c06b5e30b45b (accessed on 15 February 2018).
- SIDS Initial Assessment Report for 13th SIAM. Available online: http://www.inchem.org/documents/sids/sids/BENZOATES.pdf (accessed on 15 February 2018).
- Orlowski, R.Z.; Baldwin, A.S., Jr. NFκB as a therapeutic target in cancer. Trends Mol. Med. 2002, 8, 385–389. [Google Scholar] [CrossRef]
- Maguire, O.; Collins, C.; O’Loughlin, K.; Miecznikowski, J.; Minderman, H. Quantifying nuclear NFκB -p65 as a parameter for NF-κB activation: Correlation between ImageStream cytometry, microscopy, and Western blot. Cytometry A 2011, 6, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.W.; Karin, M.J. A cytokine mediated link between innate immunity, inflammation, and cancer. Clin. Investig. 2007, 117, 1175–1183. [Google Scholar] [CrossRef] [PubMed]
- Biswas, D.K.; Iglehart, J.D. Linkage between EGFR family receptors and nuclear factor kappaB (NFκB) signaling in breast cancer. Cell Physiol. 2006, 3, 645–652. [Google Scholar] [CrossRef] [PubMed]
- Shao, J.; Fujiwara, T.; Kadowaki, Y.; Fukazawa, T.; Waku, T.; Itoshima, T.; Yamatsuji, T.; Nishizaki, M.; Roth, J.A.; Tanaka, N. Overexpression of the wild-type p53 gene inhibits NFκB activity and synergizes with aspirin to induce apoptosis in human colon cancer cells. Oncogene 2000, 19, 726–736. [Google Scholar] [CrossRef] [PubMed]
- Stark, L.A.; Reid, K.; Sansom, O.J.; Din, F.V.; Guichard, S.; Mayer, I.; Jodrell, D.I.; Clarke, A.R.; Dunlop, M.G. Aspirin activates the NFκB signalling pathway and induces apoptosis in intestinal neoplasia in two in vivo models of human colorectal cancer. Carcinogenesis 2007, 5, 968–976. [Google Scholar]
- Parrondo, R.; de las Pozas, A.; Reiner, T.; Rai, P.; Perez-Stable, C. NFκB activation enhances cell death by antimitotic drugs in human prostate cancer cells. Mol. Cancer 2010, 9, 182. [Google Scholar] [CrossRef] [PubMed]
- Mane, S.D.; Thoh, M.; Sharma, D.; Sandur, S.K.; Naidu, K.A. Ascorbyl Stearate Promotes Apoptosis through Intrinsic Mitochondrial Pathway in HeLa Cancer Cells. Anticancer Res. 2016, 12, 6409–6417. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Gu, S.; Jiang, X.; Zhang, Z. Nuclear translocation of nuclear factor kappa B is regulated by G protein signaling pathway in arsenite-induced apoptosis in HBE cell line. Environ. Toxicol. 2016, 12, 1819–1833. [Google Scholar] [CrossRef] [PubMed]
- Kaltschmidt, B.; Kaltschmidt, C.; Hofmann, T.G.; Hehner, S.P.; Dröge, W.; Schmitz, M.L. The pro- or anti-apoptotic function of NFκB is determined by the nature of the apoptotic stimulus. Eur. J. Biochem. 2000, 12, 3828–3835. [Google Scholar] [CrossRef]
- Fan, Y.; Dutta, J.; Gupta, N.; Fan, G.; Gélinas, C. Regulation of programmed cell death by NFκB and its role in tumorigenesis and therapy. Adv. Exp. Med. Biol. 2008, 615, 223–250. [Google Scholar] [PubMed]
- Singhal, J.; Nagaprashantha, L.; Chikara, S.; Awasthi, S.; Horne, D.; Singhal, S.S. 2′-Hydroxyflavanone: A novel strategy for targeting breast cancer. Oncotarget 2017, 43, 75025–75037. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; Aman, J.M.; Tanimoto, A.; Taniguchi, H.; Shukuya, T.; Timmers, C.; Yano, S.; Shilo, K.; Carbone, D.P. Histone Deacetylase Inhibition Enhances the Antitumor Activity of a MEK Inhibitor in Lung Cancer Cells Harboring RAS Mutations. Mol. Cancer Ther. 2018, 17, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.; Isik, E.; Fernandes, S.M.; Brown, J.R.; Letai, A.; Davids, M.S. Bruton’s tyrosine kinase inhibition increases BCL-2 dependence and enhances sensitivity to venetoclax in chronic lymphocytic leukemia. Leukemia 2017, 31, 2075–2084. [Google Scholar] [CrossRef] [PubMed]
- Kowalski-Chauvel, A.; Gouaze-Andersson, V.; Vignolle-Vidoni, A.; Delmas, C.; Toulas, C.; Cohen-Jonathan-Moyal, E.; Seva, C. Targeting progastrin enhances radiosensitization of colorectal cancer cells. Oncotarget 2017, 8, 58587–58600. [Google Scholar] [CrossRef] [PubMed]
- Koç, A.; Ozkan, T.; Karabay, A.Z.; Hekmatshoar, Y.; Gurkan-Alp, A.S.; Aktan, F.; Buyukbingol, Z.; Sunguroglu, A.; Buyukbingol, E. Apoptotic effects of some tetrahydronaphthalene derivatives on K562 human chronic myelogenous leukemia cell line. Anticancer Agents Med. Chem. 2017, 17. [Google Scholar] [CrossRef]
- Ozkan, T.; Hekmatshoar, Y.; Ertan-Bolelli, T.; Hidayat, A.N.; Beksac, M.; Aki-Yalcin, E.; Yalcin, I.; Sunguroglu, A. Determination of the apoptotic effect and molecular docking of benzamide derivative XT5 in K562 cells. Anti-Cancer Agents Med. Chem. 2017. [Google Scholar] [CrossRef] [PubMed]
- Karabay, A.Z.; Koc, A.; Ozkan, T.; Hekmatshoar, Y.; Sunguroglu, A.; Aktan, F.; Buyukbingol, Z. Methylsulfonylmethane Induces p53 Independent Apoptosis in HCT-116 Colon Cancer Cells. Int. J. Mol. Sci. 2016, 17, 1123. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are available from the authors. |








© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yilmaz, B.; Karabay, A.Z. Food Additive Sodium Benzoate (NaB) Activates NF?B and Induces Apoptosis in HCT116 Cells. Molecules 2018, 23, 723. https://doi.org/10.3390/molecules23040723
Yilmaz B, Karabay AZ. Food Additive Sodium Benzoate (NaB) Activates NF?B and Induces Apoptosis in HCT116 Cells. Molecules. 2018; 23(4):723. https://doi.org/10.3390/molecules23040723
Chicago/Turabian StyleYilmaz, Betul, and Arzu Zeynep Karabay. 2018. "Food Additive Sodium Benzoate (NaB) Activates NF?B and Induces Apoptosis in HCT116 Cells" Molecules 23, no. 4: 723. https://doi.org/10.3390/molecules23040723
APA StyleYilmaz, B., & Karabay, A. Z. (2018). Food Additive Sodium Benzoate (NaB) Activates NF?B and Induces Apoptosis in HCT116 Cells. Molecules, 23(4), 723. https://doi.org/10.3390/molecules23040723