Abstract
Facing the increasingly global crisis of antibiotic resistance, it is urgent to develop new antibacterial agents and methods. Simultaneously, as research progresses, the occurrence, development, and treatment of diseases, especially some malignant cancers, are found to be closely associated with the bacterial microenvironment, prompting us to reconsider the efficiency of existing antibacterial strategies for disease treatments. Bacteriophages have been employed as antibacterial agents for an extended period owing to their high biocompatibility and particular targetability toward the host bacterial strains. Nonetheless, they are almost neglected due to their slow and limited efficacy in antibacterial practice, especially in acute and severe infectious cases. In recent years, fantastic advancements in various biochemical technologies, such as bacteriophage display technology, genetic engineering, and chemical molecular engineering, have enabled scientists to conduct a broader range of modifications and transformations on the existing bacteriophages with inherited unique characteristics of themselves. As a result, a series of novel bacteriophage platforms are designed and fabricated with significantly enhanced properties and multiplied functionalities. These offer new avenues for combating infections caused by drug-resistant bacteria and treatment of malignancies that are associated with bacterial infections, holding great significance and potential in the innovative theranostic applications.
1. Introduction
Bacteriophages, also called phages, are viruses with a unique ability to target and infect host bacteria, widely distributed on Earth [1]. Since being identified initially from Staphylococcus and Shigella bacteria in 1915 [2], bacteriophages have played a crucial role as natural antibacterial agents, contributing significantly to the advancement of biological research [3]. There are two major types of phages, i.e., lytic and temperate [4]. Phages are obligate intracellular parasites of hosts and have diverse life cycles. The life cycles include lytic, lysogenic, and pseudolysogenic cycles [5]. Lytic phages induce the lysis of host bacterial cells to release viral progeny [6]. Conversely, lysogenic or temperate phages integrate their nucleic acid (genome) into the host bacterial cell and replicate alongside the host, imparting new characteristics to the host bacteria [7]. Prompted by the growing crisis in antibiotic resistance, phage therapy is experiencing a resurgence as a potent solution against bacterial infections [8]. Although bacteriophages serve as widespread antibacterial agents, their clinical application still encounters a series of severe challenges. For instance, the effectiveness of individual bacteriophages is constrained, and obstacles like biofilms impede their penetration and weaken the antibacterial effectiveness [9]. Moreover, bacteriophages can potentially trigger adverse inflammatory responses [10]. Fortunately, genetic engineering and chemical alterations empower the modification of bacteriophages with enhanced robustness and multifunctionalities [11], expanding their host spectrum and amplifying their therapeutic capacity, presenting hopeful resolutions to these obstacles.
Bacteriophages consist mainly of proteins, forming the outer capsid, and protein-coding nucleic acids, which are protected within the capsid [12]. The protein capsid of bacteriophages contains various amino acids, each contributing distinct active functional groups [13]. Chemical modification techniques of phage rely on the interaction of functional groups on the capsid with active conjugates under specific solution pH and salt conditions [14]. By interacting with active sites like N-terminal alanine, lysine, and tyrosine, bacteriophages can acquire a variety of functionalities [15]. Bacteriophages have been tailored with various functional materials, including inorganic nanoparticles, organic molecules, fluorophores, DNA, antigens, and more, to align with their intended applications. Alternatively, genetic modification approaches entail the introduction of genes encoding amino acids, peptides, or protein sequences into either the bacteriophage genome or the capsid genes on the host plasmid, facilitating the generation of recombinant bacteriophages [16]. Utilizing advanced genome editing methods like recombination, CRISPR-Cas-assisted selection, or synthetic in vitro genome assembly allows for the precise design of particular genes or gene clusters within the bacteriophage genome, enabling the deliberate manipulation of functions such as merging, deleting, modifying, or diversifying [17]. In addition, bacteriophage display technology enables the swift and efficient identification of functional sites involved in protein–protein interactions, receptor binding, epitopes, isotopes, and antigens [18]. Through genetic engineering or chemical modification, bacteriophages acquire multifunctionality, paving the way for novel advancements in synthesizing functional materials, assembling and packing nanostructures, biosensing, biomedical applications, disease diagnosis, and treatment.
These genetic and chemical techniques for altering bacteriophages can be applied to develop diverse bacteriophage-based technologies [19]. These include the following: designing drugs and nanomaterials, synthesizing new proteins, detecting target molecules and pathogenic bacteria, constructing protein libraries, screening peptides or antibodies, treating diseases by killing bacteria, diagnosing diseases, and delivering drugs and genes [20]. Considering the close relationship of the microbiota environment with the occurrence, development, and therapeutics of cancer [21], the new-generation phage-based multifunctional platform has shown great promise in targeted bacterial killing-assisted modulation of the cancer immune environment and reversing cancer chemodrug resistance [22]. Further, bacteriophage-based nanocarriers demonstrate exceptional targeting precision, facilitating the precise delivery of drugs to specific lesion sites, thus enhancing the effectiveness, specificity, and safety of disease treatments, particularly in malignant cancer and brain disease [23]. Engineered bacteriophages can also overcome the challenges related to circulation metabolism and in vivo stability, prolonging their circulation time by evading attacks from the reticuloendothelial system and other bioactive molecules [24]. The gene of phages can be used to develop new DNA vaccines and antigen presentation systems because they can provide a highly organized and repetitive antigen presentation to immune cells, thus holding potential to rejuvenate the adaptive immune systems for combating the cancer cells. Phages allow new possibilities to target the specific molecular biomarkers of cancer cells. Phages can also be used as nanocarriers for delivery of imaging agents and therapeutic drugs/genes for cancer theranostics [25]. This review focused on the exploration of various techniques to engineer bacteriophages and expand their diverse applications across various domains, including templated synthesis of functional nanomaterials, guided assembly of nanostructures, bioanalytes assay, targeted bacterial elimination, assisted disease therapeutics, drug/gene delivery, etc., offering perspectives on the advancement of engineered bacteriophage-based technologies (Figure 1). Engineered phages that are chemically or genetically modified can be specifically localized at lesion sites (e.g., tumor tissue) that are infected by symbiotic target bacteria, which can not only contribute to their application in in vivo imaging but also serve for therapeutics by delivering gene/drug and act as immunomodulators to regulate the tumor microenvironment, thus achieving inhibition effects in the growth of tumors.
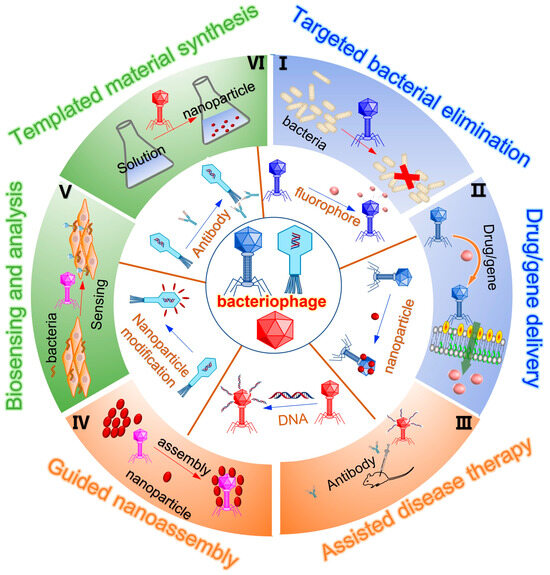
Figure 1.
Overview of bacteriophage engineering for functional material synthesis, bioanalytical sensing, and disease theranostics. Diverse bacteriophages can undergo chemical modifications to incorporate functional materials like inorganic nanoparticles, organic polymer chains, fluorophore molecules, RNA/DNA, antibodies, etc. Meanwhile, they can undergo genetic engineering to integrate foreign peptides or proteins into their capsid proteins and undergo DNA recombination to aid the production of engineered bacteriophages. These engineered bacteriophages, with their multifunctionality, find applications in diverse fields such as templated nanoparticle synthesis, guided nanostructural assembly, bioanalyte assay, identification and eradication of pathogenic bacteria, assisted disease diagnosis and treatment, as well as drug and gene delivery.
2. The Modification of Bacteriophage
Bacteriophages serve as nano-sized probes for finely tuned disease diagnostics and as therapeutic agents for targeted treatment modalities. Given their bacteria-specific nature, phages refrain from infecting eukaryotic cells, thus ensuring safety. Moreover, they can be genetically engineered to home in on nanoparticles, cells, tissues, and organs, and can even be endowed with functional non-biological nanomaterials to enhance their diagnostic and therapeutic capabilities [26]. In recent years, extensive research endeavors have been undertaken to leverage phages as platforms in nanomedicine, capitalizing on their distinctive biological attributes. The versatility of phages as multifunctional platforms hinges significantly on the functionalization of their coat proteins with diverse functionalities. While genetic manipulation of these proteins stands out as a prominent approach for such functionalization, complementary chemical modification strategies also contribute to expanding the repertoire of materials, enabling precise, site-specific incorporation of multiple functionalities onto the phage [27].
2.1. Chemical Modification of Phage
Chemical modification stands as a pivotal approach in reshaping the structure and enhancing the functionality of bacteriophages. Within the proteinaceous capsids of these phages lie an array of amino acids, each offering diverse reactive functional groups encompassing carboxylic acids, amines, phenols, and thiols, presenting a rich landscape for molecular manipulation [28]. The abundance of nucleophilic functional groups further broadens the scope, enabling multiple amino acids to engage in chemical transformations. Through interactions with reactive sites including N-terminal alanine, lysine, aspartic/glutamic acid, cysteine, N-terminal serine/threonine, and tyrosine, phages can be endowed with an array of functionalities [29], ranging from DNA strands and antibodies to nanoparticles, fluorophores, and pharmaceutical agents, opening new avenues across biosensing, biomedicine, disease diagnosis, and treatment [30]. Especially, the phage can carry and deliver genes or drugs for the treatment of malignant tumors after chemical modification.
To precisely recognize, adhere, and kill bacteria at the infection sites, Mao et al. developed an innovative nano-enzyme-armed bacteriophage system, designated as phage@palladium (Pd), for combating bacterial infections (Figure 2a) [31]. The proposed phage@Pd retains the functionality of bacteriophages to achieve precise recognition and adhesion to host Escherichia coli. Meanwhile, the ultra-small Pd nano-enzymes with pronounced pH-dependent peroxidase-like activity can generate toxic hydroxyl radicals around bacteria in acidic and hydrogen peroxide-overexpressing infectious microenvironments. In contrast, under physiological conditions, the Pd nanoenzymes remained inert, leading to significant bacterial clearance at infection sites while excellent biocompatibility in healthy tissues. Additionally, phage@Pd can not only eradicate planktonic bacteria but also kill bacteria within biofilms in vitro. In in vivo models of acute bacterial pneumonia or subcutaneous abscesses, phage@Pd demonstrates significant activity in eliminating infections and promoting tissue recovery, suggesting a safe and effective antibacterial agent. In another example, Peng et al. coupled engineered chimeric M13 phage with photothermal-active gold nanorods and realized ablation of diverse bacterial strains [32]. Meanwhile, Wang et al. prepared a photocatalytic quantum-dot-equipped phage, abbreviated as QD@phage, showing excellent performance in combating infections caused by Pseudomonas aeruginosa [33].
Given the widespread use of filamentous bacteriophages as platforms for peptide and protein displaying, the ability to attach a large number of synthetic functional groups to their capsid proteins is of significant value to improve the performance of bacteriophages. Francis’s group further decorated bacteriophages with polymer chains and imaging moieties (Figure 2b) [34]. After converting the N-terminal amines of the capsid proteins to ketones, further chemically specific modification via oxime formation with alkoxyamine groups can be achieved, as fluorescent moieties and up to 3000 polyethylene glycol molecules were attached on each bacteriophage capsid without significantly affecting the binding efficiency of bacteriophage-displayed antibody fragments to EGFR and HER2 receptors. The chemically modified fluorescent dyes, such as Alexa Fluor 488 and 647 C5-aminooxyacetamide (AF488/647-ONH2), can be easily substituted by imaging, MRI, PET, and other detection agents, allowing for various in vitro and in vivo biomedical applications. Additionally, filamentous bacteriophage M13 with N-terminal ketone can serve as an effective supramolecular scaffold for the assembly of multivalent CryA, a hyperpolarized 129Xe contrast agent composed of many assembled latent A molecular cages (Figure 2c) [35]. The saturation of a large number of latent cages with xenon gas is chemically exchanged to a water-soluble xenon pool, leading to the effective generation of ultra-chemical exchange saturation transfer (CEST) contrast. The detection limit of this bacteriophage-based ultra-CEST agent can be as low as 230 fM, which represents the optimal single-dose sensitivity for magnetic resonance imaging agents.
As nucleic acid aptamers can serve as specific biotarget-recognizable ligands, an efficient oxidative coupling strategy was developed by Tong et al. to attach them to the surface of non-genomic bacteriophage capsid carriers (Figure 2d) [36]. Up to 60 DNA strands can be attached to each capsid without significant loss of base-pairing ability or protein stability. The specific binding capability was demonstrated by attaching aptamers that target tyrosine kinase receptors expressed on Jurkat T cells. With fluorescent dyes capsulated inside the capsid, the level of binding between phage capsids and Jurkat T cells was significantly enhanced. Confocal microscopy indicated that capsids were internalized and transported to lysosomes, suggesting the potential of that aptamer-modified bacteriophage in the application of targeted delivery and controlled release of cargos in living cells at lesion sites.
The bacteriophage T4 capsid is adorned with 155 copies of Hoc, a non-essential highly antigenic outer capsid protein. To induce effective immune responses and provide protection in the viral infection, Rao’s group reported an in vitro assembly system that allows the surface display of human immunodeficiency virus (HIV) antigens, p24-gag, on the bacteriophage T4 capsid through Hoc–capsid interactions (Figure 2e) [37]. Fusion within the plasmid framework was constructed by splicing HIV genes to the 5′ or 3′ end of the Hoc gene. The Hoc fusion proteins were expressed, purified, assembled, and displayed on bacteriophage particles in the defined E. coli system. Single or multiple antigens were efficiently displayed, saturating all available capsid binding sites. In the absence of any external adjuvants, the displayed p24 demonstrated high immunogenicity in mice, eliciting strong p24-specific antibodies as well as Th1 and Th2 cell responses, providing new directions and insights for HIV vaccine development with the potential to broaden both cellular and humoral immune responses.
Phages have been modified by a range of functional moieties (e.g., fluorophores, nanoparticles, functional nucleic acids, antigens, and drugs) to serve their applications. The protein on the phage capsid can be coupled with functional molecules and materials by electrostatic adsorption or chemical reaction under appropriate pH values and ionic conditions to obtain chemically modified multifunctional phages. Modifying phage capsid protein is a promising method to obtain multifunctional phage. Flexible and diverse chemical modification strategies have expanded phage-based biomedical applications (Table 1).

Figure 2.
Chemical modification with diverse structure for reshaping the structures of bacteriophages. (a) The Pd nanocubes modified M13 phage (phage@Pd) for H2O2 catalytical hydrolysis and antibacterial study through the targeting and infection capabilities of phage toward the host bacteria [31]. Reproduced with permission from [31]; published by the Wiley Online Library, 2023. (b) Chemical modification of filamentous phage via transamination reaction. The N-termini were converted to ketone-bearing proteins, which were then reacted with aminooxy-functionalized fluorophores and aminooxy-functionalized PEG2k [34]. Reproduced with permission from [34]; published by the American Chemical Society, 2012. (c) Chemical modification of M13 phage. The N-termini of p8 proteins were transaminated to ketones along the capsids, which were then reacted with PEG-5k–ONH2 chains and aminooxy-functionalized cryptophane-A cages (CryA–ONH2) to construct M13-based biosensors [35]. Reproduced with permission from [35]; published by the Wiley Online Library, 2013. (d) Dual-surface modification of MS2 phage. For interior surfaces, an N87C mutation on the coat protein allowed for site-specific alkylation with up to 180 cargo molecules uploaded. For the exterior surface, a T19paF mutation on the capsid allowed for the attachment of the phenylene diamine group-modified aptamer via a NaIO4-mediated oxidative coupling reaction [36]. Reproduced with permission from [36]; published by the American Chemical Society, 2009. (e) Antigen spikes artificially fused to Hoc subunits. The left reconstruction showed blue spikes representing the single antigen display, and the right reconstruction showed blue, green, and pink spikes representing three antigen displays [37]. Reproduced with permission from [37]; published by the American Society for Microbiology, 2006.
Figure 2.
Chemical modification with diverse structure for reshaping the structures of bacteriophages. (a) The Pd nanocubes modified M13 phage (phage@Pd) for H2O2 catalytical hydrolysis and antibacterial study through the targeting and infection capabilities of phage toward the host bacteria [31]. Reproduced with permission from [31]; published by the Wiley Online Library, 2023. (b) Chemical modification of filamentous phage via transamination reaction. The N-termini were converted to ketone-bearing proteins, which were then reacted with aminooxy-functionalized fluorophores and aminooxy-functionalized PEG2k [34]. Reproduced with permission from [34]; published by the American Chemical Society, 2012. (c) Chemical modification of M13 phage. The N-termini of p8 proteins were transaminated to ketones along the capsids, which were then reacted with PEG-5k–ONH2 chains and aminooxy-functionalized cryptophane-A cages (CryA–ONH2) to construct M13-based biosensors [35]. Reproduced with permission from [35]; published by the Wiley Online Library, 2013. (d) Dual-surface modification of MS2 phage. For interior surfaces, an N87C mutation on the coat protein allowed for site-specific alkylation with up to 180 cargo molecules uploaded. For the exterior surface, a T19paF mutation on the capsid allowed for the attachment of the phenylene diamine group-modified aptamer via a NaIO4-mediated oxidative coupling reaction [36]. Reproduced with permission from [36]; published by the American Chemical Society, 2009. (e) Antigen spikes artificially fused to Hoc subunits. The left reconstruction showed blue spikes representing the single antigen display, and the right reconstruction showed blue, green, and pink spikes representing three antigen displays [37]. Reproduced with permission from [37]; published by the American Society for Microbiology, 2006.


Table 1.
The modification of bacteriophage for application.
Table 1.
The modification of bacteriophage for application.
| Modification Strategies | Modified Materials | Functionalization | Application | Reference |
|---|---|---|---|---|
| Amino-carboxyl group reaction | Pd nanocubes | Catalytic hydrolysis | Antibacterial study | Jin et al., 2023 [31] |
| N-succinimidyl-S-acetylthiopropionate (SATP) | Gold nanorods (AuNRs) | Photothermal | Antibacterial study | Peng et al., 2020 [32] |
| Avidin-biotin bioconjugation | Cd-based quantum dot | Photocatalytic localized reactive oxygen species (ROS) | Antibacterial study | Wang et al., 2022 [33] |
| N-termini reacted with aminooxy-functionalized fluorophores | Fluorophores and 2 kDa PEG | Fluorescence imaging | Characterization of breast cancer cells | Carrico et al., 2012 [34] |
| Site-specific protein bioconjugation | CryA cages | Hyperpolarized xenon MR contrast agent | NMR/MRI imaging | Stevens et al., 2012 [35] |
| Phenylene diamine-substituted oligonucleotides with aniline groups | Nucleic acid aptamers | Bind specific cellular targets | Targeted drug delivery of acid-labile prodrugs | Tong et al., 2009 [36] |
| Hoc–capsid interactions | HIV antigens | Highly immunogenic | HIV vaccine | Sathaliyawala et al., 2006 [37] |
2.2. Genetic Modification of Phage
The distinguishing advantage of phages over other synthetic biomaterials lies in their capacity to present proteins or peptides on specific coat proteins through genetic modification [38]. Genetic engineering techniques involve the introduction of genes encoding alternative amino acids, peptides, or protein sequences into either the bacteriophage genomes or host plasmids containing capsid genes, facilitating the generation of recombinant phages via employing the well-established “phage display” method [26,39]. These recombinant capsid proteins then create additional sites for modification by enzymes or chemicals. For accommodating large, complex foreign protein fusions, recombinant capsid decoration proteins can be further assembled in vitro onto phages with accessible binding sites [40,41].
One method for engineering phages is homologous recombination (HR), which is a process involving the exchange of nucleotide sequences between two similar or identical molecules of double-stranded DNA (dsDNA) or single-stranded RNA (ssRNA) [42]. It serves crucial roles in DNA repair, DNA replication, and telomere maintenance. Regulated by the enzyme DNA helicase, HR has the capability to exchange up to 23 base pairs between two homologous DNA sequences, making it valuable for engineering phages within their bacterial hosts. This mechanism has been utilized for the introduction of foreign genes into phages (Figure 3a) [43]. Gene insertion, deletion, and substitution in phages through HR follow the same principles as bacterial recombination. In the development of recombinant phages, the desired gene sequence is initially incorporated into a replicative plasmid, followed by homologous integration of the desired phage sequences. Similarly, for engineering phage particles, bacterium containing donor plasmids is infected with phages, leading to the packaging of heterogeneous genes within phage particles [44].
The CRISPR-Cas system, found naturally in many prokaryotes, serves as an adaptive defense mechanism against invasive nucleic acids [45]. Comprising Cas proteins and the CRISPR array, this system has garnered attention for its potential applications. For instance, the type I-E CRISPR-Cas system has recently been employed as a counter-selection mechanism by Lv et al. for engineered T7 phages (Figure 3b) [46]. Through homologous recombination-mediated editing, the phage successfully excised the dispensable gene 1.7. This approach effectively removed nonrecombinant phage genomes containing gene 1.7 while preserving recombinant phage genomes lacking this gene. Additionally, the CRISPR/Cas II-A system can facilitate in vivo editing of the phage 2972 genome, enabling modifications such as point mutations, gene deletions, and DNA exchange [47]. With these promising genetic manipulations, the CRISPR/Cas strategy shows potential for broader applications across various phage genomes. The application of CRISPR-Cas-based techniques for phage engineering is limited to bacteria with either a native CRISPR-Cas system or the capacity for genetic transformation to express a functional heterologous CRISPR-Cas system. This limitation significantly hampers the manipulation of bacteriophages targeting bacteria lacking genetic manipulability [48].
Before introducing them into host bacterial cells, the genome of phages can undergo manipulation and modification in vitro. To eliminate overlaps between genetic segments, the refactoring process has been applied to T7 phages. In one study, 73 segments were divided into six sections using bracketing restriction sites to allow for changes within each section independently, without affecting others (Figure 3c) [43]. Subsequently, the refactoring of genomes after transformation into bacteria yielded a chimeric T7 phage genome composed of various engineered sections combined with viable phages. However, recombinant phages generated through the refactoring method displayed significantly smaller plaques compared to wild-type phages. Moreover, the refactoring approach to phage engineering necessitates large-scale DNA manipulation in vitro and modification of bacterial hosts with engineered genomes to retrieve viable phages. Consequently, this process yields only a limited number of engineered phages, particularly when applied to non-domesticated bacterial hosts.
The proliferation of phages within bacterial hosts can sometimes result in toxicity, leading to reduced efficiency of homologous recombination (HR), in vivo recombineering, and bacteriophage recombineering with electroporated DNA(BRED) genetic engineering techniques [49]. To address this issue, an intermediate host such as Saccharomyces cerevisiae can be employed, playing a vital role in genetic manipulation. In the approach of Ramirez-Chamorro’s group, phages remain stable and non-toxic within yeast cells (Figure 3d) [43]. In the process of producing engineered phages, the genome of S. cerevisiae is utilized to capture the phage genome using a bacterial shuttle vector containing overhangs for recombination of the phage genome and vector. Subsequently, the newly assembled, modified, and propagated phage genomes in yeast are isolated and introduced into the host bacterium to recover recombinant phage particles. A similar strategy from Pires’s group was employed in the creation of Pseudomonas aeruginosa phage vB_PaeP_PE3, where yeast-based assembly was utilized to delete approximately 48% of the hypothetical gene protein. During this process, the phage genome was PCR-amplified, excluding hypothetical protein sequences, and assembled in yeast before being isolated and incorporated into the host bacteria to recover engineered phages. Employing this approach, engineered phages can be assembled by deleting surplus parts of their genome through the incorporation of desired sequences [50,51].
Genetic modifications that introduce genes into the phage genome or capsid genes to replace amino acid, peptide, or protein sequences on the host plasmid can promote the production of recombinant phages. These recombinant capsid proteins provide additional sites for enzymes or chemicals to react with the capsid.

Figure 3.
Genetic engineering techniques for modifying phages. (a) Developing engineered phages through homologous gene recombination. Phages 1 and 2 infect the same bacterial host, which results in the recombination of their genomes and forms new recombinant phages [43]. Reproduced with permission from [43]; published by Elsevier, ScienceDirect, 2023. (b) Phage genome engineering with CRISPR-Cas13a. The CRISPR-Cas13a selectively eliminated the harboring gene in the nonrecombinant phage genomes while sparing the recombinant phage genomes devoid of this gene [46]. Reproduced with permission from [46]; published by MDPI, 2023. (c) Rebuilding or refactoring the phage genome in vitro. The independent pieces of phage DNA via digesting the native restriction sites of purified phage DNA were further manipulated by sub-cloning. After release from the plasmid, the recombinant DNA section was ligated with the rest of the phage DNA and delivered to the host bacteria via electroporation, leading to the recovery of engineered phages [43]. Reproduced with permission from [43]; published by Elsevier, ScienceDirect, 2023. (d) Phage genome engineering by the yeast-based assembly. Phage genomic DNA is used as a template for the amplification of overlapped PCR products, except for the knockout region. For assembly of the genome, the genomic fragments are co-transformed to yeast cells along with a linearized YAC plasmid, and the phage genome is transformed into P. aeruginosa cells for the generation of engineered phages [43]. Reproduced with permission from [43]; published by Elsevier, ScienceDirect, 2023.
Figure 3.
Genetic engineering techniques for modifying phages. (a) Developing engineered phages through homologous gene recombination. Phages 1 and 2 infect the same bacterial host, which results in the recombination of their genomes and forms new recombinant phages [43]. Reproduced with permission from [43]; published by Elsevier, ScienceDirect, 2023. (b) Phage genome engineering with CRISPR-Cas13a. The CRISPR-Cas13a selectively eliminated the harboring gene in the nonrecombinant phage genomes while sparing the recombinant phage genomes devoid of this gene [46]. Reproduced with permission from [46]; published by MDPI, 2023. (c) Rebuilding or refactoring the phage genome in vitro. The independent pieces of phage DNA via digesting the native restriction sites of purified phage DNA were further manipulated by sub-cloning. After release from the plasmid, the recombinant DNA section was ligated with the rest of the phage DNA and delivered to the host bacteria via electroporation, leading to the recovery of engineered phages [43]. Reproduced with permission from [43]; published by Elsevier, ScienceDirect, 2023. (d) Phage genome engineering by the yeast-based assembly. Phage genomic DNA is used as a template for the amplification of overlapped PCR products, except for the knockout region. For assembly of the genome, the genomic fragments are co-transformed to yeast cells along with a linearized YAC plasmid, and the phage genome is transformed into P. aeruginosa cells for the generation of engineered phages [43]. Reproduced with permission from [43]; published by Elsevier, ScienceDirect, 2023.

3. Phage Mediated Synthesis and Assembly
3.1. Phage Mediated Nanomaterial Synthesis
In the last decade, biotemplates, such as bacteriophages, have emerged as powerful tools for synthesizing inorganic materials [52]. Bacteriophages can undergo genetic modification to incorporate specific material-recognizing peptides, which can induce the nucleation of materials, primarily crystals, from an aqueous precursor solution onto the phage surface under mild conditions [53]. By employing phages of diverse shapes, controlled architectures of inorganic materials can be achieved, including one-dimensional nanowires, two-dimensional nanofilms, and three-dimensional scaffolds [54]. Phages serve as versatile building blocks, capable of organizing nanomaterials with various properties. Moreover, beyond utilizing the inherent structure of phages themselves, genetic engineering enables the display of peptides that confer affinity characteristics, serving as bio-interaction motifs or forming self-assembled nanomaterials [55]. Particularly, specific recognition peptides identified through phage library have been harnessed to template the growth of a wide array of inorganic nanocrystals and nanomaterials. By fusing these crystal-nucleating peptides onto multiple phage coat proteins, an assortment of intriguing nanomaterials and nanostructures can be constructed [56].
Mao et al. reported a phage-based scaffold for synthesizing semiconductor and magnetic materials including single-crystal ZnS, CdS, and chemically ordered CoPt and FePt nanowires (Figure 4a) [57]. Peptides exhibiting composition, size, and phase control during nanoparticle nucleation processes were expressed on the highly ordered filamentous coat of evolutionarily selected M13 bacteriophage. Annealing to remove the phage template promotes the growth of oriented crystalline aggregates, forming individual crystal nanowires. This was the first example to introduce the unique ability of substrate-specific peptides into the linear self-assembled filamentous structure of M13 phage for inorganic nanomaterial synthesis.
Ferroelectric materials, like tetragonal barium titanate (BaTiO3), are widely utilized in various fields such as bioimaging, biosensing, and high-power switching devices. However, the conventional synthesis methods for tetragonal phase BaTiO3 typically involve toxic organic reagents and high-temperature treatments, rendering them environmentally unfriendly and energy-inefficient. Therefore, Mao’s group harnessed the phage display technique to devise a novel approach for producing BaTiO3 nanowires [58]. As shown in Figure 4b, through biopanning of a phage-displayed random peptide library, a short BaTiO3-binding/nucleating peptide, CRGATPMSC (named RS), was identified and genetically fused to the major coat protein (pVIII) of filamentous M13 phages to create pVIII-RS phages. These modified phages not only exhibited capability in binding with pre-synthesized BaTiO3 crystals but also inducing the nucleation of uniform tetragonal BaTiO3 nanocrystals at room temperature without the need for toxic reagents, resulting in one-dimensional polycrystalline BaTiO3 nanowires. This method offers a sustainable approach for the eco-friendly synthesis of BaTiO3 polycrystalline nanowires.
Besides nanowires being synthesized through bacteriophages, they were utilized to produce complex 3D structures of nanoparticles (NPs). M13 bacteriophage derivatives that display a ZnO-binding peptide (TMGANLGLKWPV) on either the pIII or pVIII coat protein were engineered and employed as biotemplates by Piotr Golec’s team (Figure 4c) [59]. Depending on the type of recombinant phage used (M13-pIII-ZnO or M13-pVIII-ZnO), well-separated ZnO NPs or complex 3D structures of ZnO NPs with sizes of approximately 20–40 nm were synthesized at room temperature. The synthesized ZnO nanoparticles exhibited luminescence, emitting light near the short wavelength end of the visible region (around 400 nm). The presence of a very low-intensity emission band at 530 nm indicated that the ZnO material obtained had a low concentration of surface defects.
Besides these reports that focused on single modifications of the pIII minor coat protein to synthesize ZnS and CdS nanoparticles at the tip of the bacteriophage, Nam et al. modified both of the end minor coat proteins (pIII and pIX) with six histidine and HPQ (His–Pro–Gln) streptavidin binding peptide motifs [60]. Subsequently, these engineered phages were employed to fabricate nanoscale viral ring structures by leveraging the specific binding between chemically conjugated streptavidin and Ni-NTA linkers. Through expression of semiconductor and metal-binding peptides on the major coat proteins of phages, high-quality single crystal monodisperse ZnS nanowires with promising electro-optical and magnetic properties were produced through annealing processes. By simultaneously modifying pIII and pVIII coat proteins, a variety of nanostructures were successfully produced via the template of bacteriophage (Figure 4d) [61].
The unique recognition peptides of phages can direct mineralization processes for the synthesis of many useful nanoscale electronic and medical materials. The monodispersed and long rod-shaped phages, with specific recognition patterns, enable the synthesis of various nanomaterials such as ZnO, ZnS, and CdS quantum dots, which can be used in electronic, optical, and biotechnological applications.

Figure 4.
Phage-mediated nanomaterial synthesis. (a) The nucleation, ordering, and annealing of phage–nanoparticle assembly including II–VI semiconductors ZnS and CdS and ferromagnetic CoPt and FePt nanowire on M13 phage. The capsid and ends of M13 phages were genetically modified by encoding gP3, gP8, and gP9 peptides. The resulted symmetric phage allowed for ordering of the nucleated particles along the x, y, and z directions with the aid of the expressed peptides with rigidity and packing pattern [57]. Reproduced with permission from [57]; published by Science, 2004. (b) Using engineered phages for the synthesis of BaTiO3 polycrystalline nanowires at room temperature. BaTiO3-binding peptide was selected via biopanning and construction of the pVIII-RS phage via displaying of BaTiO3-binding peptides on the side wall of phage, which can guide the nucleation of BaTiO3 on phage to form polycrystalline nanowires [58]. Reproduced with permission from [58]; published by the American Chemical Society, 2016. (c) The modified phages acted as machinery for the synthesis of both the previously selected and identified ZnO-binding peptide and ZnO nanoparticles. The exposure of a ZnO-binding peptide to a phage mediated the growth of unique ZnO nanostructures [59]. Reproduced with permission from [59]; published by the American Chemical Society, 2016. (d) Genetic modification of the phage for programmed synthesis of ZnS nanoparticles, nanowires, viral rings, and arrayed hetero-nanostructures by engineered phages with display of pIII coat protein, pVIII coat protein, pIII/pIX dual-protein, and pIII/pVIII dual-protein, respectively [61]. Reproduced with permission from [61]; published by the American Chemical Society, 2005.
Figure 4.
Phage-mediated nanomaterial synthesis. (a) The nucleation, ordering, and annealing of phage–nanoparticle assembly including II–VI semiconductors ZnS and CdS and ferromagnetic CoPt and FePt nanowire on M13 phage. The capsid and ends of M13 phages were genetically modified by encoding gP3, gP8, and gP9 peptides. The resulted symmetric phage allowed for ordering of the nucleated particles along the x, y, and z directions with the aid of the expressed peptides with rigidity and packing pattern [57]. Reproduced with permission from [57]; published by Science, 2004. (b) Using engineered phages for the synthesis of BaTiO3 polycrystalline nanowires at room temperature. BaTiO3-binding peptide was selected via biopanning and construction of the pVIII-RS phage via displaying of BaTiO3-binding peptides on the side wall of phage, which can guide the nucleation of BaTiO3 on phage to form polycrystalline nanowires [58]. Reproduced with permission from [58]; published by the American Chemical Society, 2016. (c) The modified phages acted as machinery for the synthesis of both the previously selected and identified ZnO-binding peptide and ZnO nanoparticles. The exposure of a ZnO-binding peptide to a phage mediated the growth of unique ZnO nanostructures [59]. Reproduced with permission from [59]; published by the American Chemical Society, 2016. (d) Genetic modification of the phage for programmed synthesis of ZnS nanoparticles, nanowires, viral rings, and arrayed hetero-nanostructures by engineered phages with display of pIII coat protein, pVIII coat protein, pIII/pIX dual-protein, and pIII/pVIII dual-protein, respectively [61]. Reproduced with permission from [61]; published by the American Chemical Society, 2005.

3.2. Phage-Guided Nanostructural Assembly
Previously, it was demonstrated that displaying semiconductor-binding peptides on phages can facilitate the production of single-crystalline semiconductor nanowires. This approach typically involves three key steps. Firstly, a target-binding peptide is identified through biopanning. Secondly, the peptide is genetically presented on the side wall of the phage. Finally, the resulting phage serves as a biotemplate for the assembly of nanoparticles [62]. Due to the capability of genetically fusing foreign peptides to bacteriophage coat proteins for generation of highly decorated phages and the significant influence of surface charges on bacteriophage assembly behavior, bacteriophages emerge as promising templates for controlling the assembly of nanostructures [63]. At each end of the phage nanocrystalline wires, specific receptors can be incorporated as fusions to terminal proteins. By programming the self-assembly of these constructs, complex nanowire arrays with diverse one-, two-, and three-dimensional geometries can be meticulously engineered.
Mao’s group has introduced a system harnessing the M13 phage coats to nucleate and template II–VI semiconductor nanocrystals into well-oriented quantum dot nanowires (Figure 5a) [64]. The pVIII major coat protein was genetically engineered to showcase evolutionarily selected peptides, facilitating precise control over nanocrystal growth into phage–semiconductor hybrid nanowires with crystallographic alignment spanning micrometer scales. By incorporating semiconductor nucleating peptides onto the capsids of M13 phages, directed and phase-specific nanocrystal growth was achieved. This method seamlessly integrates the use of naturally ordered and assembled phage templates with the versatility of genetic manipulation for material synthesis, enabling the controllable assembly of semiconductor wires within supramolecular hybrid structures.
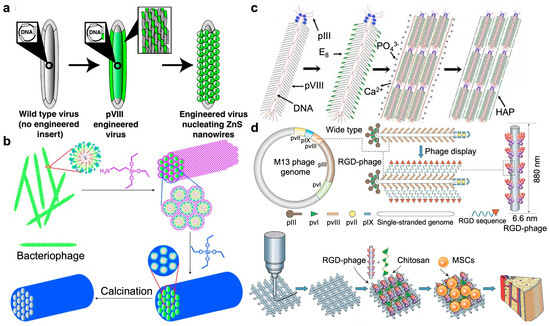
Figure 5.
Phage-guided nanostructural assembly. (a) Directed ZnS nanocrystal synthesis on A7–pVIII-engineered phages at 0 °C [64]. Reproduced with permission from [64]; published by the National Academy of Sciences, 2003. (b) Templated silica nanostructure formation via monodispersed filamentous phages that self-assembled into highly ordered hexagonal lattices. Removal of the phage assembly through calcination led to the formation of mesoporous silica fibers with pore structures precisely defined by the phage assembly [65]. Reproduced with permission from [65]; published by the Wiley Online Library, 2012. (c) Controlled assembly of hydroxylapatite (HAP) on the wild-type M13 phage. Genetic fusion of E8 peptide to major coat protein (pVIII) on the side wall. Self-assembly of anionic phage into a nanofibrous structure (bundle) in the presence of Ca2+ ions. Mineralization of the phage bundle to form a fiber [66]. Reproduced with permission from [66]; published by the Wiley Online Library, 2010. (d) Bone generation via genetically modified M13 phage. RGD peptide was fused to the solvent-exposed terminal of the major coat protein (pVIII), constituting the side wall of filamentous phages. RGD-phage nanofibers (negatively charged) were integrated into a 3D printed bioceramic scaffold along with chitosan (positively charged). The resultant scaffold was seeded with MSCs and then implanted into bone defects, inducing the formation of new bone [67]. Reproduced with permission from [67]; published by the Wiley Online Library, 2014.
Later, the same group devised a versatile approach leveraging the surface proteins of biotemplates to catalyze the hydrolysis of organosilicate precursors and the ensuing polycondensation process to yield silica (Figure 5b) [65]. Meticulously organized mesoporous silica fibers featuring hexagonally arranged pores were fabricated by utilizing fd phage as biotemplate. By exhibiting peptides with varying charges on the side wall of filamentous bacteriophage, silica nanostructures were successfully controlled through manipulation of both bacteriophage surface charge density and the accompanying bacteriophage assembly behavior. Furthermore, the resultant mesoporous silica has been successfully utilized as a medium for fabricating three-dimensional arrays of PbS nanoparticles.
Building upon this foundation, Mao further expanded research efforts for genetic modification of phages. The filamentous M13 phage was anionized through the fusion of a negatively charged peptide to its major coat protein (pVIII) [66]. In the presence of cationic precursors like Ca2+ ions, the negatively charged phage underwent self-assembly into a nanofiber that promoted the formation of oriented nanocrystalline hydroxyapatite (HAP) (Figure 5c). In co-assembly, collagen fostered the formation of a hierarchical architecture akin to the extracellular matrix (ECM), while E8-displayed phage enhanced matrix mineralization. The arrangement and alignment of collagen and HAP dictated the superior mechanical properties of bone, holding promise for developing bone-mimetic biomaterials and bone tissue engineering.
Further, the same group introduced a novel VAM (Vascularized Bone Augmentation Matrix) strategy aimed at enhancing the formation of vascularized bone (Figure 5d) [67]. Through the incorporation of RGD-phage, the VAM exhibits the ability to regulate endothelial cell (EC) migration and adhesion, thereby promoting endothelialization. Simultaneously, it triggers osteoblastic differentiation of MSCs, consequently facilitating osteogenesis and angiogenesis in vivo. The critical role of engineered phage nanofibers, specifically RGD-phage, in stimulating both angiogenesis and osteogenesis marks a significant breakthrough in phage-based nanomedicine and regenerative medicine. The specific recognition capabilities of phages can guide the growth of inorganic semiconductors and bone-like biomaterials and facilitate osteogenic formation. Such functionalized phages can self-assemble to form highly organized hierarchies.
4. Bacteriophage in Precise Bioanalytes Assay
Molecular diagnostics endeavors to identify specific biomarkers for disease diagnosis, monitoring, risk assessment, and treatment determination. Although conventional enzyme-linked immunosorbent assays (ELISAs) and polymerase chain reaction (PCR) offer standard measurement techniques, the integration of nanotechnology into molecular diagnostics in recent years has also yielded numerous novel diagnostic approaches [68]. The nanoscale-sized phages, with their DNA enclosed within a protein capsid, present an opportunity for genetic engineering to display multiple peptides on the sidewall (pVIII) and tip (pIII) of the protein capsid [69]. Consequently, they have been extensively engineered to express nanobodies or mimotopes for selective recognition of target analytes or to interact with functional nanoparticles for serving as specific nanoprobes [70]. Thus, modified phage nanoprobes have great potential in enhancing the accuracy and reliability of diagnostic schemes. It can be expected that phages modified with specific peptides or functional materials can accurately target tumors and detect tumor biomarkers.
Bruce D. Hammock et al. introduced a new approach to immune-polymerase chain reaction (IPCR) known as phage anti-immunocomplex assay real-time PCR (PHAIA-PCR) for detecting small analytes, including 3-phenoxybenzoic acid and molinate (Figure 6a) [71]. The technique employed phage anti-immunocomplex assay technology, where a short peptide was presented on the M13 bacteriophage’s surface specifically bound to the antibody–analyte complex. As the phage DNA can function as a versatile material for PCR, both universal amplification that targets the phagemid’s common element and specific amplification when the real-time PCR probe is designed to bind the phagemid DNA can be achieved, leading to the significantly improved sensitivity. A tenfold increase in sensitivity was achieved via this PHAIA-PCRs method compared to traditional PHAIA. The successful assay conducted using agricultural drain water and human urine samples further demonstrated its reliability for monitoring human exposure or environmental contamination.
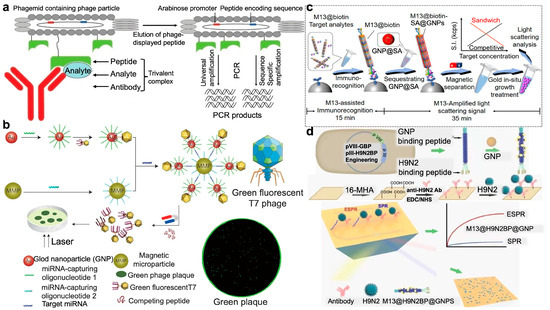
Figure 6.
Modified phage for the selective recognition and analysis of biotargets. (a) Gene detection via PHAIA-PCR technology. The M13 phage with peptide binding to an immunocomplex of an antibody–analyte was eluted and immediately neutralized for PCR detection [71]. Reproduced with permission from [71]; published by the American Chemical Society, 2011. (b) A naked-eye-counting method for miRNA quantification with sensitivity comparable to PCR. The magnetic microparticle and the T7 phage–gold nanoparticle (GNP) binding probe were linked by cohybridization with target miRNA, forming a sandwich complex with miRNA number equal to the phage number. After magnetic separation, T7 phages were released from the GNPs by adding more competitive gold-binding peptides. Counting the number of fluorescent plaques formed by T7 phages by eye can quantify the miRNA [72]. Reproduced with permission from [72]; published by Springer Nature, 2015. (c) Immunoassay strategy via integrating designed M13 phage-assisted recognition with the in situ growth of gold nanoparticle-based light scattering signal for ultrasensitive quantitative detection of low-abundance ochratoxin A and alpha-fetoprotein analytes [73]. Reproduced with permission from [73]; published by the American Chemical Society, 2023. (d) Using phage@GNP nanofibers to detect H9N2 virus. M13 with H9N2 and GNP binding peptides can attach with GNP onto the sidewall to form the M13@H9N2BP@GNP nanofiber probe, showing an enhanced surface plasmon resonance (SPR) signal upon binding with H9N2 targets on the phage tail. SPR signals can be cross-validated with the blue-color phage plaques that were naked-eye discernable [74]. Reproduced with permission from [74]; published by Elsevier, ScienceDirect, 2023.
Counting disease biomarkers like miRNAs with the naked eye is very difficult in molecular diagnostics. Gu and Mao reported a highly sensitive method for quantifying miRNAs using T7 phage, which was modified to fluoresce and bound to a DNA-modified gold nanoparticle (GNP) in a one-to-one fashion after genetic engineering (Figure 6b) [72]. Target miRNAs then crosslinked the resulting phage–GNP complex with DNA-modified magnetic microparticles, forming a sandwich complex containing equimolar amounts of phage and miRNA. After release from the complex, the phages were grown into a macroscopic fluorescent plaque for visually counting. Thus, miRNA targets can be sensitively and conveniently quantified using only the naked eye, with detection limits down to aM level for miRNA biomarkers.
Traditional immunoassay-based detection methods face limitations due to the size effect of nanoprobes, which hinders the balance between molecular recognition and signal amplification. Tang’s group presented a versatile approach by integrating immunoassay with a light-scattering signal of gold nanoparticles that were in situ grown on the M13 phage surface for quantitative detection of low-abundance analytes (Figure 6c) [73]. In the practical detection of ochratoxin A and alpha-fetoprotein, a detection limit of sub-femtomolar levels in real samples was achieved, showcasing four orders of magnitude enhancement in sensitivity compared to traditional phage-based ELISA methods. With the implementation of a biotin-streptavidin amplification scheme, further enhancement in sensitivity was realized, allowing the detection of severe acute respiratory syndrome coronavirus 2 spike protein down to the attomolar range.
Rapid and sensitive detection of pathogenic viruses is crucial for pandemic control. Hou et al. reported a swift and highly sensitive optical biosensing method for detecting avian influenza virus H9N2 (Figure 6d), utilizing a genetically modified filamentous M13 phage probe, M13@H9N2BP@AuBP, that borne an H9N2-binding peptide (H9N2BP) at its tip and a gold nanoparticle (AuNP)-binding peptide (AuBP) on its sidewall [74]. Simulation modeling demonstrated that the electric field in surface plasmon resonance (SPR) enhanced by 40-fold compared to conventional AuNPs, guaranteeing the experimentally ultrasensitive detection of H9N2 particles with a detection limit down to 6.3 copies/mL. It was even more sensitive than the quantitative polymerase chain reaction (qPCR). Further, after capturing H9N2 viruses on the sensor chip, the H9N2-binding phage nanofibers can be quantitatively converted into visible plaques, allowing for quantification through naked-eye observation. This phage-based biosensing strategy can be easily extended to the detection of other pathogens just by replacing the H9N2-binding peptides with other pathogen-binding peptides using phage display technology.
Phages have been developed as emerging nanoprobes for the detection of various biomarkers and pathogens. After displaying arbitrary sequences of exogenous peptides or antibodies, phages can specifically bind to targets, such as animal cells, bacteria, viruses, and protein molecules. Phage can also be co-modified with metal nanoparticles to detect targets by showing colorimetrical, fluorescence, and plasmonic signals.
5. Bacteriophages in Bacterial Recognition and Detection
Conventional bacterial detection primarily relies on bacterial culture, which is cumbersome and time-consuming. PCR technology and ELISA-based immunoassay offer enhanced sensitivity and efficiency in bacterial detection yet necessitate specialized instruments and skilled personnel [75]. Recently, biosensors employing specific molecular recognition agents to target bacteria have emerged as promising alternatives. Phages utilize receptor-binding proteins at the distal end of the tail to recognize and interact with bacterial surface receptors such as lipopolysaccharides, teichoic acids, and porins. This unique ability allows phages to capture specific host bacteria from complex matrices [76]. More importantly, phage-based detection schemes offer the advantage of distinguishing between live and inactivated bacteria as phages replicate exclusively within viable hosts. This feature represents an improvement over DNA- or antibody-based detection assays, which may yield false positives if the pathogen has been successfully inactivated but its genetic material or epitopes remain detectable [77,78]. Thus, increasing studies have explored the utilization of phage-based schemes for the accurate and reliable detection of pathogenic bacteria.
To develop an inexpensive, rapid, and reliable method for detection of E. coli in drinking water or food, Chen et al. reported a phage-based magnetic separation technique for the swift detection of E. coli in drinking water as the broad host range specificity of T7 phage toward E. coli (Figure 7a) [79]. After capturing and isolating E. coli BL21 from the drinking water sample, phage-mediated lysis can liberate intrinsic β-galactosidase from the adhered bacteria, which can convert the yellow-color chlorophenol red-β-D-galactopyranoside to a red color. E. coli can be successfully detected within 2.5 h with a detection limit down to 1 × 104 CFU·mL–1. This colorimetric change can be visually discerned, facilitating a straightforward and rapid quantification of target bacteria, particularly advantageous in resource-limited settings. By incorporating a pre-enrichment step in Luria–Bertani meida supplemented with isopropyl β-D-thiogalactopyranoside, even 10 CFU·mL–1 E. coli in drinking water can be detected after 6 h of pre-enrichment.
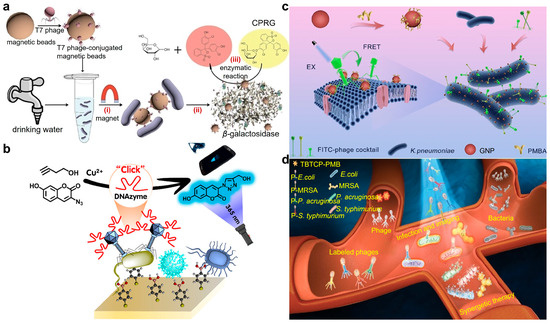
Figure 7.
Phage for bacterial recognition and detection. (a) Detection of E. coli in drinking water using a T7 bacteriophage-conjugated magnetic probe. The separated E. coli released β-gal that can catalyze chlorophenol red-β-D-galactopyranoside (CPRG) hydrolysis to produce a colorimetric signal for readout [79]. Reproduced with permission from [79]; published by the American Chemical Society, 2015. (b) M13 phage@DNAzyme fluorescent probe for portable and ultrasensitive detection of bacteria. After selective conjugation with target E. coli O157:H7, the probe can efficiently catalyze the click reaction between 3-azido-7-hydroxycoumarin (AHC) and 3-butyn-1-ol (BOL) to generate a fluorescent product in the presence of Cu(II) [80]. Reproduced with permission from [80]; published by the American Chemical Society, 2023. (c) A phage cocktail composed of three phages for wide-spectrum detection of K. pneumoniae strains. Both the FITC-labeled phage cocktail and PMBA-modified GNPs were employed to conjugate K. pneumoniae to generate a bacterial complex as they recognized the different sites of the bacterial surface. In the present of K. pneumoniae, a decreased FL signal can be observed as a fluorescence resonance energy transfer (FRET) effect occurred between GNP and FITC, enabling the quantification of target bacteria [81]. Reproduced with permission from [81]; published by the American Chemical Society, 2023. (d) AIE-photosensitizer modified phage cocktails for the early diagnosis of sepsis. Phages were modified by AIE-active TBTCP-PMB through a simple nucleophilic substitution reaction between benzyl bromide functional groups and the sulfhydryl functional groups of proteins on the phage. The perfect and simultaneously maintained advantages of phages and PSs enabled the selective identification of bacterial species in 30 min through fluorescence imaging [82]. Reproduced with permission from [82]; published by the Wiley Online Library, 2023.
The prompt and precise identification of virulent foodborne pathogens is crucial for ensuring food safety. Gan’s group introduces an ultra-sensitive and specific phage@DNAzyme probe for the detection of foodborne pathogens (Figure 7b) [80]. The carefully selected phage and functionalized DNAzyme enabled specific recognition of target foodborne pathogens at the strain level and efficient catalysis of copper(II)-based azide-alkyne cycloaddition (CuAAC) click reaction with fluorescent signal, respectively. Using virulent E. coli O157:H7 as the representative analyte, the phage@DNAzyme probes specifically bound to the captured E. coli O157:H7 and facilitated the click reaction to generate a visible fluorescent signal that can be quantified using a smartphone. A wide linear range from 102 to 108 CFU/mL was demonstrated with a detection limit of 50 CFU/mL, offering a novel approach for enhancing food safety.
Since single phages often encounter limitations in bacterial detection due to the high strain specificity of individual phages, Fu et al. prepared a cocktail comprising three Klebsiella pneumoniae (K. pneumoniae) phages to broaden the detection spectrum (Figure 7c) [81]. A total of 155 clinically isolated K. pneumoniae strains were used to evaluate the recognition spectrum. The recognition rate of 42.3–62.2% using a single phage can be significantly increased to 91.6% via phage cocktail, highlighting the complementarity of the recognition capabilities. Leveraging the wide-spectrum recognition capability of the phage cocktail, a fluorescence resonance energy transfer method was further established through labeling the phage cocktail with fluorescein isothiocyanate and Au nanoparticles with p-mercaptophenylboronic acid as energy donors and acceptors, respectively. The detection process for K. pneumoniae strains could be completed within 35 min, with a broad dynamic range of 5.0 × 102–1.0 × 107 CFU/mL, offering promising avenues for enhancing bacterial detection methodologies.
By introducing highly sensitive fluorescence imaging functionality, Gu et al. proposed a novel approach for detecting pathogenic bacteria via covalently conjugating the aggregation-induced emission photosensitizer (AIE-PS) TBTCP-PMB with a phage cocktail (Figure 7d) [82]. The host specificity of phages, the fluorescence imaging capability, and reactive oxygen species (ROS) of AIE-PS were synergized for bacteria detection and sterilization alongside favorable biocompatibility. Additionally, this approach enables rapid diagnosis of blood infections in clinical samples, offering a straightforward and precise method for pathogen detection and point-of-care diagnosis.
Rapid and reliable bacterial tests are essential for reducing the socio-economic burden associated with bacterial infections. Bacteriophages have high specificity to their host bacteria and can identify target bacteria quickly and accurately. Whether it is a single phage to recognize a specific genus of bacteria or a phage cocktail to identify a mixture of bacteria, it reflects the great potential of phage in bacterial detection. Thus, the particular specificity of phages and their widespread availability make them ideal tools for developing biosensors and bioassays for bacterial detection.
6. Bacteriophages in Targeted Bacterial Killing
Efficient Bacterial infections have long posed significant threats to public health, contributing to severe illness and persistently high mortality rates worldwide [83]. Since the discovery of penicillin, antibiotics have been the cornerstone of bacterial infection treatment [84]. However, the global misuse and overuse of antibiotics have fueled bacterial evolution, leading to the emergence of antibiotic-resistant strains. Phages that can selectively infect bacteria and cause lysis offer a promising alternative to antibiotics. They exhibit remarkable specificity towards their hosts and can adapt and evolve synchronously to infect even resistant bacteria [85]. Consequently, phages are increasingly recognized as a potent tool for antimicrobial therapy. Despite their potential, the efficacy of phages in combating infectious diseases, especially acute infections, is hindered by their relatively low bactericidal stability efficiency. To address this limitation, phages have been modified to enhance their functionality, enabling them to target and eradicate specific bacterial strains more effectively [86].
In light of the escalating global antibiotic resistance crisis, there is an urgent demand for novel agents exhibiting specific targeting toward bacterial strains and exceptional antibacterial efficacy. To address this challenge, Tang et al. introduce a groundbreaking approach that combines bacteriophage (PAP) with photodynamic inactivation (PDI)-active AIEgens (luminogens with aggregation-induced emission property) (Figure 8a) [87]. This strategy yields a unique AIE–PAP bioconjugate characterized by superior capabilities for targeted imaging and synergistic eradication of select bacterial species with maintained infection activity of the phage. Crucially, the PDI-active AIEgens function as potent in situ photosensitizers, generating high-efficiency reactive oxygen species (ROS) upon white light irradiation. Consequently, selective targeting and synergistic elimination of both antibiotic-sensitive and multi-drug-resistant (MDR) bacteria were achieved with excellent biocompatibility. This pioneering AIE–phage integrated approach diversifies the current arsenal of antibacterial agents and promises to catalyze the development of potent drug candidates for the future.
The misuse of antibiotics has spurred the rise of bacterial resistance, prompting the exploration of alternative treatments. Photodynamic antibacterial chemotherapy (PACT) holds promise but faces challenges such as inefficient ROS generation and lack of bacterial targeting. In response, Peng et al. reported a novel photodynamic antimicrobial agent (APNB) based on a cationic photosensitizer (NB) and bacteriophage (ABP) for precise bacterial eradication and effective biofilm ablation (Figure 8b) [88]. Through structural modification with a sulfur atom, NB exhibits enhanced ROS-production capability and near-infrared fluorescence. Furthermore, bacteriophages confer specific binding to pathogenic microorganisms and combinational therapeutic functionalities (PACT and phage therapy). Both in vitro and in vivo studies confirm APNB’s efficacy against A. baumannii infection, with faster recovery compared to conventional treatments like ampicillin and polymyxin B in vivo. Additionally, the combination of bacteriophages and photosensitizers showed promising results in eradicating bacterial biofilms for the first time, offering a practical solution to bacterial infections.
Intracellular bacterial infections pose significant threats to human health, evading immune defenses and displaying notable drug resistance, presenting formidable challenges for treatment. He and Tang developed a versatile living phage nanoconjugate, integrating aggregation-induced emission luminogen (AIEgen) photosensitizers and nucleic acids onto a bacteriophage framework (forming MS2-DNA-AIEgen bioconjugates) (Figure 8c) [89]. These nanoconjugates efficiently penetrate mammalian cells, specifically identifying intracellular bacteria while emitting a detectable fluorescent signal. Leveraging the photodynamic properties of AIEgen photosensitizers and the bacteriophage’s innate targeting and lysis abilities, effectively eliminate intracellular bacteria and restore infected cell activity. Furthermore, the engineered phage nanoconjugates accelerate wound healing in bacterially infected wounds observed in diabetic mouse models, enhancing immune activity within infected cells and in vivo, without evident toxicity. These multifunctional phage nanoconjugates employ AIEgen photosensitizers and spherical nucleic acids as a pioneering strategy for combating intracellular bacteria and offering potent avenues for theranostics applications in diseases associated with intracellular bacterial infections.
Increasing research results show that the occurrence and development of malignant tumors are closely related to the symbiotic bacteria in the tumor microenvironment. The treatment of tumor-associated bacteria by phages has become a promising avenue to augment the cancer therapy [90]. Emerging evidence underscores the role of gut microbiota in colorectal cancer (CRC) tumorigenesis, with symbiotic Fusobacterium nucleatum (Fn) selectively boosting immunosuppressive myeloid-derived suppressor cells (MDSCs), thereby impairing the host’s anticancer immune response. A Fn-targeting M13 phage was identified by Zhang’s group via phage display technology (Figure 8d) [91]. Subsequently, silver nanoparticles (AgNP) were electrostatically assembled on its surface capsid protein (forming M13@Ag) to achieve precise clearance of Fn and reshape the tumor-immune microenvironment. Further, antigen-presenting cells (APCs) can be activated to bolster the host immune system’s response against CRC. When combined with immune checkpoint inhibitors (α-PD1) or chemotherapeutics (FOLFIRI), M13@Ag significantly extended overall survival in an orthotopic CRC model.
The engineered bacteriophage overcomes the limited killing ability of nude bacteriophage and achieves an augmented bactericidal effect assisted by photodynamic, photoacoustic, and photothermal effects. Whether it is sensitive or drug-resistant bacteria or even intracellular bacterial infections, excellent antibacterial performance can be achieved through engineered phages.

Figure 8.
Modified phages to target and eradicate specific bacteria. (a) AIE photosensitizer modified phage for guided targeting, discriminative imaging, and synergistic killing of bacteria. Phage possessed particularly targetability toward the host bacteria that can be lit up by the AIE fluorescence and subsequently killed by the synergized effect of phage infection and photodynamic activity of AIE photosensitizers [87]. Reproduced with permission from [87]; published by the American Chemical Society, 2020. (b) The multi-functional antibacterial system (APNB) based on the ABP phage and Nile blue photosensitizer (NB) for the treatment of multi-drug-resistant A. baumannii and its biofilms [88]. Reproduced with permission from [88]; published by the Royal Society of Chemistry, 2021. (c) MS2-DNA-AIEgen bioconjugate for specific targeting and synergistic elimination of intracellular bacteria. After DNA modification on the surface, the MS2 phage was transferred to a type of spherical nucleic acid that can efficiently enter the infected immune cells and find the target bacteria. The fluorescence property and photodynamic activity of AIE photosensitizers can subsequently label and eliminate the intracellular bacteria [89]. Reproduced with permission from [89]; published by the American Chemical Society, 2024. (d) Phage-based bio/abiotic hybrid system (M13@Ag) to regulate gut microbes for cancer-specific immune therapy. Silver (Ag) nanoparticles were electrostatically assembled on the surface of M13 phages (M13@Ag). After accumulation in the tumor microenvironment, AgNP can selectively kill the protumoral F. nucleatum under the guidance of M13 phages, which possessed exquisite recognition capability toward F. nucleatum and blocked the recruitment of immunosuppressive cells [91]. Reproduced with permission from [91]; published by Science, 2020.
Figure 8.
Modified phages to target and eradicate specific bacteria. (a) AIE photosensitizer modified phage for guided targeting, discriminative imaging, and synergistic killing of bacteria. Phage possessed particularly targetability toward the host bacteria that can be lit up by the AIE fluorescence and subsequently killed by the synergized effect of phage infection and photodynamic activity of AIE photosensitizers [87]. Reproduced with permission from [87]; published by the American Chemical Society, 2020. (b) The multi-functional antibacterial system (APNB) based on the ABP phage and Nile blue photosensitizer (NB) for the treatment of multi-drug-resistant A. baumannii and its biofilms [88]. Reproduced with permission from [88]; published by the Royal Society of Chemistry, 2021. (c) MS2-DNA-AIEgen bioconjugate for specific targeting and synergistic elimination of intracellular bacteria. After DNA modification on the surface, the MS2 phage was transferred to a type of spherical nucleic acid that can efficiently enter the infected immune cells and find the target bacteria. The fluorescence property and photodynamic activity of AIE photosensitizers can subsequently label and eliminate the intracellular bacteria [89]. Reproduced with permission from [89]; published by the American Chemical Society, 2024. (d) Phage-based bio/abiotic hybrid system (M13@Ag) to regulate gut microbes for cancer-specific immune therapy. Silver (Ag) nanoparticles were electrostatically assembled on the surface of M13 phages (M13@Ag). After accumulation in the tumor microenvironment, AgNP can selectively kill the protumoral F. nucleatum under the guidance of M13 phages, which possessed exquisite recognition capability toward F. nucleatum and blocked the recruitment of immunosuppressive cells [91]. Reproduced with permission from [91]; published by Science, 2020.

7. Bacteriophage-Assisted Chemo/Radio/Immunotherapy
Emerging research highlights the significant role of the microbiota in both the development and progression of various types of cancer (e.g., colorectal cancers (CRCs) that exist in the gut with abundant bacteria). It has been demonstrated that bacteria can induce severe drug resistance in cancer chemotherapy and foster the immunosuppressive tumor microenvironment (TME) that impedes the immune cell responses [92,93]. While antibiotics have been proposed to regulate the microbiota, their broad spectrum of targets and negative impact on immunotherapy limit their efficacy. Alternatively, phages offer a promising approach to rebalance the tumor-immune microenvironment through precise manipulation of the gut microbiota and activation of innate immune cells [94]. In clinical practice, conventional cancer treatment modalities such as surgery, radiotherapy (RT), chemotherapy, endocrinotherapy, and targeted therapy are commonly employed. However, these approaches may induce resistance and severe side effects [95]. The integration of phage technology in cancer therapy presents a promising avenue for identifying tumor-targeting agents, carriers, and potential vaccines. Natural microorganisms containing pathogen-associated molecular patterns (PAMPs) effectively stimulate the host’s innate immune response, surpassing synthetic material-based vaccines in robustness and efficacy [96].
The gut microbiota in humans is closely associated with the progression of colorectal cancer (CRC) and its response to treatment. By harnessing the higher concentration of pro-tumorigenic Fusobacterium nucleatum and the deficiency of anti-tumorigenic butyrate-producing bacteria in the fecal microbiota of CRC patients, Zhang’s team discovered that oral or intravenous administration of irinotecan-loaded glucan nanoparticles with modification of bacteriophages targeting Fusobacterium nucleatum can effectively inhibit the growth of Fusobacterium nucleatum and significantly enhance the efficacy of chemotherapy for CRC in mice (Figure 9a) [97]. Additionally, the impact of bacteriophage-guided irinotecan nanoparticles on blood cell counts, immunoglobulin, and tissue protein levels, as well as liver and kidney function in piglets, was negligible. Utilizing this bacteriophage-guided nanotechnology to modulate the gut microbiota may inspire new approaches for treating colorectal cancer.
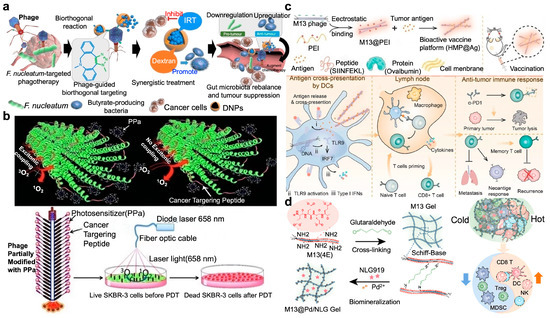
Figure 9.
Phage-assisted chemo/radio/immunotherapy. (a) The phage-guided biotic–abiotic hybrid nanosystem for CRC therapeutics. Through bioorthogonal reaction, chemodrug IRT-loaded dextran nanoparticles (IDNPs) were bound to the phages with the capability to target CRC and eliminate the pro-tumoral Fusobacterium and enhance the production of butyrate from anti-tumoral bacteria. The inhibition of pro-tumoral bacteria and the promotion of anti-tumoral bacteria together augmented the chemotherapeutic efficacy [97]. Reproduced with permission from [97]; published by Springer Nature, 2019. (b) Phage conjugated with photosensitizer for targeted photodynamic therapy (PDT). With modification of the major coat protein (pVIII) with PPa photosensitizer, the filamentous phage fd-tet nanowire was used as a vehicle for targeted delivery of photosensitizer to the SKBR-3 cells to realize cancer eradication [98]. Reproduced with permission from [98]; published by the Wiley Online Library, 2013. (c) Phage-based vaccine combined with immune checkpoint blockade (ICB) treatment for inhibition of primary and metastatic cancers. Tumor antigen (Ag) modified hybrid M13 phage (HMP@Ag)-based vaccine was internalized by DCs for antigen release and cross-presentation to promote DCs maturation, which further migrated to lymph nodes for activation and expansion of antigen-specific CD8+ T cells, representing a remarkable tumor recurrence suppression effect after surgery [99]. Reproduced with permission from [99]; published by Elsevier, ScienceDirect, 2022. (d) Construction of M13 phage-based self-adjuvant photothermal gel for antitumor therapy. The engineered M13 phage was chemically cross-linked through the Schiff base reaction to biomineralize Pd nanoparticles and load the inhibitor NLG919 of immunosuppressive factor indoleamine 2,3-dioxygenase 1 (IDO1), realizing photothermal and immunotherapy synergistically for robust antitumor response [100]. Reproduced with permission from [100]; published by the American Chemical Society, 2023.
Mao et al. conjugated a photosensitizer to a phage for specifically targeting SKBR-3 breast cancer cells and consequently PDT via fusing a unique targeting peptide (VSSTQDFP) to each of the ~3900 copies of pVIII on the side wall of the phage (Figure 9b) [98]. A well-established photosensitizer, 9-ethenyl-14-ethyl-4,6,13,18-tetramethyl-20-oxo-3-phorbinepropanoic acid, commonly known as pyropheophorbid-a (PPa), was then partially modified on the N-terminus of the phage surface. Thus, the photosensitizers can be transported to the intended site, as the targeting specificity of the phage nanowire was well maintained through the free N terminus of the pVIII that is not conjugated to PPa. These phage-conjugated photosensitizers expand the clinical repertoire of target-specific PDT.
Zhang’s group demonstrated the effectiveness of phages in immunotherapy via a bioactive vaccine platform termed HMP@Ag, which was composed by hybrid M13 phages and individualized tumor antigens (Figure 9c) [99]. The transportation of antigens to lymph nodes was facilitated, and the antigen-presenting cells (APCs) were activated via the Toll-like receptor 9 (TLR9) signaling pathway, leading to enhanced innate and adaptive immune responses. Serving as an adjuvant platform, hybrid M13 phages can easily deliver a variety of tumor-specific antigens by simple adsorption, thereby supporting the development of personalized cancer vaccines. Significantly, the HMP@Ag vaccine not only prevents tumor formation but also retards tumor growth in established (subcutaneous and orthotopic) and metastatic tumor-bearing models, particularly when used in combination with immune checkpoint blockade (ICB) therapy, with dramatically suppressed post-operative recurrence and concurrently promotes long-term immune memory. Furthermore, HMP@Ag induces a robust immune response targeting neoantigens in models of tumor-specific mutations, suggesting a potent tool for developing a bio-activated hybrid platform for personalized therapy.
The complexity and diversity within the tumor microenvironment have posed significant challenges to the efficacy of cancer treatments. Zhang et al. utilized an engineered M13 bacteriophage with immunogenic activity to remodel the tumor microenvironment (Figure 9d) [100]. Through chemical cross-linking and biomineralization, a living phage system capable of transforming the tumor microenvironment was fabricated. Through biomineralization, photothermal-active palladium nanoparticles (PdNPs) were in situ synthesized on the pVIII capsid protein of M13 phage. After chemical cross-linking and loading of NLG919 that aimed to inhibit the expression of the tryptophan metabolic enzyme indoleamine 2,3-dioxygenase 1 (IDO1), M13@Pd/NLG gel was formed. M13 bacteriophage not only acted as a carrier for cargo but also functioned as a self-immune adjuvant, effectively inducing immunogenic cell death in tumor cells and down-regulating IDO1 expression with good potential to reverse immunosuppression and significantly enhance the anti-breast cancer response.
The above studies confirm the efficacy and safety of using phage-based vectors as delivery tools for therapeutic genes and drugs in cancer therapy. Compared to the non-viral gene transfer methods, engineered phages show much higher efficiencies in gene delivery and expression in cancer cells with enhanced selectivity. However, as the interaction mechanisms between phage and eukaryotic cells are not illustrated clearly, it is necessary to carry out more in-depth investigations for further cancer diagnosis and treatment.
8. Bacteriophages for Drug/Gene Delivery
As nanomedicine advances, an array of carriers has emerged for targeted drug delivery, including liposomes, dendrimers, polymers, micelles, virus-like particles, and even stem cells [101]. Phages stand out among these carriers due to their non-infectivity to mammalian cells, small genome size, simple structure, and ease of engineering. In addition, phage display technology offers an efficient strategy to modify phages by displaying target-specific peptides [102]. Moreover, in chemical drug delivery, filamentous phages offer superior pharmacokinetics and delivery efficiency compared to spherical nanoparticles as they can accommodate a larger payload of chemical drugs. For gene delivery, foreign genes can be inserted into the phage genome or loaded onto phage-mimetic nanoparticles. Thus, phages emerge as excellent candidates for drug and gene delivery applications [103].
With advancements in covalent modification techniques for phage capsids, Francis et al. can transform them into versatile carrier systems for drugs and imaging agents (Figure 10a) [104]. Two distinct modification strategies were employed to adorn the outer surface of genome-free MS2 capsids with PEG chains while incorporating 50–70 fluorescent dye inside to mimic drug cargo. Despite extensive modification, the capsids maintained their assembled state. The efficacy of the PEG coating in obstructing access to the capsid surface by polyclonal antibodies was as high as 90%. Biotin groups positioned at the distal ends of the polymer chains retained the ability to bind to streptavidin. Additionally, a modular approach was developed for attaching small-molecule targeting moieties to the PEG chains via an efficient oxime formation reaction, facilitating the development of robust and versatile platforms for therapeutic cargo delivery.
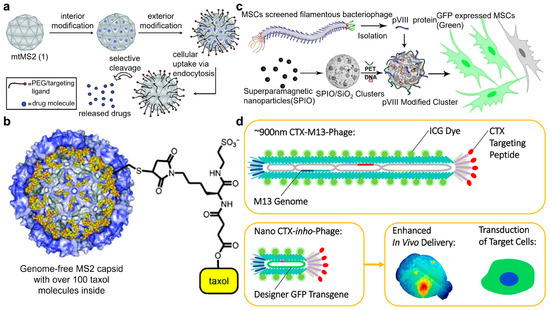
Figure 10.
Phage for drug/gene delivery. (a) Construction of multifunctional drug delivery vessels derived from genome-free (mtMS2) phage capsids. Utilizing orthogonal coupling strategies to append over 180 PEG chains to the exterior surface of phage MS2, while decorating its interior surface with 50–70 fluorescein molecules as drug cargo mimic. The loaded drug can be efficiently released after cell uptake [104]. Reproduced with permission from [104]; published by the American Chemical Society, 2007. (b) Drug delivery system with a water-soluble derivative of the chemotherapeutic agent taxol conjugated to the capsids of MS2 phages. An amino acid on the interior of MS2 capsid was mutated to a cysteine residue (N87C) to provide a sulfhydryl group for the attachment of cargo. The modified capsids remained in their native form and released taxol when incubated with MCF-7 cells [105]. Reproduced with permission from [105]; published by the Wiley Online Library, 2009. (c) Phage capsids for gene delivery to mesenchymal stem cells (MSCs). The gene cargo was incorporated into the porous structure of SPIO-embedded magnetic/silica nanoclusters (MSNCs) that was modified with PEI and pVIII proteins extracted from the MSC-targeting phage. Bearing pVIII allowed the nanoclusters to mimic phage particles and target MSCs to achieve gene delivery [106]. Reproduced with permission from [106]; published by the Wiley Online Library, 2013. (d) CTX-M13 phage for gene delivery. After p3 capsid chlorotoxin fusion with the “inho” circular single-stranded DNA (cssDNA) gene packaging system, a miniatured chlorotoxin inho (CTX-inho) phage particle was produced that can target the GBM22 glioblastoma tumors in the brains of mice, achieving enhanced efficiency in gene delivery and target cell transduction [107]. Reproduced with permission from [107]; published by the American Chemical Society, 2022.
Wu et al. attached the water-soluble derivative of the chemotherapy drug paclitaxel to the capsid of MS2 phages via covalently bioconjugation (Figure 10b) [105]. Moreover, 180 copies of the MS2 protein monomer aggregate to create a hollow spherical carrier with a diameter of 27 nm and 32 pores of 2 nm wide that can facilitate the incorporation of small drug molecules onto the interior surface. Degradation and nonspecific interactions of the cargo with healthy tissue can be well avoided. Up to 65% modification of the MS2 monomer proteins was achieved, corresponding to approximately 110 molecules of taxol per capsid, or roughly 2% loading by weight. The modified capsid retains its capsid form and can efficiently release paclitaxel with excellent cytotoxicity toward the tested MCF-7 cancer cells.
Non-viral gene therapy toward mesenchymal stem cells (MSCs) holds promise for treating various diseases, but MSCs are difficult to transfect using non-viral vectors. Mao’s group has successfully engineered phage-borne rMSC-targeting pVIII proteins on the surface of gene-carrying fd-tet phage, resulting in the formation of virus-mimetic magnetic/silica nanoclusters (VMSNCs) (Figure 10c) [106]. These VMSNCs achieved a significantly improved transfection efficiency of approximately 40% in rMSCs compared to commercial transfection reagents. Furthermore, incorporating rMSC-targeting peptides significantly enhanced transfection efficiency (up to approximately 50%) with sustained gene expression for over 170 h, indicating enhanced stem cell-based gene therapy.
The M13 bacteriophage is a highly adaptable and genetically customizable nanocarrier for disease diagnosis and therapeutics. By fusing the p3 capsid with chlorotoxin and incorporating the “inho” circular single-stranded DNA (cssDNA) gene, Belcher’s group successfully engineered miniature chlorotoxin inho (CTX-inho) phage particles with a length of at least 50 nm (Figure 10d) [107]. These particles can selectively target intracranial orthotopic patient-derived GBM22 glioblastoma tumors in mouse brains. Upon systemic administration, indocyanine green-conjugated CTX-inho phage accumulates in brain tumors by emitting out infrared fluorescence signals. Additionally, the transported cssDNA remained transcriptionally active when delivered to GBM22 glioma cells in vitro. Taking together the ability to adjust capsid display, surface loading, phage length, and cssDNA gene content, the recombinant M13 phage particle can be engineered to be an optimal delivery platform.
Chemical drugs can be modified on the phage surface, and gene drugs can also be inserted into the phage genome by recombinant DNA technology. Meanwhile, specific peptides/proteins displayed on the phage surface are able to bind to functional materials, giving them specific targeting and huge drug loading capabilities. What is more, phage peptides or proteins can be directly self-assembled into nanovehicles for self-guided drug delivery.
9. Conclusions and Prospectives
Although numerous approaches have demonstrated potential in modifying and armoring bacteriophages that bring considerable strides in the applications of bioanalysis and disease theranostics, certain constraints persist and imped their continuous advancement and utilization. First, the screening process for phages is relatively complicated, time-consuming, and labor-intensive, leading to the lack of a standardized phage library. Second, long-term examination of the genetic stability of the engineered bacteriophages that acquired through genetic modification and transformation is necessary. Despite the encouraging outcomes of applying the existing generation of engineered bacteriophages, the durability of their genetic alterations and inheritability of properties and functions are remaining unresolved problems. Thirdly, safety is the pivotal issue in the real clinical translation. Apart from a few experiments that have demonstrated the safety of engineered bacteriophages in vitro and in cell-level, persistent challenges still impede the progress of applying them in animal-level practices. These obstacles necessitate us to gain a deeper understanding of the immune response of modified bacteriophages and carry out thorough evaluations of the potential risks linked to their in vivo and clinic applications. Consequently, further research is imperative to probe the interactions between bacteriophages and the bodies, aiming to clearly illustrate the stability and degradability of engineered bacteriophages in diverse environments.
In sum, this review outlines the capabilities of chemical modification and genetic engineering to create engineered bacteriophages with enhanced properties and multiplied functionalities. Accompanying the advancements in phage display technology and molecular engineering, armored bacteriophages are extensively utilized across diverse fields. This review summarizes the outstanding achievements of engineered bacteriophages in recent years, including the synthesis and assembly of nanomaterials, detection of biotargets and pathogenic bacteria, screening of peptides or antibodies, identification and elimination of bacteria, diagnosis and treatment of diseases, as well as drug and gene delivery, offering promising avenues for future clinical research endeavors.
Author Contributions
Conceptualization, X.H.; resources, J.Z.; writing—original draft preparation, J.Z.; writing—review and editing, X.H.; supervision, X.H.; funding acquisition, X.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China, grant numbers 22274106 and 22104104; the Natural Science Foundation of Jiangsu Province, grant number BK20210701; and the Program of Suzhou Innovation and Entrepreneurship Leading Talents grant number ZXL2022513. The APC was funded by Soochow University.
Acknowledgments
The authors acknowledge funding from the National Natural Science Foundation of China (22274106 and 22104104), the Natural Science Foundation of Jiangsu Province (BK20210701), the Program of Suzhou Innovation and Entrepreneurship Leading Talents (ZXL2022513), and startup funds from Soochow University.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Hendrix, R.W. Bacteriophages: Evolution of the majority. Theor. Popul. Biol. 2002, 61, 471–480. [Google Scholar] [CrossRef] [PubMed]
- Twort, F.W. Further investigations on the nature of ultra-microscopic viruses and their cultivation. J. Hyg. 1936, 36, 204–235. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Chatterjee, S.; Datta, S.; Prasad, R.; Dubey, D.; Prasad, R.K.; Vairale, M.G. Bacteriophages and its applications: An overview. Folia Microbiol. 2017, 62, 17–55. [Google Scholar] [CrossRef] [PubMed]
- Yue, H.; Li, Y.; Yang, M.; Mao, C. T7 phage as an emerging nanobiomaterial with genetically tunable target specificity. Adv. Sci. 2022, 9, 2103645. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhang, T.; Yu, M.; Chen, Y.-L.; Jin, M. The life cycle transitions of temperate phages: Regulating factors and potential ecological implications. Viruses 2022, 14, 1904. [Google Scholar] [CrossRef] [PubMed]
- Young, R.; Wang, I.-N.; Roof, W.D. Phages will out: Strategies of host cell lysis. Trends Microbiol. 2000, 8, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Sunderland, K.S.; Yang, M.; Mao, C. Phage-enabled nanomedicine: From probes to therapeutics in precision medicine. Angew. Chem. Int. Ed. 2017, 56, 1964–1992. [Google Scholar] [CrossRef]
- Hesse, S.; Adhya, S. Phage therapy in the twenty-first century: Facing the decline of the antibiotic era; is it finally time for the age of the phage? Annu. Rev. Microbiol. 2019, 73, 155–174. [Google Scholar] [CrossRef] [PubMed]
- Pires, D.P.; Meneses, L.; Brandão, A.C.; Azeredo, J. An overview of the current state of phage therapy for the treatment of biofilm-related infections. Curr. Opin. Virol. 2022, 53, 101209. [Google Scholar] [CrossRef] [PubMed]
- Champagne-Jorgensen, K.; Luong, T.; Darby, T.; Roach, D.R. Immunogenicity of bacteriophages. Trends Microbiol. 2023, 31, 1058–1071. [Google Scholar] [CrossRef] [PubMed]
- Kortright, K.E.; Chan, B.K.; Koff, J.L.; Turner, P.E. Phage therapy: A renewed approach to combat antibiotic-resistant bacteria. Cell Host Microbe 2019, 25, 219–232. [Google Scholar] [CrossRef]
- Mochizuki, K.; Matsukura, L.; Ito, Y.; Miyashita, N.; Taki, M. A medium-firm drug-candidate library of cryptand-like structures on t7 phage: Design and selection of a strong binder for hsp90. Org. Biomol. Chem. 2021, 19, 146–150. [Google Scholar] [CrossRef]
- Schmidt, A.; Kellermann, J.; Lottspeich, F. A novel strategy for quantitative proteomics using isotope-coded protein labels. Proteomics 2005, 5, 4–15. [Google Scholar] [CrossRef] [PubMed]
- Bernard, J.M.L.; Francis, M.B. Chemical strategies for the covalent modification of filamentous phage. Front. Microbiol. 2014, 5, 734. [Google Scholar] [CrossRef]
- Abello, N.; Kerstjens, H.A.M.; Postma, D.S.; Bischoff, R. Selective acylation of primary amines in peptides and proteins. J. Proteome Res. 2007, 6, 4770–4776. [Google Scholar] [CrossRef]
- Carmody, C.M.; Goddard, J.M.; Nugen, S.R. Bacteriophage capsid modification by genetic and chemical methods. Bioconj. Chem. 2021, 32, 466–481. [Google Scholar] [CrossRef] [PubMed]
- Marinelli, L.J.; Piuri, M.; Swigoňová, Z.; Balachandran, A.; Oldfield, L.M.; van Kessel, J.C.; Hatfull, G.F. Bred: A simple and powerful tool for constructing mutant and recombinant bacteriophage genomes. PLoS ONE 2008, 3, e3957. [Google Scholar] [CrossRef]
- Tan, Y.; Tian, T.; Liu, W.; Zhu, Z.; Yang, C.J. Advance in phage display technology for bioanalysis. Biotechnol. J. 2016, 11, 732–745. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Chen, I.A. Phage engineering and the evolutionary arms race. Curr. Opin. Biotechnol. 2021, 68, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Ragothaman, M.; Yoo, S.Y. Engineered phage-based cancer vaccines: Current advances and future directions. Vaccines 2023, 11, 919. [Google Scholar] [CrossRef] [PubMed]
- Veeranarayanan, S.; Azam, A.H.; Kiga, K.; Watanabe, S.; Cui, L. Bacteriophages as solid tumor theragnostic agents. Int. J. Mol. Sci. 2022, 23, 402. [Google Scholar] [CrossRef] [PubMed]
- Li, X.-T.; Peng, S.-Y.; Feng, S.-M.; Bao, T.-Y.; Li, S.-Z.; Li, S.-Y. Recent progress in phage-based nanoplatforms for tumor therapy. Small 2024, 20, 2307111. [Google Scholar] [CrossRef] [PubMed]
- Farr, R.; Choi, D.S.; Lee, S.-W. Phage-based nanomaterials for biomedical applications. Acta Biomater. 2014, 10, 1741–1750. [Google Scholar] [CrossRef]
- Chang, C.; Guo, W.; Yu, X.; Guo, C.; Zhou, N.; Guo, X.; Huang, R.-L.; Li, Q.; Zhu, Y. Engineered m13 phage as a novel therapeutic bionanomaterial for clinical applications: From tissue regeneration to cancer therapy. Mater. Today Bio 2023, 20, 100612. [Google Scholar] [CrossRef] [PubMed]
- Rehman, S.; Ali, Z.; Khan, M.; Bostan, N.; Naseem, S. The dawn of phage therapy. Rev. Med. Virol. 2019, 29, e2041. [Google Scholar] [CrossRef] [PubMed]
- Meile, S.; Du, J.; Dunne, M.; Kilcher, S.; Loessner, M.J. Engineering therapeutic phages for enhanced antibacterial efficacy. Curr. Opin. Virol. 2022, 52, 182–191. [Google Scholar] [CrossRef] [PubMed]
- Stacey, H.J.; De Soir, S.; Jones, J.D. The safety and efficacy of phage therapy: A systematic review of clinical and safety trials. Antibiotics 2022, 11, 1340. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, D.; Mihara, K.; Takayama, R.; Nakamura, Y.; Ueda, M.; Tsumuraya, T.; Fujii, I. Chemical modification of phage-displayed helix-loop-helix peptides to construct kinase-focused libraries. ChemBioChem 2021, 22, 3406–3409. [Google Scholar] [CrossRef] [PubMed]
- Branston, S.; Stanley, E.; Ward, J.; Keshavarz-Moore, E. Study of robustness of filamentous bacteriophages for industrial applications. Biotechnol. Bioeng. 2011, 108, 1468–1472. [Google Scholar] [CrossRef] [PubMed]
- Zheng, M.; Gao, J. Phage display of two distinct warheads to inhibit challenging proteins. ACS Chem. Biol. 2023, 18, 2259–2266. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Cao, F.; Gao, Y.; Zhang, C.; Qian, Z.; Zhang, J.; Mao, Z. Microenvironment-activated nanozyme-armed bacteriophages efficiently combat bacterial infection. Adv. Mater. 2023, 35, 2301349. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Borg, R.E.; Dow, L.P.; Pruitt, B.L.; Chen, I.A. Controlled phage therapy by photothermal ablation of specific bacterial species using gold nanorods targeted by chimeric phages. Proc. Natl. Acad. Sci. USA 2020, 117, 1951–1961. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Fan, X.; Gonzalez Moreno, M.; Tkhilaishvili, T.; Du, W.; Zhang, X.; Nie, C.; Trampuz, A.; Haag, R. Photocatalytic quantum dot-armed bacteriophage for combating drug-resistant bacterial infection. Adv. Sci. 2022, 9, 2105668. [Google Scholar] [CrossRef]
- Carrico, Z.M.; Farkas, M.E.; Zhou, Y.; Hsiao, S.C.; Marks, J.D.; Chokhawala, H.; Clark, D.S.; Francis, M.B. N-terminal labeling of filamentous phage to create cancer marker imaging agents. ACS Nano 2012, 6, 6675–6680. [Google Scholar] [CrossRef] [PubMed]
- Stevens, T.K.; Palaniappan, K.K.; Ramirez, R.M.; Francis, M.B.; Wemmer, D.E.; Pines, A. Hypercest detection of a 129xe-based contrast agent composed of cryptophane-a molecular cages on a bacteriophage scaffold. Magn. Reson. Med. 2013, 69, 1245–1252. [Google Scholar] [CrossRef] [PubMed]
- Tong, G.J.; Hsiao, S.C.; Carrico, Z.M.; Francis, M.B. Viral capsid DNA aptamer conjugates as multivalent cell-targeting vehicles. J. Am. Chem. Soc. 2009, 131, 11174–11178. [Google Scholar] [CrossRef]
- Sathaliyawala, T.; Rao, M.; Maclean, D.M.; Birx, D.L.; Alving, C.R.; Rao, V.B. Assembly of human immunodeficiency virus (hiv) antigens on bacteriophage t4: A novel in vitro approach to construct multicomponent hiv vaccines. J. Virol. 2006, 80, 7688–7698. [Google Scholar] [CrossRef]
- Guan, J.; Oromí-Bosch, A.; Mendoza, S.D.; Karambelkar, S.; Berry, J.D.; Bondy-Denomy, J. Bacteriophage genome engineering with CRISPR–Cas13a. Nat. Microbiol. 2022, 7, 1956–1966. [Google Scholar] [CrossRef] [PubMed]
- Paul, B.; Montoya, G. CRISPR-Cas12a: Functional overview and applications. Biomed. J. 2020, 43, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Park, J.Y.; Moon, B.Y.; Park, J.W.; Thornton, J.A.; Park, Y.H.; Seo, K.S. Genetic engineering of a temperate phage-based delivery system for CRISPR/Cas9 antimicrobials against staphylococcus aureus. Sci. Rep. 2017, 7, 44929. [Google Scholar] [CrossRef] [PubMed]
- Braun, P.; Raab, R.; Bugert, J.J.; Braun, S. Recombinant reporter phage rTUN1::nLuc enables rapid detection and real-time antibiotic susceptibility testing of klebsiella pneumoniae K64 strains. ACS Sens. 2023, 8, 630–639. [Google Scholar] [CrossRef] [PubMed]
- Marinelli, L.J.; Hatfull, G.F.; Piuri, M. Recombineering. Bacteriophage 2012, 2, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Hussain, W.; Yang, X.; Ullah, M.; Wang, H.; Aziz, A.; Xu, F.; Asif, M.; Ullah, M.W.; Wang, S. Genetic engineering of bacteriophages: Key concepts, strategies, and applications. Biotechnol. Adv. 2023, 64, 108116. [Google Scholar] [CrossRef]
- Ramirez-Chamorro, L.; Boulanger, P.; Rossier, O. Strategies for bacteriophage T5 mutagenesis: Expanding the toolbox for phage genome engineering. Front. Microbiol. 2021, 12, 667332. [Google Scholar] [CrossRef] [PubMed]
- Martel, B.; Moineau, S. CRISPR-Cas: An efficient tool for genome engineering of virulent bacteriophages. Nucleic Acids Res. 2014, 42, 9504–9513. [Google Scholar] [CrossRef] [PubMed]
- Lv, S.; Wang, Y.; Jiang, K.; Guo, X.; Zhang, J.; Zhou, F.; Li, Q.; Jiang, Y.; Yang, C.; Teng, T. Genetic engineering and biosynthesis technology: Keys to unlocking the chains of phage therapy. Viruses 2023, 15, 1736. [Google Scholar] [CrossRef] [PubMed]
- Shivram, H.; Cress, B.F.; Knott, G.J.; Doudna, J.A. Controlling and enhancing CRISPR systems. Nat. Chem. Biol. 2021, 17, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Malone, L.M.; Birkholz, N.; Fineran, P.C. Conquering CRISPR: How phages overcome bacterial adaptive immunity. Curr. Opin. Biotechnol. 2021, 68, 30–36. [Google Scholar] [CrossRef] [PubMed]
- da Silva, J.L.; Piuri, M.; Broussard, G.; Marinelli, L.J.; Bastos, G.M.; Hirata, R.D.C.; Hatfull, G.F.; Hirata, M.H. Application of BRED technology to construct recombinant D29 reporter phage expressing EGFP. FEMS Microbiol. Lett. 2013, 344, 166–172. [Google Scholar] [CrossRef] [PubMed]
- Jaschke, P.R.; Lieberman, E.K.; Rodriguez, J.; Sierra, A.; Endy, D. A fully decompressed synthetic bacteriophage øX174 genome assembled and archived in yeast. Virol. J. 2012, 434, 278–284. [Google Scholar] [CrossRef]
- Assad-Garcia, N.; D’Souza, R.; Buzzeo, R.; Tripathi, A.; Oldfield, L.M.; Vashee, S.; Fouts, D.E. Cross-genus “boot-up” of synthetic bacteriophage in staphylococcus aureus by using a new and efficient DNA transformation method. Appl. Environ. Microbiol. 2022, 88, e01486-21. [Google Scholar] [CrossRef] [PubMed]
- Dhanker, R.; Hussain, T.; Tyagi, P.; Singh, K.J.; Kamble, S.S. The emerging trend of bio-engineering approaches for microbial nanomaterial synthesis and its applications. Front. Microbiol. 2021, 12, 638003. [Google Scholar] [CrossRef]
- Asar, M.; Newton-Northup, J.; Soendergaard, M. Improving pharmacokinetics of peptides using phage display. Viruses 2024, 16, 570. [Google Scholar] [CrossRef]
- Zhang, H.; Xu, Y.; Huang, Q.; Yi, C.; Xiao, T.; Li, Q. Natural phage nanoparticle-mediated real-time immuno-PCR for ultrasensitive detection of protein marker. Chem. Commun. 2013, 49, 3778–3780. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-H.; Moody, I.; Zeng, Z.; Fleischer, E.B.; Weiss, G.A.; Shea, K.J. Synthesis of a high affinity complementary peptide–polymer nanoparticle (NP) pair using phage display. ACS Appl. Bio Mater. 2021, 4, 2704–2712. [Google Scholar] [CrossRef] [PubMed]
- Bush, J.; Singh, S.; Vargas, M.; Oktay, E.; Hu, C.-H.; Veneziano, R. Synthesis of DNA origami scaffolds: Current and emerging strategies. Molecules 2020, 25, 3386. [Google Scholar] [CrossRef] [PubMed]
- Mao, C.; Solis, D.J.; Reiss, B.D.; Kottmann, S.T.; Sweeney, R.Y.; Hayhurst, A.; Georgiou, G.; Iverson, B.; Belcher, A.M. Virus-based toolkit for the directed synthesis of magnetic and semiconducting nanowires. Science 2004, 303, 213–217. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Cao, B.; Yang, M.; Zhu, Y.; Suh, J.; Mao, C. Identification of novel short BaTiO3-binding/nucleating peptides for phage-templated in situ synthesis of BaTiO3 polycrystalline nanowires at room temperature. ACS Appl. Mater. Interfaces 2016, 8, 30714–30721. [Google Scholar] [CrossRef] [PubMed]
- Żelechowska, K.; Karczewska-Golec, J.; Karczewski, J.; Łoś, M.; Kłonkowski, A.M.; Węgrzyn, G.; Golec, P. Phage-directed synthesis of photoluminescent zinc oxide nanoparticles under benign conditions. Bioconjugate Chem. 2016, 27, 1999–2006. [Google Scholar] [CrossRef] [PubMed]
- Nam, K.T.; Peelle, B.R.; Lee, S.-W.; Belcher, A.M. Genetically driven assembly of nanorings based on the M13 virus. Nano Lett. 2004, 4, 23–27. [Google Scholar] [CrossRef]
- Merzlyak, A.; Lee, S.-W. Phage as templates for hybrid materials and mediators for nanomaterial synthesis. Curr. Opin. Chem. Biol. 2006, 10, 246–252. [Google Scholar] [CrossRef]
- Tang, J.; Fraden, S. Isotropic-cholesteric phase transition in colloidal suspensions of filamentous bacteriophage fd. Liq. Cryst. 1995, 19, 459–467. [Google Scholar] [CrossRef]
- Moon, J.-S.; Choi, E.J.; Jeong, N.-N.; Sohn, J.-R.; Han, D.-W.; Oh, J.-W. Research progress of M13 bacteriophage-based biosensors. J. Nanomater. 2019, 9, 1448. [Google Scholar] [CrossRef] [PubMed]
- Mao, C.; Flynn, C.E.; Hayhurst, A.; Sweeney, R.; Qi, J.; Georgiou, G.; Iverson, B.; Belcher, A.M. Viral assembly of oriented quantum dot nanowires. Proc. Natl. Acad. Sci. USA 2003, 100, 6946–6951. [Google Scholar] [CrossRef] [PubMed]
- Mao, C.; Wang, F.; Cao, B. Controlling nanostructures of mesoporous silica fibers by supramolecular assembly of genetically modifiable bacteriophages. Angew. Chem. Int. Ed. 2012, 51, 6411–6415. [Google Scholar] [CrossRef] [PubMed]
- He, T.; Abbineni, G.; Cao, B.; Mao, C. Nanofibrous bio-inorganic hybrid structures formed through self-assembly and oriented mineralization of genetically engineered phage nanofibers. Small 2010, 6, 2230–2235. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yang, M.; Zhu, Y.; Wang, L.; Tomsia, A.P.; Mao, C. Phage nanofibers induce vascularized osteogenesis in 3D printed bone scaffolds. Adv. Mater. 2014, 26, 4961–4966. [Google Scholar] [CrossRef] [PubMed]
- Taton, T.A.; Mirkin, C.A.; Letsinger, R.L. Scanometric DNA array detection with nanoparticle probes. Science 2000, 289, 1757–1760. [Google Scholar] [CrossRef]
- Wang, L.; Li, H.; Wang, X.; Yang, X.; Tian, C.; Sun, D.; Liu, L.; Li, J. Modification of low-energy surfaces using bicyclic peptides discovered by phage display. J. Am. Chem. Soc. 2023, 145, 17613–17620. [Google Scholar] [CrossRef]
- Guliy, O.I.; Evstigneeva, S.S.; Khanadeev, V.A.; Dykman, L.A. Antibody phage display technology for sensor-based virus detection: Current status and future prospects. Biosensors 2023, 13, 640. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-J.; McCoy, M.; Gee, S.J.; González-Sapienza, G.G.; Hammock, B.D. Noncompetitive phage anti-immunocomplex real-time polymerase chain reaction for sensitive detection of small molecules. Anal. Chem. 2011, 83, 246–253. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Cao, P.; Zhu, Y.; Lu, W.; Gu, N.; Mao, C. Phage-mediated counting by the naked eye of mirna molecules at attomolar concentrations in a petri dish. Nat. Mater. 2015, 14, 1058–1064. [Google Scholar] [CrossRef]
- Fang, H.; Zhou, Y.; Ma, Y.; Chen, Q.; Tong, W.; Zhan, S.; Guo, Q.; Xiong, Y.; Tang, B.Z.; Huang, X. M13 bacteriophage-assisted recognition and signal spatiotemporal separation enabling ultrasensitive light scattering immunoassay. ACS Nano 2023, 17, 18596–18607. [Google Scholar] [CrossRef]
- Hou, J.; Qian, X.; Xu, Y.; Guo, Z.; Thierry, B.; Yang, C.-T.; Zhou, X.; Mao, C. Rapid and reliable ultrasensitive detection of pathogenic H9N2 viruses through virus-binding phage nanofibers decorated with gold nanoparticles. Biosens. Bioelectron. 2023, 237, 115423. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.-X.; Wang, C.-Y.; Li, Y.-Y.; Li, J.; Wan, Q.-Q.; Chen, J.-H.; Tay, F.R.; Niu, L.-N. Considerations and caveats in combating ESKAPE pathogens against nosocomial infections. Adv. Sci. 2020, 7, 1901872. [Google Scholar] [CrossRef] [PubMed]
- Nobrega, F.L.; Vlot, M.; de Jonge, P.A.; Dreesens, L.L.; Beaumont, H.J.E.; Lavigne, R.; Dutilh, B.E.; Brouns, S.J.J. Targeting mechanisms of tailed bacteriophages. Nat. Rev. Microbiol. 2018, 16, 760–773. [Google Scholar] [CrossRef] [PubMed]
- Paczesny, J.; Richter, Ł.; Hołyst, R. Recent progress in the detection of bacteria using bacteriophages: A review. Viruses 2020, 12, 845. [Google Scholar] [CrossRef] [PubMed]
- Richter, Ł.; Janczuk-Richter, M.; Niedziółka-Jönsson, J.; Paczesny, J.; Hołyst, R. Recent advances in bacteriophage-based methods for bacteria detection. Drug Discov. Today 2018, 23, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Alcaine, S.D.; Jiang, Z.; Rotello, V.M.; Nugen, S.R. Detection of Escherichia coli in drinking water using T7 bacteriophage-conjugated magnetic probe. Anal. Chem. 2015, 87, 8977–8984. [Google Scholar] [CrossRef]
- You, H.; Wang, M.; Wang, S.; Xu, J.; Hu, S.; Li, T.; Yu, Z.; Tang, D.; Gan, N. Ultrasensitive and specific phage@DNAzyme probe-triggered fluorescent click chemistry for on-site detection of foodborne pathogens using a smartphone. Anal. Chem. 2023, 95, 11211–11218. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Gou, Z.; Yuan, H.; Yue, X.; Lu, S.; Fu, Z. Employment of the phage cocktail as a species-specific recognition agent for wide-spectrum detection of bacterial strains. Anal. Chem. 2023, 95, 10008–10016. [Google Scholar] [CrossRef]
- Wu, M.-Y.; Chen, L.; Chen, Q.; Hu, R.; Xu, X.; Wang, Y.; Li, J.; Feng, S.; Dong, C.; Zhang, X.-L.; et al. Engineered phage with aggregation-induced emission photosensitizer in cocktail therapy against sepsis. Adv. Mater. 2023, 35, 2208578. [Google Scholar] [CrossRef] [PubMed]
- Brown, E.D.; Wright, G.D. Antibacterial drug discovery in the resistance era. Nature 2016, 529, 336–343. [Google Scholar] [CrossRef] [PubMed]
- Kupferschmidt, K. Resistance fighters. Science 2016, 352, 758–761. [Google Scholar] [CrossRef] [PubMed]
- Thiel, K. Old dogma, new tricks—21st century phage therapy. Nat. Biotechnol. 2004, 22, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhang, J.; Wei, J.; Jiang, L.; Jiang, L.; Sun, Y.; Zeng, Z.; Wang, Z. Phage-inspired strategies to combat antibacterial resistance. Crit. Rev. Microbiol. 2024, 50, 196–211. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Yang, Y.; Guo, Y.; Lu, S.; Du, Y.; Li, J.-J.; Zhang, X.; Leung, N.L.C.; Zhao, Z.; Niu, G.; et al. Phage-guided targeting, discriminative imaging, and synergistic killing of bacteria by AIE bioconjugates. J. Am. Chem. Soc. 2020, 142, 3959–3969. [Google Scholar] [CrossRef] [PubMed]
- Ran, B.; Yuan, Y.; Xia, W.; Li, M.; Yao, Q.; Wang, Z.; Wang, L.; Li, X.; Xu, Y.; Peng, X. A photo-sensitizable phage for multidrug-resistant Acinetobacter baumannii therapy and biofilm ablation. Chem. Sci. 2021, 12, 1054–1061. [Google Scholar] [CrossRef]
- Zhang, J.; He, X.; Tang, B.Z. Aggregation-induced emission-armored living bacteriophage-DNA nanobioconjugates for targeting, imaging, and efficient elimination of intracellular bacterial infection. ACS Nano 2024, 18, 3199–3213. [Google Scholar] [CrossRef] [PubMed]
- Jiang, T.; Yang, T.; Chen, Y.; Miao, Y.; Xu, Y.; Jiang, H.; Yang, M.; Mao, C. Emulating interactions between microorganisms and tumor microenvironment to develop cancer theranostics. Theranostics 2022, 12, 2833–2859. [Google Scholar] [CrossRef]
- Dong, X.; Pan, P.; Zheng, D.-W.; Bao, P.; Zeng, X.; Zhang, X.-Z. Bioinorganic hybrid bacteriophage for modulation of intestinal microbiota to remodel tumor-immune microenvironment against colorectal cancer. Sci. Adv. 2020, 6, eaba1590. [Google Scholar] [CrossRef] [PubMed]
- Geller, L.T.; Barzily-Rokni, M.; Danino, T.; Jonas, O.H.; Shental, N.; Nejman, D.; Gavert, N.; Zwang, Y.; Cooper, Z.A.; Shee, K.; et al. Potential role of intratumor bacteria in mediating tumor resistance to the chemotherapeutic drug gemcitabine. Science 2017, 357, 1156–1160. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, H.; Bording-Jorgensen, M.; Dijk, S.; Wine, E. The complex interplay between chronic inflammation, the microbiome, and cancer: Understanding disease progression and what we can do to prevent it. Cancers 2018, 10, 83. [Google Scholar] [CrossRef] [PubMed]
- Federici, S.; Kredo-Russo, S.; Valdés-Mas, R.; Kviatcovsky, D.; Weinstock, E.; Matiuhin, Y.; Silberberg, Y.; Atarashi, K.; Furuichi, M.; Oka, A.; et al. Targeted suppression of human IBD-associated gut microbiota commensals by phage consortia for treatment of intestinal inflammation. Cell 2022, 185, 2879–2898.e2824. [Google Scholar] [CrossRef] [PubMed]
- Baudino, A.T. Targeted cancer therapy: The next generation of cancer treatment. Curr. Drug Discov. Technol. 2015, 12, 3–20. [Google Scholar] [CrossRef]
- Chen, X.; Mendes, B.G.; Alves, B.S.; Duan, Y. Chapter four—Phage therapy in gut microbiome. In Progress in Molecular Biology and Translational Science; Academic Press: Cambridge, MA, USA, 2023; Volume 201, pp. 93–118. [Google Scholar]
- Zheng, D.-W.; Dong, X.; Pan, P.; Chen, K.-W.; Fan, J.-X.; Cheng, S.-X.; Zhang, X.-Z. Phage-guided modulation of the gut microbiota of mouse models of colorectal cancer augments their responses to chemotherapy. Nat. Biomed. Eng. 2019, 3, 717–728. [Google Scholar] [CrossRef] [PubMed]
- Gandra, N.; Abbineni, G.; Qu, X.; Huai, Y.; Wang, L.; Mao, C. Bacteriophage bionanowire as a carrier for both cancer-targeting peptides and photosensitizers and its use in selective cancer cell killing by photodynamic therapy. Small 2013, 9, 215–221. [Google Scholar] [CrossRef]
- Dong, X.; Pan, P.; Ye, J.-J.; Zhang, Q.-L.; Zhang, X.-Z. Hybrid M13 bacteriophage-based vaccine platform for personalized cancer immunotherapy. Biomaterials 2022, 289, 121763. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Pan, P.; Zhang, Q.; Ye, J.-J.; Zhang, X.-Z. Engineered living bacteriophage-enabled self-adjuvanting hydrogel for remodeling tumor microenvironment and cancer therapy. Nano Lett. 2023, 23, 1219–1228. [Google Scholar] [CrossRef]
- Qamar, Z.; Qizilbash, F.F.; Iqubal, K.M.; Ali, A.; Narang, K.J.; Ali, J.; Baboota, S. Nano-based drug delivery system: Recent strategies for the treatment of ocular disease and future perspective. Recent. Pat. Drug Deliv. Formul. 2019, 13, 246–254. [Google Scholar] [CrossRef] [PubMed]
- Sioud, M. Phage display libraries: From binders to targeted drug delivery and human therapeutics. Mol. Biotechnol. 2019, 61, 286–303. [Google Scholar] [CrossRef] [PubMed]
- Kalarical Janardhanan, S.; Narayan, S.; Abbineni, G.; Hayhurst, A.; Mao, C. Architectonics of phage-liposome nanowebs as optimized photosensitizer vehicles for photodynamic cancer therapy. Mol. Cancer Ther. 2010, 9, 2524–2535. [Google Scholar] [CrossRef] [PubMed]
- Kovacs, E.W.; Hooker, J.M.; Romanini, D.W.; Holder, P.G.; Berry, K.E.; Francis, M.B. Dual-surface-modified bacteriophage MS2 as an ideal scaffold for a viral capsid-based drug delivery system. Bioconjugate Chem. 2007, 18, 1140–1147. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Hsiao, S.C.; Carrico, Z.M.; Francis, M.B. Genome-free viral capsids as multivalent carriers for taxol delivery. Angew. Chem. Int. Ed. 2009, 48, 9493–9497. [Google Scholar] [CrossRef]
- Gandra, N.; Wang, D.-D.; Zhu, Y.; Mao, C. Virus-mimetic cytoplasm-cleavable magnetic/silica nanoclusters for enhanced gene delivery to mesenchymal stem cells. Angew. Chem. Int. Ed. 2013, 52, 11278–11281. [Google Scholar] [CrossRef]
- Tsedev, U.; Lin, C.-W.; Hess, G.T.; Sarkaria, J.N.; Lam, F.C.; Belcher, A.M. Phage particles of controlled length and genome for in vivo targeted glioblastoma imaging and therapeutic delivery. ACS Nano 2022, 16, 11676–11691. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).