Abstract
This study employs Principal Component Analysis to examine cranial measurements from both extant and fossil specimens spanning 38 species and comprising over 300 individuals within the subfamily Cervinae. Our findings elucidate craniometric distinctions among cervids characterized by varying body sizes and certain evolutionary adaptations. While our results generally corroborate previous assertions regarding the limited craniometric variability among plesiometacarpal deer, they also unveil specific cranial adaptations within certain genera and species. The Principal Component Analysis of craniometric data revealed that giant and large-sized deer display significantly broader ecomorphological diversity in cranial shape compared to small-sized deer. Secondly, small-sized deer exhibit greater uniformity in their cranial shape, appearing densely clustered on the factorial map. Thus, we conclude that body size imposes ecological constraints, limiting the available niches due to eco-physiological factors. This study demonstrates that endemic insular deer do not evolve consistent craniometric features attributable to insular isolation, while the cranial proportions of medium-to-small-sized deer delineate a ubiquitous morphological archetype characteristic of numerous cervid taxa spanning diverse phylogenetic lineages and sharing comparable body sizes. This group of “Dama-like” deer, characterized by similar body size, metabolic rates, ecological needs, and cranial morphometry, is referred to here as the fundamental eco-physiological type, typical of warm regions within the Palearctic and Oriental biogeographic realms.
1. Introduction
Cranial material holds crucial importance in the taxonomy and systematics of cervids, as it provides most of the essential characteristics used in zoological classification. The implication of craniometrics in cervid taxonomy generally carries a more nuanced meaning, as cranial morphology in deer, unlike in other ruminants, has consistently been considered conservative and less diversified [1,2,3,4]. It was commonly assumed that cervids followed the evolutionary trend typical of ruminant herbivores, transitioning from small-sized forms with a relatively long braincase and short splanchnocranium to larger forms with a shortened braincase and elongated splanchnocranium [4,5,6].
Vislobokova [2] delineated general evolutionary trends in cervid cranial shape at the subfamily level, noting an increase in splanchnocranium length and neurocranial shortening from the most primitive cervids (subfamilies Dremotheriinae and Lagomerycinae) to the most advanced deer (Cervinae and Capreolinae). Mejaard and Groves [7] cautiously suggested that elements of skull shape, as determined by morphometrics, could be used to reveal phylogenetic relationships among deer species within the subfamily Cervini, although they acknowledged that the use of this type of data might be challenging due to the broad size variation within a species.
Lowe and Gardiner [8] endeavored to utilize craniometric data to distinguish subspecies of red deer. However, their findings did not support the concept of subspeciation in Red deer based on the measured skull features. Put simply, they found no significant differences or discontinuities among the various subspecies sampled.
In a separate attempt, Markov and Danilkin [9] proposed the use of absolute and relative craniometric measurements to differentiate modern subspecies of Eurasian elaphine deer (Cervus elaphus and Cervus canadensis). However, their conclusions were contradicted by molecular phylogenetic data [10,11]. Consequently, the value and significance of craniometric data in cervids remain unclear. It is uncertain to what extent these data can be utilized in cervid taxonomy and systematics. Additionally, it is unknown whether cervid craniometric data can provide insights into cranial ecomorphology, antler weight, or specific evolutionary processes, such as adaptation to insular environments, often considered a form of specific insular “paedomorphosis” involving general changes in cranial proportions [12,13].
The current study proposes the utilization of Principal Component Analysis to explore morphometric cranial diversity among deer of the subfamily Cervinae, employing craniometric data. It aims to elucidate the potential significance of cranial measurements in cervid taxonomy, systematics, ecomorphology, and the description of evolutionary processes. The focus of this study is directed towards the subfamily Cervinae due to its representation by a nearly intact initial evolutionary radiation, unlike the Capreolinae, and its abundant availability of cranial material in the fossil record [4].
This study aims to test several hypotheses on the evolutionary tendencies of cranial morphology and proportions previously proposed in scientific literature:
- (I)
- The hypothesis of general uniformity of cranial morphology within the subfamily Cervinae [4,5]. The PCA analysis is the most suitable research method in this case.
- (II)
- (The hypothesis of a universal evolutionary pattern in cranial modifications in insular deer (testing the uniformity of the ‘island rule’ in plesiometacarpal deer) [12,13].
- (III)
- The hypothesis of a gradual increase in the relative length of the splanchnocranium in deer from the most primitive to the most advanced forms [2,4,5].
The subfamily Cervinae is particularly suitable for this study as it provides a rich cranial material from very small tropical forest ‘hiders’ (as per the terms introduced by Geist [4]), such as Muntiacus, to open landscape giants such as Megaloceros and Praemegaceros. This systematic diversity of modern plesiometacarpal deer represents a practically intact evolutionary diversification and includes forms at various stages of evolutionary specialization, while the fossil record of the subfamily Cervinae is exceptionally rich [2,4,5]. The subfamily Cervinae also provides numerous well-documented instances of evolution under insular isolation conditions [12,13].
Special attention is devoted to the comparative craniometry of the most specialized representatives of cervines included in this study. This focus aims to facilitate a comprehensive analysis of cranial morphology specialization within a taxonomic group traditionally considered conservative in cranial morphology.
2. Materials and Methods
The cranial material comprises over 400 skulls of both modern and fossil representatives of the subfamily Cervinae. The fossil and modern osteological specimens utilized in this study are curated at various institutions, including the Institute of Zoology of the State University of Moldova (IZM), Republic of Moldova; the Institute of Zoology of the University of Wroclaw (IZW), Poland; the Museum of Geology and Paleontology of the University of Florence (MGUF), Italy; the Muséum national d’Histoire naturelle (MNHN), Paris, France; the Museum of Paleontology of the Department of Earth Sciences (MPS), University of Rome “Sapienza”, Italy; the Natural History Museum “Grigore Antipa” (NHMB), Bucharest, Romania; the Natural History Museum of London (NHML), United Kingdom; the Natural History Museum “La Specola” (NHMF), University of Florence, Italy; the Paleontological Museum of the University “Claude Bernard” Lyon 1 (PMUL), France; the Faculty of Geography and History of the University of Barcelona (FGUB); and the Stuttgart State Museum of Natural History (SMNS), Germany.
Some cranial measurements of Cervus elaphus are adapted from Heptner and Tzalkin [14], cranial measurements of Candiacervus ropalophorus are adapted from Schilling and Roessner [15], data on Cervus elaphus siciliae are adapted from Gliozzi et al. [16], while data on Praedama giulii are adapted from Breda et al. [17]. Detailed information regarding cranial specimens, their collection numbers, institutions of curation, and measurements can be found in the Supplementary Materials (Table S1).
The following cranial measurements were obtained (Figure 1):
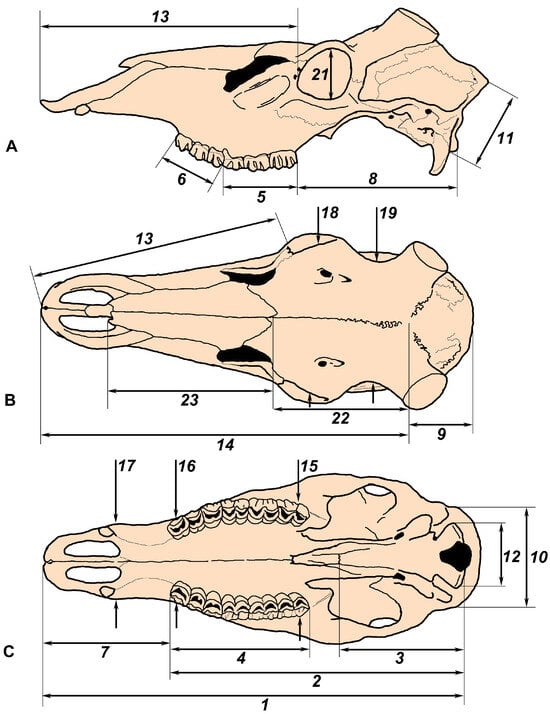
Figure 1.
The scheme of cranial measurements used in the present study shown of a skull of Cervus elaphus maral: (A,B) side and dorsal views (skull drawings adapted from Flerov [6]); (C) palatal view (adapted from Sokolov [18]).
- (1)
- Condylobasal length of the skull (CBL): Measured from the prosthion point to the posterior edges of the occipital condyles;
- (2)
- Condylodental length of the skull (L_P2_occiput): Measured from the anterior edge of the P2 alveolus to the posterior edges of the occipital condyles;
- (3)
- Length of basioccipitale (L_basiocc): Measured from the posterior edge of the occipital condyles to the suture between basioccipital and basisphenoid;
- (4)
- Length of the upper tooth row (L_P2_M3): Measured from the anterior edge of the P2 crown to the posterior edge of the M3 crown;
- (5)
- Length of the upper molar series (L_M1_M3): Taken as the maximal value from the anterior edge of the M1 crown to the posterior edge of the M3 crown;
- (6)
- Length of the upper premolar series (L_P2_P4): Taken as the maximal value from the anterior edge of the P2 crown to the posterior edge of the P4 crown;
- (7)
- Length of the anterior part of the skull before the tooth row (L_P2_prosth): The distance between the anterior edge of the alveolus of P2 and the prosthion point;
- (8)
- Length of the posterior part of the skull (L_M3_occ_cond): The distance between the anterior edge of the alveolus of M3 and the posterior edge of the occipital condyles;
- (9)
- Length of the braincase (L_bregma_inion): The distance between the bregma point and the inion point;
- (10)
- Occipital breadth (W_occiput): Measured as the maximum breadth of the occiput;
- (11)
- Occipital height (H_occiput): Measured from the basicranium to the opisthocranion;
- (12)
- Breadth of occipital condyles (W_occ_cond);
- (13)
- Length of face (L_orbit_prosth): Measured from the anterior edge of the orbits to the prosthion;
- (14)
- Length of the splanchocranium (L_bregma_prosth): Measured from the bregma to the prosthion;
- (15)
- Skull breadth at M3 (W_at_M3): The measurement taken between the alveolar edges in the middle of the right and left M3;
- (16)
- Skull breadth at P2 (W_at_P2): The measurement taken between the alveolar edges in the middle of the right and left P2;
- (17)
- Breadth of the snout (W_rostrum): Measured behind the upper canines or at the suture between the maxillary and premaxillary bones;
- (18)
- Breadth above orbits (W_at_orbits): Measured above the orbital centers;
- (19)
- Frontal breadth (W_frontal; in males only): Measurement taken at the frontal constriction before the pedicles;
- (20)
- Breadth of the neurocranium (W_braincase): Measurement taken behind the pedicles in males and as the maximal breadth of the braincase in females;
- (21)
- Diameter of the orbit (D_orbit; vertical);
- (22)
- Length of the frontal suture (L_front_sut): From bregma to nasion;
- (23)
- Length of the nasalia (L_nasal): From nasion to rhinion.
Some skulls, particularly fossils, have varying proportions of missing data (Figure S1). The majority of specimens analyzed in this study are from male individuals, with female specimens included only when their measurements provided valuable insights minimally influenced by sexual dimorphism. Mean values of cranial measurements for species and subspecies (for instance, in the case of red deer Cervus elaphus, which is represented by subspecies with significantly different body sizes) were utilized for analysis (Figure S2). While certain species are represented by only a single specimen or a small sample size, the behavior of data from these cases within the context of the entire dataset suggests that this limitation has minimal impact on this study’s results. The correlation matrix, based on all cranial material, both complete and fragmentary, demonstrates, with few exceptions, a very high correlation among cranial measurements (Figure 2).
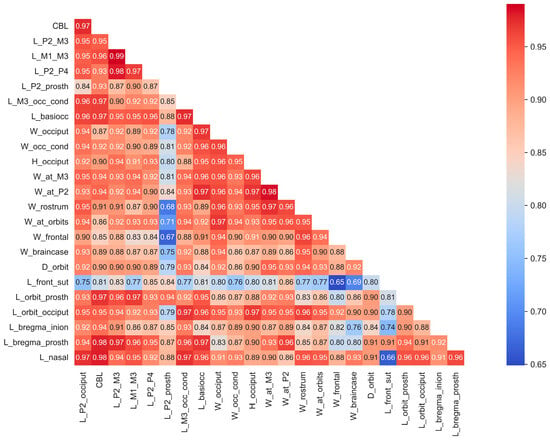
Figure 2.
Correlations of craniometrical data in the studied total sample of cranial material before missing data imputation.
Considering the known or estimated body mass of deer species categorized within these informal groups, it can be inferred that the body size class of “small-sized deer” encompasses deer with a body size comparable to that of modern Muntiacus, hog deer, and fallow deer, i.e., less than 90 kg (approximately corresponding to a condylobasal length of 160 to 300 mm) [1,4,6,19]. The body size class of “medium-small deer” comprises deer with a body size similar to that of sika deer and Eld’s deer, as well as small subspecies of red deer, ranging from 90 to 175 kg (with a condylobasal length of 300–350 mm) [1,4,14,16,19]. The size class of “large-sized deer” includes modern rusa deer, large subspecies of red deer, and wapiti, with body masses ranging from 175 to 350 kg (and condylobasal lengths ranging from 350 to 450 mm) [4,14,16,19]. The size class of “giant deer” encompasses species with a body mass above 350 kg and a condylobasal length of the skull exceeding 450 mm [4,20,21].
The body mass of extinct deer species was estimated based on craniodental variables, employing the methodology proposed by Janis [22,23].
Due to the high degree of correlation among most cranial measurements, linear and multilinear regressions were employed to estimate missing measurements in certain specimens or samples. A step-by-step imputation strategy was implemented: initially, data were imputed for measurements with fewer missing values, followed by those with the highest number of missing data points. This sequential approach to imputation was based on data completeness and the functional or topological relationships among measurements. Overall, the imputation process preserved the initial structure of relationships within the data (Figure 3).
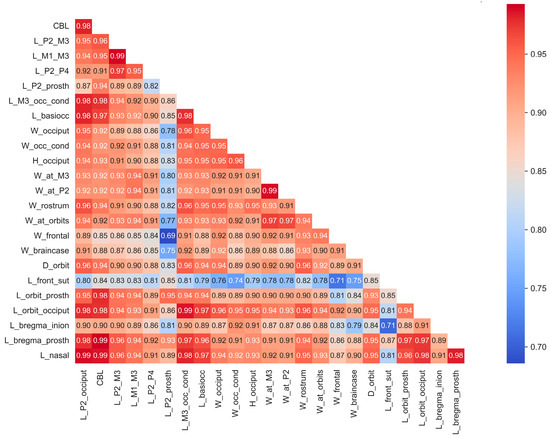
Figure 3.
Correlations of craniometrical data in the studied total sample of cranial material after missing data imputation.
The significant correlation observed among cranial measurements within the subfamily Cervinae was the key factor driving the decision to narrow the scope of this study exclusively to that subfamily. Members of the Capreolinae subfamily are noted for developing more distinct cranial morphological adaptations [4], posing potential challenges to the predictive accuracy of linear and multilinear regressions.
For further analysis, only species that retained more than 50% of their cranial measurements were included in this study. The selected deer species encompass various evolutionary specializations, including extinct giant forms (Megaloceros giganteus and Praemegaceros obscurus), modern and extinct Palearctic species, less specialized modern species from the Oriental zoogeographic realm, and modern and extinct insular deer (Rusa timorensis, Cervus elaphus siciliae, Praemegaceros cazioti, Candiacervus ropalophorus). Some species, such as Praemegaceros verticornis, were included despite having less complete original data to assess their position among other genera and species. For data analysis, Principal Component Analysis (PCA) was utilized as the most effective multivariate analysis in this context (Figures S3 and S4). The data include species represented by large samples (more than 10 individuals), smaller samples (5–10 individuals), and a certain number of species (mostly extinct) that are represented by a single individual. All data are transformed by applying the normalization, which brings the variation within the samples to the same scale. Dimensionality reduction influences the variation within a sample by reducing the number of variables while attempting to retain as much important variation as possible. The key impact is a trade-off: retaining significant variation while potentially losing less important or noisy variation. By reducing the number of dimensions, dimensionality reduction can help eliminate noise and redundant features. This can lead to a clearer understanding of the underlying structure of the data. Therefore, it is difficult to estimate to what extent the size of the sample is important, since the PSA technique in this study deals with dimensionality reduction, not with descriptive statistics. The dimensionality reduction simplifies the data and removes the “variation noise.” While dimensionality reduction can mitigate some issues related to small sample sizes, it does not completely eliminate the need for adequate sample sizes for robust analysis. Nonetheless, according to the obtained results, the behavior of the data are adequate and corresponds to the known characteristics of the deer species involved in this study. Therefore, the size of samples has little or practically no influence on results.
Certain cranial measurements were further examined by plotting their relative values against condylobasal length. The analysis was conducted using the Python programming language (Python 3.7.9) within the Jupyter Notebook environment. The Jupyter Notebook, along with craniometric data (CSV and EXCEL files) and Supplementary Figures S1–S4, are available in the Supplementary Materials.
3. Results
The cranial measurements of all specimens collectively exhibit a very high degree of correlation, thus supporting Geist’s statement [4] that cervids, particularly the subfamily Cervinae, possess a relatively conservative and uniform cranial shape. This finding is particularly intriguing as the database encompasses nearly the entire evolutionary diversity of cervids, including modern ‘primitive’ Muntiacus, continental giant forms such as Megaloceros, and both modern and extinct insular small-sized deer. It suggests that despite variations in body size, antler weight, and dentition specialization, deer generally share similar skull shapes and proportions throughout their evolutionary history.
The results indicate that body size is often the primary characteristic distinguishing species. However, there are two measurements that exhibit lower (moderate) correlation with others: the length of the interfrontal suture and the length of the premaxillary bones measured from P2 to the prosthion point. The reduced correlation of these measurements with other craniometric characteristics is attributable to different factors in each case.
The length of the interfrontal suture is associated with the degree of caudal extension of the nasal bones. This caudal extension of nasal bones represents an advanced morphological feature [2] that evolved independently in several evolutionary lineages among deer species of varying body sizes. Short nasal bones that do not reach the level of the anterior edges of the orbits represent a morphological condition found in species such as Cervus and Muntiacus [6]. Conversely, significant caudal extension of nasal bones evolved independently in several species with small body sizes, such as the fallow deer Dama dama, the extinct Axis-like Metacervocerus rhenanus, and the giant deer Megaloceros giganteus.
The relative length of premaxillary bones is an exosomatic feature representing a feeding adaptatin and is therefore morphologically more variable than other parts of the skull.
The observed moderate correlation between braincase breadth and braincase length, measured as the distance between the bregma point and the inion (Figure 2 and Figure 3), is intriguing as it indicates a certain diversity in braincase shape within the subfamily Cervinae. Vislobokova [2] posited that the evolution of the family Cervidae is characterized by the evolutionary shortening of the braincase. Unspecialized forms of deer typically exhibit elongated oval-shaped neurocrania and a relatively short facial portion of the skull, while the most advanced deer display a relatively short braincase and a long splanchnocranium. The analyzed data in the present study indicate that cervine species often considered the most primitive (such as Muntiacus reevesi, Muntiacus muntjak, and Hyelaphus porcinus) are actually characterized by relatively advanced short braincase proportions, where the braincase breadth behind the pedicles exceeds the bregma-inion distance. On the other hand, the two evolutionary most specialized species within Cervinae, Dama dama and Megaloceros giganteus, retain elongated braincases, with the bregma-inion measurement surpassing the braincase breadth behind the pedicles.
The Principal Component Analysis is particularly well-suited for this study, as the first two principal components, capturing 94% of the data variance, retain the maximum variance. Generally, the first Principal Component reflects skull length measurements, while the second Principal Component reflects breadth measurements. Although these components are related to body size, they do not correspond exactly, as some skulls exhibit relatively shortened or elongated proportions (Figure 4). From the obtained diagram, we can conclude that the first factorial plane adequately represents the data. The variable factor map confirms the varying degree of positive correlation between all cranial measurements (Figure 5). The positioning of species on the factorial plane is influenced by the contribution of their cranial breadth and length measurements. Unexpectedly, the resulting plot cloud exhibits complexity, yet it is perfectly explainable.
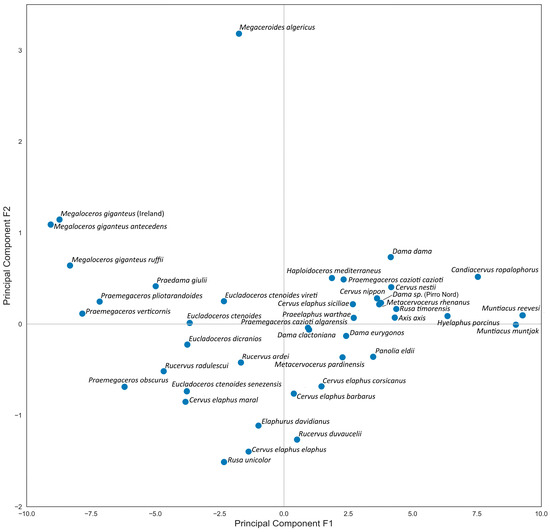
Figure 4.
Factorial plane 1: distribution of deer species following Principal Component Analysis of craniometric data.
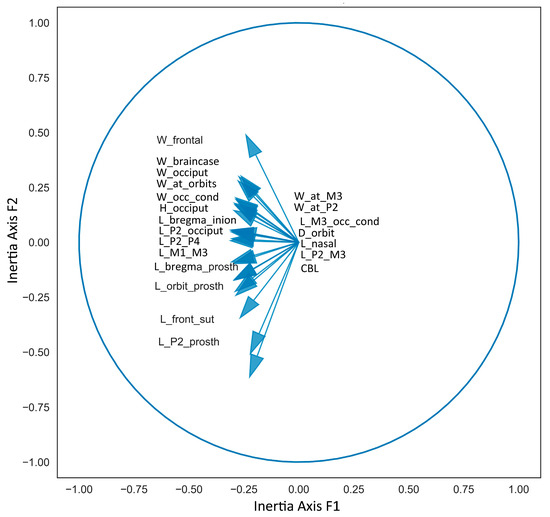
Figure 5.
Variable factor map of cranial measurements used in Principal Component Analysis.
The Factorial Plan depicts a distinctive zigzag-shaped distribution of species (with Megaceroides algericus as a single exception), revealing the variability in the craniometric parameters of deer within the subfamily and indicating the trajectory of evolutionary changes in skull morphology (Figure 4). The species situated in the upper left end of the PCA plot are characterized by generally robust skull proportions, notably broad frontals, and a relatively short snout, as seen in Megaloceros giganteus. Interestingly, Praedama giulii also demonstrates a trend toward this region of the dot cloud. The craniometric data for Praedama giulii can be considered relatively comprehensive, as they are based on a complete female skull from Untermassfeld and male cranial fragments of Praedama dupuisi, which apparently is a senior synonym of P. giulii.
In the lower left part of the diagram, large deer with relatively slender and elongated cranial proportions, particularly noticeable in the elongated facial part of the skull, are placed: Elaphurus davidianus, Cervus elaphus, and Rusa unicolor. On the right side of the plot, small subspecies of red deer (Cervus elaphus) are situated. These forms of red deer are characterized primarily by the shortening of the facial part of the skull, particularly the orbitofrontal portion. The so-called “Dama-like” deer represent a compact group that includes representatives of several extinct taxa, such as Metacervocerus, Praeelaphus, Haploidoceros, and insular Praemegaceros (Nesoleipoceros), as well as modern and fossil forms of the genus Dama, modern axis deer (Axis axis), sika deer (Cervus nippon), and Javan rusa (Rusa timorensis).
The extreme position of Muntiacus at the left extremity of the factorial plane mirrors its small size; however, the skull proportions of species within the genus Muntiacus are rather close to the average type of the subfamily Cervinae.
The outlier position of Megaceroides algericus is determined by the predominant importance of its breadth measurements, particularly the breadth of the frontlet and the breadth of the braincase, with very insignificant length parameters, especially the length of the facial part, such as P2—prosthion length and orbit—prosthion distance.
A closer examination of the ratios between certain measurements provides a better understanding of the evolutionary or ecomorphological significance of some features. Of particular interest is the ratio between the bregma-inion distance and the bregma-prosthion distance, which exhibits a step-like distribution when plotted against the condylobasal length (Figure 6). Small-sized and medium-small deer (below 100–120 kg of body mass) have this index varying between 32 and 38%, suggesting that the bregma point is somewhat shifted rostrally. In large-sized and giant deer, this ratio is below 32%, indicating a caudal shift of the bregma point. Several factors may influence the position of the bregma point, such as the relative length of the rostrum or the shortening or elongation of the orbitofrontal portion of the skull; however, this characteristic is clearly related to body size. Perhaps this is a better characteristic for demonstrating Vislobokova’s [2] observation on the shortening of the braincase and elongation of the splanchnocranium in more advanced (large-sized) deer. However, one possible cause of the caudal shifting of the bregma point in large-sized deer could be the increase in frontal bone size and robustness related to the increased weight of antlers and the increased robustness of pedicles. The transition from a more rostrally positioned bregma point to its more caudal position is not linear and has a rather broken, steplike character, possibly mirroring a certain biomechanical threshold. The position of the bregma point has no taxonomical significance and may vary within a genus or even between subspecies of the same species. This is evident in the subspecies of red deer: the insular subspecies Cervus elaphus siciliae has a more rostral position of the bregma point, unlike all other subspecies of Cervus elaphus. A similar difference can be observed in the Corso-Sardinian species Praemegaceros cazioti, where the less specialized nominotypical subspecies has a caudally shifted bregma point, presumably similar to its larger continental form, while in the more specialized P. cazioti algarensis, the bregma point is shifted rostrally.
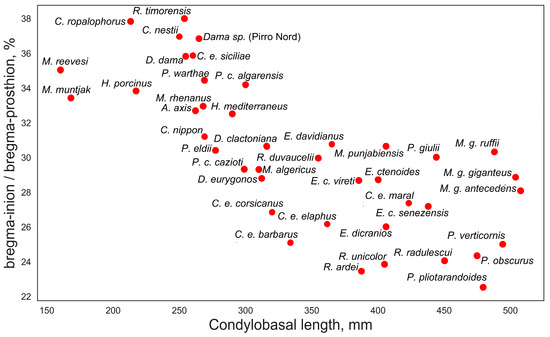
Figure 6.
Bregma position in various deer species: ratio of bregma-inion to bregma-prosthion measurements plotted against condylobasal length.
The relationship between the position of the bregma and the relative length of the facial part of the skull is not so straightforward. The relative length of the splanchnocranium is a feature related to the degree of evolutionary specialization of the skull in cervids: large body size is positively correlated with a relatively longer splanchnocranium in deer [2]. However, the analyzed data do not show such a clear relationship. Generally, the relative length of the face, measured from the anterior edge of the orbit to the prosthion in most studied cervids (including Muntiacus and Megaloceros), varies from 55% to 65%. Extreme shortening of the face is recorded only in Megaceroides algericus, with the facial portion attaining less than half of the condylobasal length, and in the highly specialized insular dwarf Candiacervus ropalophorus, which has a facial length attaining only 30% of the condylobasal length (see Supplementary Materials).
The relative breadth of the frontal bones, plotted against the condylobasal length, does not show a clear relationship with antler size (Figure 7). Megaloceros giganteus from Ireland is characterized by the broadest frontal bones among the large-sized deer with more or less heavy antlers. However, the relatively broadest frontal bones are found in specialized cervids, primarily in the North African endemic species Megaceroides algericus, which shows the broadest frontal bones in accordance with the generally short and broad proportions of its skull (Figure 8). Exceptionally broad frontal bones are also characteristic of some insular forms, such as Praemegaceros cazioti and Cervus elaphus siciliae.
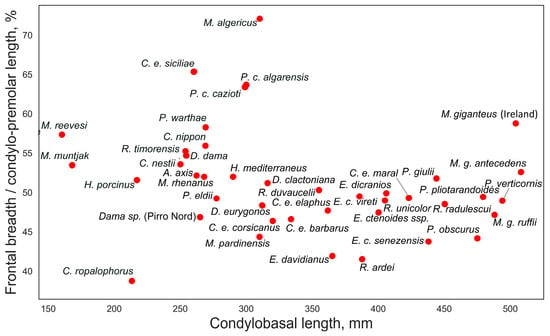
Figure 7.
Relative frontal breadth: ratio of frontal breadth at constriction to cranial length from upper second premolar to posterior side of occipital condyles plotted against condylobasal length.
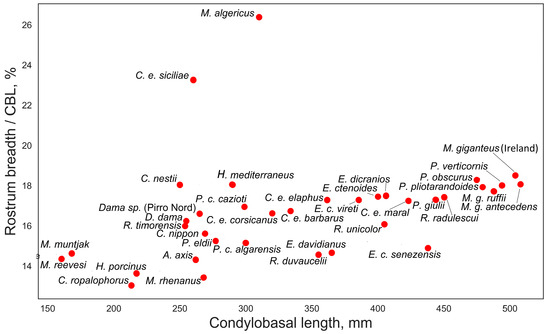
Figure 8.
The relative breadth of rostrum, calculated as the ratio of rostrum breadth to condylobasal length, plotted against condylobasal length.
The relative breadth of the snout, measured at the upper canines or the premaxillary/maxillary suture, is moderately positively correlated with condylobasal length (Figure 8), but this relationship between measurements is likely due to increased cranial robustness in larger deer. However, there are some interesting exceptions that may have ecomorphological significance. Eucladoceros ctenoides senezensis is characterized by a relatively narrow snout, approximately 14.5% of the condylobasal length. Megaceroides algericus, on the other hand, is characterized by the broadest snout among cervids, approximately 26.5% of CBL. It is also necessary to note the unusually broad snout in the insular red deer Cervus elaphus siciliae, approximately 23% of CBL.
4. Discussion
4.1. Craniometric Diversity within the Subfamily Cervinae
The distribution pattern of species on the PCA plot suggests several conclusions regarding the relationship between body size and ecomorphological diversity. Firstly, giant and large deer display significantly broader ecomorphological diversity in cranial shape compared to small-sized deer. Secondly, small-sized deer exhibit greater uniformity in their cranial shape, appearing densely clustered on the PCA plot. Thirdly, there is a gap in the continuum of cranial morphology that contributes to the V-shaped distribution of cervid species on the PCA plot (Figure 4).
The first two observations can be attributed to the unique digestive physiology of ruminants. Body size in ruminants imposes a corresponding rate of metabolism, which is lower in larger species and higher in smaller species [24].
The complex anatomy of the four-chambered stomach includes the rumen, which serves as the primary site for microbial fermentation of ingested feed in ruminants. The duration of fermentation depends on the quality of forage, particularly the relative content of cellulose fibers in the ingested plant material. Fermented food particles can progress through the digestive tract via the reticulum only if they are small enough. Lower-quality forage increases fermentation time in the rumen, and in species with high metabolism, such as small ruminants, animals may experience starvation despite their rumen being filled with low-quality, hard-to-ferment forage [24]. This limitation in forage quality is even more stringent in small male cervids, which require high-quality forage each year for antler growth.
As a result, smaller deer have a narrower range of digestible food resources, primarily limited to higher-quality forage. This eco-physiological constraint results in reduced diversity of cranial shape among small-sized cervids, which are adapted to similar forage, feeding height, and biomes. Given that cervid dentition and postcranial bones are even more uniform, distinguishing fragmentary remains of small cervids with similar body sizes becomes challenging. Differential and taxonomically meaningful characters become very subtle. As a result, the taxonomy and systematics of the so-called “small-sized deer” or “Dama-like deer” remain subjects of long-lasting debates and disagreements [19,23,25,26,27,28,29,30]. We will delve into this issue in detail in the next chapter.
In contrast, large and giant deer exhibit broader tolerance to forage quality due to their lower metabolic rates. Consequently, larger deer can occupy more diverse ecological niches, such as the modern large-sized tropical forest-dwelling Rusa unicolor, the extinct giant woodland dweller Praemegaceros obscurus, and the giant open landscape inhabitant Megaloceros giganteus.
The narrower availability of ecological niches in smaller cervids likely contributes to lower diversity in cranial shape. Conversely, the broader ecological tolerance of large and giant deer enables them to occupy a wider range of ecological niches, which underpins the greater diversity in cranial shape observed in large-sized deer.
The cause of the observed gap within the eco-morphological continuum of cervid cranial shape is not yet clear. It may be attributable to biomechanical limitations or ecological constraints.
The alignment of the skull craniometric characteristics of Muntiacus with the average type of the subfamily Cervinae is interesting because we do not find here a specific “primitive”, “archaic”, or paedomorphic craniometric condition that could be expected in very small tropical deer. This serves as one more argument in favor of the viewpoint that Muntiacus is a quite specialized genus [5] characterized, besides the extreme reduction of lateral digits, by typical Cervinae cranial proportions, including a relatively short braincase.
The relative length of the face in deer of the subfamily Cervinae varies from 55% to 65% and rather represents a species-specific feature that is not correlated or correlated very little with body size.
4.2. Craniometric Features of Giant Deer
One of the most interesting results is the cranial shape specialization observed in Megaloceros, which deviates from Vislobokova’s [2] rule of ‘larger body size—longer splanchnoranium’. According to the position of the giant deer on the factorial map, it seems that the facial portion of Megaloceros giganteus is relatively shorter than expected for a deer of such size (Figure 4). However, the relative length of the snout, measured between the orbit and prosthion with respect to condylobasal length, in the giant deer Megaloceros giganteus is actually the same as in Praemegaceros obscurus. It measures 56.8% in the only complete skull of Praemegaceros obscurus (IGF4024 from Pietrafitta); in Megaloceros giganteus, this index varies between 54.1% and 59.4% (34 individuals, with a mean value of 56.9%).
Partially, this could be related to the relatively long and unflexed braincase (Figure 9). Thus, the relative length of the braincase, expressed as a ratio bregma–inion length to braincase breadth, ranges from 96.1% to 140% in Praemegaceros (with 22 individuals belonging to P. obscurus, P. pliotarandoides, and P. verticornis; the mean value being 118.2%), while in Megaloceros giganteus, this ratio ranges from 85.4% to 99.7% (with 42 individuals, females excluded; the mean value being 90.9%) (Figure 10). Overlapping values are represented by a very young individual of Praemegaceros verticornis (Nr 1/229, Sucleia, Moldova) and an old individual of Megaloceros giganteus (SMNS18360, Murr, Rhine Valley, Germany).
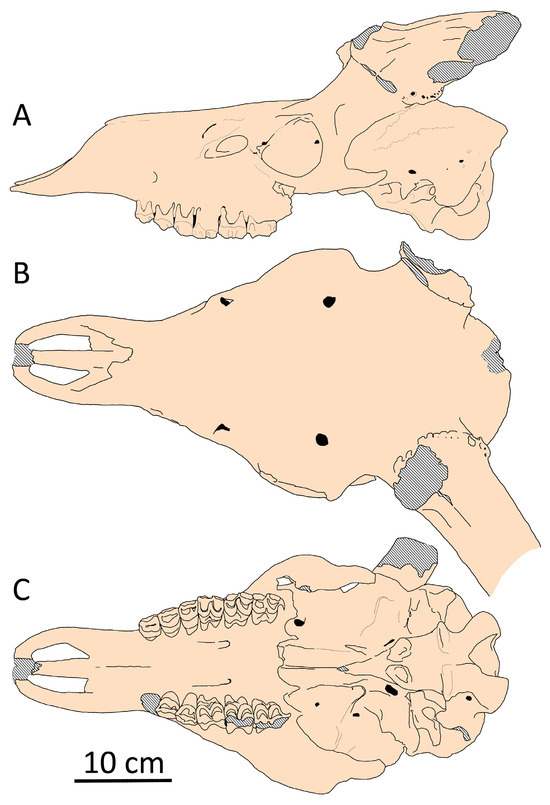
Figure 9.
Cranial shape of Megaloceros giganteus from Brühl (Koller), Rhine Valley, Germany (SMNS, collection number 166.16.17.11.80.37): (A) side view; (B) dorsal view; (C) palatal view.
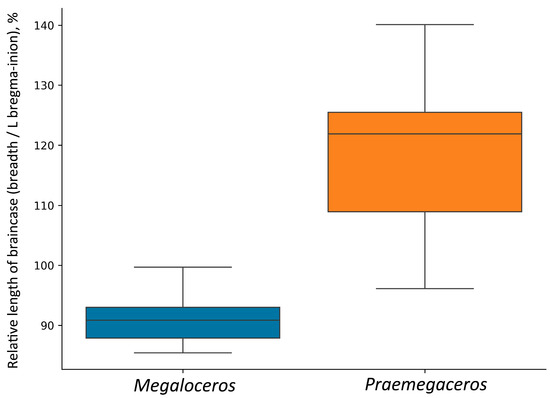
Figure 10.
Relative length of braincase in Megaloceros and Praemegaceros, calculated as the ratio of braincase breadth behind pedicles to bregma-inion distance.
Partially, the small relative length of the splanchnocranium in Megaloceros giganteus is related to its shortened orbitofrontal portion, a feature that distinguishes the giant deer from the long-snoutd Cervus elaphus and Elaphurus davidianus. Thus, the anterior edge of the orbit in Megaloceros giganteus is situated above M3 (Figure 9), while in red deer and Pere David’s deer the downward projection of the anterior edge of orbit falls far behind M3. However, the bregma point in giant deer is shifted caudally, as in all large-sized deer with a relatively long face (Figure 6). Possibly, this feature is related to the increased size of the heavy antler-bearing frontal bones, which are also relatively broad, especially in the giant deer from Ireland. The pedicles in giant deer maintain the common length and position observed in cervids; however, the frontal bones between pedicles are reinforced by extreme thickening, forming a strong bulge that creates a specific concave shape of the forehead before the pedicles [21].
The most significant evolutionary specialization of Megaloceros giganteus is its cranial pachyostosis, where the bones of the skull are significantly thickened compared to other cervid species. This adaptation most likely serves as a mineral storage mechanism necessary for the rapid growth of large antlers, particularly in environments with short vegetation periods [31]. The cranial pachyostosis results in several specific features of the giant deer’s skull morphology. For instance, advanced ossification of the vomer [32], previously misunderstood as a feature aligning plesiometacarpal Megaloceros with telemetacarpal representatives of the subfamily Capreolinae, is characterized by a completely ossified vomer dividing the choanae [33]. This feature is plesiomorphic for telemetacarpal deer and is not related to pachyostosis. Other cranial features associated with pachyostosis in giant deer include a reduced size of the foramen ovale, extremely reduced or completely closed ethmoidal openings, and the extreme thickening of mandibles, sometimes reaching an almost circular cross-section [31,32].
Despite these adaptations, cranial pachyostosis has had minimal influence on the general cranial shape and proportions of Megaloceros giganteus. Specific morphological specializations include the strong caudal extension of nasal bones beyond the level of the anterior edges of the orbits [32]. However, in many aspects, the cranial morphology of the giant deer remains primitive: the braincase remains unflexed, relatively long, and narrow compared to the genus Praemegaceros (Figure 10). These morphological features, according to Vislobokova [2], are primitive among cervids.
4.3. Specialization of Cranial Morphology in Megaceroides
Among cervid species, one of the most intriguing is Megaceroides algericus from the Late Pleistocene of Northwestern Africa, which occupies an outlier position on the factorial plane (Figure 4). This medium-sized endemic relative of Megaloceros giganteus exhibits unique characteristics compared to other representatives of the subfamily Cervinae (Figure 11). Similar to Eurasian Megaloceros giganteus, Megaceroides algericus displays significant cranial and mandibular pachyostosis [34,35]. However, the cranial pachyostosis in Megaceroides serves a different adaptive purpose. Unlike Megaloceros, Megaceroides lacks a frontal bulge, with frontals appearing notably flat and broad, and its neurocranium exhibits moderate flexion. The overall cranial pachyostosis is more pronounced in Megaceroides: eye socket rims are markedly thick, premaxillary bones are exceptionally broad and robust, and bone suture obliteration occurs early. In contrast, the zygomatic arches are thin and fragile, while cheek teeth are brachyodont and significantly reduced in size, particularly the third upper and lower molars [35]. Megaceroides’ outlier position is attributed to its distinctive cranial shape, characterized by a broad and flattened braincase and frontals and a relatively short snout—the shortest among continental deer (Figure 4). The facial portion between the orbits and prosthion is notably short, not exceeding half the length of the condylobasal measurement. The combination of a robust and broad skull with small cheek teeth and fragile, thin zygomatic arches is highly unusual, suggesting specific eco-morphological adaptations that differ significantly from those of most cervids. It is noteworthy that Megaceroides algericus is the only cervid species to have successfully entered and established itself on the African continent. The dispersal of Megaceroides algericus into Africa occurred during the Middle Pleistocene via the East-Mediterranean dispersal route, indicating a lengthy evolutionary process in Northwestern Africa, which allowed for extreme specializations distinguishing this deer from Eurasian Cervinae.
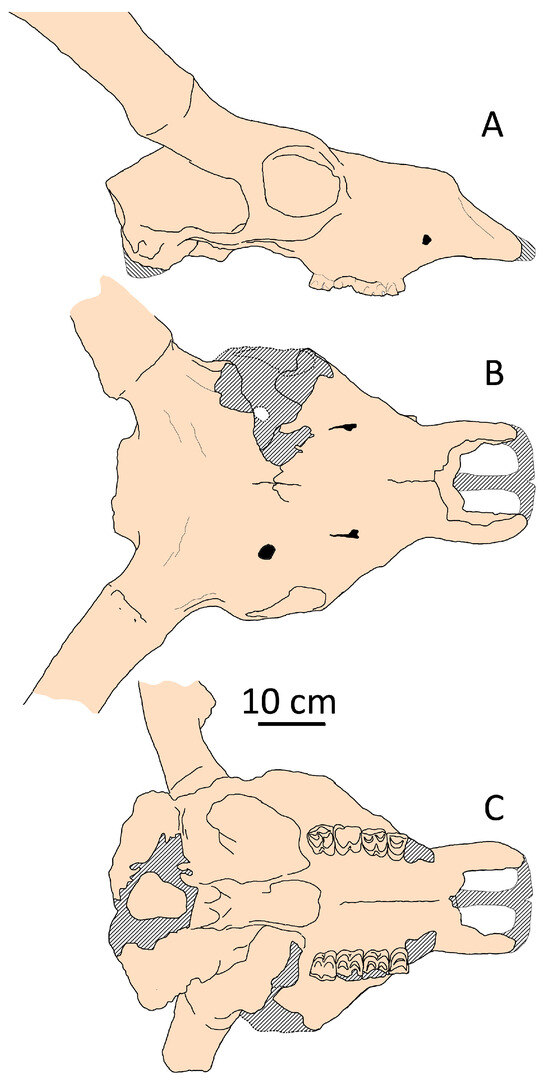
Figure 11.
Cranial shape of Megaceroides algericus from Guyotville (now Ain-Benian), Algeria (NMNH, “Collection Arambourg”’, no number): (A) side view; (B) dorsal view; (C) palatal view.
The primary obstacle to cervid dispersal into Africa is ecological competition with the diverse and rich community of African bovids, which have evolved high ecomorphological specializations, posing strong competition for less specialized and opportunistic Cervidae [4]. Consequently, successful cervid dispersal into Africa would require adaptation to a marginal ecological niche unoccupied by local African bovids. The combination of an extremely pachyostotic, broad, and flattened skull with small and partially reduced cheek teeth, along with thin zygomatic arches, may be interpreted as an adaptation to a periaquatic or semiaquatic ecological niche rich in soft, nutritious water plants but also fraught with the constant danger of attacks by dangerous water predators such as crocodiles [35]. The broad premaxillary bones indicate an adaptation to feeding on soft water plants. Thus, the extremely thick and hard cranial bones serve as a protective helmet, a preadaptation inherited from its Eurasian predecessor, Megaloceros giganteus, with initially different adaptation significance [35].
4.4. Insular Deer
The analyzed data comprise craniometric measurements of several species that are either considered insular or presumed to have been insular: modern Rusa timorensis from the islands of Java, Bali, and Timor; modern Cervus elaphus corsicanus from Corsica and Sardinia; the Late Pleistocene Cervus elaphus siciliae from Sicily; Praemegaceros cazioti from the Late Pleistocene of Corsica and Sardinia; and Candiacervus ropalophorus from the Late Pleistocene and Holocene of Crete. The results obtained did not reveal a specific “morphological syndrome” unique to all endemic insular deer.
The evolution of deer in conditions of insular isolation has long been recognized as a distinctive mode of evolution characterized by a suite of particular adaptations, forming a syndrome of responses to insular isolation. Among these adaptations, researchers often cite diminished body size, reduced and simplified antlers, shortened limb bones, abbreviated splanchnocranium, and generally paedomorphic cranial features [12,15,36,37]. Researchers have paid particularly close attention to Pleistocene insular deer of the Mediterranean. Azzaroli [12] proposed that all extinct Mediterranean insular deer belonged to only two genera: Megaloceros and Cervus. Radulescu and Samson [38] suggested that all extinct endemics from the Mediterranean islands should be grouped into a special genus, Nesoleipoceros. However, subsequent studies revealed that the taxonomic situation in Pleistocene endemic deer of Mediterranean islands is highly complex, with more than two phylogenetically distant lineages that independently colonized Mediterranean islands [36,37,39,40]. Do insular cervids indeed follow a specific, predefined evolutionary trajectory across all cervid lineages that colonized islands? This study seeks to provide an answer.
Cervus elaphus corsicanus. Some Mediterranean insular deer, as noted by Azzaroli [12], indeed belong to the genus Cervus. Two insular forms of red deer are included in the present study: modern Cervus elaphus corsicanus and extinct Cervus elaphus siciliae. Cervus elaphus corsicanus is one of the smallest modern forms of the red deer, characterized by simplified antlers with a diminished number of antler tines (missing bez tine and simplified crown) and a somewhat shorter facial portion of the splanchnocranium compared to the largest subspecies of red deer such as Cervus elaphus maral [6,14]. Specifically, C. elaphus corsicanus is characterized by a relatively shorter orbitofrontal skull portion. Thus, in C. elaphus maral, the anterior edge of the orbit projects downward behind the third upper molar (Figure 1), whereas in C. elaphus corsicanus, the anterior edge of the orbit is positioned above the posterior half of the second upper molar ([6]: Figure 50). According to Flerov [6], C. elaphus corsicanus is actually not an insular deer and should be synonymized with Atlas deer Cervus elaphus barbarus. This opinion was confirmed by archaeozoological studies [41] and genetic studies [42], which demonstrated that modern red deer from Corsica and Sardinia were introduced by humans. The mitochondrial haplotype of Corso-Sardinian red deer and North Africa reveals their relationship with the indigenous red deer population from the central part of the Italian Peninsula [42]. Therefore, C. elaphus corsicanus is a diminished form of the continental red deer. The shortened orbitofrontal portion of the skull in C. elaphus corsicanus is rather a paedomorphic feature. On the factorial plane, the Corso-Sardinian red deer is situated close to Atlas red deer, and both cervid forms are placed among continental deer (Figure 4). According to our results, C. elaphus corsicanus and C. elaphus barbarus show very close cranial characteristics: moderate breadth of occiput with respect to condylopremolar length, much narrower than in many insular deer; frontal bones are moderately narrow and slightly narrower than in C. elaphus elaphus and C. elaphus maral. However, the frontal bones of Corsican and Atlas deer are significantly narrower than in other fossil Mediterranean deer and significantly narrower than in specialized insular endemic deer, such as C. elaphus siciliae and Praemegaceros cazioti. So, the craniometric data reveal the close cranial resemblance between supposed “insular” C. elaphus corsicanus and continental C. elaphus barbarus, thus confirming the possible synonymy between those two subspecies, if we can still speak about subspecies taking into account their artificial translocation from the Italian Peninsula [41,42]. Geist [4] described the cranial shape of Atlas red deer as exhibiting paedomorphism, a characteristic also noted in C. elaphus corsicanus [6,14]. So, the craniometric data reveal that C. elaphus corsicanus maintains the cranial shape and proportions characteristic of continental deer and therefore does not show specific insular adaptations. The relative length of the facial part in C. elaphus corsicanus is very close to that of continental red deer C. elaphus elaphus. Concerning the simplified shape of antlers, they are known to be very sensitive to limited ecological resources [43]. Therefore, the diminished and simplified antlers of Corsican deer rather represent a phenetic response to environmental conditions than a genuine evolutionary adaptation to insular isolation. The body size of both C. elaphus barbarus and C. elaphus corsicanus generally corresponds to the size of continental Cervinae from the same latitudes (such as Cervus nippon, Panolia eldii, etc.), and therefore we cannot speak about a particular body size reduction.
Cervus elaphus siciliae. Craniometric features of C. elaphus siciliae reveal a distinct pattern suggestive of deep evolutionary transformation. This subspecies of red deer evolved in geographic isolation on Sicily Island during the Middle and Late Pleistocene. It is characterized by a diminished body size (approximately 80 kg) and reduced size of antlers, although they typically retain all elements such as the brow, bez, trez, and crown tines. The three-pointed crown of C. elaphus siciliae indicates its relation to crowned forms of red deer, akin to C. elaphus maral or Cervus elaphus aretinus. On the factorial map (Figure 4), this deer is positioned among insular species such as Praemegaceros cazioti and Rusa timorensis, as well as Mediterranean continental small-sized deer such as Dama dama and Haploidoceros mediterraneus.
As noted by Gliozzi et al. [16], the braincase of C. elaphus siciliae is relatively short and shares proportions with continental European red deer forms. The authors also observed a notably narrow and high occiput, along with a relatively short snout, in this insular deer. According to our data, the relative length of the face in C. elaphus siciliae, measured as the distance between the anterior edges of the orbit to the prosthion, closely resembles that of Cervus nippon, Rusa timorensis, and Dama dama. Remarkably, C. elaphus siciliae is the only subspecies of red deer with a rostrally shifted bregma point (Figure 6). One of its most striking morphological features is the relatively broad frontals, broader even than those of giant deer from Ireland (Figure 7). Some features of C. elaphus siciliae suggest an adaptation to grazing: the relative breadth of the rostrum is extreme for cervines, reaching approximately 23% of the condylobasal length (Figure 8). Additionally, premaxillary bones are square-shaped, while maxillary bones exhibit strongly developed rugosity for the attachment of musculus masseter [16]. The ecomorphological features of C. elaphus siciliae are quite similar to those seen in Praemegaceros (Nesoleipoceros) cazioti from the Late Pleistocene of Corsica and Sardinia.
Praemegaceros cazioti. Praemegaceros (Nesoleipoceros) cazioti evolved from the giant continental deer Praemegaceros (Nesoleipoceros) solilhacus, which is known from the Early Middle Pleistocene of Europe [36]. The evolutionary relationship between the continental giant P. solilhacus with large palmated antlers (approximately 400 kg in body mass [32]) and the insular Corso-Sardinian endemic P. cazioti is primarily inferred from the similarity of antler shape, and both species are classified within the subgenus Nesoleipoceros of the genus Praemegaceros. Praemegaceros cazioti is characterized by a significantly reduced body size (estimated body mass approximately 80–100 kg [36]) and a vestigial character of antler palmation. Thus, during the evolutionary transition from P. solilhacus to P. cazioti, the lineage of Nesoleipoceros experienced a 4–5-fold reduction in body size.
Unfortunately, we do not have at our disposal a complete skull of P. solilhacus, but the cranial material of the better-known Praemegaceros obscurus [44] can provide insight into the cranial transformations during the reduction in body size as an adaptation to insular isolation.
Both subspecies, P. cazioti cazioti and P. cazioti algarensis, are positioned among “Dama-like” deer on the factorial plan (Figure 4). The frontal bones in Praemegaceros cazioti are flat and relatively broad, comparable in breadth to those of C. elaphus siciliae (Figure 7). Unlike C. elaphus siciliae, P. cazioti is characterized by an exceptionally broad occiput (Figure 12), comparable to that of Megaceroides algericus in its relative breadth calculated to premolar-occipital length (excluding the variable part of the skull between P2 and prosthion). A similarly broad occiput is also observed in Rusa timorensis. The functional significance of such extreme occipital breadth is not immediately evident. Geist [4] explained the relatively broad occiput in Hyelaphus porcinus as having a functional role in accommodating strong neck muscles needed to stabilize the animal’s head during fast locomotion through tall and dense grass. While the occiput in hog deer is indeed relatively broader than in most cervines, it is not as extremely broad as in P. cazioti and R. timorensis. Perhaps the relatively broad occiput in P. cazioti is another morphological remnant from its giant ancestor. The broad occiput in large-sized deer with large heavy antlers (Megaloceros, Praemegaceros, Cervalces) is an adaptation that enhances head stability by increasing the strength of the neck muscles supporting the head [2,21]. However, the presence of this feature in R. timorensis suggests that this morphological peculiarity may have a more complex evolutionary origin.
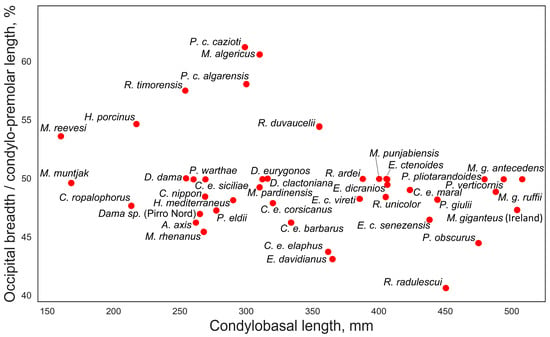
Figure 12.
Relative breadth of occiput calculated as the ratio of occipital breadth to cranial length measured from P2 to the posterior side of occipital condyles, plotted against condylobasal length.
The nasal bones in P. cazioti are short, with their posterior edge not reaching the line connecting the anterior edges of the eye sockets [36]. The area of musculus masseter insertion is clearly defined and extends forward until the level of M1. The orbitofrontal region is shorter compared to continental species of the genera Eucladoceros and Praemegaceros [36,45,46]; in lateral view, the anterior edge of the orbit is situated at the level of M2. The premaxillary bones are broad, robust, and square-shaped [45,46,47], resembling those of Cervus elaphus siciliae, suggesting similar adaptations to grazing (Figure 13). One of the most interesting morphological features of P. cazioti cazioti is the more caudal position of the bregma point (Figure 6), indicating the retention of some characteristics typical of giant continental deer.
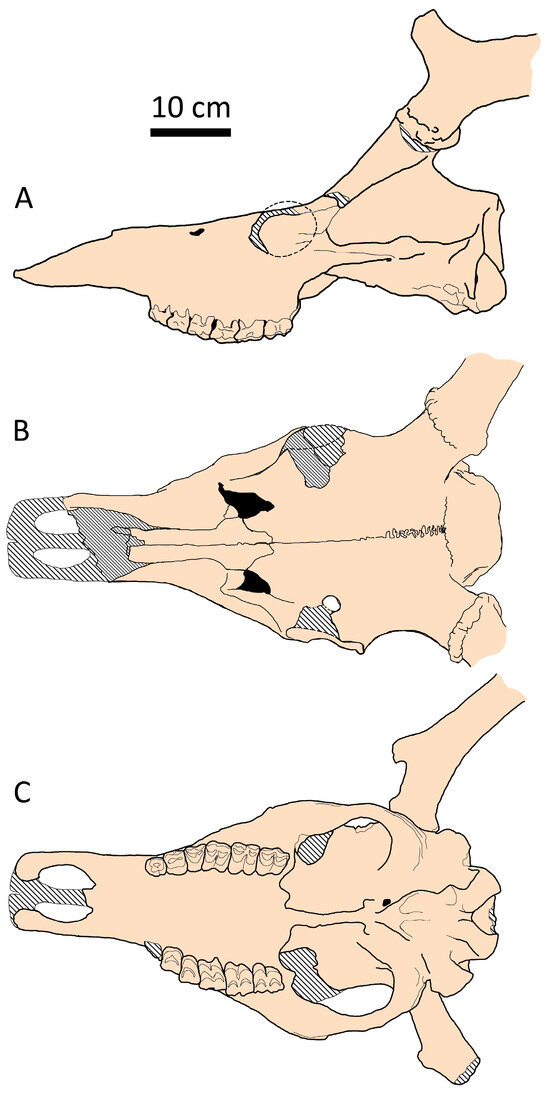
Figure 13.
Cranial shape of Praemegaceros cazioti cazioti from Coscia (Corsica): (A) side view (MNHN, no number); (B) dorsal view (Cos-6930; adapted from [36]; (C) palatal view (MNHN, no number).
However, the grazing adaptations in P. cazioti cazioti are still moderate and do not significantly alter its craniodental morphology [36]. Thus, the upper cheek tooth-row exhibits a relative length typical of Cervinae, remaining below 30% of the condylobasal skull length. The skull proportions of P. cazioti cazioti from Coscia (Corsica) remain largely similar to those of P. obscurus from Pietrafitta, except for a slightly shorter face, which nonetheless maintains a similar relative length compared to mainland deer of similar size. Both insular P. cazioti cazioti and mainland small-sized species share similarities in the position of the upper tooth row, the relative length of the P2-prosthion distance, and the relative length of the upper dentition [36,45,46,47].
Praemegaceros cazioti algarensis from Dragonara (Sardinia) exhibits more evolved grazing adaptations: its square-shaped praemaxillare are notably broad; the area of musculus masseter insertion is extensive, bordered with prominent ribs, and extends forward until the level of P4 [46,47]. The upper tooth row is relatively long, reaching 33.7% of the condylobasal length, and the cheek teeth have remarkably higher crowns, classified as mesodontous [36,46]. The orbitofrontal portion is very short, with the anterior edge of the orbit positioned at the level of the posterior edge of M2 (when projected onto the upper tooth row plane), and the distance between P2 and the prosthion is notably short (24.5% of the condylobasal length). Unlike P. cazioti cazioti, the bregma point in P. cazioti algarensis is shifted rostrally, as observed in all small-sized deer (Figure 6). The difference in the position of the bregma point between P. cazioti cazioti and P. cazioti algarensis is likely related to the shortening of the facial part: the relative length of the snout, measured from the anterior edges of the orbits to the prosthion with respect to the condylobasal length, is 56% in the former subspecies and approximately 50% in the latter.
The morphological and craniometric similarities between C. elaphus siciliae and P. cazioti should be considered more as a convergence than parallelism. Both deer exhibit exceptionally broad frontal bones, a trait likely inherited from their large-sized or giant ancestors as a consequence of significant body size reduction. This disproportionally broad feature of the frontal bones may share a similar nature to that observed in Megaceroides algericus. Thus, in small-sized deer, disproportionally broad frontal bones may serve as an indicator of considerable evolutionary size reduction.
Candiacervus ropalophorus. The genus Candiacervus encompasses insular deer native to Crete, which have undergone extreme reduction in body size and significant cranial shape transformation due to evolution in isolated island conditions [12,15,37,39]. The body size of Candiacervus species falls between that of Hyelaphus and Muntiacus. Possibly, the genus Candiacervus evolved from the continental giant deer Praemegaceros obscurus or Praemegaceros verticornis. The evolutionary relationship between Candiacervus and Praemegaceros is apparent from the antler shape of Candiacervus listeri, considered the less specialized representative of the genus Candiacervus, which retains key characteristics of antler morphology from its continental precursor. The antlers of Candiacervus listeri preserve all structural elements seen in Praemegaceros [32,38,44], including the dorsal (outer) tine, middle tine, and the sharp bend in the area of the posterior tine, albeit reduced in C. listeri [37]. Additionally, C. listeri maintains the shape and position of compressed and divergent pedicles [37] observed in advanced continental Praemegaceros [32].
The antlers of Candiacervus ropalophorus are the most specialized within the genus, retaining only the middle tine and featuring a very long beam with a mace-shaped distal end [37]. This deer exhibits advanced cranial adaptations to insular conditions, positioning it somewhat marginally on the factorial map, though not as an extreme outlier (Figure 4).
Candiacervus ropalophorus is characterized by a significantly reduced relative length of the facial part of the skull [15]. The measurement from the orbit to the prosthion point attains 30% of the condylobasal length, representing an outlier value among cervines, where the relative length of the face typically ranges from 50 to 65%. The position of the bregma point is strongly shifted rostrally, with the ratio of bregma-inion to bregma-prosthion amounting to 38%, which is also an extreme value among Cervinae, shared only with Rusa timorensis (Figure 6). The relative length of the cheek teeth attains 30% of the condylobasal length, a common value among representatives of the subfamily Cervinae.
Frontal bones in C. ropalophorus are flat, with moderately diverged and backward-pointing pedicles, resembling those in advanced continental Praemegaceros. However, frontal bones are relatively narrow, showing the lowest value among Cervines. Possibly, the peculiar narrowness of frontals in C. ropalophorus is related to the shortened and diminished facial portion of the cranium. Similarly, C. ropalophorus has the narrowest rostrum among Cervinae, although its premaxillary bones are square-shaped, as seen in other endemic Mediterranean insular deer with grazing adaptations.
Among the morphological peculiarities of the splanchnocranium in C. ropalophorus, one can notice its low profile. This morphological peculiarity is shared with Praemegaceros cazioti (Figure 13) and is supposed to be related to the diminished volume of the nasal cavity. One plausible explanation for this peculiar feature in insular deer is their diminished olfactory capacities in the absence of carnivores in the insular ecosystem [36]. The braincase of C. ropalophorus is relatively short, as in continental Praemegaceros [15], while its occiput maintains a normal relative breadth for cervinae (Figure 12).
Thus, our study could not reveal a universal type of insular dwarfed deer, as each insular species inherits specific morphology from its continental ancestor and is characterized by its own degree of evolutionary adaptation to insular conditions. The character and completeness of the local insular fauna (the so-called “balanced insular faunas”, i.e., faunas containing predators that are very rare in endemic insular faunas) are another important factor that shapes the evolution of insular deer [13,48]. However, we can recognize some common morphological and craniometric features that mark the evolutionary diminishment of body size, such as disproportionately broad frontal bones and occiput in the early stages of evolutionary insular dwarfing, as well as the progressive shortening of the splanchnocranium and rostral shift of the bregma point.
4.5. “Dama-like” Deer
The so-called Plio-Pleistocene “Dama-like” deer (a term introduced by Di Stefano and Petronio [26]) or “small-sized deer” (a term introduced by Azzaroli [19]) represents an arbitrary group of cervids that includes about half a dozen forms with body masses ranging from 60 to 90 kg. This group of Villafranchian species presents a taxonomical issue that continues to confuse researchers. The main problem is that the postcranial and dental morphology of deer is quite uniform, making it challenging to distinguish isolated fossil fragments of deer with similar body sizes. This poses a practical inconvenience for biostratigraphic and faunistic studies, and a clear, simple taxonomical solution seems desirable. Thus, Azzaroli [12] introduced the genus Pseudodama, with the genotype Dama nestii nestii Azzaroli, 1947, grouping all Villafranchian “Dama-like” deer based on general antler morphology. These small-sized deer are characterized by three- and four-tined stages of antler evolution, which are quite universal and may be found in all lineages of the subfamily Cervinae [4]. This taxonomical solution has sparked extensive discussions, as many authors doubt that the genus Pseudodama represents a genuine monophyletic group [23,25,26,27,28,29,30,40].
The situation with Pseudodama is reminiscent of the historical development of views on the genus Megaceros/Megaloceros, which, during the late 19th century and the first half of the 20th century, was used as a taxonomical container for all giant deer and their supposed relatives [20,21,32,38]. However, simple and convenient taxonomical solutions are not always the best, as they can obscure the real evolutionary diversity of a systematic group and hide important evolutionary processes, as was the case with the genus Megaloceros [2,20,21,32,34,35,36,38,47].
Di Stefano and Petronio [26] approached Villafranchian deer from Europe to modern Asian genera Axis and Rusa. Their conclusions are based solely on general antler shape, which represents a stage of antler evolution [4], rather than a genuine phylogenetic relationship. Cranial morphology, however, is a much more valuable source of information for determining systematic positions and phylogenetic relationships among evolutionary lineages. It is widely used in zoological systematics, including the systematics of modern Cervidae [2,6,14,18,23,27,33].
The analysis of diagnostic cranial morphological characters of “Dama-like” deer—such as the shape and flexion of the braincase, the position of orbits relative to the upper tooth row, the length and position of nasal bones, the length and orientation of frontal pedicles, the presence or absence of upper canines, the shape of the ethmoidal opening, and other characteristics—revealed that the genotype of Pseudodama, Dama nestii nestii, shows a great affinity with modern European red deer Cervus elaphus [27]. This similarity is enhanced by antler features such as the pearled surface and the transverse orientation of the distal fork of their four-tined antlers relative to the sagittal plane [28].
The taxonomical importance of cranial diagnostic characters is well illustrated by hierarchical clustering, which includes the “Dama-like” deer and modern “typical” representatives of genera such as Cervus elaphus, Dama dama, Rusa unicolor, and Axis axis [30]. The results of hierarchical cluster analysis show that Dama nestii nestii is closely grouped with Cervus elaphus, while Dama eurygonos from the Late Villafranchian of Italy is clustered with modern fallow deer. Therefore, Pseudodama is a junior synonym of Cervus, and its type species should be regarded as Cervus nestii [27,28].
Other species previously included in the genus Pseudodama have no closely related forms in modern faunas and are now classified in the extinct genera Metacervocerus Dietrich, 1938 (M. pardinensis and M. rhenanus) and Praeelaphus Portins, 1920 (P. lyra) [28]. Metacervocerus had a vast distribution area, ranging from Eastern Asia and the Indian subcontinent to Western Europe, and included species with primitive cranial morphology, lacking upper canines, and possessing uniform three-pointed antlers with a smooth surface [29,30]. Superficially, Metacervocerus resembles the genus Axis; however, its antlers have a different bauplan, with the anterior tine of the distal fork being inserted on the anterior side of the antler beam while the posterior tine of the distal fork continues along the antler axis. The anterior tine of the distal fork of Metacervocerus is an evolutionary homologue of the trez tine in Cervus and Dama. It is likely that Metacervocerus is related to the origin of the genus Dama, which is endemic to the Mediterranean area of Eurasia [19,30].
Praeelaphus lyra is an endemic species from the Late Pliocene to Early Pleistocene of the Italian Peninsula, with origins related to Praeelaphus perrieri from the Pliocene of Western Europe [28]. Despite differences in evolutionary history and cranial morphology, the “Dama-like” deer from the Mediterranean area show similar biometric characteristics of the skull and similar body size.
The body size, similar to that of modern fallow deer, appears to hold pivotal importance in the subfamily Cervinae and likely within the entire Cervid family. This body size is common among continental species inhabiting warm latitudes of the Holarctic and the Oriental zoogeographic realm [23]. In the examples of insular deer discussed in this study, we have observed that body size reduction—especially in the early stages of evolution under insular isolation—can lead to deer reaching a size similar to that of fallow deer. However, insular isolation is not a prerequisite for evolutionary body size reduction to this extent. Examples from the continental paleontological record also exist. One such example is Haploidoceros mediterraneus, a small to medium-sized deer that evolved in the northwestern Mediterranean region during the Middle and Late Pleistocene [49]. The genus Haploidoceros underwent at least three million years of evolution in the warm and relatively dry conditions of the Pliocene in Western Europe and in the Mediterranean conditions of the Pleistocene glacial refugium of the Iberian Peninsula. Based on cranial variables, its body mass is estimated to be around 70–80 kg [40], considerably smaller than its presumed ancestor, Rucervus (Arvernoceros) ardei, with a body mass of approximately 180 kg [50]. Therefore, the evolution of the Haploidoceros lineage involved roughly a two-fold reduction in body mass. This reduction in body size appears to be an adaptation to the ecological niche of “Dama-like” deer within the Early Pleistocene Mediterranean ecosystems.
H. mediterraneus is characterized by unusually specialized antlers, consisting solely of a very long sickle-shaped beam and a single large basal tine [40]. The evolutionary reasons for antler simplification in Haploidoceros are not yet understood. The craniodental morphology of H. mediterraneus is also peculiar and does not provide a straightforward answer regarding the species’ dietary specialization. The relatively large cheek teeth (32–33% of condylobasal length) and short diastema in H. mediterraneus from Cova del Rinoceront (Spain) resemble the grazing adaptations of P. cazioti. However, the shape of narrow and pointed premaxillary bones (Figure 14) differs from the broad and rectangular-shaped premaxillaries of P. cazioti. The premaxillary bones of H. mediterraneus are even narrower and more pointed than those of fossil and modern fallow deer (Figure 14), approaching those of specialized browsers. Thus, the masticatory apparatus of H. mediterraneus represents several superimposed morphological adaptations resulting in an unusual combination of features: relatively large cheek teeth and narrow, pointed premaxillary bones, akin to browsing herbivores. The protruding telescopic (or tube-shaped) orbits are a distinguishing morphological feature of H. mediterraneus. The development of tubular orbits is most likely related to the long hair on males’ heads and necks, which could serve as protection against rivals’ simplified antlers during intraspecific combat [40]. Among other advanced cranial morphological specializations are short and diverged pedicles and a flexed braincase (Figure 14). However, despite numerous highly specialized morphological features, the skull of H. mediterraneus maintains generalized proportions characteristic of most less specialized small-sized Cervinae and is grouped with small-sized deer such as Axis and Dama on the factorial plane (Figure 4).
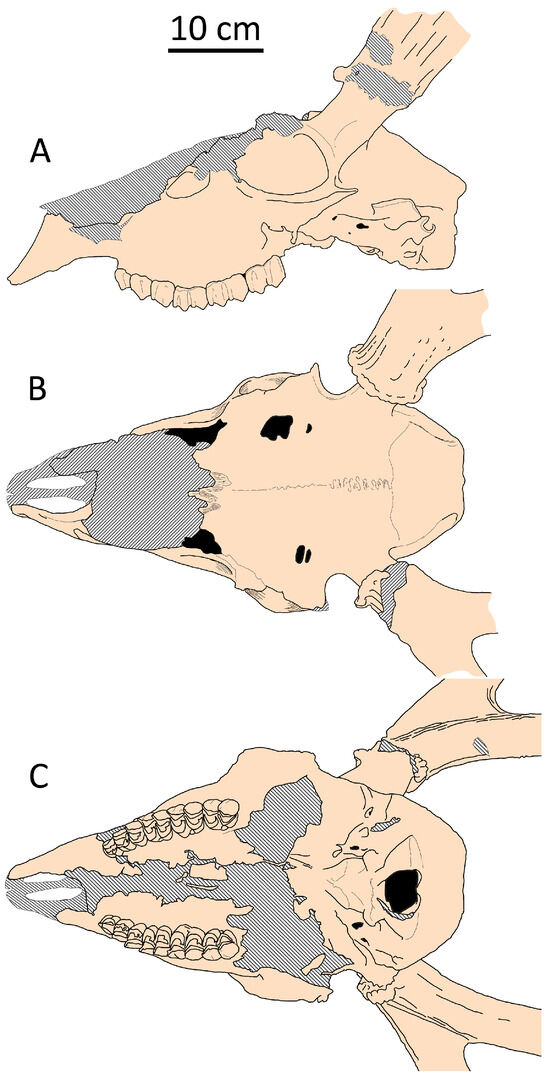
Figure 14.
Cranial shape of Haploidoceros mediterraneus from Cova del Rinoceront, Spain (CR-2413, FGUB): (A) side view; (B) dorsal view; (C) palatal view.
The term “Dama-like deer” carries deeper significance, as it refers to an eco-morphological type of small-sized deer that evolved independently in several phylogenetic lineages due to common eco-physiological and biomechanical constraints shared by all cervids. However, despite the similarity in craniometric characteristics, the systematic position of these small-sized deer can be deduced from morphological details of their skull, dentition, and antlers, which hold taxonomic and systematic significance [27].
The “Dama-like deer” represents a fundamental eco-physiological type among cervids, encapsulating key ecological and physiological traits relevant to their roles within ecosystems. This concept describes a set of characteristics essential for optimal metabolism, reproduction rates, and antipredator strategies tailored to specific ecological conditions. Associated with this eco-physiological type are distinct craniometric features influenced by shared ecological niches and food resource characteristics.
Given that the ecological and evolutionary strategy of the Cervinae subfamily often revolves around nutrient-rich foraging grounds, this fundamental eco-physiological type holds universal significance, occupying a central position within cervid diversity. However, external factors such as predation pressure, changes in food quality, and interactions with other herbivores can lead to the evolution of various forms within the cervid lineage, including giant forms, small Muntiacus-like forms, or other eco-physiological types, depending on the new ecological niches they inhabit. Yet, when these external pressures diminish, cervid lineages tend to revert to their fundamental eco-physiological type, as, for instance, in the case of endemic Mediterranean small-sized cervids discussed in the present study.
Thus, the “Dama-like” deer represents the fundamental eco-physiological type prevalent in warm latitudes, including the Mediterranean biome. The presence of several species of similar body size in the paleontological record has sparked prolonged debates and taxonomical uncertainties. Azzaroli [19] proposed a radical taxonomical solution by grouping all deer of small-medium body size from the Pliocene and Early Pleistocene of Europe into the genus Pseudodama, offering an unclear and vague diagnosis for this taxon. However, these cervids exhibit varying degrees of cranial shape specialization and antler evolution, displaying different stages of morphological and ethological development [27]. For instance, the modern fallow deer (Dama dama) showcases the most advanced cranial morphology among cervine species [4]. Similarly, the antler shapes of small-sized deer display a wide diversity, ranging from the highly complex palmated antlers of fallow deer to the simpler three-tined antlers in Axis axis [4]. Based on our taxonomic and systematic analyses, it is evident that the genus Pseudodama Azzaroli, 1992, is an artificial polyphyletic taxon encompassing cervid forms from various lineages, including Cervus, Dama, Metacervocerus, and Praeelaphus [23,27,28,29,30].
Thus, while “Dama-like deer” share similar eco-physiological and feeding adaptations, they each possess distinct evolutionary histories, biological characteristics, and behavioral traits. Therefore, the fundamental eco-physiological type should not be conflated with parallelism or convergence but should be viewed as the central, pivotal amalgamation of ecological and physiological characteristics within the subfamily Cervinae.
5. Conclusions
Concerning the hypotheses on the evolutionary trends in cranial morphology and proportions previously proposed in scientific literature, the following conclusions can be drawn:
(I) The results obtained support earlier findings regarding the conservative nature of cranial shape in cervines [2,4], highlighting a relatively low degree of craniodental specialization compared to bovids or even representatives of the subfamily Capreolinae, which exhibit a deeper level of cranial morphological diversification [4,51]. This fundamental conservatism in cervid cranial morphology reflects their generally opportunistic ecological strategy [4]. This observation is especially true for medium- and small-sized deer.
Body size remains the primary eco-morphological specialization in Cervinae, facilitating ecological partitioning among sympatric deer species [52]. Differences in body size are crucial for paleontologists in identifying sympatric fossil deer species from fossil faunas, as this approach remains reliable and methodologically straightforward [53]. Modern unmodified faunas rarely contain sympatric deer of the same body size, underscoring the importance of body size for ecological partitioning among deer species [4].
Craniometric data offer valuable information for taxonomic diagnoses at the subspecies, species, and genus levels [2,6,7,14,18,23,27,30,32]. However, due to numerous instances of parallelisms and convergences, their implication to higher taxonomic levels and to phylogenetic relationships between genera and groups is very limited. Thus, the present study was unable to resolve cervid systematics at the tribal level.
The Principal Component Analysis of craniometric data revealed that giant and large-sized deer display significantly broader ecomorphological diversity in cranial shape compared to small-sized deer. Secondly, small-sized deer exhibit greater uniformity in their cranial shape, appearing densely clustered on the factorial map. Thus, we conclude that body size imposes ecological constraints, limiting the available niches due to eco-physiological factors that restrict the range of food resources. Large-sized and giant deer exhibit broader ecological tolerance to food resources and can occupy a wider variety of ecological niches, resulting in a broader craniometric diversity. Conversely, smaller deer face more limited ecological niches, leading to more uniform craniometric characteristics, aligning with the perception of cervids having conservative and uniform cranial morphology.
The concept of the fundamental eco-physiological type elucidates the phenomenon of “Dama-like” deer, frequently evolved species across various cervid lineages with a body mass around 70–90 kg. These deer species may have different evolutionary histories and diverse biological and ethological adaptations. The “Dama-like” fundamental eco-physiological type is characterized by body mass that ensures optimal metabolism, reproduction rates, and antipredator strategies within specific ecological conditions. Additionally, this eco-physiological type is associated with specific craniometric characteristics dictated by similar ecological niches and food resource traits. In the Mediterranean biome, “Dama-like” deer represent the fundamental eco-physiological type, evolving equally in continental climate refugia and on isolated islands.
(II) The craniometric data and analysis of morphological cranial features of fossil and modern insular deer, or deer presumed to be adapted to insular isolation conditions, reveal a more complex picture that does not support a simplistic understanding of the “insular rule”. The results confirm that the assumed insular form of the red deer, Cervus elaphus corsicanus, is actually a paedomorphic form of continental red deer, as previously suggested [4,6]. Cervus elaphus siciliae and Praemegaceros cazioti exhibit convergent adaptations to Mediterranean insular environments with limited resources and are characterized by similar features (e.g., relatively broad frontal bones) that indicate a strong body size reduction during adaptation to insular conditions. The general cranial biometric characteristics align Cervus elaphus siciliae and Praemegaceros cazioti with other “Dama-like” cervids of the Mediterranean area. Candiacervus ropalophorus is the only cervid that fully corresponds to the “insular rule” and has undergone deep evolutionary modifications of cranial shape and morphology.
(III) The obtained results did not support the hypothesis of a gradual evolutionary shortening of the braincase from assumed primitive’ deer with small body size and simple antlers to large and giant specialized deer within the subfamily Cervinae. The relative length of the braincase appears to be influenced by multiple factors, including evolutionary history and superimposed ecomorphological adaptations.
Supplementary Materials
The following supporting information can be downloaded at: https://github.com/Praemuntiacus/Craniometric_Analysis_of_the_Subfamily_Cervinae (accessed on 7 March 2024); Figure S1: Missing data in the craniometric dataset; Figure S2: Missing data in the dataset of selected species; Figure S3: Individual and Cumulative Explained Variance; Figure S4: Hierarchical clustering of deer species based on scaled data; Table S1: Cervidae cranial measurements.
Funding
This study was conducted as part of the State Program Project No. 010701 at Moldova State University, Republic of Moldova.
Data Availability Statement
The raw data and Jupyter Notebook containing the Python code for data imputation, processing, analysis, and visualization are publicly accessible on GitHub: https://github.com/Praemuntiacus/Craniometric_Analysis_of_the_Subfamily_Cervinae (accessed on 7 March 2024).
Acknowledgments
I would like to express my sincere gratitude to the colleagues who provided access to the paleontological and osteological collections: the late Danilo Torre, Lorenzo Rook, Paul Peter Mazza (MGUF), Dumitru Murariu (NHMB), Andrew Currant (NHML), Paolo Agnelli (NHMF), Carmello Petronio, Maria Rita Palombo (MPS), the late Theresa Wiszniowska, Krzysztof Stefaniak (IZW), Pascal Tassy, Christine Argot, Claire Sagne (MNHN), Reinhard Ziegler (SMNS), Abel Prieur (PMUL), Montserrat Sanz, and Joan Daura (FGUB). I also wish to thank the three anonymous reviewers for their time and efforts, which significantly contributed to improving the quality of this paper.
Conflicts of Interest
The author declares no conflicts of interest.
References
- Lydekker, R. Deer of All Lands: A History of the Family Cervidae Living and Extinct; Rowland Ward Ltd.: London, UK, 1898; 329p. [Google Scholar]
- Vislobokova, I.A. The fossil deer of Eurasia. Transact. Paleont. Inst. 1990, 240, 1–208. (In Russian) [Google Scholar]
- Köhler, M. Skeleton and Habitat of recent and fossil Ruminants. Münch. Geowiss. Abh. A 1993, 25, 1–88. [Google Scholar]
- Geist, V. Deer of the World: Their Evolution, Behaviour, and Ecology; Stackpole Books: Mechanicsburg, PA, USA, 1998; pp. 1–421. [Google Scholar]
- Gentry, A.W.; Rössner, G.E.; Heizmann, E.P.J. Suborder Ruminantia. In The Miocene Land Mammals of Europe; Rössner, G.E., Heissig, K., Eds.; Verlag Dr. Friedrich Pfeil: München, Germany, 1999; pp. 225–258. [Google Scholar]
- Flerov, K.K. Musk deer and deer. In The Fauna of USSR. Mammals 1(2); Novikov, G.A., Ed.; Academy of Sciences of USSR: Moscow, USSR, 1952; pp. 1–256. (In Russian) [Google Scholar]
- Meijaard, E.; Groves, C.P. Morphometrical relationships between South-east Asian deer (Cervidae, tribe Cervini): Evolutionary and biogeographic implications. J. Zool. 2004, 263, 179–196. [Google Scholar] [CrossRef]
- Lowe, V.P.W.; Gardiner, A.S. A re-examination of the subspecies of Red deer (Cervus elaphus) with particular reference to the stocks in Britain. J. Zool. 1974, 174, 185–201. [Google Scholar] [CrossRef]
- Markov, G.G.; Danilkin, A.A. Geographic craniometrics variability and intraspecific taxonomy of the red deer (Cervus elaphus L.) in Palearctic. In Advances in Deer Biology, Proceedings of the 4th International Deer Biology Congress, Kaposvár, Hungary, 30 June–4 July 1998; Zomborszky, Z., Ed.; Pannon University of Agriculture Faculty of Animal Science: Kaposvár, Hungary, 1999; pp. 119–120. [Google Scholar]
- Polziehn, R.O.; Strobeck, C. Phylogeny of Wapiti, Red Deer, Sika Deer, and Other North American Cervids as Determined from Mitochondrial DNA. Mol. Phylogenet. Evol. 1998, 10, 249–258. [Google Scholar] [CrossRef]
- Ludt, C.; Schroeder, W.; Rottmann, O.; Kuehn, R. Mitochondrial DNA phylogeography of red deer (Cervus elaphus). Mol. Phylogenet. Evol. 2004, 31, 1064–1083. [Google Scholar] [CrossRef]
- Azzaroli, A. Il nanismo nei cervi insulari. Palaeontogr. Ital. 1961, 56, 1–32. [Google Scholar]
- Sondaar, P.Y. Insularity and its effects on mammal evolution. In Major Patterns in Vertebrate Evolution; Hect, M.N., Goody, P.L., Hect, B.M., Eds.; Plenum Press: New York, NY, USA, 1977; pp. 671–707. [Google Scholar]
- Heptner, V.G.; Zalkin, V.I. Deer of the USSR (Systematics and Zoogeography). Trans. Study Fauna Flora USSR Mosc. Soc. Nat. 1947, 10, 1–176. (In Russian) [Google Scholar]
- Schilling, A.M.; Roessner, G.E. New skull material of Pleistocene dwarf deer from Crete (Greece). Comptes Rendus Palevol 2021, 20, 141–164. [Google Scholar] [CrossRef]
- Gliozzi, E.; Malatesta, A.; Scalone, E. Revision of Cervus elaphus siciliae Pohlig, 1893, Late Pleistocene endemic seer of the Siculo-Maltese District. Geol. Rom. 1993, 29, 307–353. [Google Scholar]
- Breda, M.; Lister, A.; Kahlke, R.D. New results on cervids from the Early Pleistocene site of Untermassfeld. In The Pleistocene of Untermassfeld near Meiningen (Thüringen, Germany), Part 4. Monographien des Römisch-Germanisches Zentralmuseum; Kahlke, R.D., Ed.; Verlag des Römisch-Germanischen Zentralmuseums: Mainz, Germany, 2020; Volume 40, pp. 1197–1249. [Google Scholar]
- Sokolov, I.I. Hoofed Animals (Orders Perissodactyla and Artiodactyla). In Fauna of the USSR: Mammals, Volume 1, No. 3; Pavlovskij, E.N., Ed.; Academy of Sciences of USSR: Moscow, USSR, 1959; pp. 1–640. (In Russian) [Google Scholar]
- Azzaroli, A. The cervid genus Pseudodama n.g. in the Villafranchian of Tuscany. Palaeontogr. Ital. 1992, 79, 1–41. [Google Scholar]
- Azzaroli, A. Critical Remarks on some Giant Deer (genus Megaceros Owen) from the Pleistocene of Europe. Palaeontogr. Ital. 1979, 71, 7–16. [Google Scholar]
- Vislobokova, I.A. Giant Deer: Origin, Evolution, Role in the Biosphere. Paleontol. J. 2012, 46, 643–775. [Google Scholar] [CrossRef]
- Janis, C. Correlation of cranial and dental variables with body size in ungulates and macropodoids. In Body Size in Mammalian Paleobiology: Estimation and Biological Implications; Damuth, J., MacFadden, B.J., Eds.; Cambridge University Press: Cambridge, UK, 1990; pp. 255–299. [Google Scholar]
- Croitor, R. Plio-Pleistocene Deer of Western Palearctic: Taxonomy, Systematics, Phylogeny; Academy of Sciences of Moldova: Chişinău, Moldova, 2018; 140p. [Google Scholar]
- Janis, C. The evolutionary strategy of the Equidae and the origins of rumen and cecal digestion. Evolution 1976, 30, 757–774. [Google Scholar] [CrossRef]
- Vos, J.D.; Mol, D.; Reumer, J.W.F. Early Pleistocene Cervidae (Mammalia, Artiodactyla) from the Oosterschelde (the Netherlands), with a revision of the cervid genus Eucladoceros Falconer, 1868. Deinsea 1995, 2, 95–121. [Google Scholar]
- Di Stefano, G.; Petronio, C. Systematics and evolution of the Eurasian Plio-Pleistocene tribe Cervini (Artiodactyla, Mammalia). Geol. Rom. 2002, 36, 311–334. [Google Scholar]
- Croitor, R. Early Pleistocene small-sized deer of Europe. Hell. J. Geosci. 2006, 41, 89–117. [Google Scholar]
- Croitor, R. Deer from Late Miocene to Pleistocene of Western Palearctic: Matching fossil record and molecular phylogeny data. Zitteliana B 2014, 32, 115–153. [Google Scholar]
- Croitor, R.; Bonifay, M.-F. Etude preliminaire des cerfs du gisement Pleistocene inferieur de Ceyssaguet (Haut-Loire). Paleo 2001, 13, 129–144. [Google Scholar] [CrossRef]
- Croitor, R.; Robinson, C. A revision of “Cervus” punjabiensis Brown, 1926 (Cervidae, Mammalia) from the Upper Siwaliks of Chandigarh, India. Quat. Int. 2020, 550, 147–158. [Google Scholar] [CrossRef]
- Lister, A.M. The evolution of the giant deer, Megaloceros giganteus (Blumenbach). Zool. J. Linn. Soc. 1994, 112, 65–100. [Google Scholar] [CrossRef]
- Croitor, R. Taxonomy and systematics of large-sized deer of the genus Praemegaceros Portis, 1920 (Cervidae, Mammalia). Cour. Forsch.-Inst. Senckenberg 2006, 256, 91–116. [Google Scholar]
- Lönnberg, E. Which is the Taxonomic Position of the Irish Giant Deer and Allied Races? Almqvist Zool. 1906, 3, 1–8. [Google Scholar]
- Hadjiouis, J. Megaceroides algericus (Lydekker, 1890), du gisement des Phacochères (Alger, Algérie). Etude critique de la position systematique de Megaceroides. Quaternaire 1990, 3, 247–258. [Google Scholar] [CrossRef]
- Croitor, R. Systematical position and paleoecology of the endemic deer Megaceroides algericus Lydekker, 1890 (Cervidae, Mammalia) from the late Pleistocene-early Holocene of North Africa. Geobios 2016, 49, 265–283. [Google Scholar] [CrossRef]
- Croitor, R.; Bonifay, M.F.; Bonifay, E. Origin and evolution of the late Pleistocene island deer Praemegaceros (Nesoleipoceros) cazioti (Depéret) from Corsica and Sardinia. Bull. Mus. Anthropol. Préhist. Monaco 2006, 46, 35–68. [Google Scholar]
- Van der Geer, A.A.E. Uniformity in variety: Antler morphology and evolution in a predator-free environment. Palaeontol. Electron. 2018, 21.1.9A, 1–31. [Google Scholar] [CrossRef]
- Radulescu, C.; Samson, P. Sur un nouveau cerf megacerin du pleistocene moyen de la depression de Brasov (Roumanie). Geol. Rom. 1967, 6, 317–344. [Google Scholar]
- Kuss, S.E. Die pleistozänen Hirsche der ostmediterranen Inseln Kreta, Kasos, Karpathos und Rhodos (Griechenland). Berichte Naturforsch. Ges. Freibg. Breisgau 1975, 65, 25–79. [Google Scholar]
- Abbazzi, L.; Bonfiglio, L.; Marra, A.; Masini, F. A revision of medium and small sized deer from the Middle and Late Pleistocene of Calabria and Sicily. Boll. Soc. Paleontol. Ital. 2001, 40, 115–126. [Google Scholar]
- Vigne, J.-D. Les mammifères post-glaciaires de Corse. Étude Archéozoologique; Gallia Prehistoire: Paris, France, 1988; Volume 26, pp. 1–337. [Google Scholar]
- Doan, K.; Zachos, F.E.; Wilkens, B.; Vigne, J.D.; Piotrowska, N.; Stanković, A.; Jędrzejewska, B.; Stefaniak, K.; Niedziałkowska, M. Phylogeography of the Tyrrhenian red deer (Cervus elaphus corsicanus) resolved using ancient DNA of radiocarbon-dated subfossils. Sci. Rep. 2017, 7, 2331. [Google Scholar]
- Bubenik, A.B. Epigenetical, morphological, physiological, and behavioral aspects of evolution of horns, pronghorns, and antlers. In Horns, Pronghorns, and Antlers: Evolution, Morphology, Physiology, and Social Significance; Bubenik, G.A., Bubenik, A.B., Eds.; Springer: New York, NY, USA, 1990; pp. 3–113. [Google Scholar]
- Azzaroli, A.; Mazza, P. Large early Pleistocene deer from Pietrafitta lignite mine, Central Italy. Palaeontogr. Ital. 1993, 80, 1–24. [Google Scholar]
- Caloi, L.; Palombo, M.-R. Functional aspects and ecological implications in Pleistocene endemic cervids of Sardinia, Sicily and Crete. Geobios 1995, 28, 247–258. [Google Scholar] [CrossRef]
- Caloi, L.; Palombo, M.-R. Il megacerino di Dragonaria: Interpretazione funzionale di alcuni caratteri cranici. Geol. Rom. 1995, 31, 29–40. [Google Scholar]
- Caloi, L.; Malatesta, A. Il Cervo Pleistocenico di Sardegna. Mem. Ist. Ital. Paleontol. Uman. 1974, 2, 162–247. [Google Scholar]
- Palombo, M.R. Insular mammalian fauna dynamics and paleogeography: A lesson from the Western Mediterranean islands. Integr. Zool. 2018, 13, 2–20. [Google Scholar] [CrossRef]
- Croitor, R.; Sanz, M.; Daura, J. The endemic deer Haploidoceros mediterraneus (Bonifay)(Cervidae, Mammalia) from the Late Pleistocene of Cova del Rinoceront (Iberian Peninsula): Origin, ecomorphology, and paleobiology. Hist. Biol. 2020, 32, 409–427. [Google Scholar] [CrossRef]
- Croitor, R. A description of two new species of the genus Rucervus (Cervidae, Mammalia) from the Early Pleistocene of Southeast Europe, with comments on Hominin and South Asian ruminants dispersals. Quaternary 2018, 1, 17. [Google Scholar] [CrossRef]
- Gustafson, E.P. An Early Pliocene North American deer: Bretzia pseudalces, its osteology, biology, and place in cervid history. Bull. Mus. Nat. Hist. Univ. Or. 2015, 25, 1–75. [Google Scholar]
- Kaiser, T.M.; Croitor, R. Ecological interpretations of early Pleistocene deer (Mammalia, Cervidae) from Ceyssaguet (Haute-Loire, France). Geodiversitas 2004, 26, 661–674. [Google Scholar]
- Heintz, E. Les cervidés villafranchiens de France et d’Espagne. Mém. Mus. Natl. Hist. Nat. Sér. C Sci. Terre 1970, 22, 303. [Google Scholar]
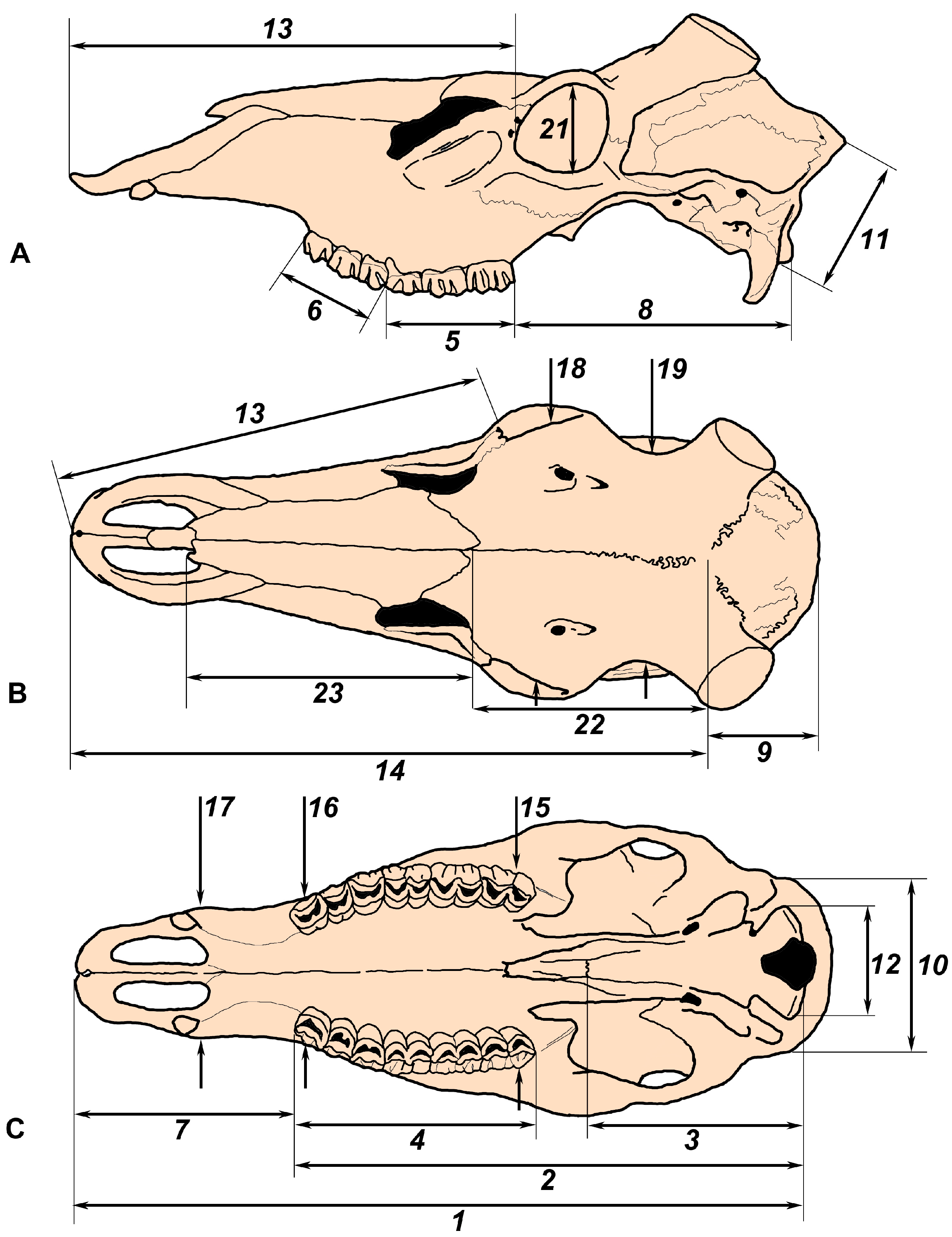
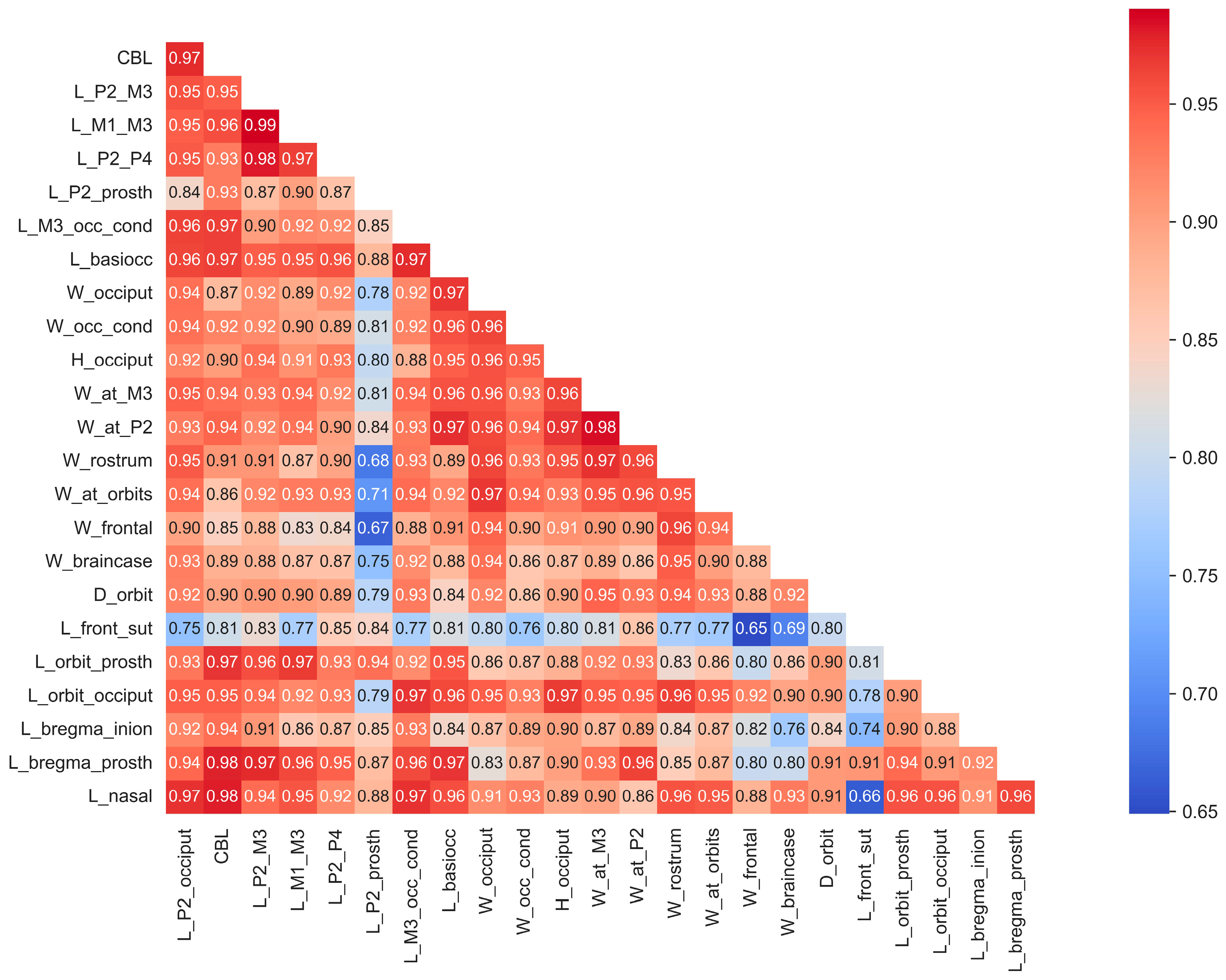
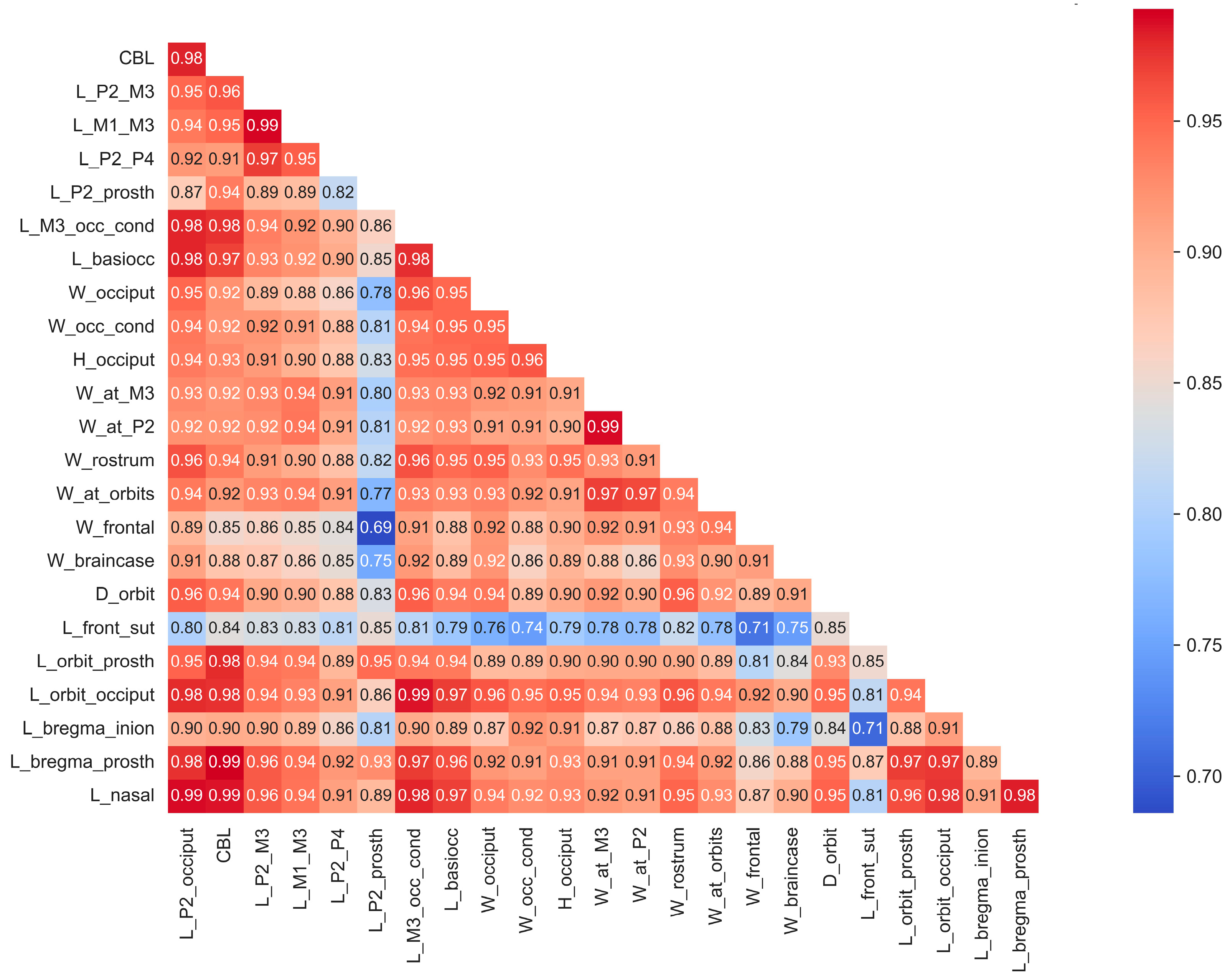
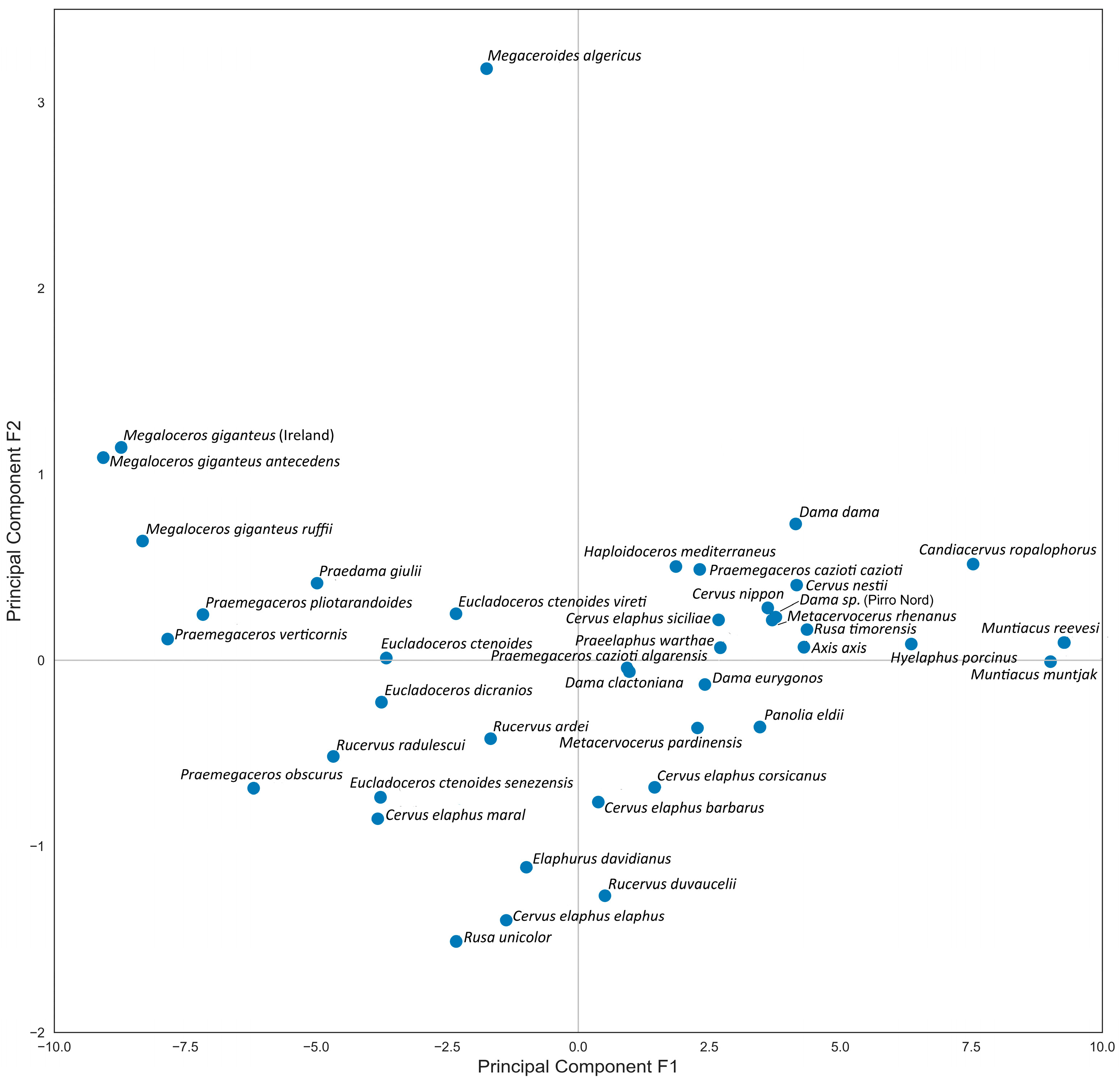
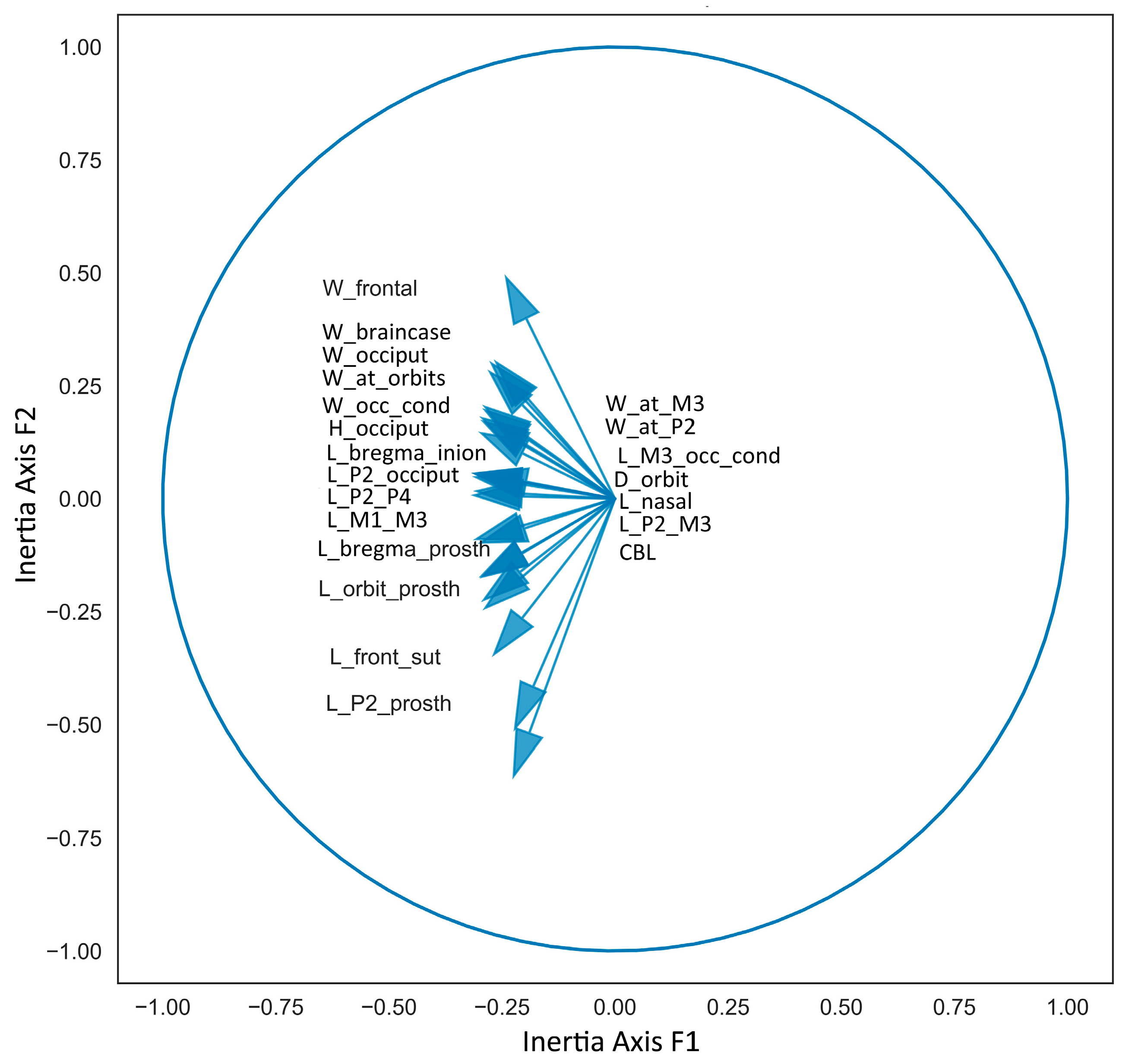
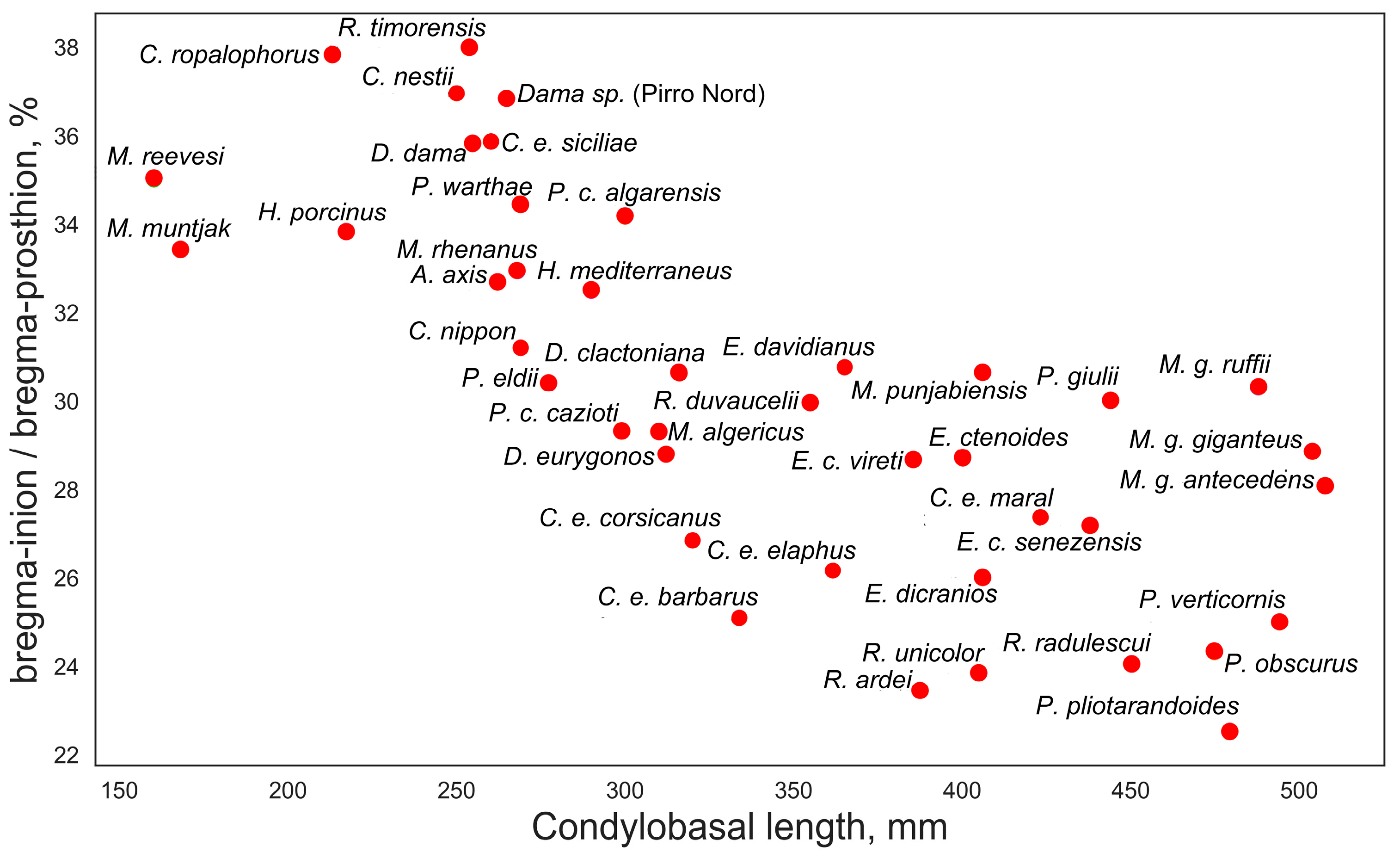
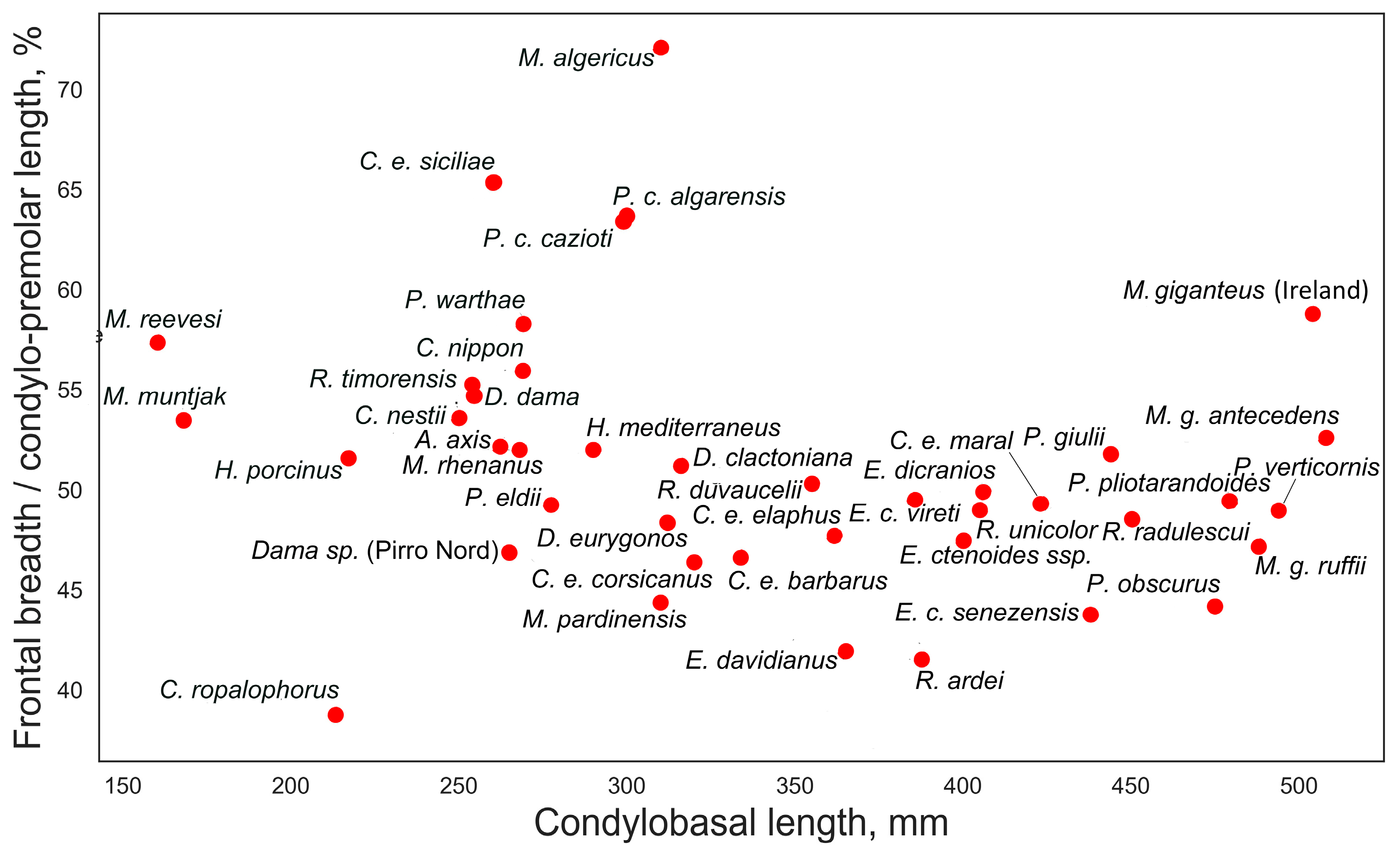
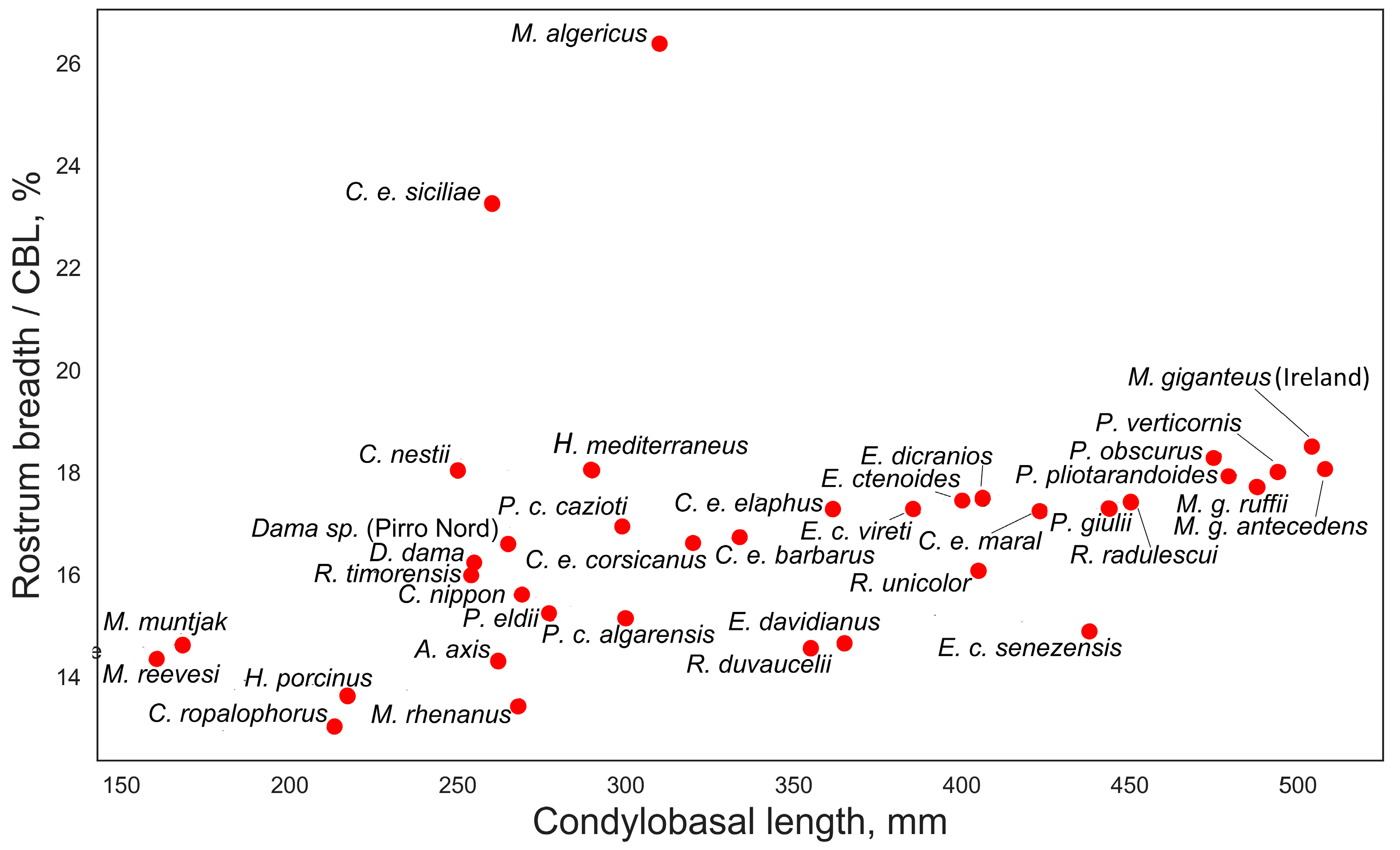
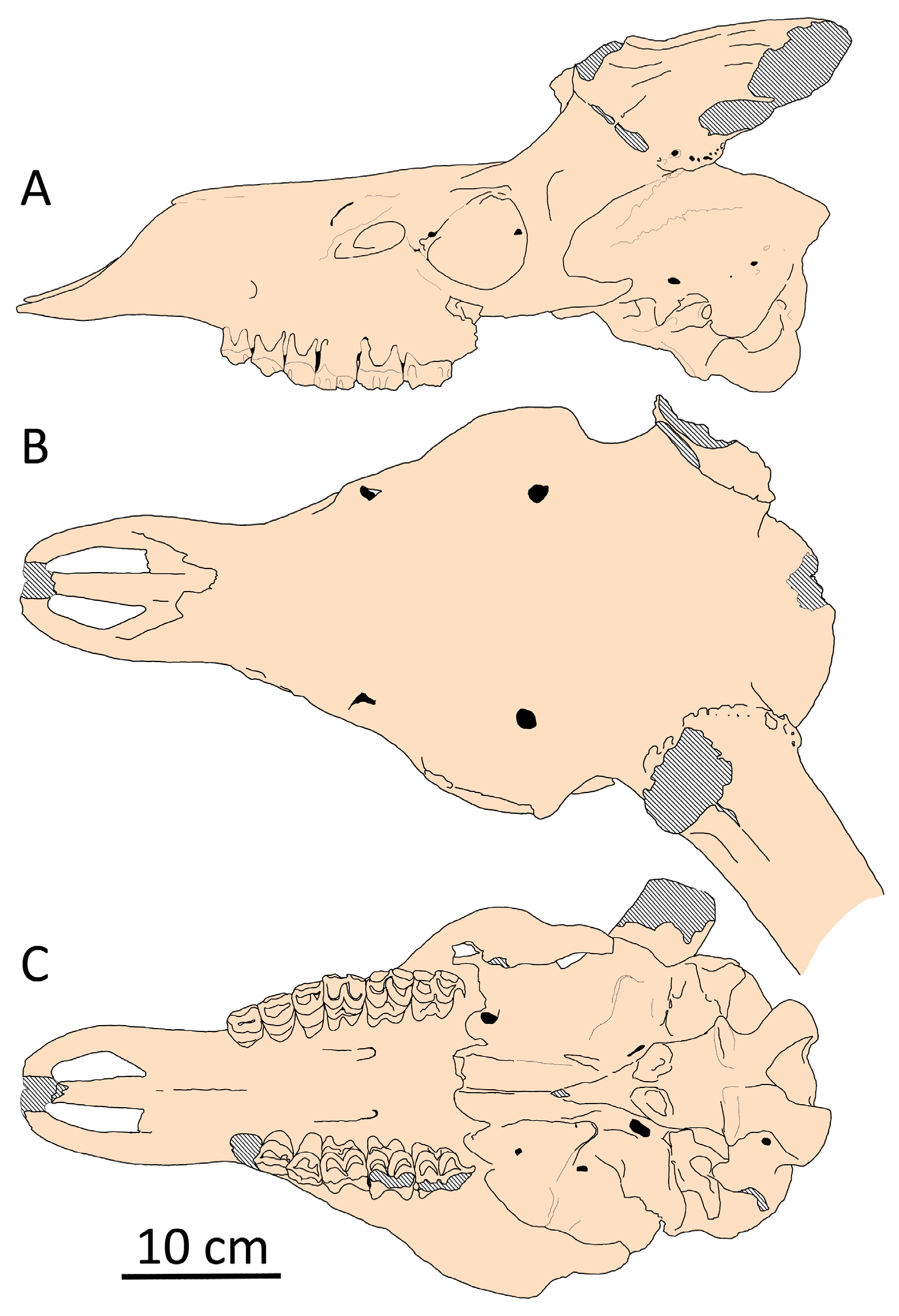
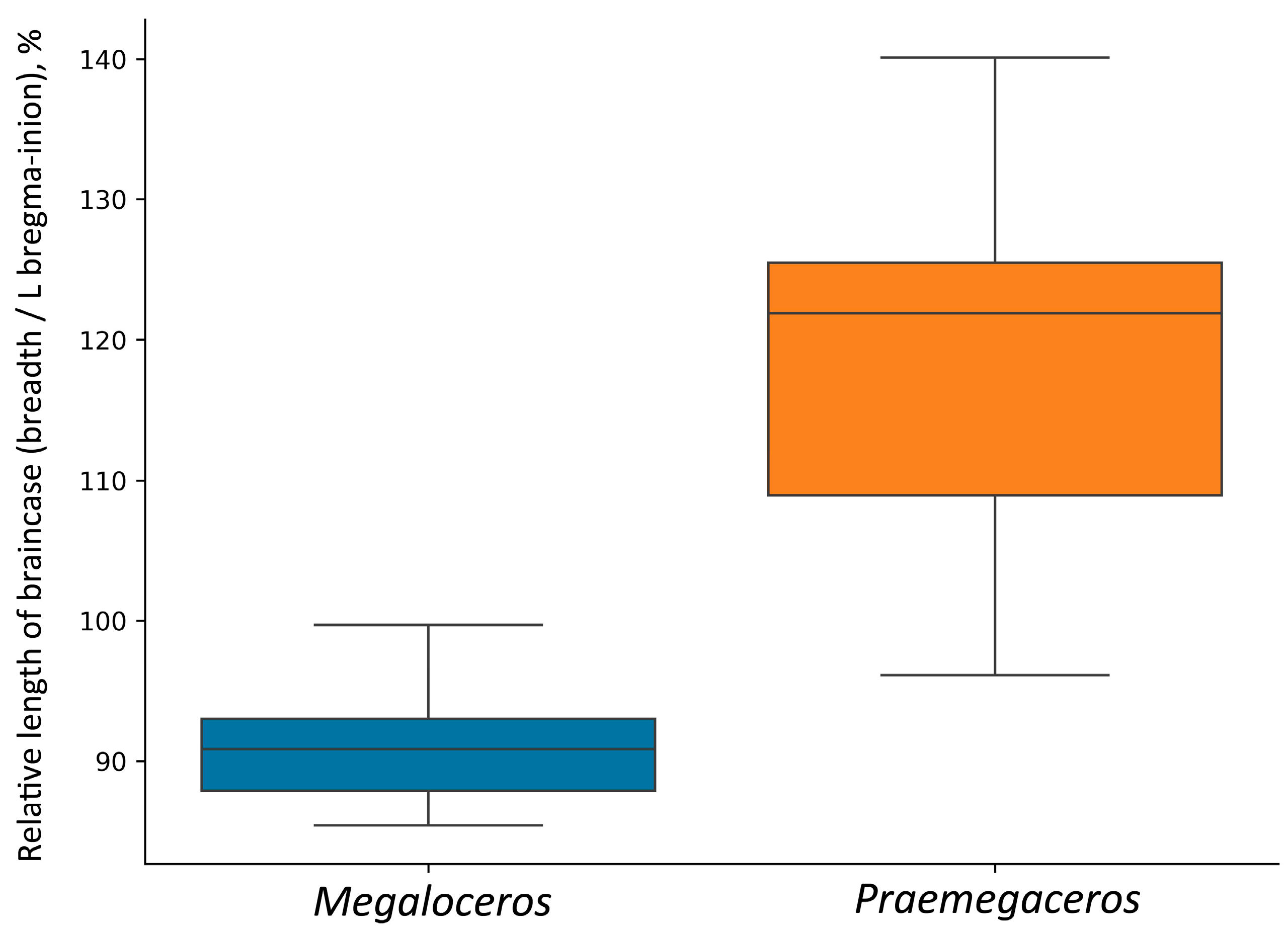
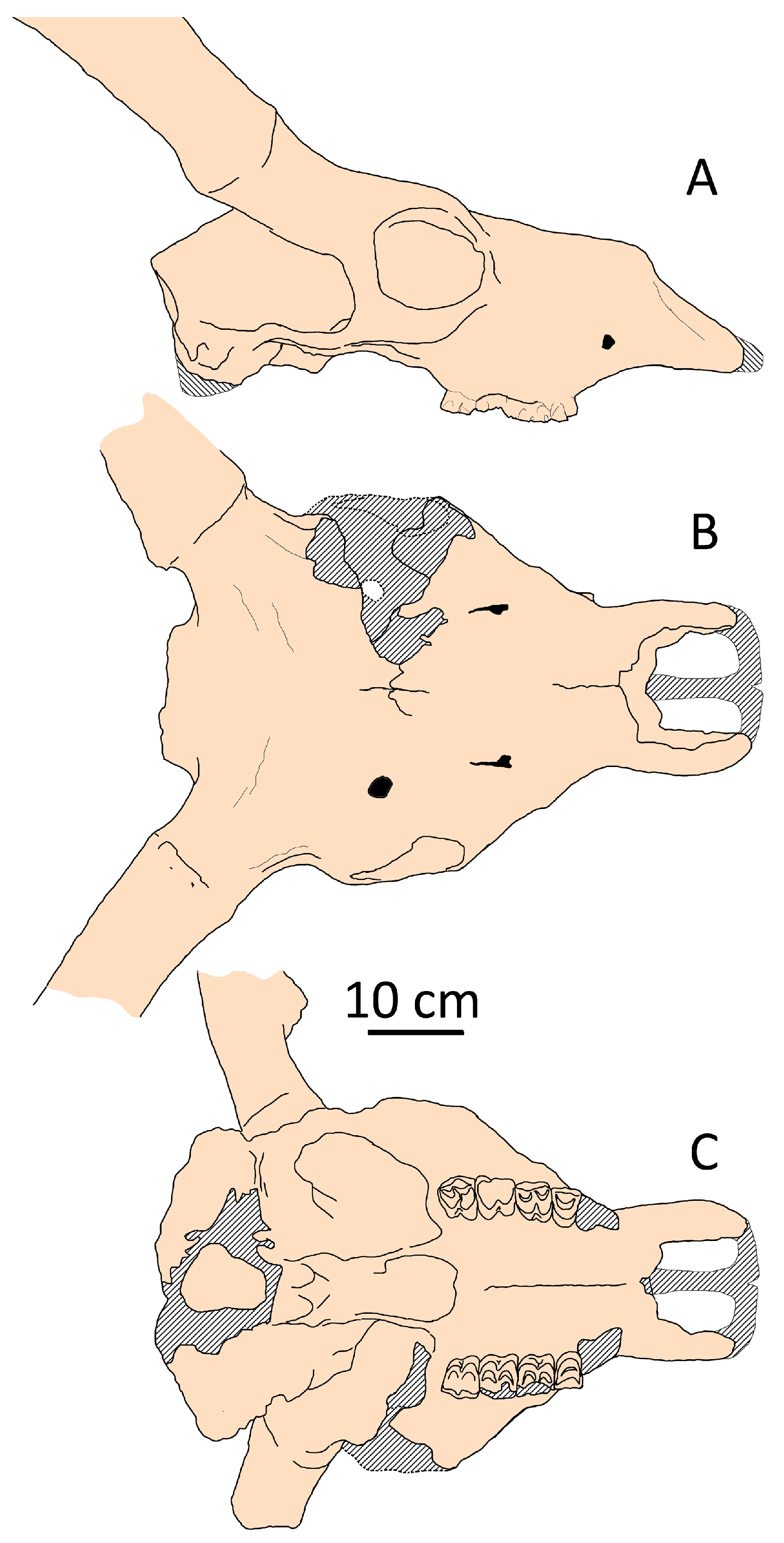
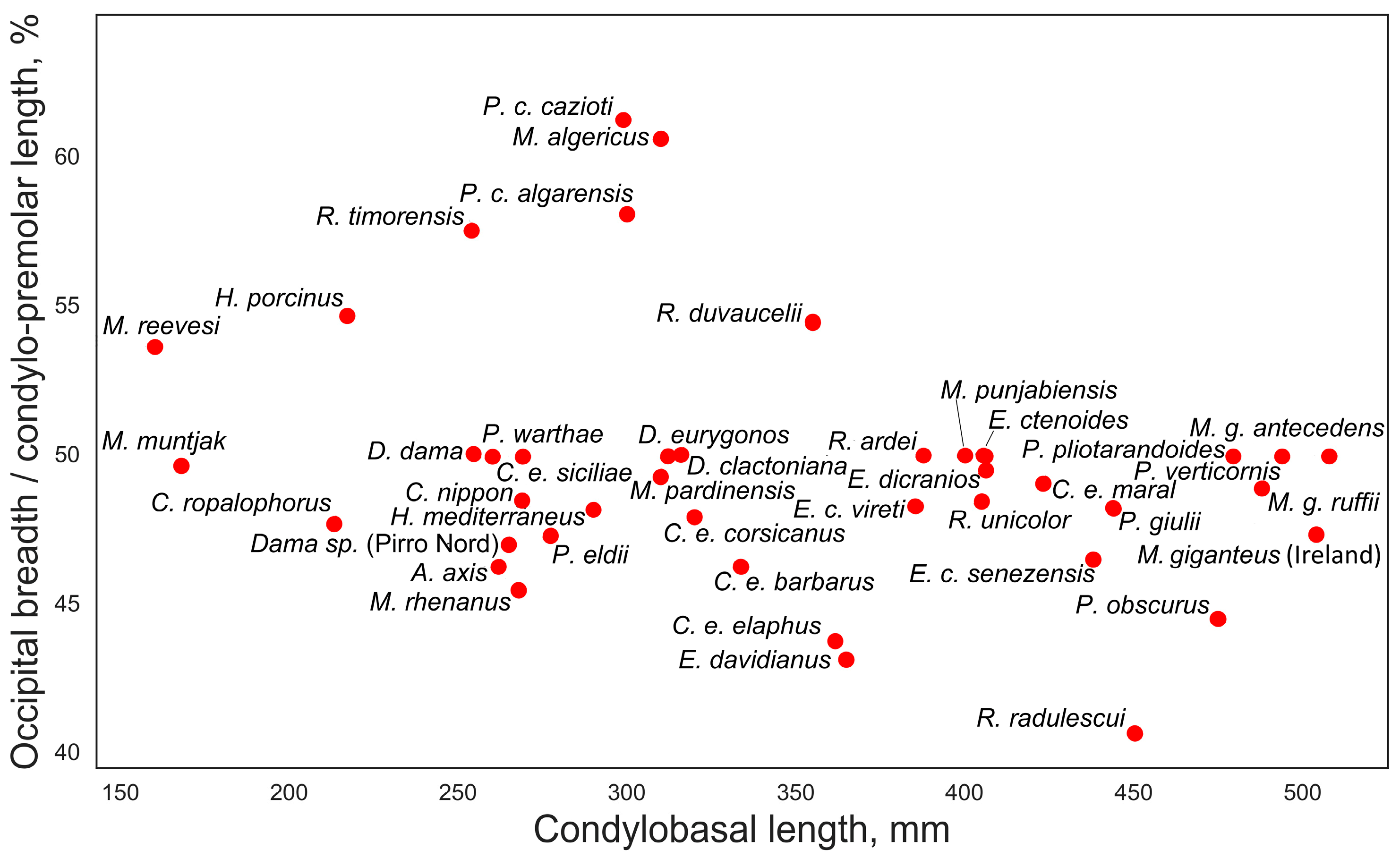
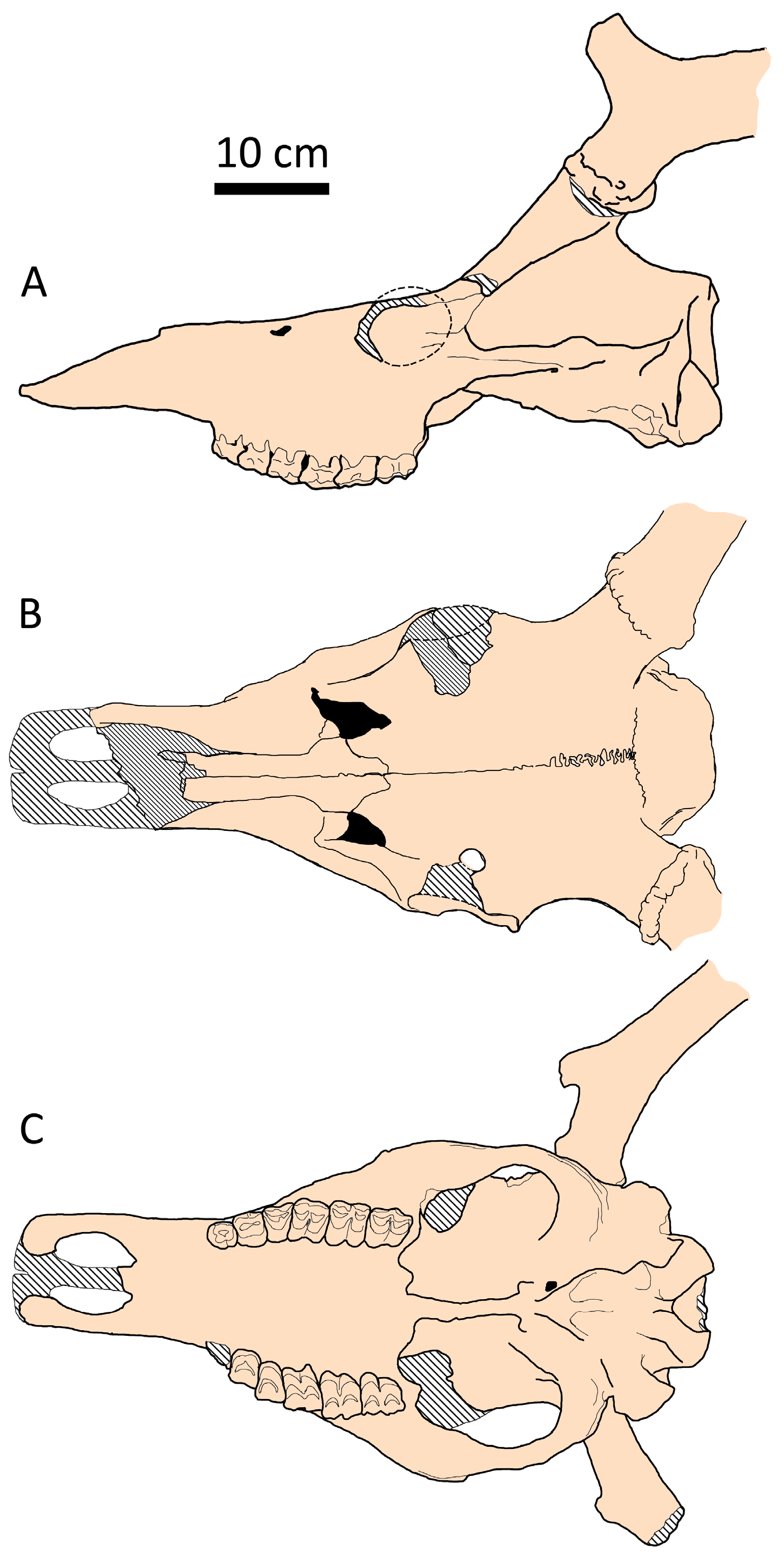
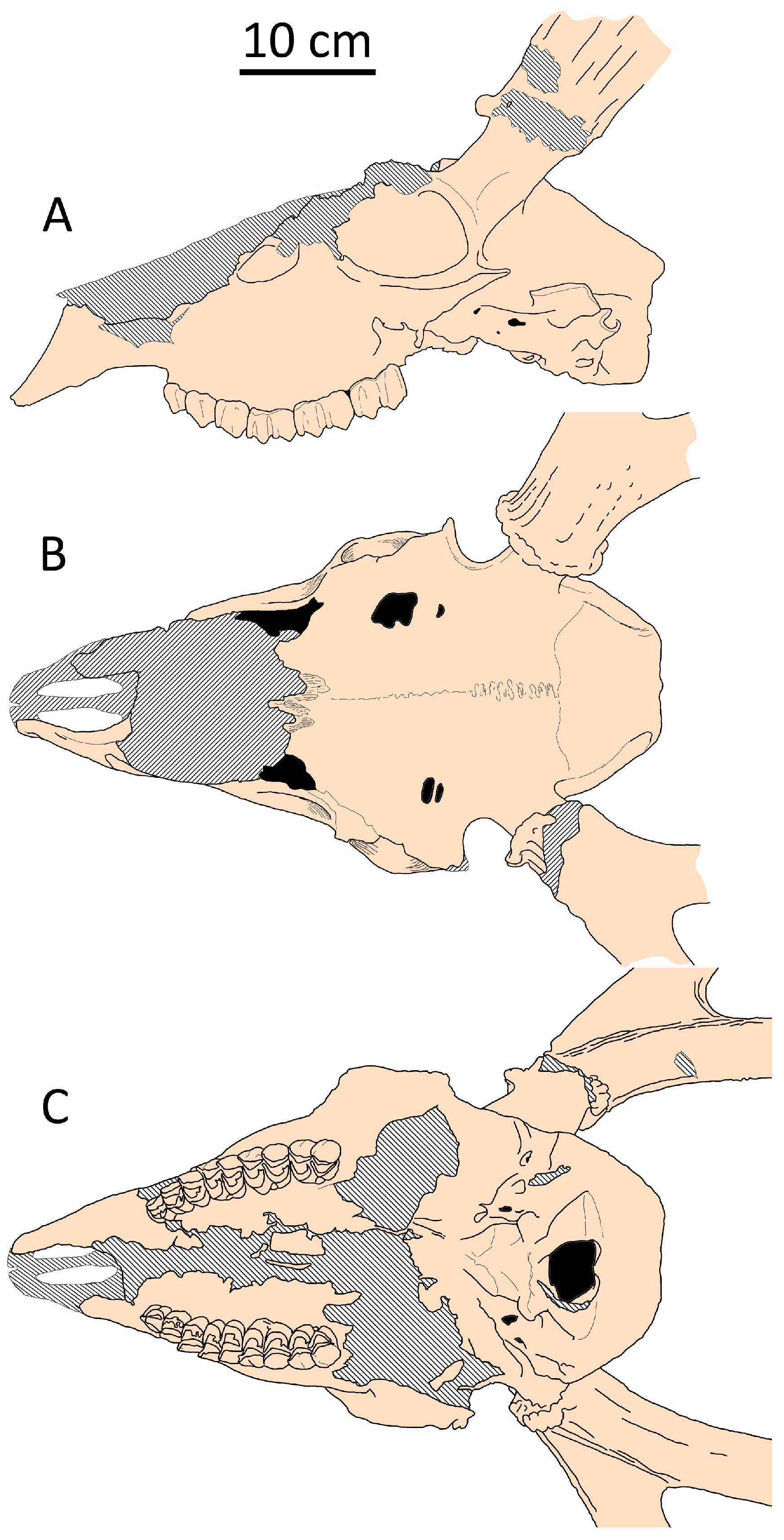
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).