Abstract
Uruguayan ammonoids are preserved in phosphate and siderite nodules found at the basalmost tillite-like conglomerates of the San Gregorio Formation. This lithostratigraphic unit was deposited under glacial conditions and its age (as well as that of the nodules) has been highly debated because glaciations were intermittent in Gondwana during the Late Paleozoic. Reef-builder organisms (e.g., Rugosa and Tabulata), goniatite and orthoceratid cephalopods, brachiopods, sponges, actinopterygians and other indeterminate gnatostomes, as well as fragmentary stems and roots of cf. Lycopsida are the most frequent fossils in the nodules. According to new biostratigraphic and paleoclimatic evidence, these taxa are representative of a reefal environment of a preliminary Devonian age including species that are common in the underlying Early Devonian (Emsian) Durazno Group. Among the ammonoid remains, more than 40 clusters of hatchling goniatites were found in the nodules. Each cluster contains a variable number of shells similar in shape to some of the adults also preserved within the nodules, representing a single species preserved at the same developmental stage (3 mm on size average). The strongly packed shells are enveloped by a substance with a different chemical composition and microstructure with respect to that of the nodule matrix, possibly indicating the presence of a gelatinous-like substance reminiscent of that secreted by the females of some extant cuttlefish and octopuses at the time of the egg spawn. Differing from previously described ammonoid accumulations, our clusters are unique in containing individuals of just a single species preserved in the same ontogenetic stage. That allows us to suggest that they represent a mode of reproduction in which hatchlings were morphologically similar to their parents and occupied the same habitat. Our results are thus one of the oldest known records of reproductive strategies in Paleozoic ammonoids and the phosphate and siderite nodules from the San Gregorio Formation are here classified as a new Konservat-Lagerstätte, which is the oldest known for South America.
1. Introduction
In Uruguay, fossil cephalopods are exclusively represented by ammonoids and nautiloids preserved within phosphate and siderite nodules included at the basalmost levels of the San Gregorio Formation. This formation is well represented along both the banks of the Negro River, encompassing the Tacuarembó and Durazno counties, and extends to the East, cropping up at the Cerro Largo department, close to the Melo city (Figure 1). The deposition of the San Gregorio Formation has been traditionally related to the Late Paleozoic Ice Age, which was apparently represented by short and subtle events in Gondwana, followed by episodes of more attenuated but still cold conditions [1]. The age of the San Gregorio Formation has been constrained as Early Permian based on palynological data from boreholes and surface samples taken from the deposits that crop out near the Melo city (Cerro Largo Department) [1,2], among others); such deposits lack fossiliferous nodules (Figure 1). Indeed, the phosphate and siderite nodules containing goniatite ammonoids [3] and actinopterygians [4,5] were thought to have an Upper Carboniferous age and they are common along the banks of the Negro River in Tacuarembó and Durazno counties (Figure 1).
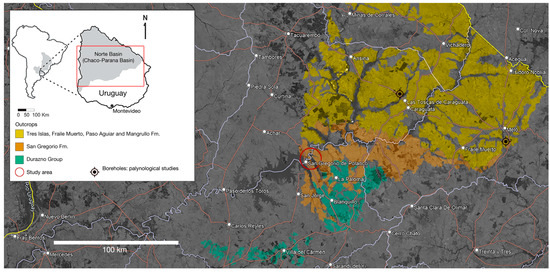
Figure 1.
Map showing the distribution of the studied areas of the San Gregorio Formation of Uruguay in the vicinity of the San Gregorio de Polanco city along both the banks of the Negro River (Tacuarembó and Durazno Departments, red circle), and close to the Melo city in the Cerro Largo Department (black symbols) [1,2,6]. The fossiliferous phosphate and siderite nodules described herein are limited to the former area.
The well-preserved cephalopod specimens initially studied by [3,7,8] led to the description of the goniatite Eosianites (Glaphyrites) rionegrensis [7] and the orthoceratid nautiloid Dolorthoceras chubutense [8]. However, no other records of such well-preserved specimens were made since and the research was reduced to better define the paleoenvironmental settings and to try solving the controversy about the age of the San Gregorio Formation. However, the fossil content of the nodules does not provide relevant evidence for these studies.
Recent investigations in newly discovered outcrops of the San Gregorio Formation in the area of the Negro River banks have yielded new cephalopod materials preserved in phosphate and siderite nodules that may represent new species, but direct (at hand) comparisons with the Glaphyrites rionegrensis-type specimen and paratypes previously reported by [7], would be required to formally describe them.
The high potential of the San Gregorio nodules to preserve soft tissues is well known. We have corroborated this taphonomic feature in a recent preliminary contribution [9], where we described masses of small ammonoids preserving soft tissues and a possible evidence of ammonoid reproductive biology.
The aim of this paper is to analyze the meaning of these ammonoid concentrations containing a high number of tiny shells and to describe the morphology of the ammonitella stage from the best preserved specimens. Furthermore, we will analyze the environmental conditions suggested for the San Gregorio Formation in contrast to the nature of the fossil community preserved by the nodules in order to shed light about the age of the specimens and that of the nodule formation.
1.1. Geological Setting
Paleozoic sediments in Uruguay are primarily located in what is known as the Norte Basin, which covers the north and northeastern region of the country as part of the southern extension of the Brazilian Paraná Basin [10]. The oldest sedimentary deposits of the Norte Basin are represented by the Durazno Group and date back to the Lower Devonian (Emsian) [11]. This group comprises the base to the top, the Cerrezuelo, Cordobés and the La Paloma formations [12], distributed along the southern central edge of the Norte Basin and spreading northwest over the right margin of the Negro River.
Overlying the Durazno Group is the glacio-marine San Gregorio Formation, a sedimentary sequence distinguished mainly by the presence of diamictites, sandstones, and rhythmic shales (varves) with dropstones. These typical glacial to postglacial environments are described for several Gondwanan continents and today represent the Late Paleozoic Ice Age (LPIA) e.g., [13] (Figure 2). Recent studies have shown that the glacial and postglacial deposits of the San Gregorio Formation originated from the weathering of Lower to Upper Proterozoic Terranes, and received a substantial sedimentary contribution from the underlying Lower Devonian (Emsian) formations [13,14,15,16,17]. The Lower Devonian sequence was deposited unconformably over crystalline basement rocks with a slight northwest inclination [16].
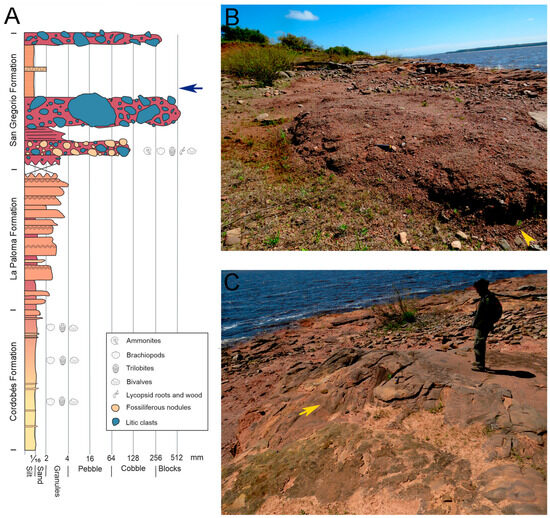
Figure 2.
Basic stratigraphy of the nodule bearing San Gregorio Formation of Uruguay and the underlying Early Devonian Durazno Group. (A). Schematic stratigraphic section of one of the outcrops where we found the specimens described herein. The arrow points to the level where striate bedrock surfaces from glacier displacement were found. (B). Photograph of the basal level of tillites that contain the reworked fossiliferous nodules (head arrow). (C). Detail of the bedrock surface scratched by rocks embedded in glacial ice during glacier displacement. It is worth noting that these striae (yellow arrow) are stratigraphically above the tillites that are bringing the nodules, evidencing the periglacial and postglacial origin of these deposits. Scale: Engineer Sebastian Marmol.
Previous research on the age of the San Gregorio Formation has suggested deposition during the Carboniferous-Permian or close to this boundary. This age was mostly calibrated by palynostratigraphy based on microfossil assemblages obtained from boreholes of the eastern section of the San Gregorio Formation at the Cerro Largo county i.e., [1,2,6], which is lithologically different from the deposits found at the central region of the country, over the Negro River banks which are characterized by the presence of fossiliferous phosphate and siderite nodules (see Figure 1).
The Upper Carboniferous-Lower Permian age suggested by the microfossils from the eastern boreholes is congruent with previously published age estimates based on actinopterygians i.e., [5] and on cephalopods [7,8] from the nodules of the central region. However, the actinopterygians are represented by endemic species not found from other Pangean localities [18], and goniatite ammonoids are mostly known from the Lower Devonian. It was also proposed an Upper Carboniferous age based on the presence of radiolarians while leaving open the possibility that the nodules were older [19]. Thus, the age assigned to the formation is in fact based on a single relying biostratigraphic tool (palynostratigraphy), which is not optimal.
Recent interdisciplinary studies have suggested a more complex scenario [9,20], of multiple glaciations that impacted Gondwana over a nearly 100-million-year interval, ranging from the Famennian (Late Devonian) to the Visean (Early Carboniferous) and possibly affecting part of the Capitanian (Middle Permian), but not uniformly affecting all regions of Gondwana [21]. Regional correlations indicate that glaciations in the Paraná basin originated during the Mid-Visean (Early Carboniferous) and persisted intermittently through the Early Permian [18,22].
1.2. Ammonoid Reproductive Strategies
According to most authors, the ammonoid ontogenetic development lacked larval stages, implying that hatchlings should resemble adults except for size [23]. Nevertheless, the successive embryonic stages as occurring before the hatching are represented by a small, generally bulbous protoconch that is the first living chamber followed by a short first whorl extending up to the primary constriction, a shallow notch that marks the position of the embryonic aperture [24]. Collectively, these initial ontogenetic forms are referred as the ammonitella embryonic stage [23,25,26,27,28], (among others), which represents the early developmental phase of ammonoids. As an embryonic stage, ammonitella should be developed inside the egg capsule.
The ammonitellae are morphologically different from their parents because growth of the phragmocone occurs after hatching and juvenile configuration (miniatures similar in shape to their parents) may have arisen relatively soon in post-embryonic development. It has been proposed that ammonoids hatched at the initial stage of ontogenetic development and thus they were represented by only the initial chamber, considered as a planktonic larval stage that transformed into a juvenile through some kind of metamorphosis [29,30]. According to this theory, ammonitellae are a larval rather than an embryonic stage. However, other evidence would indicate that at least some ammonoids may have been ovoviviparous, as suggested by [30]. These authors discovered clusters of “embryonic” ammonitellae in the lower Aptian (Cretaceous) of Russia, associated with the decomposing shell of an adult ammonoid. Intriguingly, each tiny embryonic specimen carried an already mineralized aptychi, a condition not previously observed at such an early stage of ammonoid development. After a detailed study of the taphonomic conditions affecting fossil preservation and a comparative analysis of the known reproductive traits in extant and extinct cephalopods, [30] concluded that ovoviviparity was the most plausible explanation for the association of embryonic masses with the shell of an adult ammonoid. In this scenario, hatchling ammonitellae would complete their development inside the female’s body or within protective gelatinous sacs secreted by the mother’s glands to avoid predation as it is observed in some extant cuttlefishes and octopuses [31].
Nevertheless, these hypotheses, particularly that of [29] have been widely rejected by most ammonoid specialists, primarily because a single planktonic and dispersive “protoconch” has never been found in the fossil record [24].
Following the common developmental concept that embryonic means not born organisms, the characterization of the ammonitella stage has to be defined as a pre-hatching stage and as a consequence, it would be difficult to find it in the fossil record. Accordingly, the predicted size of the egg in fossil ammonoids would be equivalent to the ammonitella size and not to that of the initial chamber [32].
Despite many authors have described the presence of a larval (paralarva) phase in the development of some cephalopod groups, the prevailing view is that the reproductive strategy of cephalopods follows direct development, because the structural features of the adults are present in the larva, and the morphologic changes between the larva and the adult are gradual as in direct development meaning that there is not a true metamorphosis [33]. Although the controversy about larval stage different from the juvenile or larval stage as an intermediary phase in the development of cephalopods is not completely resolved, taking into account what is known about extant cephalopods, a direct development can be inferred for the extinct groups e.g., [23,25,26,28,34].
According to previous studies e.g., [28,35], there was a notable evolutionary change leading to a reduction in the ammonitella size from the Early Devonian when they were relatively large (averaging 0.54–2.6 mm, but up to 5 mm) to the Late Cretaceous when their size was minimal (0.7–1 mm) [36]. Furthermore, a notorious reduction in ammonitella size from the Early to the Late Devonian was also proposed [28,36,37] but see also [32]. These size variations can be linked to episodes of fluctuating water temperatures, with the largest ammonitellae associated with temperate and warm climates during periods of global warming. Increase in shell size from the embryonic ammonitella stage to the hatchling or early postembryonic individuals, involves the addition of 2.5 to 3.3 whorls, resulting in a shell diameter of 3 to 5 mm [35].
2. Materials and Methods
Five of the more than forty clusters of tiny ammonoid shells (JMMO-IMT 0215, 0304, 0305, 0311 and 312; see Figure 3 and Figure 4) housed at the Museo de Las Bochas (San Gregorio de Polanco city) are here described. The specimens were found preserved inside phosphate and siderite nodules of the San Gregorio Formation of Uruguay (Figure 1 and Figure 2). The specimens were analyzed using a Jeol JSM 5900LV SEM Scanning Electron Microscopy and in all cases the samples were mounted and examined with the microscope operated at 20 kV. The studied specimens are part of the Juan Manuel Mendez Ortiz collection (JMMO-IMT), recently formalized by the Municipality of Tacuarembó County, central Uruguay. Photographs and microphotographs of the specimens were taken by a Canon SX 60 HS Power Shot camera, PC 2154, Canon INC, Japan, with a Full HD 65 x optical zoom.
The microstructure of each ammonoid specimen within the masses (Figure 5) was studied under SEM using energy-dispersive X-ray spectroscopy analysis (EDS) to determine the chemical composition of the shells by a Thermo Scientific Ultra Dray EDS detector with NORAN System 7 X-Ray Microanalysis System. This method was also used to study the microstructure and chemical composition of the surrounding nodule matrix. Measurements of initial chamber and ammonitella diameters were taken following [36,38] (Figure 5B). The specimens in the clusters remained uncoated during their study with SEM.
3. Results
3.1. The Ammonitella Stage in the Early Juvenile Ammonoid Clusters
The study of the five nodules containing clusters of very small ammonoids (Figure 3 and Figure 4), allowed us to determine the morphology of the shell at the ammonitella stage and compare it with that described for Glaphyrites, the only previously known ammonoid taxon for Uruguay that was described by [7,8] as Eosianites (Glaphyrites) rionegrensis. Complete clusters have an oval outline and their sizes range from 30 to 42 mm in length, a variation possibly due to the incompleteness of some specimens. The largest one, measuring 55 mm in length preserves the mineralized shells of most of the represented individuals (Figure 3A) while other clusters contain only molds of a reduced number of larger individuals. Nevertheless, both kinds of fossils are generally well preserved, many even showing perfectly preserved siphuncle tubes, representing the oldest record of this structure in ammonoids. Individual shells in eight of the clusters were measured and a size variation between 2.9 to 5 mm in diameter was detected from one cluster to other, probably representing natural intraspecific or developmental stage variations (Figure 5). Such size variation in individuals believed to have approximately the same ontogenetic age was also observed in Mesozoic ammonitella clusters see [24,32] and reflects differences in growth rates.
The general morphology of the hatchling shells only differs in size and number of whorls, meaning in the phragmocone length, from that observed in adult and subadults ammonoids also preserved within the San Gregorio Formation nodules. The clusters are easily delimited inside the nodule matrix by exhibiting a different microstructure of long and intricate tubules and a different chemical composition (Figure 6).
These structures (Figure 6C1 and Figure 7A) are not found around other fossil groups within the nodules. Despite the high density of the ammonoids inside the clusters will avoid the complete shell delimitation, we estimate that each one would have enclosed a variable number of individuals (between 10 and 70) and those containing only a few individuals are represented by larger shells than those preserving the largest number. SEM images taken of some well-preserved individuals from the available studied clusters have facilitated the identification of key structures in goniatite ammonoids from Uruguay, which were previously used to reconstruct the microanatomy of the ammonitella stage from elsewhere species.
The embryonic shell (ammonitella) includes a semicircular to elliptical initial chamber in transverse cross section (Figure 7 and Figure 8), which is not easily distinguishable from the first whorl in external view (Figure 3, Figure 4 and Figure 7).

Figure 3.
Photographs of early juvenile ammonoid clusters from the Devonian of Uruguay. (A). Specimen JMMO-IMT 0215 represents the largest cluster found (approximately 55 mm in length) and it shows the closely packed ammonoids shells embedded in a possible gelatinous substance that protected and feed the individuals before hatching [39]. The ammonitellae average diameter at this stage is near 0.4 to 0.5 mm, and the shell has reached the three additional whorls that would characterize the hatchling phase outside the gelatinous envelope. Scale bar: 10 mm. (B,C). JMMO-IMT 0311. Part and counterpart of a cluster containing well-preserved, early juvenile tiny ammonoids, which allowed us to studied them by SEM and took good microphotographs of the initial chamber, as well as study the morphology that characterize the ammonitella stage for this new species of Uruguayan ammonoids (see Figure 7 and Figure 8). Scale bars: 5 mm.
Figure 3.
Photographs of early juvenile ammonoid clusters from the Devonian of Uruguay. (A). Specimen JMMO-IMT 0215 represents the largest cluster found (approximately 55 mm in length) and it shows the closely packed ammonoids shells embedded in a possible gelatinous substance that protected and feed the individuals before hatching [39]. The ammonitellae average diameter at this stage is near 0.4 to 0.5 mm, and the shell has reached the three additional whorls that would characterize the hatchling phase outside the gelatinous envelope. Scale bar: 10 mm. (B,C). JMMO-IMT 0311. Part and counterpart of a cluster containing well-preserved, early juvenile tiny ammonoids, which allowed us to studied them by SEM and took good microphotographs of the initial chamber, as well as study the morphology that characterize the ammonitella stage for this new species of Uruguayan ammonoids (see Figure 7 and Figure 8). Scale bars: 5 mm.


Figure 4.
JMMO-IMT 0230. Photograph of early juvenile ammonoid clusters from the Devonian of Uruguay. The image shows some shells inside a phosphate nodule of the San Gregorio Formation (Central Uruguay). Following [40], a possible gelatinous substance enclosing the shells seems to have been broken, maybe representing the moment in which the individuals abandon it and escape to the water-column, or the image might show the thin gelatinous envelop unexpectedly broken by a strong environmental event or by a predator attack. Scale bar: 5 mm.
Figure 4.
JMMO-IMT 0230. Photograph of early juvenile ammonoid clusters from the Devonian of Uruguay. The image shows some shells inside a phosphate nodule of the San Gregorio Formation (Central Uruguay). Following [40], a possible gelatinous substance enclosing the shells seems to have been broken, maybe representing the moment in which the individuals abandon it and escape to the water-column, or the image might show the thin gelatinous envelop unexpectedly broken by a strong environmental event or by a predator attack. Scale bar: 5 mm.

A fortuitous finding revealing the internal surface of the initial chamber showed the presence of a septum that separates it from the beginning of the first whorl and contributes to its elliptical or semicircular outline. The initial chamber has an average diameter of 0.21 mm (Figure 5B and Figure 7, Figure 8 and Figure 9), it is devoid of ornamentation or has very weakly marked lines. It seems to retain an organic wall, according with the data obtained from the SEM-EDS study for chemical composition (see Section 2).

Figure 5.
Size variation in the shells (a) and ammonitella stage (b) recording at the ammonoid clusters found in phosphate and siderite nodules of the San Gregorio Formation of Uruguay. Measurements of the shells were taken from the best preserved specimens in dorsal view from eight clusters (MMO 0156, 171, 215, 225, 230, 304, 305, 311), trying to cover all the represented sizes, ranging from 3 to 5 mm in average. Measurements of diameters of the ammonitella stage were mostly allowed from the best preserved specimens studied by SEM and were taken following [36,38].
Figure 5.
Size variation in the shells (a) and ammonitella stage (b) recording at the ammonoid clusters found in phosphate and siderite nodules of the San Gregorio Formation of Uruguay. Measurements of the shells were taken from the best preserved specimens in dorsal view from eight clusters (MMO 0156, 171, 215, 225, 230, 304, 305, 311), trying to cover all the represented sizes, ranging from 3 to 5 mm in average. Measurements of diameters of the ammonitella stage were mostly allowed from the best preserved specimens studied by SEM and were taken following [36,38].


Figure 6.
Comparative chemical composition and microstructure of the substance that encloses the ammonoid early juvenile specimens herein studied with respect of that of the nodule matrix containing them. (A). SEM-EDS analysis of an area of the matrix in specimen JMMO-IMT 0215. The main elements are calcium and phosphorus, with a variable content of iron, which turns the nodules as phosphate or siderite according to the amount of Fe that they contain. (B). Typical composition found in the possible gelatinous substance that encloses the clusters. (C1,C2) are SEM-EDS analyses that show the high content of carbon that is found in the mentioned substance, denoting the high preservation of soft tissues.
Figure 6.
Comparative chemical composition and microstructure of the substance that encloses the ammonoid early juvenile specimens herein studied with respect of that of the nodule matrix containing them. (A). SEM-EDS analysis of an area of the matrix in specimen JMMO-IMT 0215. The main elements are calcium and phosphorus, with a variable content of iron, which turns the nodules as phosphate or siderite according to the amount of Fe that they contain. (B). Typical composition found in the possible gelatinous substance that encloses the clusters. (C1,C2) are SEM-EDS analyses that show the high content of carbon that is found in the mentioned substance, denoting the high preservation of soft tissues.

However, original aragonite layers could have been replaced by apatite during the diagenesis as is common for preservation in phosphorite concretions. The caecum has a bulbous shape as in most goniatite cephalopods and it is positioned against the external surface of the initial chamber, near the beginning of a muscle scar. It is more or less well identified in the specimens depicted in Figure 7 and Figure 8C,D, but other structures are less clear due to the accumulation of debris produced by shell fracturing. The proseptum (1) and the second septum (2) (Figure 7A and Figure 8D) are not easily observable in most of the available materials. The prosiphon is preserved and appears as comparatively thicker and longer than the proseptum (Figure 7D). Following the initial chamber, in which a crystalline layer is not observable, the phragmocone could be at the second stage of mineralization; however, the external wall of the first whorl is covered by a single crystalline layer while the region near the umbilical area remains unmineralized. Part of the successive two whorls appears to be partially mineralized and the proseptum (1) and the second septum (2) which seem to be similar in length and positioned parallel to each other when preserved, initiate their mineralization at this stage. This aspect of the shell mineralization will need to be corroborated by a more thorough analysis involving more specimens.

Figure 7.
Microanatomy of the ammonitella stage of tiny ammonoid shells inside the clusters from the Devonian of Uruguay. (A). JMMO-IMT 0305. SEM images of one of the best-preserved shells in lateral view. The initial chamber is poorly delimited but the caecum is well identified. (B). Close up of other shell preserved in the same cluster (JMMO-IMT 0305) to show the internal morphology of the initial chamber well delimited by a septum that separates it from the beginning of the first whorl. (C,D). Other specimens from cluster JMMO-IMT 0304 also show the semicircular shape of the initial chamber and the caecum and prosiphon shapes. Abbreviations: c, caecum; ic, initial chamber; prs, proseptum; psi, prosiphon.
Figure 7.
Microanatomy of the ammonitella stage of tiny ammonoid shells inside the clusters from the Devonian of Uruguay. (A). JMMO-IMT 0305. SEM images of one of the best-preserved shells in lateral view. The initial chamber is poorly delimited but the caecum is well identified. (B). Close up of other shell preserved in the same cluster (JMMO-IMT 0305) to show the internal morphology of the initial chamber well delimited by a septum that separates it from the beginning of the first whorl. (C,D). Other specimens from cluster JMMO-IMT 0304 also show the semicircular shape of the initial chamber and the caecum and prosiphon shapes. Abbreviations: c, caecum; ic, initial chamber; prs, proseptum; psi, prosiphon.

The siphuncular tube and the siphuncle are well preserved in many individuals and are positioned marginally and ventrally. It is enclosed by the shell septa, which forms a median semicircular opening (the siphuncular lobe) that seems to be continuous through the phragmocone (Figure 9). Intriguingly, the siphuncular tube can be seen as passing through more central or marginal openings of the septa and thus it appears to shift its position even within the phragmocone of the same individual (Figure 9A), but it may be a taphonomic bias.
The ornamentation of the external shell surface consists of transverse fine ribs known as lirae, which are more prominent on the outermost whorls (Figure 8C,D and Figure 9A). Delicate lirae can also be seen on the first whorl, although only with SEM, along with a delicate transverse striate surface (Figure 10C). Towards the outer whorls, these thinned ribs become widely spaced and are well exposed in various specimens (Figure 8C,D and Figure 10A). As observed by other authors, the last two chambers are filled with organic matter, likely representing the fossilized remains of the body of the animal that inhabited them. A primary constriction or primary varix is not present or it may have been very weakly marked. The prosiphon is also observed in some specimens and it appears to be short and thick (see Figure 7D and Figure 8C,D).

Figure 8.
(A). Dorsal view of the initial chamber and the first whorl showing the position and morphology of the proseptum and the second septum (small black arrow). (B). Close up of the proseptum and second septum shown in (A). (C). Ammonitella stage morphology of a specimen found in the cluster JMMO-IMT 0305, where can be seen that the ornamentation of the shell shows a gradual change at the level of what we interpreted is the proseptum 1. (D). Interpretive drawing of (C) at the same scale. Abbreviations: c, caecum, ic, initial chamber, prs, proseptum, psi, prosiphon.
Figure 8.
(A). Dorsal view of the initial chamber and the first whorl showing the position and morphology of the proseptum and the second septum (small black arrow). (B). Close up of the proseptum and second septum shown in (A). (C). Ammonitella stage morphology of a specimen found in the cluster JMMO-IMT 0305, where can be seen that the ornamentation of the shell shows a gradual change at the level of what we interpreted is the proseptum 1. (D). Interpretive drawing of (C) at the same scale. Abbreviations: c, caecum, ic, initial chamber, prs, proseptum, psi, prosiphon.

Glaphyrites rionegrensis is the only ammonoid species known for Uruguay and it is exclusively represented in the phosphate and siderite nodules of the San Gregorio Formation [7,8,9,20,41,42]. Some similarities can be noted between the ammonitella stage of the herein described tiny ammonoids and that of Glaphyrites sp. from the Carboniferous of North America [26] particularly in the globular morphology of the caecum and possibly in the configuration of the proseptum and the second septum, although is not clear if the later was present or mineralized (Figure 7A,D and Figure 8C,D). However, the referred characters are very common in Palaeozoic ammonoids and thus differences in the shape of the initial chamber are rather more relevant. The initial chamber is apparently semicircular to elliptical in the new Uruguayan specimens (although it is not easy to discern externally), and it is rounded in Glaphyrites [26]. Another difference lies in the relative position of the caecum, which is separated from the initial chamber by the prosiphon in Glaphyrites sp., but it is very close to that chamber in the Uruguayan specimens, possibly due to the shorter length of the prosiphon and siphuncle (see Figure 7A,D and Figure 8C,D). Additionally, the configuration of the siphuncle in our specimens differs from that described by [26] for Glaphyrites, as it extends along to the outer border of the chambers in the former (Figure 10), while it is preserved separated from that border in the specimen figured by [26]. This last configuration may reflect taphonomic differences or a more effective support of the organs by the septa in the Uruguayan species.

Figure 9.
SEM images showing the morphology and configuration of the siphuncle and the siphuncular tube in the ammonitella ontogenetic stage of the best-preserved specimens in clusters from the Devonian of Uruguay. All specimens shown are part of the cluster JMMO-IMT 0311. (A). Almost complete specimen showing the siphuncle across the last whorl passing through the chambers close to the outer wall of the shell at the middle region of the chamber. Interestingly, it can be seen that the septum is deformed/opened both centrally and dorsally. (B–D). Images taken from different shells confirming the common configuration of the siphuncle by passing through the median opening formed by the septa, particularly clear in (D).
Figure 9.
SEM images showing the morphology and configuration of the siphuncle and the siphuncular tube in the ammonitella ontogenetic stage of the best-preserved specimens in clusters from the Devonian of Uruguay. All specimens shown are part of the cluster JMMO-IMT 0311. (A). Almost complete specimen showing the siphuncle across the last whorl passing through the chambers close to the outer wall of the shell at the middle region of the chamber. Interestingly, it can be seen that the septum is deformed/opened both centrally and dorsally. (B–D). Images taken from different shells confirming the common configuration of the siphuncle by passing through the median opening formed by the septa, particularly clear in (D).


Figure 10.
Ornamentation of the external surface of the ammonitella and early juvenile stages in individual shells from clusters of the Devonian of Uruguay. (A). JMMO-IMT 0304. Photograph under Camera Lucida of part of the gelatinous substance covering the shells. An almost complete and well preserved specimen displays the lirae-like ornamentation across the shell. Note that it is very fine-ribbed at the first whorl, increasing the space between ribs in direction to the aperture. Scale bar: 30 mm. (B). JMMO-IMT 0304. SEM image where both the mold and the outer ornamentation of the ammonitella shell can be appreciated. (C). JMMO-IMT 0311. SEM photograph of one of the cluster components showing the faint lirae-like ornamentation pattern characterizing the first whorls of the phragmocone, which is different from the well marked ribs observed in the outermost whorls.
Figure 10.
Ornamentation of the external surface of the ammonitella and early juvenile stages in individual shells from clusters of the Devonian of Uruguay. (A). JMMO-IMT 0304. Photograph under Camera Lucida of part of the gelatinous substance covering the shells. An almost complete and well preserved specimen displays the lirae-like ornamentation across the shell. Note that it is very fine-ribbed at the first whorl, increasing the space between ribs in direction to the aperture. Scale bar: 30 mm. (B). JMMO-IMT 0304. SEM image where both the mold and the outer ornamentation of the ammonitella shell can be appreciated. (C). JMMO-IMT 0311. SEM photograph of one of the cluster components showing the faint lirae-like ornamentation pattern characterizing the first whorls of the phragmocone, which is different from the well marked ribs observed in the outermost whorls.

3.2. Chemical Composition of the Shells in Ammonoid Clusters from Uruguay
Our SEM analyses of the strongly packed shells in the clusters yielded new data on their chemical composition (Figure 6). The analyses revealed significant differences in the matrix structure and chemical composition of the phosphate nodules with respect to these found at the clusters. Ca and P along with Fe are the most abundant components of the nodule matrix, whereas only P and Ca are present at the space surrounding the ammonoid shells, where a distinctive structure of intricate tubules interwoven within the matrix and the shells is observed (see Figure 7A). It is important to note that this particular microanatomical feature is found just in the nodules containing the ammonoid clusters, and was not yet observed surrounding other preserved fossil groups such for instance the actinopterygians, the adult ammonoids and nautiloids or around the plant remains.
4. Discussion
While several examples of clusters of purposed eggs, embryonic ammonitella, early postembryonic, and juvenile ammonoids, have been discovered in Mesozoic (Jurassic and Cretaceous) deposits [30,43,44,45], older records of early ammonoid masses, particularly from the Devonian have been less documented. Examples of the oldest found records include clusters of shells containing individuals at different stages of the ontogenetic development (e.g., ammonitellae mixed with postembryonic and juvenile specimens), and accumulations mixed with remains of other taxa, see [24,27,30,32,46]. As a result, these findings did not allow identifying reproductive strategies in ancient ammonoids [30]. In contrast, the clusters described herein comprise small ammonoid shells, all belonging to the same species and all in each cluster preserved at the same ontogenetic stage, although showing a small difference in size (Figure 5), possibly related to particular environmental or physiological conditions that may have impacted the growth rates.
As an attempt to reconstruct reproductive traits and lifestyles of Paleozoic ammonoids, the relationship between egg size and the size of the initial chamber, as well as the rates of growth of the initial chamber and the ammonitella size were highlighted in previous discussions e.g., [32,36]. However, the impact of these parameters on ammonoid evolution could not be conclusively demonstrated because patterns are not persistent and show a high variability, suggesting that the proposed evolutionary changes in the egg and ammonitella size through time may have affected only particular groups and may be linked to environmental and climate change variability [47]. Accordingly, an intraspecific size variation in ammonitellae of Cretaceous species was observed by [48], who concluded that it is due to different regional environmental factors (epigenetic effects plus selection), and should not be linked to chronological (or evolutionary) changes.
4.1. Available Evidence of Ammonoid Reproductive Strategies
Reproductive strategies in ammonoids have been hypothesized but the evidence is still weak. Putative egg-masses have been discovered e.g., [24,27] and see also [40] but the ammonoid status in such masses is uncertain see [27,30]. It has been suggested the reproduction of ammonoids as involving egg masses laid by the female suspended in a gelatinous substance secreted by special nidamental glands, based on what is known for extant cephalopod species and from the study of ammonoids found in a Lower Carboniferous locality at the United States [40]. These gelatinous masses may have dispersed for long distances by floating in the water column, or they may have been attached to floating debris as is observed in some octopuses and cuttlefishes, hypothesized [40]. The eggs hatch protected by the gelatinous masses and the embryonic ammonitellae would have been part of the plankton until they reached a diameter of 20 or 20,5 mm, when they finally acquired a necktoplantonic or necktobentonic lifestyle [40]. However, [30] found evidence to suggest ovoviviparity among other reproductive strategies for ammonoids because embryonic individuals were found in the living chamber of one adult aconeceratid ammonoid, which also might suggest some kind of parental care.
To evaluate these previous hypotheses on the reproductive behavior of early ammonoids, it is necessary to consider the strategies known for extant cephalopods which certainly, will shed light on this attempt.
According to a recent research, cephalopods have two developmental strategies: (i) they hatch as planktonic paralarvae which are morphologically different from the adults and occupy different niches during their early ontogenetic development or (ii) they hatch as juveniles that are similar to the adults and both share the same habitat [36,45]. The latter authors also highlight the fact that cephalopods possess a direct development, independently of the mode followed by the species.
4.2. Ammonoid Clusters Being Gastric Contents or Coprolites
Even suspecting a strong reproductive significance for the clusters described in this contribution, there is yet a plausible hypothesis that should be considered to explain the preservation of several masses of small ammonoids in the Uruguayan nodules. Such alternative possibility is that they could represent gastric contents, regurgitations or coprolites. Indeed, some of the features already described for our specimens lead us to dismiss this hypothesis because of the delicate preservation of the samples plus the fact that they are accumulations of individuals of a single taxon representing the same ontogenetic stage, and because the notable packaging of the shells that gives the impression that they have been enclosed by a substance or envelope at the moment of their death (Figure 4). In addition, the fracture of the shells observed in some of the clusters is mainly produced by the process of opening of the nodules. Therefore, it is highly unlikely that a predator consumes such a well-selected set of prey in terms of size and specific composition and then defecate its prey in the good condition observed in the samples.
4.3. Taxonomic Identification of the Ammonoid Clusters
We have insufficient evidence to assign the tiny specimens preserved inside the nodules to Glaphyrites rionegrensis, which is the only species previously described for the Uruguayan record. All the specimens display the same overall morphology and identical pattern of sutures (Figure 11), which is notably more consistent with the pattern described for species of the early to middle Devonian [49], suggesting that they belong to a unique species, a new one for Uruguay.
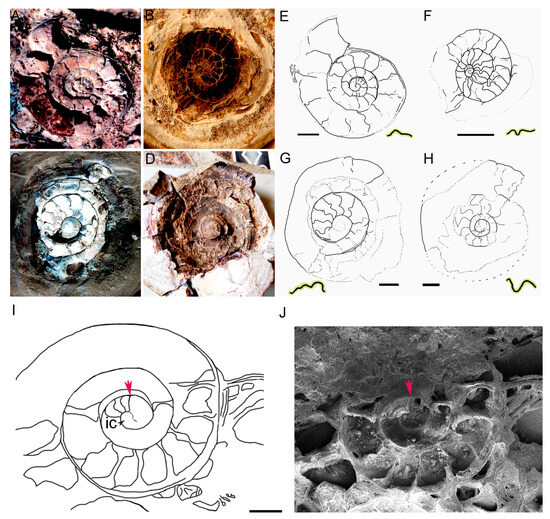
Figure 11.
Morphology of the suture in ammonoids preserved in siderite and phosphate nodules of the San Gregorio Formation, Devonian of Uruguay. (A): JMMO-IMT 0215, small ammonoid, possible early juvenile individual. (B). JMMO-IMT 0016, young adult individual. (C,D). JMMO-IMT 0092 and 0091, adult individuals. The pattern of the sutures is clearly related to the ontogenetic stage of the analyzed specimens. It seems to be not constant because the walls of the septa form three-dimensional folds that are more or less accentuated or possess more or less irregularities, depending on the height at which the specimen is sectioned. The sutures at the central chambers are not always visible, i.e., they are not always exposed if the cut does not pass exactly to the center of these sections. (E–H). Interpretive drawings of (A–D). (I). JMMO-IMT 0305. Interpretive drawing of the shell of one of the individuals that are components of the cluster showing the first four phragmocone cameral septa and the sutures for these septa, thus delimitating the end of the ammonitella shell (red arrow). (J). Photograph of the specimen drawn at I clearly showing the first septum (red arrow) delimitation the shape of the ammonitella stage. Scale bars: (E). 5 mm; (F–H). 10 mm; (J). 1 mm. See also Figure 8 as a complement.
The main differences with respect to Glaphyrites sp. from the Early Carboniferous of North America [26] are noted in the morphology of ammonitella stage of the best-preserved specimens, such as the semicircular shape of the initial chamber, which is poorly differentiated with respect to the first whorl; the slightly marked change in ornamentation from ammonitella to the beginning of the phragmocone represented by only a slightly marked lirae-type ornamentation in the first whorl with respect to the ribbed pattern observed in the outer whorls (Figure 9), the presence of a short and curved prosiphon and the apparent absence of a primary constriction and primary varix [50]. These characters have been considered as phylogenetically plesiomorphic for ammonoids (i.e., they were acquired early in the evolution of the group).
In order to investigate the taxonomic affinities of the Uruguayan ammonoids described herein we will have to test the following three hypotheses, (a) there is indeed more than one species preserved in the reworked nodules of the San Gregorio Formation, (b) the assignment to Glaphyrites of the previously described Uruguayan ammonoid specimens is wrong, and (c) if there is only one species and the taxonomic assignment to Glaphyrites is correct, it can be suggested that the biochron for this taxon would be extended back to the Devonian. All hypotheses will be analyzed in a forthcoming paper.
4.4. The Devonian Age of the Nodules from Biostratigraphic and Paleogeographic Evidence
The age of the San Gregorio Formation has remained poorly constrained due to the absence of radiometric dating in both the Norte and Paraná Basins. Paleontological records (i.e., palynological assemblages from boreholes in the eastern section of the formation at Cerro Largo Department, and the content of fossiliferous nodules at the central area, in Tacuarembó and Durazno departments) provided controversial data that need to be contrasted to the paleogeographic and paleoclimatic scenario in which the deposition of the San Gregorio Formation occurred.
Notably, in Uruguay there is a tendency of rejuvenate the Paleozoic sequences and all the deposits overlying the San Gregorio Formation (e.g., Tres Islas, Frayle Muerto and the Mangrullo formations) were placed in the Lower Permian. For instance, recent studies performed at the Mangrullo Formation attributed an Upper Carboniferous or at most a Carboniferous-Permian age to this unit e.g., [51,52,53,54], among others, which had been previously considered as early Upper Permian e.g., [55]. The Mangrullo Formation is a fossil-rich Konservat-Lagerstätte containing the remains of a continental lagoonal community documented mostly by mesosaur amniotes and pygocephalomorph crustaceans, although algal matts, insects, ichnofossils tentatively assigned to cf. Chondrites isp. and plants, are commonly found [51]. That fossil association is equivalent to Upper Carboniferous sequences of North America, mainly based on pygocephalomorph and insect biostratigraphy and the identity of the well-preserved plant remains, see [51,52].
A Carboniferous age is also suggested by the plesiomorphic features of the mesosaur skeleton [53,54,56] and their phylogenetic position as one of the basalmost amniotes [54,57].
The fossiliferous nodules, whether isolated or loosely cemented to the basal fine-to middle grained tillite conglomerates, have been clearly reworked from underlying sequences and the original unit/level where these nodules formed could have been entirely barren and eroded by the glacial movements (see Figure 2). This suggests a greater resistance of the nodules to erosion than their matrix, which, however, could have also been mixed with the glacial tillites from the base of the San Gregorio Formation.
Recent fieldwork studies also provided relevant evidence to suggest that the nodules originated in the underlying Devonian sequences of the Durazno Group that bear similar, although no yet fossiliferous nodules, which can be considered as an evidence to be better developed in future studies.
We highlight the presence of an association that includes Actinopterygians, probable and yet indeterminate acanthodians and placoderms, cephalopods, trilobites, brachiopods, corals, sponges and cf. Lycopsida as the main represented taxa (Figure 12). Rugose and tabulate corals (Figure 12E and Figure 12F, respectively) are well preserved and represent the most common fossils in the nodules (up to 70% of the sample) including not only solitary corallites but colonial complex ecosystems. These reef-building organisms usually live in temperate to warm shallow water conditions [58,59], which therefore are strongly contradictory to the glacial climates suggested for the deposition of the San Gregorio Formation [1,10], among many others.
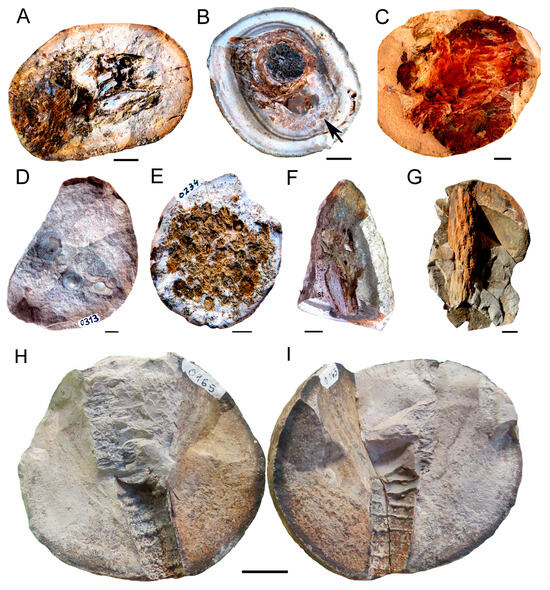
Figure 12.
Main taxa represented in the reef community preserved in siderite and phosphate nodules of the San Gregorio Formation, Devonian of Uruguay. (A). JMM-IMT 0006, Actinopterygii (skull and anterior region of the trunk). (B). JMM-IMT 0056. Ammonoidea (goniatite ammonoid adult associated to corallites of cf. Rugosa (arrow). (C). JMM-IMT 0331. Trilobita (characteristic red color and badly preserved; could be a molten exhubia). (D). JMM-IMT 0313. Brachiopoda (clusters of Orbiculoidea baini). (E). JMM-IMT 0234. Rugosa (colonial corallites). (F). JMMO-IMT 0380. Well-preserved coral specimen tentatively assigned to Tabulata, showing internal morphology of the wall (w) and trabeculate septal region (st). (G). JMM-IMT 0422 cf. Lycopsida stem or root. (H,I). JMM-IMT165. Cephalopoda Orthoceridae. Fragmentary cone-shaped shell of a nautiloid cephalopod. Scale bars: (A–C,E–G): 10 mm; (D): 3 mm; and (H–I): 20 mm.
In addition, the nodules contain goniatite cephalopods (Figure 12B), which are a basal ammonoid group commonly related to warm environments and abundant specimens of orthoceratid nautiloids (Figure 12H,I), which represent a group more frequently described as to be a component of Silurian and Devonian faunas [60]. Actinopterygians seem to be so far, the most abundant gnathostome group present in the nodules (Figure 12A). Other taxa less represented are sponges, conulariids and radiolariids [19], as well as inarticulate brachiopods assigned to cf. Orbiculoidea baini [61] (Figure 12D), a species very well represented in the underlying Devonian units, particularly in the Cordobés Formation, but recently also found by our team, as a component of the Cerrezuelo Formation [9]. Many of the mentioned taxa preserved in the nodules are in association with coral remains, showing a close temporal correspondence for the high mortality in the community and the nodule formation.
While a Lower Carboniferous age was recently suggested for the San Gregorio Formation e.g., [22], our findings suggest that the nodules cannot be younger than Middle-Upper Devonian. The fossil assemblage found within these nodules is taxonomically different from the cool Malvinokaffric fauna that dominated the marine realm during the Early Devonian in Gondwana, but a few of its characteristic taxa remains until the Middle Devonian, such as the discinid brachiopods Orbiculoidea baini and O. excentrica [62]. The Malvinokaffric community could have been affected by three ecologically significant extinction events registered at the Early Eifelian, at the Eifelian-Givetian boundary and at the Middle Givetian [63], the later possibly coincident with one of the three glaciations events suggested by the Late Devonian-Early Carboniferous of Gondwana [18]. These crises produced a gradual change in the biota composition, affecting the diversity of benthic taxa and reducing the endemism observed in the Early Devonian communities, which turned cosmopolitan before the Frasnian–Famennian mass extinction event [63]. The presence of O. baini in the nodules (Figure 12D), constrains their age to the Middle Devonian [62]. The presence of sponges was also detected with SEM and the spicules are very similar to that observed in Devonian stromatoporoids described by [64], but these materials need further studies and will be described in forthcoming papers.
The goniatite ammonoids display a suture that is typical of Devonian and Lower Carboniferous taxa e.g., [49,65,66,67] (Figure 11) and they are found in association with orthoceratid nautiloids (Figure 12H,I) as commonly observed in Lower to Middle Devonian marine communities i.e., [68]. Furthermore, the association of basal ammonoids to orthoceratids and to specimens assignable to Rugosa and Tabulata corals has been also described for Devonian sequences of Bolivia and Argentina [69], Morocco [68,70] and Uzbekistan [71], among others.
The high representation of stems and roots preliminary assigned to cf. Lycopsida (Figure 12G), are a confident evidence of the shallow-depth sea where the biota preserved in the nodules, lived. Colonial corals are another common fossils in the nodules and along with the cephalopods, they are good paleoenvironmenal and paleoclimate indicators, suggesting warm-water conditions, as those proposed for the Givetian and Late Frasnian, when temperatures could have reached 28 to 32 °C in tropical regions [72,73]; coincidently with the major reef dispersion reaching their southernmost known Devonian distribution [70]. Therefore, based on paleoclimate, paleoenvironmental and biostratigraphic evidence we propose a preliminary Devonian age for the nodule formation. Given also the excellent preservation of most of the fossils that they contain, a near synchronous massive death of the organisms and nodule formation was possible supported, because although some kind of time-averaging may have existed, it does not substantially affect our chronostratigraphic conclusions.
5. Conclusions
Masses of small spherical structures have been previously identified as putative cephalopod eggs [24] and aggregations containing high amounts of ammonoids including possible ammonitellae mixed with hatchling, juvenile and subadult individuals, and even containing other taxa such as orthoceratid nautiloids and brachiopods, are frequently found in the Devonian deposits [47]. They are interpreted as produced by some kind of catastrophic events that have produced high mass mortality in communities where these taxa were very well represented [47].
Following the interpretation of some previous authors, the clusters of tiny ammonoid shells described herein from nodules of the San Gregorio Formation are supporting the hypothesis that fossil cephalopods, particularly ammonoids, adopted the strategy of reproduce by laying the eggs in possible floating or ?fixed gelatinous masses where they hatch as morphologically similar miniatures of their parents. The clusters contain highly packed shells averaging 3 mm in diameter that suggest another life history for the ammonitellae, which are mostly considered an embryonic stage that migrate to the water column to become part of the plankton. Instead, our findings suggest that they would remain in the gelatinous envelopes at least until they grow to acquire three additional whorls, when they abandon the masses resembling the morphology of their parents, except for the smaller size. This interpretation is consistent with one of the developmental modes described by cephalopods, where hatchings are similar copies of the adults [33] and may explain the poor representation of the ammonitellae in the fossil record, which indeed should have been very abundant given the high diversity and wide geographic distribution of ammonoids in both Paleozoic and Mesozoic deposits [74]. The fact that none of the clusters herein described contain ammonitellae but hatchling or early juvenile ammonoids at the same stage of development may be indicative that the nodules preserved a mortality event that affected the community during a single reproductive season. We interpreted these findings as reproductive mass mortality (possibly involving semelparity strategies as was suggested by [44]).
On the other hand, the good number of postembryonic and juvenile ammonoid specimens reported both as isolated individuals or forming part of concentrations, which can include even adults of other taxa represented in the marine community, is more consistent with our hypothesis of ammonitellae being a transitional embryonic stage that remains inside floating or fixed gelatinous masses, where they grew to reach a morphology more similar to their progenitors. They might have become part of the community where their parents lived, adopting a similar lifestyle as was previously suggested for some extant cephalopod groups [33].
We collected enough evidence from the fossiliferous content in the nodules, as well as from biostratigraphic, paleoenvironmental and paleoclimatic data, to argue that the nodules of the San Gregorio Formation were reworked from underlying Devonian levels, possibly by glaciers movements. The fossil association represented in the nodules supports the existence of a new marine reefal assemblage not described before for the Durazno Group but containing some of the taxa, such as brachiopods and trilobites, already found in the Lower Devonian Cerrezuelo and Cordobés formations. Part of the lost levels where the nodules formed may even have been redistributed over other surrounding rock sequences, a possibility that will have to be analyzed.
The remarkably high fossiliferous potential for preservation of soft and delicate structures shown by the phosphate and siderite nodules described herein significantly enhanced our understanding of the nature and taxonomic composition of a particular shallow marine reef community that developed before the Paleozoic glaciations described for Uruguay and the Gondwanic region. This unique preservation mode provides a new hypothesis about the reproductive strategies of extinct cephalopods and facilitates the identification of various developmental ontogenetic stages. Given this exceptional quality of preservation, we propose that the phosphate and siderite nodules found at the base of the San Gregorio Formation be classified as a Konservat-Lagerstätte, following the criteria laid out by [75]. Notably, this would make it the oldest known Konservat-Lagerstätte of South America, representing a valuable scientific heritage for a small population in our country and for the overall paleontological record.
Author Contributions
Conceptualization, G.P. and P.N.D.; methodology, G.P. and P.N.D.; software, M.R.; validation, G.P., M.R. and P.N.D.; formal analysis, G.P.; investigation, G.P.; resources, G.P.; data curation, G.P.; writing—original draft preparation, G.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The specimens studied herein are part of the Juan Manuel Méndez Collection and are deposited at the Municipal De las Bochas Museum in the San Gregorio de Polanco city, Tacuarembó department, Uruguay. The original contributions presented in the study are included in the article, further inquiries can be directed to the corresponding author.
Acknowledgments
We thank Juan Manuel Méndez Ortiz for having collected the phosphate and siderite nodules along the Negro river margins. We also thank Neil Landman (Natural History Museum, New York, USA), Ana María Ribeiro (SEMA, Brasil) and Darío Lazo (University of Buenos Aires, Argentina) for providing helpful bibliography. Neil Landman, Christian Klug and Kenneth De Baets revised the manuscript, and Isabelle Kruta, Alan Pradel and Stephen Kershaw are thanked for interesting discussions on the subject of this work. Michel Laurin is warmly thanked for English language revision and for his valuable criticism on basic aspects of this article. Two anonymous reviewers made valuable contributions that highly improved the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Beri, Á.; Martínez-Blanco, X.; Mourelle, D. A synthesis of palynological data from the Lower Permian Cerro Pelado Formation (Paraná Basin, Uruguay); A record of warmer climate stages during Gondwanan glaciations. Geol. Acta 2010, 8, 419–429. [Google Scholar] [CrossRef]
- De Santa Ana, H.; Beri, A.; Goso, C.; Daners, G. Análisis estratigráfico de la Formación San Gregorio (Pérmico Inferior), en los testigos del pozo Cerro Largo Sur Nº4 (Di.Na.Mi.Ge.), Uruguay. Rev. Bras. Geociências 1993, 23, 347–351. [Google Scholar] [CrossRef]
- Closs, D.; Gordon, M.J. An upper Paleozoic goniatite radula. Esc. Geol. Notas E Estud. 1966, 1, 73–75. [Google Scholar]
- Beltán, L. Dècuverte d’une ichtyofaune dans le Carbonifère Superieur d’Uruguay. Rapport avec les faunes ichthyologiques contemporaines des autres régions du Gondwana. Ann. Société Géologique du Nord. Lille 1977, 97, 351–355. [Google Scholar]
- Beltán, L. Coccocephalithchys tessellatus n. sp. (Pisces, Actinopterygii) from the Upper Carboniferous of Uruguay. An. Do II Congr. Lat.-Am. Paleontol. 1981, 1, 95–105. [Google Scholar]
- Beri, A.; Cernuschi, F.; Martínez-Blanco, X. Palynology of the San Gregorio and Melo Formations in Borehole 254 (Dinamige), Late Paleozoic, Paraná Basin, Uruguay. Ameghiniana 2011, 48, 480–495. [Google Scholar] [CrossRef]
- Closs, D. Goniatiten mit Radula und Kieferapparat in der Itararé Formation von Uruguay. Paläontologische Z. 1967, 41, 19–37. [Google Scholar]
- Closs, D. Upper Carboniferous anaptychi from Uruguay. Ameghiniana (Rev. Asoc. Paleontológica Argent.) 1967, 5, 145–148. [Google Scholar]
- Piñeiro, G.; Estramil, C.; Méndez Ortiz, J.M.; Núñez Demarco, P. First evidence of reproductive strategies in Devonian cephalopods preserved in siderite nodules from Uruguay. In Proceedings of the IV Palaeontological Virtual Congress, Thematic Section: Palaeontological Scenarios of the Paraná Basin, Virtual, 1–22 May 2023. [Google Scholar]
- De Santa Ana, H.; Goso, C.; Daners, G. Cuenca Norte: Estratigrafía del Carbonífero-Pérmico. In Cuencas Sedimentarias de Uruguay: Geología, Paleontología y Recursos Naturales; Veroslavsky, G., Ubilla, M., Martínez, S., Eds.; Facultad de Ciencias de Uruguay (DIRAC): Montevideo, Uruguay, 2006; pp. 147–208. [Google Scholar]
- Bossi, J. Geología del Uruguay. In Colección Ciencias, 2nd ed.; Universidad de la República: Montevideo, Uruguay, 1966; 419p. [Google Scholar]
- Caorsi, J.; Goñi, J. Geología Uruguaya. Inst. Geológico Urug. Boletín Montev. 1958, 37, 1–73. [Google Scholar]
- Fedorchuk, N.D.; Griffisc, N.P.; Isbell, J.L.; Goso CRosa, E.L.M.; Montañez, I.P.; Yinc, Q.Z.; Huyskensc, M.H.; Sanbornc, M.E.; Mundile, R.; Vesely, F.F.; et al. Provenance of late Paleozoic glacial/post-glacial deposits in the eastern Chaco-Paraná Basin, Uruguay and southernmost Paraná Basin, Brazil. J. S. Am. Earth Sci. 2021, 106, 102989. [Google Scholar] [CrossRef]
- Martin, H. The Late Palaeozoic Gondwana glaciations. Geol. Rundsch. 1981, 70, 488–496. [Google Scholar] [CrossRef]
- Sprechmann, P.; Montaña, J.; Gaucher, C. Devónico. In Geología del Departamento de Durazno; Boletin de Facultad de Agronomía: Montevideo, Uruguay, 1993; pp. 25–55. [Google Scholar]
- Uriz, N.J.; Cingolani, C.A.; Basei, M.A.S.; Blanco, G.; Abre, P.; Potillo, N.S.; Sicardi, A. Provenance and paleogeography of the Devonian Durazno Group, southern Parana Basin in Uruguay. J. S. Am. Earth Sci. 2016, 66, 248–267. [Google Scholar] [CrossRef]
- Griffis, N.P.; Mundil, R.; Montañez, I.P.; Isbell, J.L.; Fedorchuk, N.; Vesely, F.; Iannuzzi, R.; Yin, Q.Z. A new stratigraphic framework built on U-Pb single-zircon TIMS ages and implications for the timing of the penultimate icehouse (Paraná Basin, Brazil). GSA Bull. 2018, 130, 848–858. [Google Scholar] [CrossRef]
- Césari, S.; Limarino, C.O.; Gulbranson, E.L. An Upper Paleozoic bio-chronostratigraphic schem for the western margin of Gondwana. Earth-Sci. Rev. 2011, 106, 149–160. [Google Scholar] [CrossRef]
- Braun, A.; Sprechmann, P.; Gaucher, C. Stratigraphic age of phosphorite-nodules from the San Gregorio Formation of Uruguay. Neues Jahrb. Geol. Paläontologie Monatshefte 2003, 12, 739–748. [Google Scholar] [CrossRef]
- Pirez, E.; Méndez, J.M.; Estramil, C.; Piñeiro, G. El Río Hum (atual Río Negro) antes y después de la Represa de Rincón del Bonete en San Gregorio de Polanco (Tacuarembó, Uruguay). J. Anim. Environ. Res. (Edición Espec.) 2022, 5, 131–155. [Google Scholar]
- Isbell, J.L.; Henry, L.C.; Gulbranson, E.L.; Limarino, C.O.; Fraiser, M.L.; Koch, Z.J.; Ciccioli, P.L.; Dineen, A.A. Glacial paradoxes during the late Paleozoic ice age: Evaluating the equilibrium line altitude as a control on glaciation. Gondwana Res. 2012, 22, 1–19. [Google Scholar] [CrossRef]
- Iannuzzi, R.; di Pasquo, M.M.; Vesely, F.F.; Scherer, C.M.; Andrade, L.S.; Mottin, T.; Kifumbi, C. Pennsylvanian glacial cycles in western Gondwana: An overview. Geol. Soc. Lond. Spec. Publ. 2023, 535, 101–116. [Google Scholar] [CrossRef]
- Drushchits, V.V.; Doguzhayeva, L.A.; Mikhaylova, I.A. The structure of the ammonitella and the direct development of ammonites. Palaeontol. J. 1977, 2, 188–199. [Google Scholar]
- Etches, S.; Clarke, J.; Callomon, J. Ammonite eggs and ammonitellae from the Kimmeridge Clay Formation (Upper Jurassic) of Dorset, England. Lethaia 2009, 42, 204–217. [Google Scholar] [CrossRef]
- Landman, N.H.; Tanabe, K.; Shigeta, Y. Ammonoid embryonic development. In Ammonoid Paleobiology; Landman, N.H., Tanabe, K., Eds.; Springer: Boston, MA, USA, 1996; Volume 67, pp. 361–437. [Google Scholar]
- Landman, N.H.; Mapes, R.H.; Tanabe, K. Internal features of the internal shell of Late Carboniferous Goniatitina. In Advanced Research on Living and Fossil Cephalopods; Rodríguez-Tovar, O., Ed.; Kluver Academic/Plenum Publishers: New York, NY, USA, 1999; pp. 242–254. [Google Scholar]
- Tanabe, K.; Landman, N.H.; Mapes, R.H.; Faulkner, C.J. Analysis of a Carboniferous embryonic ammonoid assemblage, implications for ammonoid embryology. Lethaia 1993, 26, 215–224. [Google Scholar] [CrossRef]
- De Baets, K.; Landman, N.H.; Tanabe, K. Ammonoid embryonic development. In Ammonoid Paleobiology: From Anatomy to Ecology; Klug, C., Korn, D., De Baets, K., Kruta, I., Mapes, R.N., Eds.; Springer: Amsterdam, The Netherlands, 2015; pp. 113–205. [Google Scholar]
- Erben, H.K. Bactritoidea. In Treatise on Invertebrate Paleontology, Part K, Mollusca 3, Cephalopoda; Moore, R.C., Ed.; Geological Society of America and University of Kansas Press: Lawrence, Kansas, 1964; pp. K491–K519. [Google Scholar]
- Mironenko, A.A.; Rogov, M.A. First direct evidence of ammonoid ovoviviparity. Lethaia 2016, 49, 245–260. [Google Scholar] [CrossRef]
- Westermann, G.E.G. Ammonoid life and habitat. In Ammonoid Paleobiology; Landman, N.H., Tanabe, K., Davis, R.A., Eds.; Topics in Geobiology; Plenum Press: New York, NY, USA, 1996; Volume 13, pp. 607–707. [Google Scholar]
- Laptikhovsky, V.; Nikolaeva, S.; Rogov, M. Cephalopod embryonic shells as a tool to reconstruct reproductive strategies in extinct taxa. Biol. Rev. 2017, 93, 270–283. [Google Scholar] [CrossRef] [PubMed]
- Vidal, E.A.; Shea, E.K. Cephalopod ontogeny and life cycle patterns. Frontiers of Marine Sciences. Sec. Mar. Biol. 2023, 10, 1162735. [Google Scholar] [CrossRef]
- Landman, N.H. Preserved ammonitellas of Scaphites (Ammonoidea, Ancyloceratina). Am. Mus. Novit. 1985, 2815, 1–10. [Google Scholar]
- Lukeneder, A. Ammonoid habitats and life history. In Ammonoid Paleobiology: From Anatomy to Ecology; Klug, C., Korn, D., De Baets, K., Kruta, I., Mapes, R.H., Eds.; Springer: Amsterdam, The Netherlands, 2015; pp. 689–791. [Google Scholar]
- Wani, R. Geological duration of ammonoids controlled their geographical range of fossil distribution. PeerJ 2017, 5, e4108. [Google Scholar] [CrossRef][Green Version]
- Laptikhovsky, V.L.; Rogov, M.A.; Nikolaeva, S.E.; Arkhipkin, A.I. Environmental impact on ectocochleate cephalopod reproductive strategies and the evolutionary significance of cephalopod egg size. Bull. Geosci. 2013, 88, 83–93. [Google Scholar] [CrossRef][Green Version]
- Tanabe, K.; Sasaki, T.; Mapes, R.H. Soft-part anatomy of the siphuncle in ammonoids. In Ammonoid Paleobiology: From Anatomy to Ecology; Klug, C., Korn, D., De Baets, K., Kruta, I., Mapes, R.H., Eds.; Springer: Amsterdam, The Netherlands, 2015; pp. 531–544. [Google Scholar] [CrossRef]
- Vidal, E.A.; Haimovici, M. Feeding and the possible roll of the proboscis and mucus cover in the ingestion of microor ganisms by Rhynchoteuthion paralarvae (Cephalopoda: Ommastrephidae). Bull. Mar. Sci. 1998, 63, 305–316. [Google Scholar]
- Mapes, R.H.; Nützel, A. Late Palaeozoic mollusc reproduction: Cephalopod egg-laying behavior and gastropod larval palaeobiology. Lethaia 2009, 42, 341–356. [Google Scholar] [CrossRef]
- Closs, D. Intercalation of Goniatites in the Gondwanic Glacial Beds of Uruguay. Gondwana Stratigraphy. IUGS Symposium Buenos Aires, 1–15 October 1967; Paris Unesco, 1969. pp. 197–212. Available online: https://catalog.library.vanderbilt.edu/discovery/fulldisplay/alma991017205269703276/01VAN_INST:vanui (accessed on 18 June 2024).
- Lhemann, J.; Klug, C.; Wild, F. Did ammonoids possess opercula? Reassessment of phosphatized soft tissues in Glaphyrites of the Carboniferous of Uruguay. Palaeontol. Z. 2014, 89, 63–77. [Google Scholar] [CrossRef]
- Kulicki, C.; Doguzhaeva, L.A. Development and calcification of the ammonitella shell. Acta Palaeontol. Pol. 1994, 39, 17–44. [Google Scholar]
- Stephen, D.A.; Bylund, K.G.; García, P.; McShinsky, R.D.; Carter, H.J. Taphonomy of dense concentrations of juvenile ammonoids in the Upper Cretaceous Mancos Shale, east-central Utah, USA. Geobios 2012, 45, 121–128. [Google Scholar] [CrossRef]
- Stinnesbeck, W.; Frey, E.; Zell, P. Evidence for semi-sessile early juvenile life history in Cretaceous ammonoids. Palaeogeogr. Palaeoclimatol. Palaeocol. 2016, 457, 186–194. [Google Scholar] [CrossRef]
- Klug, C.; De Baets, K.; Korn, D. Exploring the limits of morphospace: Ontogeny and ecology of late Viséan ammonoids from the Tafilalt, Morocco. Acta Palaeontol. Pol. 2016, 61, 1–14. [Google Scholar] [CrossRef]
- Greif, M.; Nebelsick, J.H.; Klug, C. Extreme abundance of ammonoids in mass accumulations from the Late Devonian of the Moroccan Anti-Atlas. Acta Palaeontol. Pol. 2022, 67, 667–684. [Google Scholar] [CrossRef]
- Tajika, A.; Wani, R. Intraspecific variation of hatchling size in Late Cretaceous ammonoids from Hokkaido, Japan: Implication for planktic duration at early ontogenetic stage. Lethaia 2011, 44, 287–298. [Google Scholar] [CrossRef]
- Korn, D. A key for the description of Paleozoic ammonoids. Foss. Rec. 2010, 13, 5–12. [Google Scholar] [CrossRef]
- De Baets, K.; Klug, C.; Korn, D.; Landman, N.H. Early evolutionary trends in ammonoid embryonic development. Evolution 2012, 66, 1788–1806. [Google Scholar] [CrossRef] [PubMed]
- Piñeiro, G.; Ramos, A.; Goso, C.; Scarabino, F.; Laurin, M. Unusual environmental conditions preserve a Permian mesosaur-bearing Konservat- Lagerstätte from Uruguay. Acta Palaeontol. Pol. 2012, 57, 299–318. [Google Scholar] [CrossRef]
- Calisto, V.; Piñeiro, G. A large cockroach from the mesosaur-bearing Konservat-Lagerstätte (Mangrullo Formation), Late Paleozoic of Uruguay. PeerJ 2019, 7, e6289. [Google Scholar] [CrossRef]
- Piñeiro, G.; Demarco, P.N.; Meneghel, M.D. The ontogenetic transformation of the mesosaurid tarsus: A contribution to the origin of the primitive amniotic astragalus. PeerJ 2016, 4, e2036. [Google Scholar] [CrossRef]
- Núñez Demarco, P.; Ferigolo, J.; Piñeiro, G. Isometry in mesosaurs: Implications for growth patterns in early amniotes. Acta Palaeontol. Pol. 2022, 67, 509–542. [Google Scholar] [CrossRef]
- Bossi, J.; Navarro, R. Geología del Uruguay; Departamento de Publicaciones, Universidad de la República: Montevideo, Uruguay, 1991; 970p. [Google Scholar]
- von Huene, F. Osteologie und systematische Stellung von Mesosaurus. Palaeontographica 1941, 92, 45–58. [Google Scholar]
- Laurin, M.; Piñeiro, G. A reassessment of the taxonomic position of mesosaurs, and a surprising phylogeny of early amniotes. Front. Earth Sci. 2017, 5, 88. [Google Scholar] [CrossRef]
- Bridge, T.C.L.; Baird, A.H.; Pandolfi, J.M.; McWilliam, M.J.; Zapalski, M.K. Functional consequences of Palaeozoic reef collapse. Sci. Rep. 2022, 12, 1386. [Google Scholar] [CrossRef] [PubMed]
- Zapalski, M.K.; Baird, A.H.; Bridge, T.; Jakubowicz, M.; Daniell, J. Unusual shallow water Devonian coral community from Queensland and its recent analogues from the inshore Great Barrier Reef. Coral Reefs 2021, 40, 417–431. [Google Scholar] [CrossRef]
- Dzik, J. Phylogeny od the Nautiloidea. Paleontol. Pol. 1984, 45, 323. [Google Scholar]
- Sharpe, D. Descriptions of Paleozoic Mollusca from South Africa. Trans. Geol. Soc. Lond. 1856, 2, 206–215. [Google Scholar]
- Carbonaro, F.A.; Comniskey, J.C.; Corral Stok, H.; Pirani Ghilardi, R. Orbiculoidea baini and Orbiculoidea excentrica (Brachiopoda, Discinidae) from the Middle Devonian (Alto Garças Sub-basin, Paraná Basin) of Caiapônia, Goiás (Brazil). Rev. Inst. Geociências—USP Série Cient. 2018, 18, 11–20. [Google Scholar] [CrossRef]
- Penn-Clarke, C.R.; Harper, D.A.T. The rise and fall of the Malvinoxhosan (Malvinokaffric) bioregion in South Africa: Evidence for Early-Middle Devonian biocrises at the South Pole. Earth Sci. Rev. 2023, 246, 104595. [Google Scholar] [CrossRef]
- Da Silva, A.-C.; Kershaw, S.; Boulvain, F.; Hubert, B.; Mistiaen, B.R.; Reynolds, A.; Reitner, J. Indigenous demosponge spicules in a Late Devonian stromatoporoid basal skeleton from the Frasnian of Belgium. Lethaia 2014, 47, 365–375. [Google Scholar] [CrossRef]
- House, M.R. Devonian goniatites from Nevada. Newes Jahrb. Geol. Paläontologie Abh. 1965, 122, 339–342. [Google Scholar]
- Klug, C. Quantitative stratigraphy and taxonomy of late Emsian and Eifelian ammonoids of the eastern Anti-Atlas (Morocco). Cour. Forschungsinstitut Senckenberg 2002, 238, 1–109. [Google Scholar]
- Becker, R.T.; Hartenfels, S.; Klug, C.; Aboussalam, Z.S.; Afhüppe, L. The cephalopod-rich Famennian and Tournaisian of the Aguelmous Syncline (southern Maïder). Münstersche Forschungsberichte Geol. Paläontologie 2018, 110, 273–306. [Google Scholar]
- Becker, R.T.; House, M.R. Emsian and Eifelian ammonoid succession at Bou Tchrafine (Tafilalt platform, Anti-Atlas, Morocco). Notes Mémoires Serv. Geol. 2000, 399, 21–26. [Google Scholar]
- Carrera, M.G.; Montoya, E.; Rustán, J.J.; Halpern, K. Silurian-Devonian coral associations across a sequence stratigraphic boundary in the Argentine Precordillera. Geol. J. 2013, 48, 256–269. [Google Scholar] [CrossRef]
- Majchrzyk, A.; Jakubowicz, M.; Berkowski, B.; Król, J.J.; Zato, M.; Zapalski, M.K. Modern-type reef in ancient time—Palaeoecology of a Middle Devonian coral community from Madène el Mrakib (Anti-Atlas, Morocco). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2024, 633, 111876. [Google Scholar] [CrossRef]
- Naglik, C.; De Baets, K.; Klug, C. Early Devonian ammonoid faunas in the Zeravshan Mountains (Uzbekistan and Tadjikistan) and the transition from a carbonate platform setting to pelagic sedimentation. Bull. Geosci. 2019, 94, 337–368. [Google Scholar] [CrossRef]
- Zapalski, M.K.; Nowicki, J.; Jakubowicz, M.; Berkowski, B. Tabulate corals across the Frasnian/Famennian boundary: Architectural turnover and its possible relation to ancient photosymbiosis. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2017, 487, 416–429. [Google Scholar] [CrossRef]
- Scotese, C.R.; Song, H.; Mills, B.J.; van der Meer, D.G. Phanerozoic paleotemperatures: The earth’s changing climate during the last 540 million years. Earth Sci. Rev. 2021, 215, 103503. [Google Scholar] [CrossRef]
- Becker, R.T.; Kullmann, J. Paleozoic ammonoids in space and time. In Ammonoid Paleobiology; Landman, N.H., Davis, R.A., Tanabe, K., Eds.; Topics in Geobiology; Plenum Press: New York, NY, USA, 1996; Volume 13, pp. 711–753. [Google Scholar]
- Seilacher, A. Begriff und Bedeutung der Fossil−Lagerstätten. Neues Jahrb. Geol. Paläontologie Monatshefte 1970, 1970, 34–39. [Google Scholar]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).