

First Evidence of Reproductive Strategies in Cephalopods Preserved in Phosphate and Siderite Nodules from the Devonian of Uruguay


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
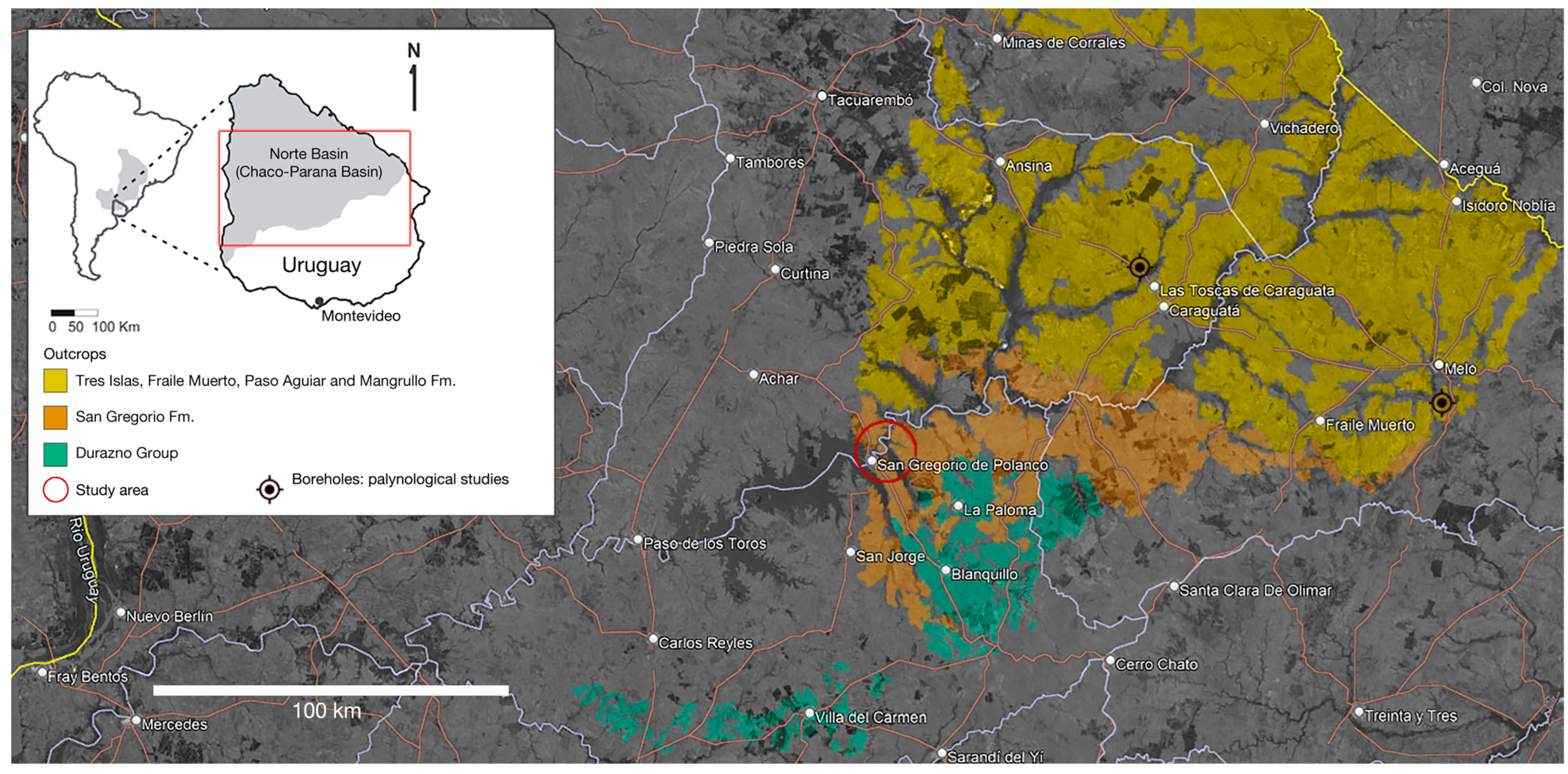
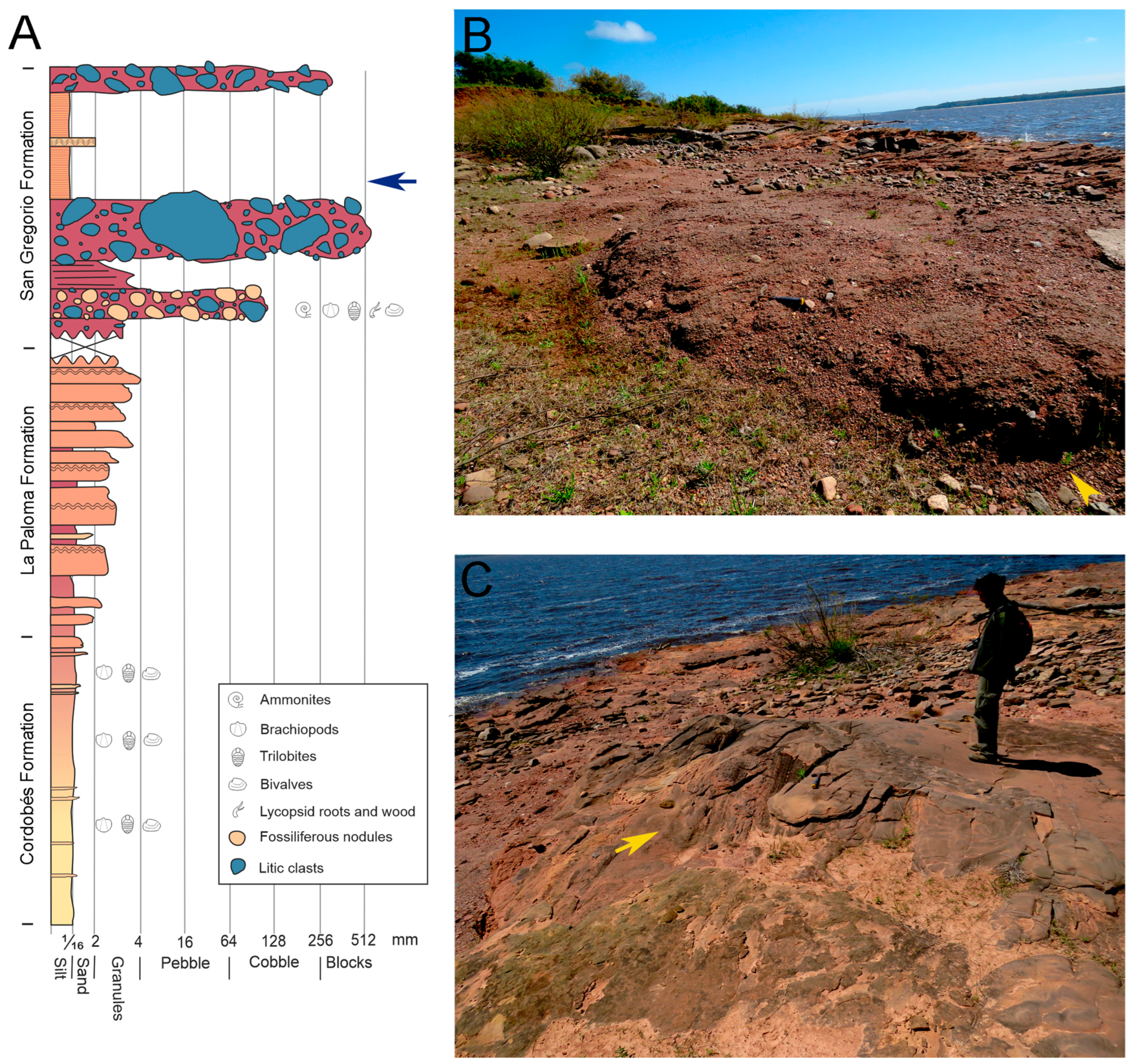
1.1. Geological Setting
1.2. Ammonoid Reproductive Strategies
2. Materials and Methods
3. Results
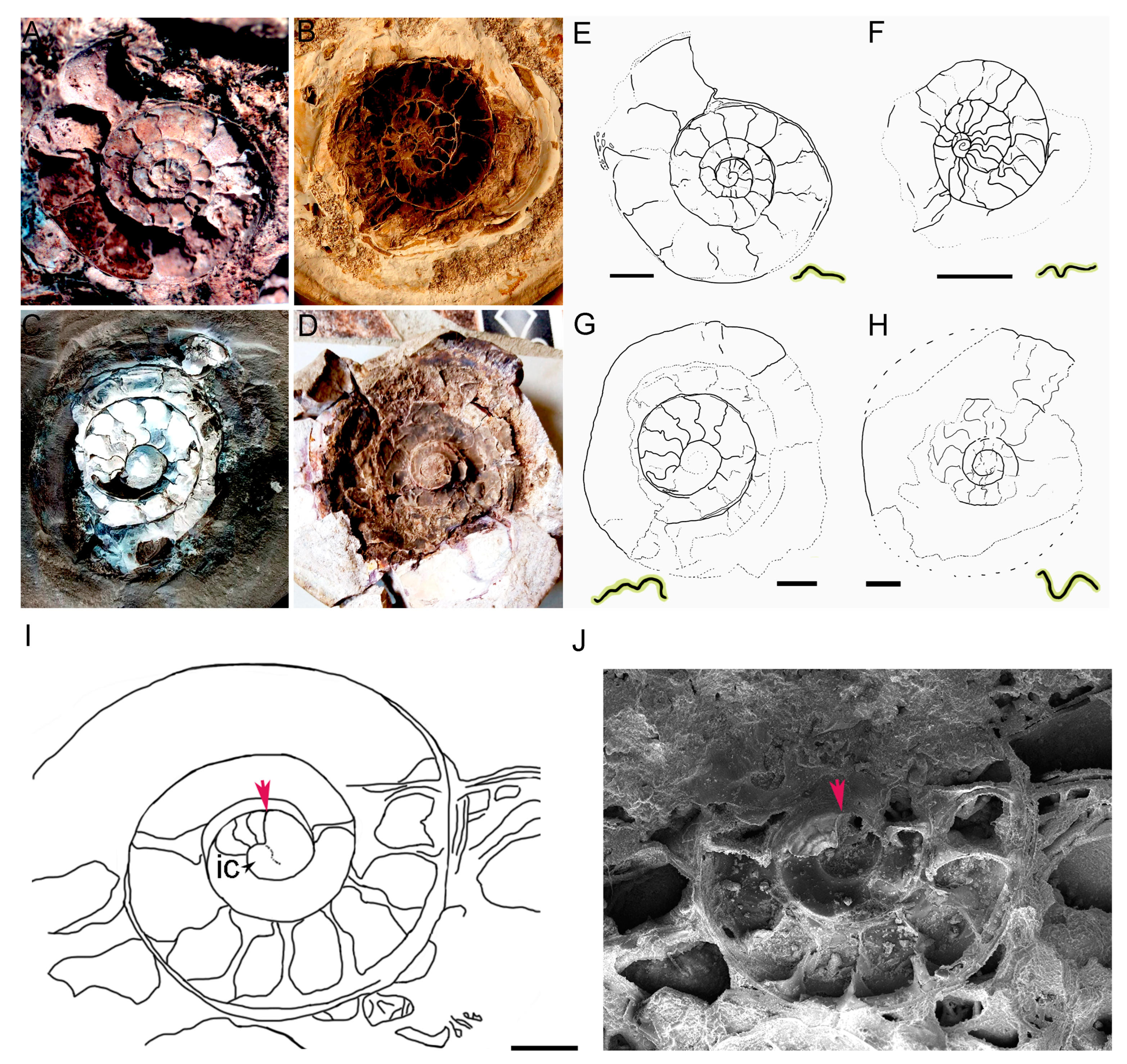
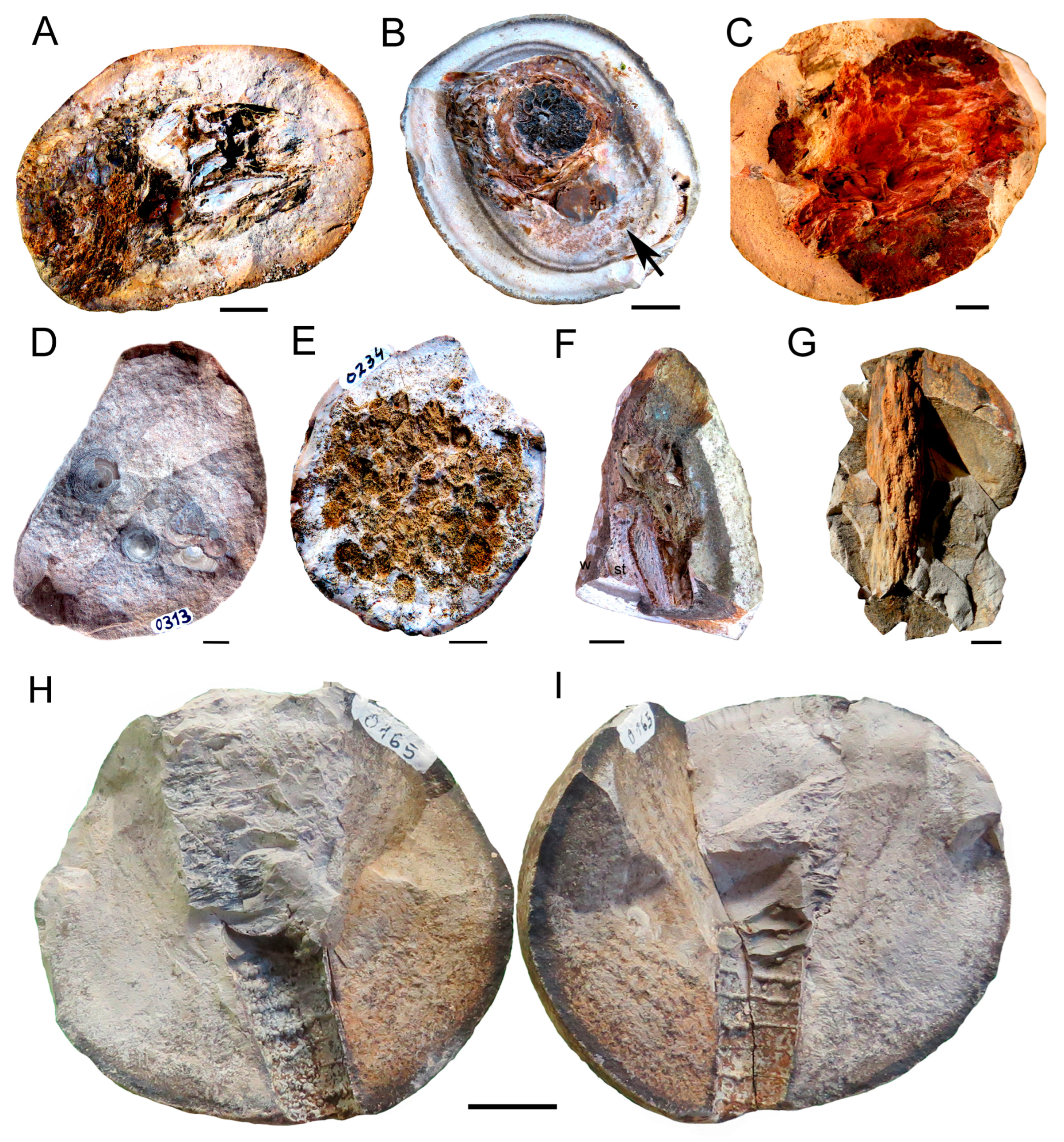
3.1. The Ammonitella Stage in the Early Juvenile Ammonoid Clusters

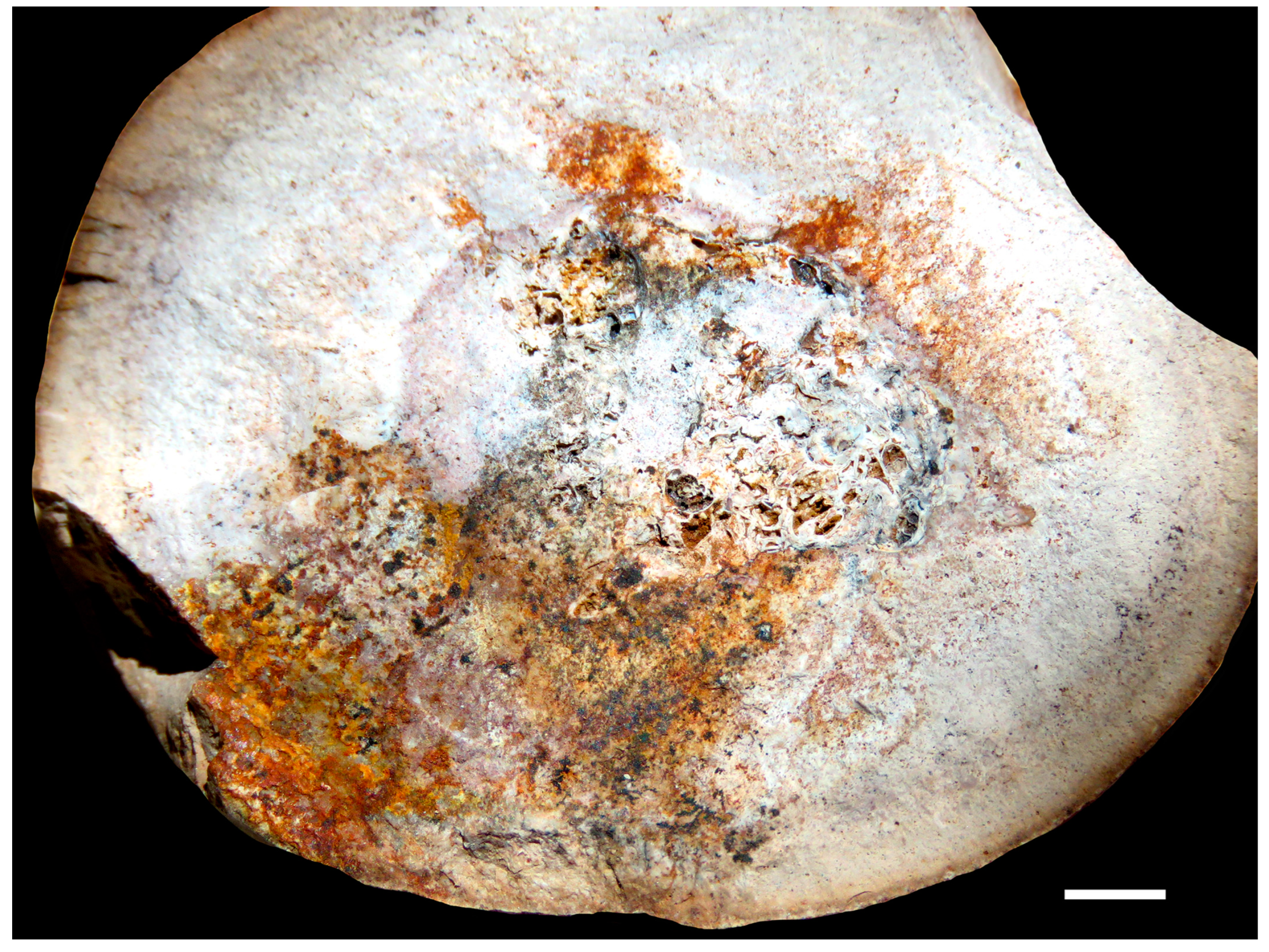

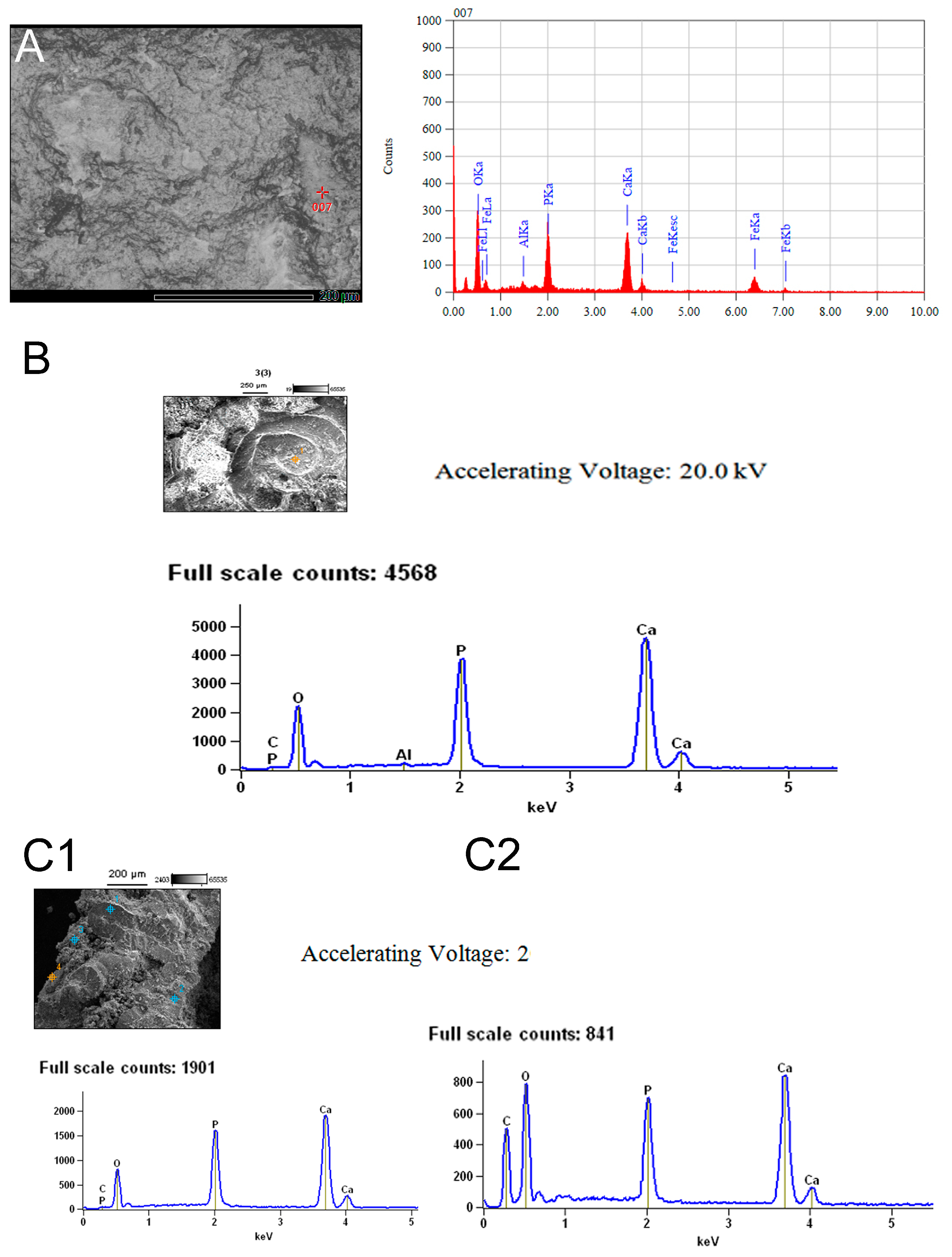
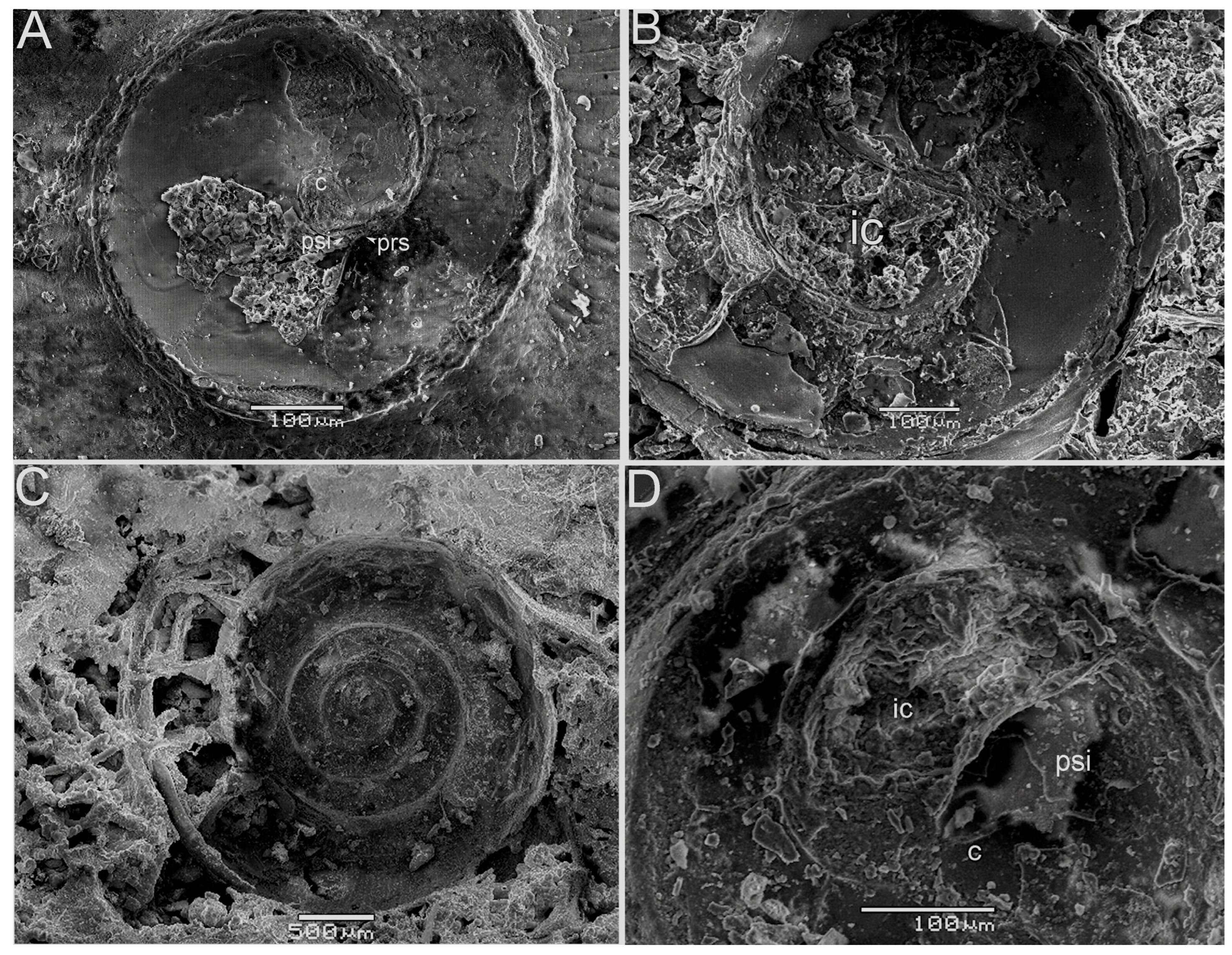

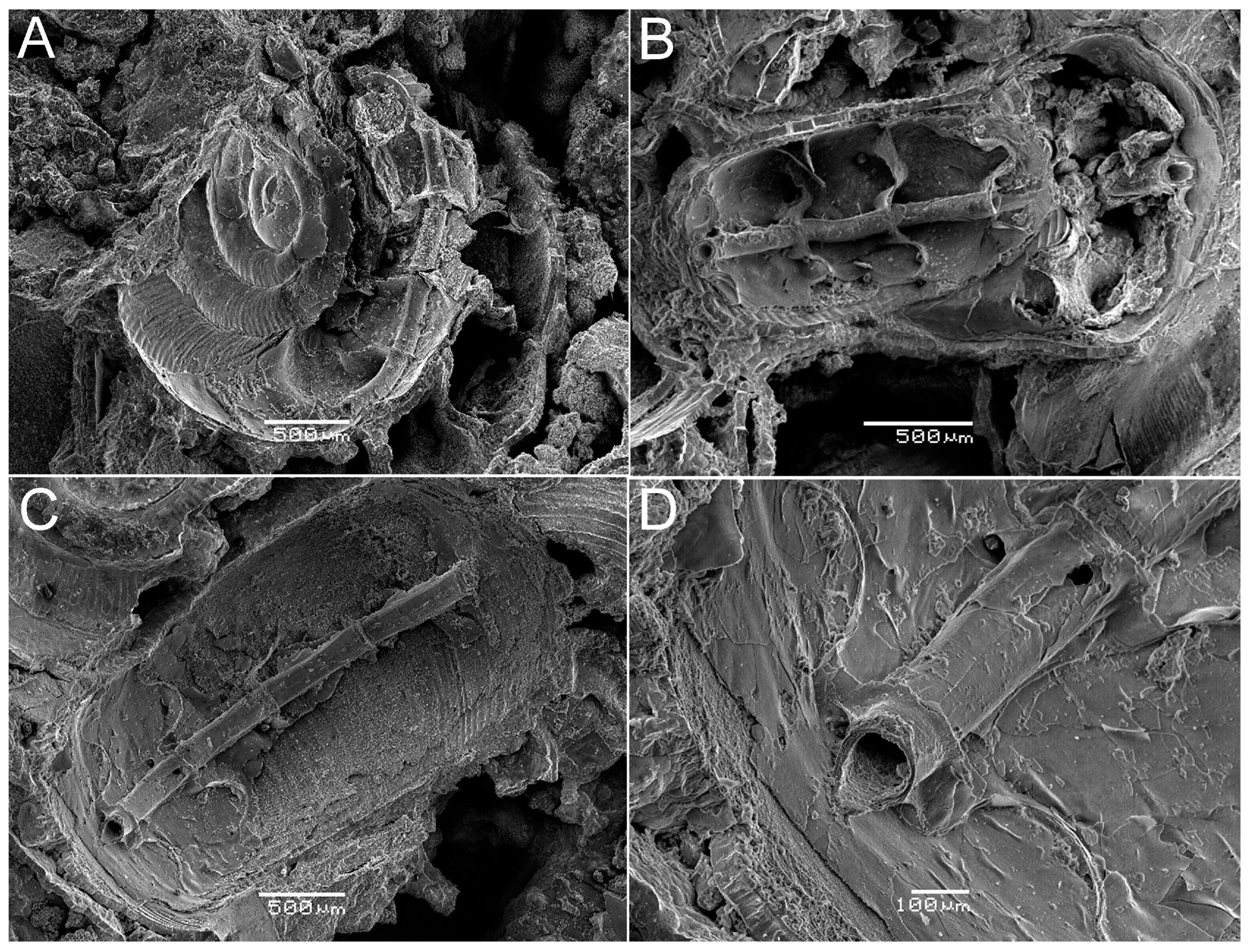

3.2. Chemical Composition of the Shells in Ammonoid Clusters from Uruguay
4. Discussion
4.1. Available Evidence of Ammonoid Reproductive Strategies
4.2. Ammonoid Clusters Being Gastric Contents or Coprolites
4.3. Taxonomic Identification of the Ammonoid Clusters
4.4. The Devonian Age of the Nodules from Biostratigraphic and Paleogeographic Evidence
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Beri, Á.; Martínez-Blanco, X.; Mourelle, D. A synthesis of palynological data from the Lower Permian Cerro Pelado Formation (Paraná Basin, Uruguay); A record of warmer climate stages during Gondwanan glaciations. Geol. Acta 2010, 8, 419–429. [Google Scholar] [CrossRef]
- De Santa Ana, H.; Beri, A.; Goso, C.; Daners, G. Análisis estratigráfico de la Formación San Gregorio (Pérmico Inferior), en los testigos del pozo Cerro Largo Sur Nº4 (Di.Na.Mi.Ge.), Uruguay. Rev. Bras. Geociências 1993, 23, 347–351. [Google Scholar] [CrossRef]
- Closs, D.; Gordon, M.J. An upper Paleozoic goniatite radula. Esc. Geol. Notas E Estud. 1966, 1, 73–75. [Google Scholar]
- Beltán, L. Dècuverte d’une ichtyofaune dans le Carbonifère Superieur d’Uruguay. Rapport avec les faunes ichthyologiques contemporaines des autres régions du Gondwana. Ann. Société Géologique du Nord. Lille 1977, 97, 351–355. [Google Scholar]
- Beltán, L. Coccocephalithchys tessellatus n. sp. (Pisces, Actinopterygii) from the Upper Carboniferous of Uruguay. An. Do II Congr. Lat.-Am. Paleontol. 1981, 1, 95–105. [Google Scholar]
- Beri, A.; Cernuschi, F.; Martínez-Blanco, X. Palynology of the San Gregorio and Melo Formations in Borehole 254 (Dinamige), Late Paleozoic, Paraná Basin, Uruguay. Ameghiniana 2011, 48, 480–495. [Google Scholar] [CrossRef]
- Closs, D. Goniatiten mit Radula und Kieferapparat in der Itararé Formation von Uruguay. Paläontologische Z. 1967, 41, 19–37. [Google Scholar]
- Closs, D. Upper Carboniferous anaptychi from Uruguay. Ameghiniana (Rev. Asoc. Paleontológica Argent.) 1967, 5, 145–148. [Google Scholar]
- Piñeiro, G.; Estramil, C.; Méndez Ortiz, J.M.; Núñez Demarco, P. First evidence of reproductive strategies in Devonian cephalopods preserved in siderite nodules from Uruguay. In Proceedings of the IV Palaeontological Virtual Congress, Thematic Section: Palaeontological Scenarios of the Paraná Basin, Virtual, 1–22 May 2023. [Google Scholar]
- De Santa Ana, H.; Goso, C.; Daners, G. Cuenca Norte: Estratigrafía del Carbonífero-Pérmico. In Cuencas Sedimentarias de Uruguay: Geología, Paleontología y Recursos Naturales; Veroslavsky, G., Ubilla, M., Martínez, S., Eds.; Facultad de Ciencias de Uruguay (DIRAC): Montevideo, Uruguay, 2006; pp. 147–208. [Google Scholar]
- Bossi, J. Geología del Uruguay. In Colección Ciencias, 2nd ed.; Universidad de la República: Montevideo, Uruguay, 1966; 419p. [Google Scholar]
- Caorsi, J.; Goñi, J. Geología Uruguaya. Inst. Geológico Urug. Boletín Montev. 1958, 37, 1–73. [Google Scholar]
- Fedorchuk, N.D.; Griffisc, N.P.; Isbell, J.L.; Goso CRosa, E.L.M.; Montañez, I.P.; Yinc, Q.Z.; Huyskensc, M.H.; Sanbornc, M.E.; Mundile, R.; Vesely, F.F.; et al. Provenance of late Paleozoic glacial/post-glacial deposits in the eastern Chaco-Paraná Basin, Uruguay and southernmost Paraná Basin, Brazil. J. S. Am. Earth Sci. 2021, 106, 102989. [Google Scholar] [CrossRef]
- Martin, H. The Late Palaeozoic Gondwana glaciations. Geol. Rundsch. 1981, 70, 488–496. [Google Scholar] [CrossRef]
- Sprechmann, P.; Montaña, J.; Gaucher, C. Devónico. In Geología del Departamento de Durazno; Boletin de Facultad de Agronomía: Montevideo, Uruguay, 1993; pp. 25–55. [Google Scholar]
- Uriz, N.J.; Cingolani, C.A.; Basei, M.A.S.; Blanco, G.; Abre, P.; Potillo, N.S.; Sicardi, A. Provenance and paleogeography of the Devonian Durazno Group, southern Parana Basin in Uruguay. J. S. Am. Earth Sci. 2016, 66, 248–267. [Google Scholar] [CrossRef]
- Griffis, N.P.; Mundil, R.; Montañez, I.P.; Isbell, J.L.; Fedorchuk, N.; Vesely, F.; Iannuzzi, R.; Yin, Q.Z. A new stratigraphic framework built on U-Pb single-zircon TIMS ages and implications for the timing of the penultimate icehouse (Paraná Basin, Brazil). GSA Bull. 2018, 130, 848–858. [Google Scholar] [CrossRef]
- Césari, S.; Limarino, C.O.; Gulbranson, E.L. An Upper Paleozoic bio-chronostratigraphic schem for the western margin of Gondwana. Earth-Sci. Rev. 2011, 106, 149–160. [Google Scholar] [CrossRef]
- Braun, A.; Sprechmann, P.; Gaucher, C. Stratigraphic age of phosphorite-nodules from the San Gregorio Formation of Uruguay. Neues Jahrb. Geol. Paläontologie Monatshefte 2003, 12, 739–748. [Google Scholar] [CrossRef]
- Pirez, E.; Méndez, J.M.; Estramil, C.; Piñeiro, G. El Río Hum (atual Río Negro) antes y después de la Represa de Rincón del Bonete en San Gregorio de Polanco (Tacuarembó, Uruguay). J. Anim. Environ. Res. (Edición Espec.) 2022, 5, 131–155. [Google Scholar]
- Isbell, J.L.; Henry, L.C.; Gulbranson, E.L.; Limarino, C.O.; Fraiser, M.L.; Koch, Z.J.; Ciccioli, P.L.; Dineen, A.A. Glacial paradoxes during the late Paleozoic ice age: Evaluating the equilibrium line altitude as a control on glaciation. Gondwana Res. 2012, 22, 1–19. [Google Scholar] [CrossRef]
- Iannuzzi, R.; di Pasquo, M.M.; Vesely, F.F.; Scherer, C.M.; Andrade, L.S.; Mottin, T.; Kifumbi, C. Pennsylvanian glacial cycles in western Gondwana: An overview. Geol. Soc. Lond. Spec. Publ. 2023, 535, 101–116. [Google Scholar] [CrossRef]
- Drushchits, V.V.; Doguzhayeva, L.A.; Mikhaylova, I.A. The structure of the ammonitella and the direct development of ammonites. Palaeontol. J. 1977, 2, 188–199. [Google Scholar]
- Etches, S.; Clarke, J.; Callomon, J. Ammonite eggs and ammonitellae from the Kimmeridge Clay Formation (Upper Jurassic) of Dorset, England. Lethaia 2009, 42, 204–217. [Google Scholar] [CrossRef]
- Landman, N.H.; Tanabe, K.; Shigeta, Y. Ammonoid embryonic development. In Ammonoid Paleobiology; Landman, N.H., Tanabe, K., Eds.; Springer: Boston, MA, USA, 1996; Volume 67, pp. 361–437. [Google Scholar]
- Landman, N.H.; Mapes, R.H.; Tanabe, K. Internal features of the internal shell of Late Carboniferous Goniatitina. In Advanced Research on Living and Fossil Cephalopods; Rodríguez-Tovar, O., Ed.; Kluver Academic/Plenum Publishers: New York, NY, USA, 1999; pp. 242–254. [Google Scholar]
- Tanabe, K.; Landman, N.H.; Mapes, R.H.; Faulkner, C.J. Analysis of a Carboniferous embryonic ammonoid assemblage, implications for ammonoid embryology. Lethaia 1993, 26, 215–224. [Google Scholar] [CrossRef]
- De Baets, K.; Landman, N.H.; Tanabe, K. Ammonoid embryonic development. In Ammonoid Paleobiology: From Anatomy to Ecology; Klug, C., Korn, D., De Baets, K., Kruta, I., Mapes, R.N., Eds.; Springer: Amsterdam, The Netherlands, 2015; pp. 113–205. [Google Scholar]
- Erben, H.K. Bactritoidea. In Treatise on Invertebrate Paleontology, Part K, Mollusca 3, Cephalopoda; Moore, R.C., Ed.; Geological Society of America and University of Kansas Press: Lawrence, Kansas, 1964; pp. K491–K519. [Google Scholar]
- Mironenko, A.A.; Rogov, M.A. First direct evidence of ammonoid ovoviviparity. Lethaia 2016, 49, 245–260. [Google Scholar] [CrossRef]
- Westermann, G.E.G. Ammonoid life and habitat. In Ammonoid Paleobiology; Landman, N.H., Tanabe, K., Davis, R.A., Eds.; Topics in Geobiology; Plenum Press: New York, NY, USA, 1996; Volume 13, pp. 607–707. [Google Scholar]
- Laptikhovsky, V.; Nikolaeva, S.; Rogov, M. Cephalopod embryonic shells as a tool to reconstruct reproductive strategies in extinct taxa. Biol. Rev. 2017, 93, 270–283. [Google Scholar] [CrossRef] [PubMed]
- Vidal, E.A.; Shea, E.K. Cephalopod ontogeny and life cycle patterns. Frontiers of Marine Sciences. Sec. Mar. Biol. 2023, 10, 1162735. [Google Scholar] [CrossRef]
- Landman, N.H. Preserved ammonitellas of Scaphites (Ammonoidea, Ancyloceratina). Am. Mus. Novit. 1985, 2815, 1–10. [Google Scholar]
- Lukeneder, A. Ammonoid habitats and life history. In Ammonoid Paleobiology: From Anatomy to Ecology; Klug, C., Korn, D., De Baets, K., Kruta, I., Mapes, R.H., Eds.; Springer: Amsterdam, The Netherlands, 2015; pp. 689–791. [Google Scholar]
- Wani, R. Geological duration of ammonoids controlled their geographical range of fossil distribution. PeerJ 2017, 5, e4108. [Google Scholar] [CrossRef]
- Laptikhovsky, V.L.; Rogov, M.A.; Nikolaeva, S.E.; Arkhipkin, A.I. Environmental impact on ectocochleate cephalopod reproductive strategies and the evolutionary significance of cephalopod egg size. Bull. Geosci. 2013, 88, 83–93. [Google Scholar] [CrossRef]
- Tanabe, K.; Sasaki, T.; Mapes, R.H. Soft-part anatomy of the siphuncle in ammonoids. In Ammonoid Paleobiology: From Anatomy to Ecology; Klug, C., Korn, D., De Baets, K., Kruta, I., Mapes, R.H., Eds.; Springer: Amsterdam, The Netherlands, 2015; pp. 531–544. [Google Scholar] [CrossRef]
- Vidal, E.A.; Haimovici, M. Feeding and the possible roll of the proboscis and mucus cover in the ingestion of microor ganisms by Rhynchoteuthion paralarvae (Cephalopoda: Ommastrephidae). Bull. Mar. Sci. 1998, 63, 305–316. [Google Scholar]
- Mapes, R.H.; Nützel, A. Late Palaeozoic mollusc reproduction: Cephalopod egg-laying behavior and gastropod larval palaeobiology. Lethaia 2009, 42, 341–356. [Google Scholar] [CrossRef]
- Closs, D. Intercalation of Goniatites in the Gondwanic Glacial Beds of Uruguay. Gondwana Stratigraphy. IUGS Symposium Buenos Aires, 1–15 October 1967; Paris Unesco, 1969. pp. 197–212. Available online: https://catalog.library.vanderbilt.edu/discovery/fulldisplay/alma991017205269703276/01VAN_INST:vanui (accessed on 18 June 2024).
- Lhemann, J.; Klug, C.; Wild, F. Did ammonoids possess opercula? Reassessment of phosphatized soft tissues in Glaphyrites of the Carboniferous of Uruguay. Palaeontol. Z. 2014, 89, 63–77. [Google Scholar] [CrossRef]
- Kulicki, C.; Doguzhaeva, L.A. Development and calcification of the ammonitella shell. Acta Palaeontol. Pol. 1994, 39, 17–44. [Google Scholar]
- Stephen, D.A.; Bylund, K.G.; García, P.; McShinsky, R.D.; Carter, H.J. Taphonomy of dense concentrations of juvenile ammonoids in the Upper Cretaceous Mancos Shale, east-central Utah, USA. Geobios 2012, 45, 121–128. [Google Scholar] [CrossRef]
- Stinnesbeck, W.; Frey, E.; Zell, P. Evidence for semi-sessile early juvenile life history in Cretaceous ammonoids. Palaeogeogr. Palaeoclimatol. Palaeocol. 2016, 457, 186–194. [Google Scholar] [CrossRef]
- Klug, C.; De Baets, K.; Korn, D. Exploring the limits of morphospace: Ontogeny and ecology of late Viséan ammonoids from the Tafilalt, Morocco. Acta Palaeontol. Pol. 2016, 61, 1–14. [Google Scholar] [CrossRef]
- Greif, M.; Nebelsick, J.H.; Klug, C. Extreme abundance of ammonoids in mass accumulations from the Late Devonian of the Moroccan Anti-Atlas. Acta Palaeontol. Pol. 2022, 67, 667–684. [Google Scholar] [CrossRef]
- Tajika, A.; Wani, R. Intraspecific variation of hatchling size in Late Cretaceous ammonoids from Hokkaido, Japan: Implication for planktic duration at early ontogenetic stage. Lethaia 2011, 44, 287–298. [Google Scholar] [CrossRef]
- Korn, D. A key for the description of Paleozoic ammonoids. Foss. Rec. 2010, 13, 5–12. [Google Scholar] [CrossRef]
- De Baets, K.; Klug, C.; Korn, D.; Landman, N.H. Early evolutionary trends in ammonoid embryonic development. Evolution 2012, 66, 1788–1806. [Google Scholar] [CrossRef] [PubMed]
- Piñeiro, G.; Ramos, A.; Goso, C.; Scarabino, F.; Laurin, M. Unusual environmental conditions preserve a Permian mesosaur-bearing Konservat- Lagerstätte from Uruguay. Acta Palaeontol. Pol. 2012, 57, 299–318. [Google Scholar] [CrossRef]
- Calisto, V.; Piñeiro, G. A large cockroach from the mesosaur-bearing Konservat-Lagerstätte (Mangrullo Formation), Late Paleozoic of Uruguay. PeerJ 2019, 7, e6289. [Google Scholar] [CrossRef]
- Piñeiro, G.; Demarco, P.N.; Meneghel, M.D. The ontogenetic transformation of the mesosaurid tarsus: A contribution to the origin of the primitive amniotic astragalus. PeerJ 2016, 4, e2036. [Google Scholar] [CrossRef]
- Núñez Demarco, P.; Ferigolo, J.; Piñeiro, G. Isometry in mesosaurs: Implications for growth patterns in early amniotes. Acta Palaeontol. Pol. 2022, 67, 509–542. [Google Scholar] [CrossRef]
- Bossi, J.; Navarro, R. Geología del Uruguay; Departamento de Publicaciones, Universidad de la República: Montevideo, Uruguay, 1991; 970p. [Google Scholar]
- von Huene, F. Osteologie und systematische Stellung von Mesosaurus. Palaeontographica 1941, 92, 45–58. [Google Scholar]
- Laurin, M.; Piñeiro, G. A reassessment of the taxonomic position of mesosaurs, and a surprising phylogeny of early amniotes. Front. Earth Sci. 2017, 5, 88. [Google Scholar] [CrossRef]
- Bridge, T.C.L.; Baird, A.H.; Pandolfi, J.M.; McWilliam, M.J.; Zapalski, M.K. Functional consequences of Palaeozoic reef collapse. Sci. Rep. 2022, 12, 1386. [Google Scholar] [CrossRef] [PubMed]
- Zapalski, M.K.; Baird, A.H.; Bridge, T.; Jakubowicz, M.; Daniell, J. Unusual shallow water Devonian coral community from Queensland and its recent analogues from the inshore Great Barrier Reef. Coral Reefs 2021, 40, 417–431. [Google Scholar] [CrossRef]
- Dzik, J. Phylogeny od the Nautiloidea. Paleontol. Pol. 1984, 45, 323. [Google Scholar]
- Sharpe, D. Descriptions of Paleozoic Mollusca from South Africa. Trans. Geol. Soc. Lond. 1856, 2, 206–215. [Google Scholar]
- Carbonaro, F.A.; Comniskey, J.C.; Corral Stok, H.; Pirani Ghilardi, R. Orbiculoidea baini and Orbiculoidea excentrica (Brachiopoda, Discinidae) from the Middle Devonian (Alto Garças Sub-basin, Paraná Basin) of Caiapônia, Goiás (Brazil). Rev. Inst. Geociências—USP Série Cient. 2018, 18, 11–20. [Google Scholar] [CrossRef]
- Penn-Clarke, C.R.; Harper, D.A.T. The rise and fall of the Malvinoxhosan (Malvinokaffric) bioregion in South Africa: Evidence for Early-Middle Devonian biocrises at the South Pole. Earth Sci. Rev. 2023, 246, 104595. [Google Scholar] [CrossRef]
- Da Silva, A.-C.; Kershaw, S.; Boulvain, F.; Hubert, B.; Mistiaen, B.R.; Reynolds, A.; Reitner, J. Indigenous demosponge spicules in a Late Devonian stromatoporoid basal skeleton from the Frasnian of Belgium. Lethaia 2014, 47, 365–375. [Google Scholar] [CrossRef]
- House, M.R. Devonian goniatites from Nevada. Newes Jahrb. Geol. Paläontologie Abh. 1965, 122, 339–342. [Google Scholar]
- Klug, C. Quantitative stratigraphy and taxonomy of late Emsian and Eifelian ammonoids of the eastern Anti-Atlas (Morocco). Cour. Forschungsinstitut Senckenberg 2002, 238, 1–109. [Google Scholar]
- Becker, R.T.; Hartenfels, S.; Klug, C.; Aboussalam, Z.S.; Afhüppe, L. The cephalopod-rich Famennian and Tournaisian of the Aguelmous Syncline (southern Maïder). Münstersche Forschungsberichte Geol. Paläontologie 2018, 110, 273–306. [Google Scholar]
- Becker, R.T.; House, M.R. Emsian and Eifelian ammonoid succession at Bou Tchrafine (Tafilalt platform, Anti-Atlas, Morocco). Notes Mémoires Serv. Geol. 2000, 399, 21–26. [Google Scholar]
- Carrera, M.G.; Montoya, E.; Rustán, J.J.; Halpern, K. Silurian-Devonian coral associations across a sequence stratigraphic boundary in the Argentine Precordillera. Geol. J. 2013, 48, 256–269. [Google Scholar] [CrossRef]
- Majchrzyk, A.; Jakubowicz, M.; Berkowski, B.; Król, J.J.; Zato, M.; Zapalski, M.K. Modern-type reef in ancient time—Palaeoecology of a Middle Devonian coral community from Madène el Mrakib (Anti-Atlas, Morocco). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2024, 633, 111876. [Google Scholar] [CrossRef]
- Naglik, C.; De Baets, K.; Klug, C. Early Devonian ammonoid faunas in the Zeravshan Mountains (Uzbekistan and Tadjikistan) and the transition from a carbonate platform setting to pelagic sedimentation. Bull. Geosci. 2019, 94, 337–368. [Google Scholar] [CrossRef]
- Zapalski, M.K.; Nowicki, J.; Jakubowicz, M.; Berkowski, B. Tabulate corals across the Frasnian/Famennian boundary: Architectural turnover and its possible relation to ancient photosymbiosis. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2017, 487, 416–429. [Google Scholar] [CrossRef]
- Scotese, C.R.; Song, H.; Mills, B.J.; van der Meer, D.G. Phanerozoic paleotemperatures: The earth’s changing climate during the last 540 million years. Earth Sci. Rev. 2021, 215, 103503. [Google Scholar] [CrossRef]
- Becker, R.T.; Kullmann, J. Paleozoic ammonoids in space and time. In Ammonoid Paleobiology; Landman, N.H., Davis, R.A., Tanabe, K., Eds.; Topics in Geobiology; Plenum Press: New York, NY, USA, 1996; Volume 13, pp. 711–753. [Google Scholar]
- Seilacher, A. Begriff und Bedeutung der Fossil−Lagerstätten. Neues Jahrb. Geol. Paläontologie Monatshefte 1970, 1970, 34–39. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piñeiro, G.; Rodao, M.; Núñez Demarco, P. First Evidence of Reproductive Strategies in Cephalopods Preserved in Phosphate and Siderite Nodules from the Devonian of Uruguay. Foss. Stud. 2024, 2, 223-244. https://doi.org/10.3390/fossils2030011
Piñeiro G, Rodao M, Núñez Demarco P. First Evidence of Reproductive Strategies in Cephalopods Preserved in Phosphate and Siderite Nodules from the Devonian of Uruguay. Fossil Studies. 2024; 2(3):223-244. https://doi.org/10.3390/fossils2030011
Chicago/Turabian StylePiñeiro, Graciela, Magela Rodao, and Pablo Núñez Demarco. 2024. "First Evidence of Reproductive Strategies in Cephalopods Preserved in Phosphate and Siderite Nodules from the Devonian of Uruguay" Fossil Studies 2, no. 3: 223-244. https://doi.org/10.3390/fossils2030011