
A New Tyrant Dinosaur from the Late Campanian of Mexico Reveals a Tribe of Southern Tyrannosaurs †



Abstract
:1. Introduction
Geology

2. Materials and Methods
3. Results
- Systematic paleontology
- Etymology
- Holotype
- Referred specimens
- Diagnosis
- Horizon and Locality
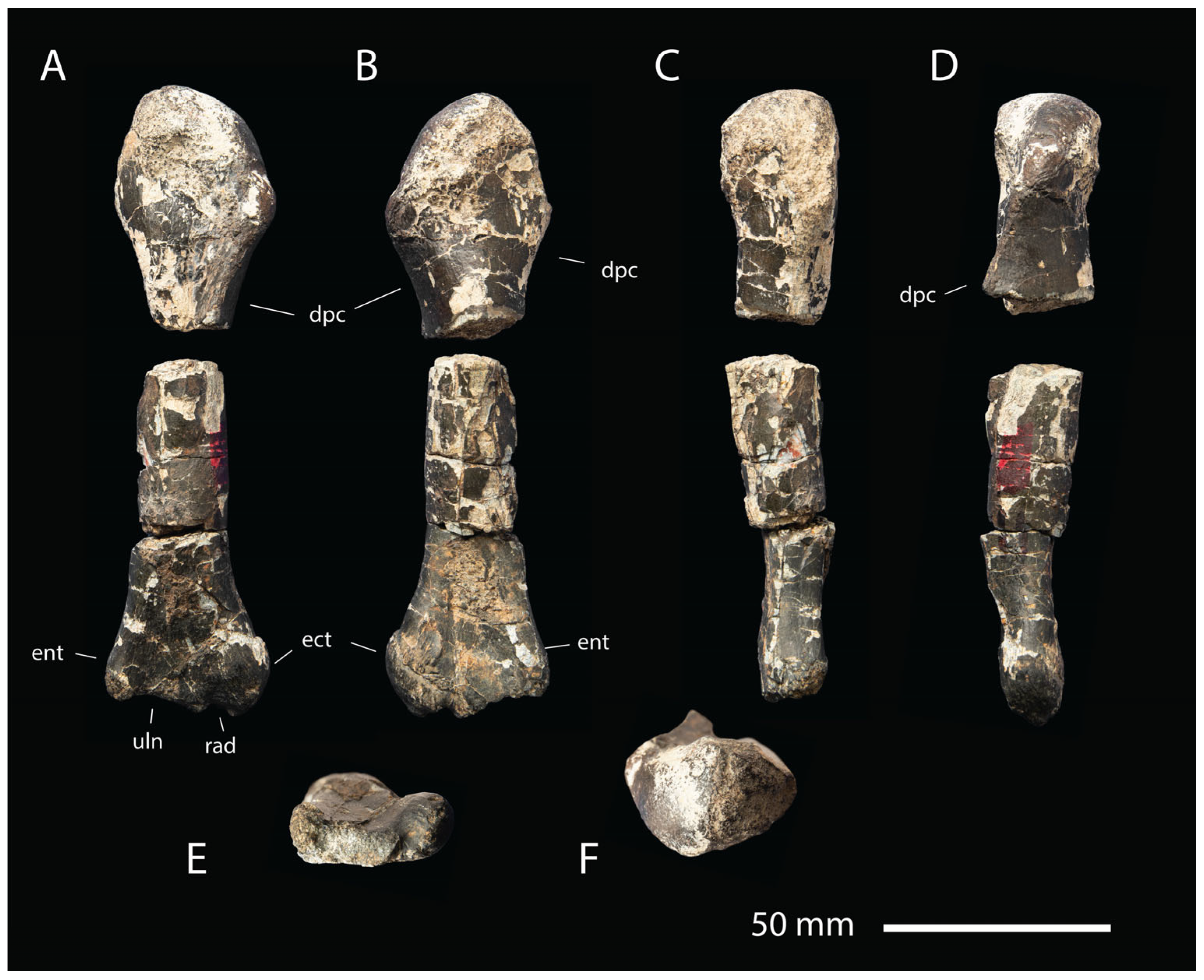
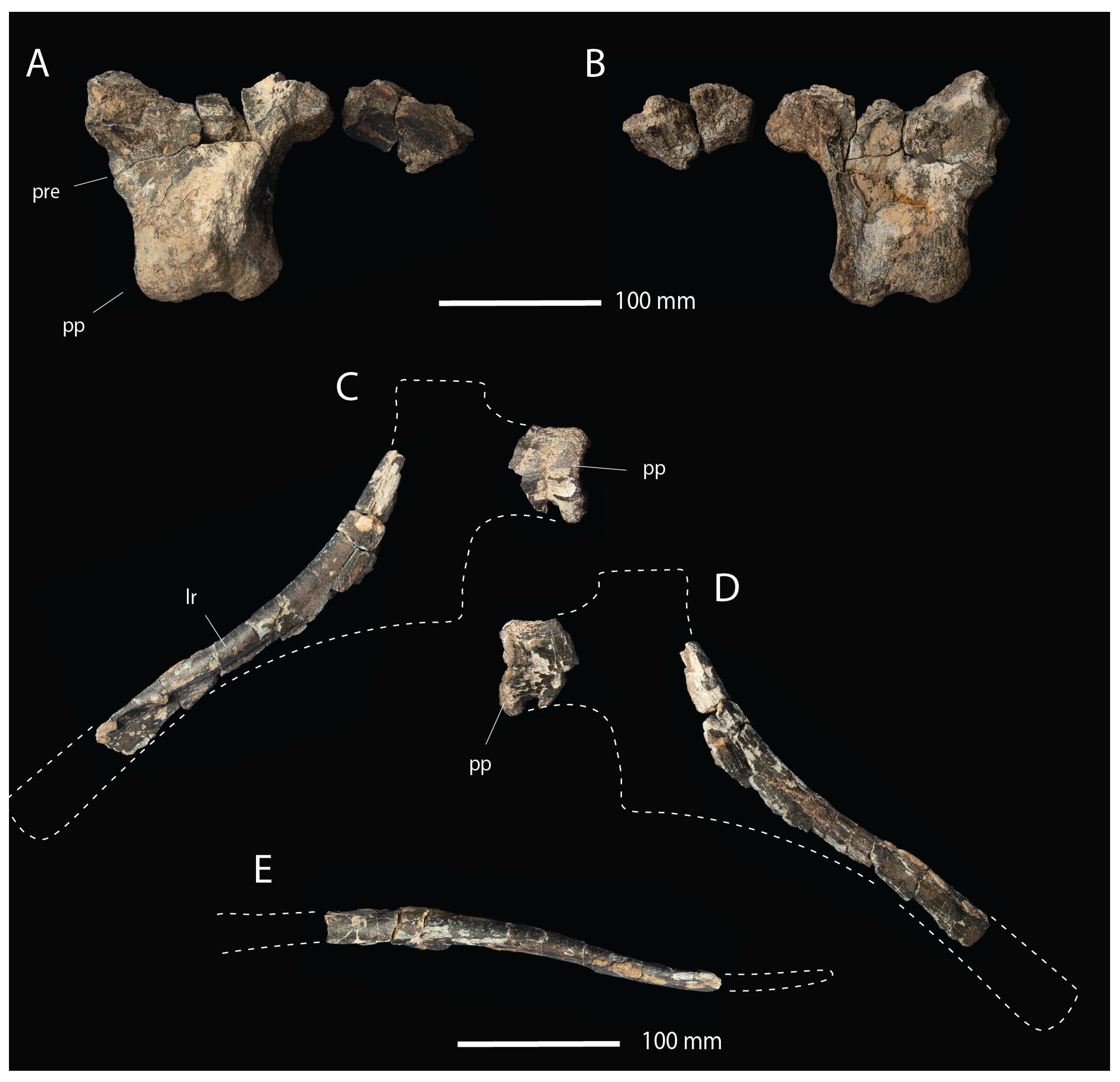
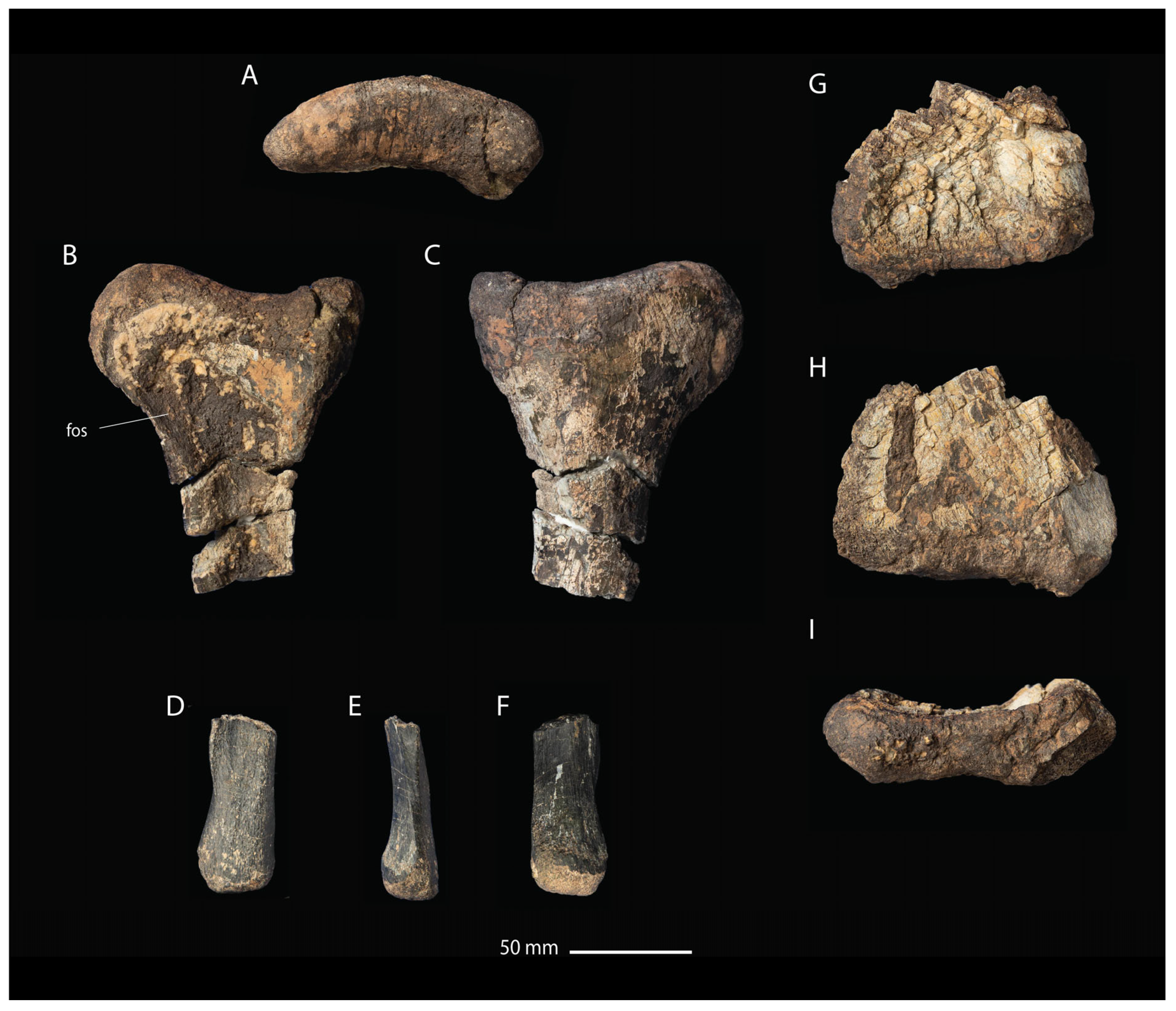
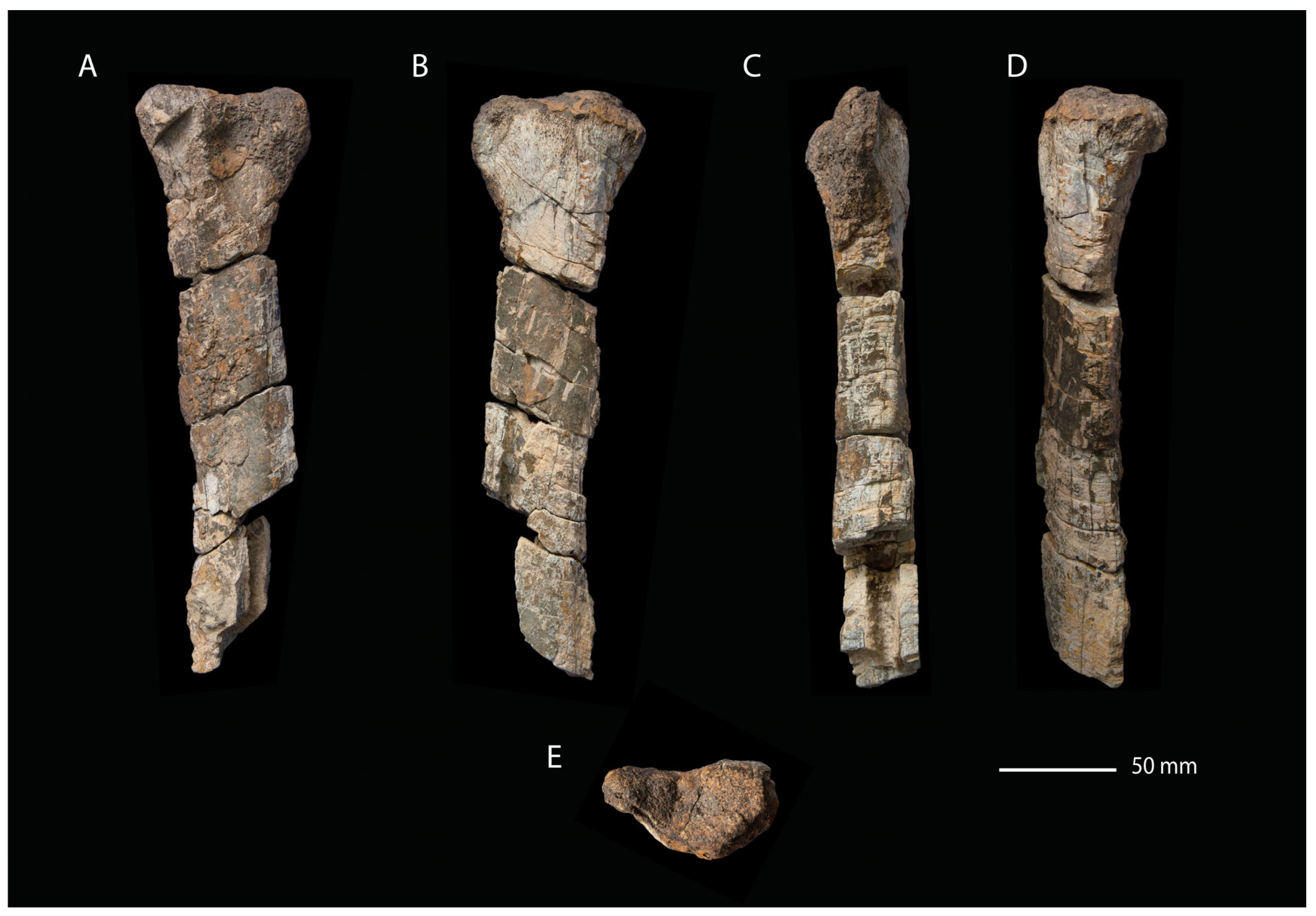
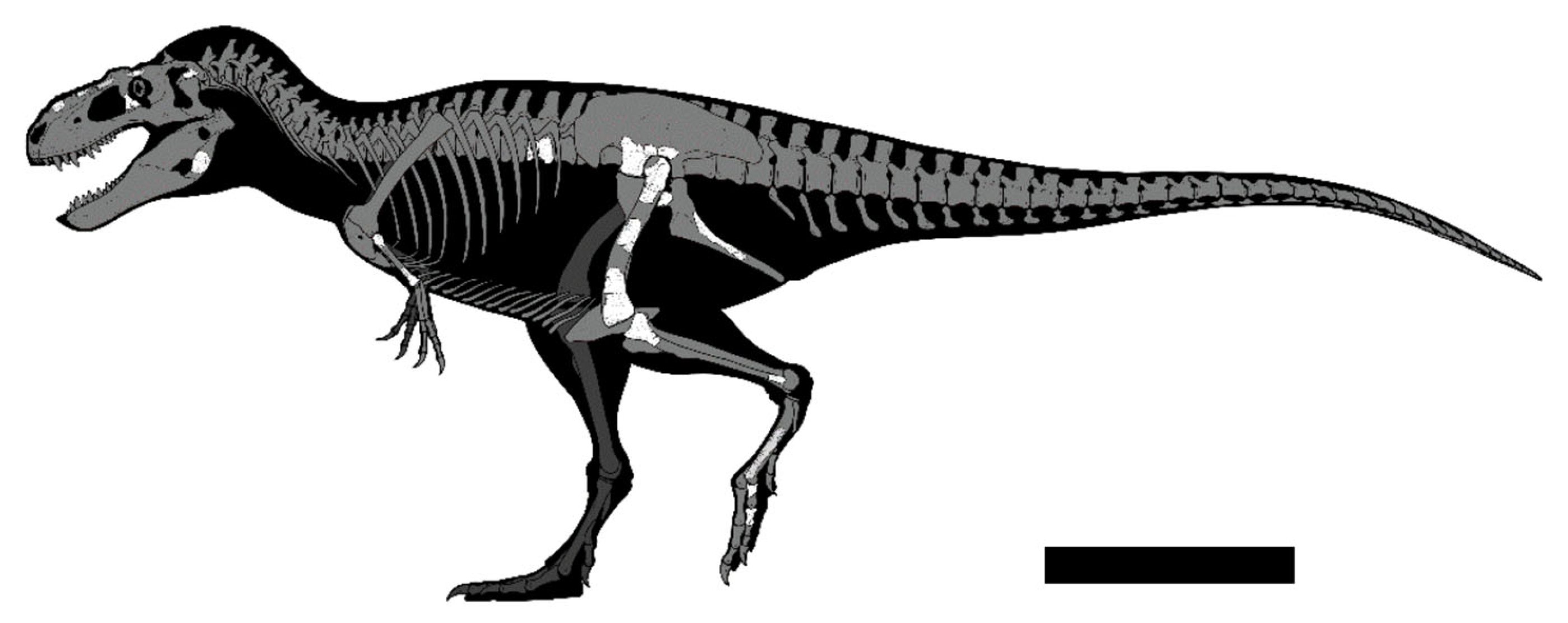
- Description





Phylogenetic Analysis
4. Discussion
4.1. Ontogenetic Status
4.2. Systematics
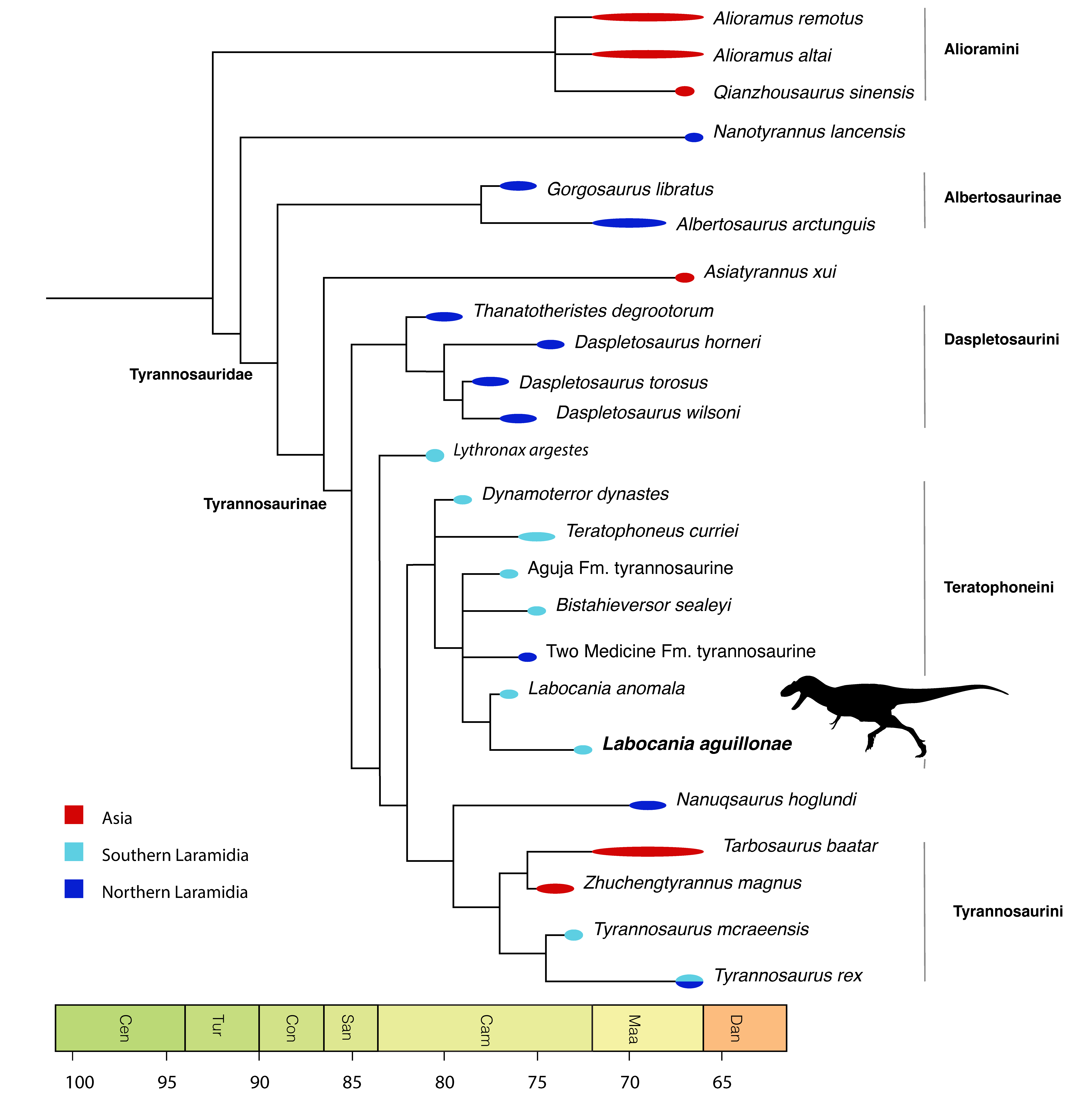
4.3. Diagnosis and Recognition of L. aguillonae
4.4. Affinities and Age of Labocania anomala
- Biogeography

5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Brusatte, S.L.; Norell, M.A.; Carr, T.D.; Erickson, G.M.; Hutchinson, J.R.; Balanoff, A.M.; Bever, G.S.; Choiniere, J.N.; Makovicky, P.J.; Xu, X. Tyrannosaur paleobiology: New research on ancient exemplar organisms. Science 2010, 329, 1481–1485. [Google Scholar] [CrossRef] [PubMed]
- Holtz, T.R. Tyrannosauroidea. In The Dinosauria, 2nd ed.; Weishampel, D.B., Dodson, P., Osmolska, H., Eds.; University of California Press: Berkeley, CA, USA, 2004; pp. 111–136. [Google Scholar]
- Weishampel, D.B.; Barrett, P.M.; Coria, R.A.; Le Loeuff, J.; Xu, X.; Zhao, X.; Sahni, A.; Gomani, E.; Noto, C.R. Dinosaur Distribution. In The Dinosauria, 2nd ed.; Weishampel, D.B., Dodson, P., Osmolska, H., Eds.; University of California Press: Berkeley, CA, USA, 2004; pp. 517–606. [Google Scholar]
- Dalman, S.G.; Loewen, M.A.; Pyron, R.A.; Jasinski, S.E.; Malinzak, D.E.; Lucas, S.G.; Fiorillo, A.R.; Currie, P.J.; Longrich, N.R. A giant tyrannosaur from the Campanian–Maastrichtian of southern North America and the evolution of tyrannosaurid gigantism. Sci. Rep. 2024, 14, 22124. [Google Scholar] [CrossRef] [PubMed]
- Brusatte, S.L.; Carr, T.D. The phylogeny and evolutionary history of tyrannosauroid dinosaurs. Sci. Rep. 2016, 6, 20252. [Google Scholar] [CrossRef] [PubMed]
- Zanno, L.E.; Tucker, R.T.; Canoville, A.; Avrahami, H.M.; Gates, T.A.; Makovicky, P.J. Diminutive fleet-footed tyrannosauroid narrows the 70-million-year gap in the North American fossil record. Commun. Biol. 2019, 2, 64. [Google Scholar] [CrossRef] [PubMed]
- Nesbitt, S.J.; Denton, R.K., Jr.; Loewen, M.A.; Brusatte, S.L.; Smith, N.D.; Turner, A.H.; Kirkland, J.I.; McDonald, A.T.; Wolfe, D.G. A mid-Cretaceous tyrannosauroid and the origin of North American end-Cretaceous dinosaur assemblages. Nat. Ecol. Evol. 2019, 3, 892–899. [Google Scholar] [CrossRef]
- Bakker, R.; Williams, M.; Currie, P.J. Nanotyrannus, a new genus of pygmy tyrannosaur, from the latest Cretaceous of Montana. Hunteria 1988, 1, 1–30. [Google Scholar]
- Longrich, N.R.; Saitta, E.T. Taxonomic status of Nanotyrannus lancensis (Dinosauria: Tyrannosauroidea)—A distinct taxon of small-bodied tyrannosaur. Foss. Stud. 2024, 2, 1–65. [Google Scholar] [CrossRef]
- Currie, P.J. Cranial anatomy of tyrannosaurid dinosaurs from the Late Cretaceous of Alberta, Canada. Acta Palaeontol. Pol. 2003, 48, 191–226. [Google Scholar]
- Brochu, C.A. Osteology of Tyrannosaurus rex: Insights from a nearly complete skeleton and high-resolution computed tomographic analysis of the skull. J. Vertebr. Paleontol. Mem. 2003, 7, 1–138. [Google Scholar] [CrossRef]
- Hutchinson, J.R.; Bates, K.T.; Molnar, J.; Allen, V.; Makovicky, P.J. A computational analysis of limb and body dimensions in Tyrannosaurus rex with implications for locomotion, ontogeny, and growth. PLoS ONE 2011, 6, e26037. [Google Scholar] [CrossRef]
- Gaston, K.J.; Blackburn, T.M. Conservation implications of geographic range size-body size relationships. Conserv. Biol. 1996, 10, 638–646. [Google Scholar] [CrossRef]
- Nowak, R.M. Walker’s Mammals of the World, 6th ed.; JHU Press: Baltimore, MD, USA, 1999. [Google Scholar]
- Currie, P.J. Theropods, including birds. In Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed; Currie, P.J., Koppelhus, E.B., Eds.; Indiana University Press: Bloomington, IN, USA, 2005; pp. 367–397. [Google Scholar]
- Warshaw, E.A.; Fowler, D.W. A transitional species of Daspletosaurus Russell, 1970 from the Judith River Formation of eastern Montana. PeerJ 2022, 10, e14461. [Google Scholar] [CrossRef] [PubMed]
- Carr, T.D.; Varricchio, D.J.; Sedlmayr, J.C.; Roberts, E.M.; Moore, J.R. A new tyrannosaur with evidence for anagenesis and crocodile-like facial sensory system. Sci. Rep. 2017, 7, srep44942. [Google Scholar] [CrossRef] [PubMed]
- Loewen, M.A.; Irmis, R.B.; Sertich, J.J.; Currie, P.J.; Sampson, S.D. Tyrant dinosaur evolution tracks the rise and fall of Late Cretaceous oceans. PLoS ONE 2013, 8, e79420. [Google Scholar] [CrossRef] [PubMed]
- Carr, T.D.; Williamson, T.E.; Britt, B.B.; Stadtman, K. Evidence for high taxonomic and morphologic tyrannosauroid diversity in the Late Cretaceous (Late Campanian) of the American Southwest and a new short-skulled tyrannosaurid from the Kaiparowits Formation of Utah. Naturwissenschaften 2011, 98, 241–246. [Google Scholar] [CrossRef]
- Carr, T.D.; Williamson, T.E. Bistahieversor sealeyi, gen. et sp. nov., a new tyrannosauroid from New Mexico and the origin of deep snouts in Tyrannosauroidea. J. Vertebr. Paleontol. 2010, 30, 1–16. [Google Scholar] [CrossRef]
- Lambe, L.M. On a new genus and species of carnivorous dinosaur from the Belly River Formation of Alberta, with a description of the skull of Stephanosaurus marginatus from the same horizon. Ott. Nat. 1914, 28, 13–20. [Google Scholar]
- Osborn, H.F. Tyrannosaurus and other Cretaceous carnivorous dinosaurs. Bull. Am. Mus. Nat. Hist. 1905, 35, 733–771. [Google Scholar]
- Rivera-Sylva, H.E.; Frey, E.; Palomino-Sánchez, F.J.; Guzmán-Gutiérrez, J.R.; Ortiz-Mendieta, J.A. Preliminary report on a Late Cretaceous vertebrate fossil assemblage in northwestern Coahuila, Mexico. Boletín Soc. Geológica Mex. 2009, 61, 239–244. [Google Scholar] [CrossRef]
- Torres-Rodríguez, E.; Montellano-Ballesteros, M.; Hernández-Rivera, R.; Benammi, M. Dientes de terópodos del Cretácico Superior del estado de Coahuila, México. Rev. Mex. Cienc. Geol. 2010, 27, 72–83. [Google Scholar]
- Lucas, S.; González-León, C. Dinosaurios del Cretácico Tardío del Grupo Cabullona, Sonora. Geol. Noroeste 1996, 1, 20–25. [Google Scholar]
- Peecook, B.R.; Wilson, J.A.; Hernández-Rivera, R.; Montellano-Ballesteros, M.; Wilson, G.P. First tyrannosaurid remains from the Upper Cretaceous “El Gallo” Formation of Baja California, México. Acta Palaeontol. Pol. 2014, 59, 71–80. [Google Scholar] [CrossRef]
- Molnar, R.E. A distinctive theropod dinosaur from the Upper Cretaceous of Baja California (Mexico). J. Paleontol. 1974, 48, 1009–1017. [Google Scholar]
- Rivera-Sylva, H.E.; Carpenter, K. Mexican saurischian dinosaurs. In Dinosaurs and Other Reptiles from the Mesozoic of Mexico; Indiana University Press: Bloomigton, IN, USA, 2014; pp. 143–155. [Google Scholar]
- Aguillón-Martínez, M. Fossil Vertebrates from the Cerro del Pueblo Formation, Coahuila, Mexico, and the Distribution of Late Campanian (Cretaceous) Terrestrial Vertebrate Faunas. Master’s Thesis, Southern Methodist University, Dallas, TX, USA, 2010; 148p. [Google Scholar]
- Rivera-Sylva, H.E.; Frey, E.; Stinnesbeck, W.; Amezcua Torres, N.; Flores Huerta, D. Terrestrial vertebrate paleocommunities from the Cerro del Pueblo Formation (Late Cretaceous; Late Campanian) at Las Águilas, Coahuila, Mexico. Palaeovertebrata 2019, 42, e1. [Google Scholar] [CrossRef]
- Vega-Vera, F.J.; Salazar, L.M.M.; Hernández, E.M. Contribución al conocimiento de la estratigrafía del Grupo Difunta (Cretácico Superior-Terciario) en el noreste de México. Rev. Mex. Cienc. Geol. 1989, 8, 179–187. [Google Scholar]
- Soegaard, K.; Ye, H.; Halik, N.; Daniels, A.T.; Arney, J.; Garrick, S. Stratigraphic evolution of latest Cretaceous to early Tertiary Difunta foreland basin in northeast Mexico: Influence of salt withdrawal on tectonically induced subsidence by the Sierra Madre Oriental fold and thrust belt. In The Circum-Gulf of Mexico and the Caribbean; Hydrocarbon Habitats, Basin Formation, and Plate Tectonics; Bartolini, C., Buffler, R.T., Blickwede, J.F., Eds.; American Association of Petroleum Geologists: Tulsa, OK, USA, 2003; pp. 364–394. [Google Scholar]
- Ifrim, C.; Stinnesbeck, W.; Garza, R.R.; Ventura, J.F. Hemipelagic cephalopods from the Maastrichtian (Late Cretaceous) Parras Basin at La Parra, Coahuila, Mexico, and their implications for the correlation of the lower Difunta Group. J. S. Am. Earth Sci. 2010, 29, 597–618. [Google Scholar] [CrossRef]
- de Jesús, C.R.D.; Eberth, D.A.; de la Rosa, R.A.R.; Lerbekmo, J.F.; Brinkman, D.B.; Sampson, S.D. Cerro del Pueblo Fm (Difunta Group, Upper Cretaceous), Parras Basin, southern Coahuila, Mexico: Reference sections, age, and correlation. Rev. Mex. Cienc. Geol. 2004, 21, 335–352. [Google Scholar]
- Serrano-Brañas, C.I.; Espinosa-Chávez, B.; Maccracken, S.A.; Torres-Rodríguez, E. The Cerro del Pueblo Formation, Unlocking the Environmental Data of an Extraordinary Ancient Ecosystem from Mexico. In Past Environments of Mexico: Unveiling the Past Environments of a Megadiverse Country Through Its Fossil Record; Springer: Berlin/Heidelberg, Germany, 2024; pp. 405–426. [Google Scholar]
- Vogt, M.; Stinnesbeck, W.; Zell, P.; Kober, B.; Kontny, J.; Herzer, N.; Frey, E.; Rivera-Sylva, H.E.; Gutierrez, J.M.P.; Amezcua, N. Age and depositional environment of the “dinosaur graveyard” at Las Águilas, southern Coahuila, NE Mexico. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2016, 441, 758–769. [Google Scholar] [CrossRef]
- Vivas González, R. Paleoecología de Dinosaurios Hadrosauridos (Ornithischia: Ornithopoda) de la Formación Cerro del Pueblo (Cretácico Tardío: Campaniano), Coahuila, México. Master’s Thesis, Universidad Autónoma de Nuevo León, San Nicolás de los Garza, Mexico, 2013. [Google Scholar]
- Swofford, D.L. PAUP*. Phylogenetic Analysis Using Parsimony (*and Other Methods); 4.0b10; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Yun, C.Y. Probable juvenile frontal of Daspletosaurus horneri (Dinosauria: Theropoda) from the Two Medicine Formation of Montana, with implications for tyrannosaurid ontogeny. Acta Palaeontol. Rom. 2023, 19, 3–11. [Google Scholar] [CrossRef]
- Owen, R. Report on British fossil reptiles. Part II. Rep. Br. Assoc. Adv. Sci. 1842, 11, 60–204. [Google Scholar]
- Marsh, O.C. Classification of the Dinosauria. Am. J. Sci. 1881, 23, 81–86. [Google Scholar] [CrossRef]
- Huene, F.v. The dinosaurs not a natural order. Am. J. Sci. 1914, 38, 145–146. [Google Scholar] [CrossRef]
- Osborn, H.F. Tyrannosaurus, Upper Cretaceous carnivorous dinosaur (second communication). Bull. Am. Mus. Nat. Hist. 1906, 22, 281–296. [Google Scholar]
- Scherer, C.R.; Voiculescu-Holvad, C. Reanalysis of a dataset refutes claims of anagenesis within Tyrannosaurus-line tyrannosaurines (Theropoda, Tyrannosauridae). Cretac. Res. 2024, 155, 105780. [Google Scholar] [CrossRef]
- Tumarkin-Deratzian, A.R.; Vann, D.R.; Dodson, P. Bone surface texture as an ontogenetic indicator in long bones of the Canada goose Branta canadensis (Anseriformes: Anatidae). Zool. J. Linn. Soc. 2006, 148, 133–168. [Google Scholar] [CrossRef]
- Brochu, C.A. Closure of neurocentral sutures during crocodylian ontogeny: Implication for maturity assessment in fossil archosaurs. J. Vertebr. Paleontol. 1996, 16, 49–62. [Google Scholar] [CrossRef]
- Fiorillo, A.R.; Tykoski, R.S. A diminutive new tyrannosaur from the top of the world. PLoS ONE 2014, 9, e91287. [Google Scholar] [CrossRef]
- Yun, C.-G.; Peters, G.F.; Currie, P.J. Allometric growth in the frontals of the Mongolian theropod dinosaur Tarbosaurus bataar. Acta Palaeontol. Pol. 2022, 67, 601–615. [Google Scholar] [CrossRef]
- McDonald, A.T.; Wolfe, D.G.; Dooley, A.C., Jr. A new tyrannosaurid (Dinosauria: Theropoda) from the Upper Cretaceous Menefee Formation of New Mexico. PeerJ 2018, 6, e5749. [Google Scholar] [CrossRef]
- Lehman, T.M.; Wick, S.L. Tyrannosauroid dinosaurs from the Aguja Formation (Upper Cretaceous) of Big Bend National Park, Texas. Earth Environ. Sci. Trans. R. Soc. Edinb. 2012, 103, 471–485. [Google Scholar] [CrossRef]
- Perry, Z.R. A Reinterpretation of Nanuqsaurus hoglundi (Tyrannosauridae) from the Late Cretaceous Prince Creek Formation, Northern Alaska. Master’s Thesis, University of Alaska Fairbanks, Fairbanks, AK, USA, 2023. [Google Scholar]
- Voris, J.T.; Therrien, F.; Zelenitsky, D.K.; Brown, C.M. A new tyrannosaurine (Theropoda: Tyrannosauridae) from the Campanian Foremost Formation of Alberta, Canada, provides insight into the evolution and biogeography of tyrannosaurids. Cretac. Res. 2020, 110, 104388. [Google Scholar] [CrossRef]
- Yun, C.-g. Frontal bone anatomy of Teratophoneus curriei (Theropoda: Tyrannosauridae) From the Upper Cretaceous Kaipariwits Formation of Utah. Acta Palaeontol. Rom. 2022, 18, 51–64. [Google Scholar] [CrossRef]
- Evans, D.C.; Larson, D.W.; Cullen, T.M.; Sullivan, R.M. “Saurornitholestes” robustus is a troodontid (Dinosauria: Theropoda). Can. J. Earth Sci. 2014, 51, 730–734. [Google Scholar] [CrossRef]
- Hurum, J.H.; Sabath, K. Giant theropod dinosaurs from Asia and North America: Skulls of Tarbosaurus bataar and Tyrannosaurus rex compared. Acta Palaeontol. Pol. 2003, 48, 161–190. [Google Scholar]
- Brusatte, S.L.; Carr, T.D.; Norell, M.A. The osteology of Alioramus, a gracile and long-snouted tyrannosaurid (Dinosauria: Theropoda) from the Late Cretaceous of Mongolia. Bull. Am. Mus. Nat. Hist. 2012, 2012, 1–197. [Google Scholar] [CrossRef]
- Carr, T.D. Significant geographic range extension for the sympatric tyrannosaurids Albertosaurus libratus and Daspletosaurus torosus from the Judith River Formation (Late Campanian) of Northern Montana. In Proceedings of the Society of Vertebrate Paleontology 2018 Annual Meeting, Albuquerque, NM, USA, 17–20 October 2018; p. 102. [Google Scholar]
- Mallon, J.C.; Bura, J.R.; Schumann, D.; Currie, P.J. A problematic tyrannosaurid (Dinosauria: Theropoda) skeleton and its implications for tyrannosaurid diversity in the Horseshoe Canyon Formation (Upper Cretaceous) of Alberta. Anat. Rec. 2020, 303, 673–690. [Google Scholar] [CrossRef]
- Paul, G.S. The Princeton Field Guide to Dinosaurs, 2nd ed.; Princeton University Press: Princeton, NJ, USA, 2016. [Google Scholar]
- Longrich, N.R. Small theropod teeth from the Lance Formation of Wyoming. In The Unique Role of Vertebrate Microfossil Assemblages in Paleoecology and Paleobiology; Sankey, J.T., Baszio, S., Eds.; Indiana University Press: Bloomington, IN, USA, 2008; pp. 135–158. [Google Scholar]
- Larson, P. The case for Nanotyrannus. In Tyrannosaurid Paleobiology; Parrish, J.M., Molnar, R.E., Currie, P.J., Koppelhus, E.B., Eds.; Indiana University Press: Bloomington, IN, USA, 2013; pp. 15–53. [Google Scholar]
- Tumarkin-Deratzian, A.R. Evaluation of long bone surface textures as ontogenetic in centrosaurine ceratopsids. Anat. Rec. 2009, 292, 1485–1500. [Google Scholar] [CrossRef]
- Ramezani, J.; Beveridge, T.L.; Rogers, R.R.; Eberth, D.A.; Roberts, E.M. Calibrating the zenith of dinosaur diversity in the Campanian of the Western Interior Basin by CA-ID-TIMS U–Pb geochronology. Sci. Rep. 2022, 12, 16026. [Google Scholar] [CrossRef]
- Mallon, J.C.; Evans, D.C.; Ryan, M.J.; Anderson, J.S. Megaherbivorous dinosaur turnover in the Dinosaur Park Formation (upper Campanian) of Alberta, Canada. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2012, 350–352, 124–138. [Google Scholar] [CrossRef]
- Paul, G.S.; Persons, W.S.; Van Raalte, J. The Tyrant Lizard King, Queen and Emperor: Multiple Lines of Morphological and Stratigraphic Evidence Support Subtle Evolution and Probable Speciation within the North American Genus Tyrannosaurus. Evol. Biol. 2022, 49, 156–179. [Google Scholar] [CrossRef]
- Carr, T.D.; Napoli, J.G.; Brusatte, S.L.; Holtz, T.R.; Hone, D.W.; Williamson, T.E.; Zanno, L.E. Insufficient Evidence for Multiple Species of Tyrannosaurus in the Latest Cretaceous of North America: A Comment on “The Tyrant Lizard King, Queen and Emperor: Multiple Lines of Morphological and Stratigraphic Evidence Support Subtle Evolution and Probable Speciation Within the North American Genus Tyrannosaurus”. Evol. Biol. 2022, 49, 327–341. [Google Scholar]
- Cau, A. A Unified Framework for Predatory Dinosaur Macroevolution. Boll. Soc. Paleontol. Ital. 2024, 63, 2. [Google Scholar]
- Kane, I.A.; Hodgson, D.M.; Hubbard, S.M.; McArthur, A.D.; Poyatos-Moré, M.; Soutter, E.L.; Flint, S.S.; Matthews, W. Deep-water Tectono-Stratigraphy at a Plate Boundary Constrained by Large N-Detrital Zircon and Micropaleontological Approaches: Peninsular Ranges Forearc, Baja California, Mexico. Sediment. Rec. 2022, 20, 37652. [Google Scholar] [CrossRef]
- Barrera-Guevara, D.; Espinosa Chávez, B.; Serrano Brañas, C.I.; de León Dávila, C.; Posada Martinez, D.; Freedman Fowler, E.; Fowler, D. Stratigraphic Reassessment of the Mexican Chasmosaurine Coahuilaceratops magnacuerna as the First Diagnostic Dinosaur Remains from the Cerro Huerta Formation (Lower Maastrichtian) Supporting the Southern Origin of the Triceratopsini. Diversity 2024, 16, 390. [Google Scholar] [CrossRef]
- Lawson, D.A. Tyrannosaurus and Torosaurus, Maestrichtian dinosaurs from Trans-Pecos, Texas. J. Paleontol. 1976, 50, 158–164. [Google Scholar]
- Russell, D.A. Tyrannosaurs from the Late Cretaceous of Western Canada. Natl. Mus. Nat. Sci. Publ. Palaeontol. 1970, 1, 1–34. [Google Scholar]
- Eberth, D.A.; Kamo, S.L. High-precision U–PB CA–ID–TIMS dating and chronostratigraphy of the dinosaur-rich Horseshoe Canyon Formation (Upper Cretaceous, Campanian–Maastrichtian), Red Deer River Valley, Alberta, Canada. Can. J. Earth Sci. 2020, 57, 1220–1237. [Google Scholar] [CrossRef]
- Lehman, T.M. Late Campanian dinosaur biogeography in the Western Interior of North America. In Dinofest International, Proceedings of a Symposium Sponsored by Arizona State University, Special Publication, Academy of Natural Sciences; Wolberg, D.A., Stump, E., Eds.; Academy of Natural Sciences: Philidelphia, PA, USA, 1997; pp. 223–240. [Google Scholar]
- Lehman, T.M. Late Cretaceous Dinosaur Provinciality. In Mesozoic Vertebrate Life; Tanke, D.H., Carpenter, K., Eds.; Indiana University Press: Bloomington, IN, USA, 2001; pp. 310–328. [Google Scholar]
- Gates, T.; Sampson, S.; Zanno, L.; Roberts, E.; Eaton, J.; Nydam, R.; Hutchison, J.; Smith, J.; Loewen, M.; Getty, M. Biogeography of terrestrial and freshwater vertebrates from the Late Cretaceous (Campanian) Western Interior of North America. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2010, 291, 371–387. [Google Scholar] [CrossRef]
- Longrich, N.R. Titanoceratops ouranos, a giant horned dinosaur from the Late Campanian of New Mexico. Cretac. Res. 2011, 32, 264–276. [Google Scholar] [CrossRef]
- Sampson, S.D.; Loewen, M.A.; Farke, A.A.; Roberts, E.M.; Forster, C.A.; Smith, J.A.; Titus, A.L. New horned dinosaurs from Utah provide evidence for intracontinental dinosaur endemism. PLoS ONE 2010, 5, e12292. [Google Scholar] [CrossRef]
- Loewen, M.A.; Sampson, S.D.; Lund, E.K.; Farke, A.A.; Aguillón-Martínez, M.C.; de Leon, C.A.; Rodríguez-de la Rosa, R.A.; Getty, M.A.; Eberth, D.A. Horned dinosaurs (Ornithischia: Ceratopsidae) from the Upper Cretaceous (Campanian) Cerro del Pueblo Formation, Coahila, Mexico. In New Perspectives on Horned Dinosaurs: The Royal Tyrrell Museum Ceratopsian Symposium; Ryan, M.J., Chinnery, B.J., Eberth, D.A., Eds.; Indiana University Press: Bloomington, IN, USA, 2010; pp. 99–116. [Google Scholar]
- Dalman, S.G.; Lucas, S.G.; Jasinski, S.E.; Longrich, N.R. Sierraceratops turneri, a new chasmosaurine ceratopsid from the Hall Lake Formation (Upper Cretaceous) of south-central New Mexico. Cretac. Res. 2022, 130, 105034. [Google Scholar] [CrossRef]
- Gates, T.A.; Sampson, S.D.; Delgado de Jesús, C.R.; Zanno, L.E.; Eberth, D.; Hernandez-Rivera, R.; Aguillón Martínez, M.C.; Kirkland, J.I. Velafrons coahuilensis, a new lambeosaurine hadrosaurid (Dinosauria: Ornithopoda) from the Late Campanian Cerro Del Pueblo Formation, Coahila, Mexico. J. Vertebr. Paleontol. 2007, 27, 917–930. [Google Scholar] [CrossRef]
- Gates, T.A.; Sampson, S. A new species of Gryposaurus (Dinosauria: Hadrosauridae) from the late Campanian Kaiparowits Formation, southern Utah, USA. Zool. J. Linn. Soc. 2007, 151, 351–376. [Google Scholar] [CrossRef]
- Velasco, A.A.R.; Aguilar, F.J.; Hernández-Rivera, R.; Maussán, J.L.G.; Rodríguez, M.L.; Alvarado-Ortega, J. Tlatolophus galorum, gen. et sp. nov., a parasaurolophini dinosaur from the upper Campanian of the Cerro del Pueblo Formation, Coahuila, northern Mexico. Cretac. Res. 2021, 126, 104884. [Google Scholar] [CrossRef]
- Parks, W.A. Parasaurolophus walkeri: A new genus and species of crested trachodont dinosaur. Univ. Tor. Stud. Geol. Ser. 1922, 13, 5–32. [Google Scholar]
- Gates, T.A.; Evans, D.C.; Sertich, J.J. Description and rediagnosis of the crested hadrosaurid (Ornithopoda) dinosaur Parasaurolophus cyrtocristatus on the basis of new cranial remains. PeerJ 2021, 9, e10669. [Google Scholar] [CrossRef]
- Lowi-Merri, T.M.; Evans, D.C. Cranial variation in Gryposaurus and biostratigraphy of hadrosaurines (Ornithischia: Hadrosauridae) from the Dinosaur Park Formation of Alberta, Canada. Can. J. Earth Sci. 2020, 57, 765–779. [Google Scholar] [CrossRef]
- Lehman, T.M. New data on the ceratopsian dinosaur Pentaceratops sternbergii Osborn from New Mexico. J. Paleontol. 1993, 67, 279–288. [Google Scholar] [CrossRef]
- Osborn, H.F. A new genus of Ceratopsia from New Mexico, Pentaceratops sternbergii. Am. Mus. Novit. 1923, 93, 3. [Google Scholar]
- Longrich, N.R. The horned dinosaurs Pentaceratops and Kosmoceratops from the upper Campanian of Alberta and implications for dinosaur biogeography. Cretac. Res. 2014, 51, 292–308. [Google Scholar] [CrossRef]
- Sampson, S.D. Two new horned dinosaurs from the Upper Cretaceous Two Medicine Formation of Montana; with a phylogenetic analysis of the Centrosaurinae (Ornithischia: Ceratopsidae). J. Vertebr. Paleontol. 1995, 15, 743–760. [Google Scholar] [CrossRef]
- Ryan, M.J.; Evans, D.C. Ornithischian Dinosaurs. In Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed; Currie, P.J., Koppelhus, E.B., Eds.; Indiana University Press: Bloomington, IN, USA, 2005; pp. 312–348. [Google Scholar]
- Fiorillo, A.R.; Tykoski, R.S. A new Maastrichtian species of the centrosaurine ceratopsid Pachyrhinosaurus from the North Slope of Alaska. Acta Palaeontol. Pol. 2012, 57, 561–573. [Google Scholar] [CrossRef]
- Sternberg, R.M. Pachyrhinosaurus canadensis, representing a new family of the Ceratopsia, from southern Alberta. Natl. Mus. Can. Bull. 1950, 118, 109–120. [Google Scholar]
- Sampson, S.D.; Lund, E.K.; Loewen, M.A.; Farke, A.A.; Clayton, K.E. A remarkable short-snouted horned dinosaur from the Late Cretaceous (late Campanian) of southern Laramidia. Proc. R. Soc. B 2013, 280, 20131186. [Google Scholar] [CrossRef] [PubMed]
- Dalman, S.G.; Lucas, S.G.; Jasinski, S.E.; Lichtig, A.J.; Dodson, P. The oldest centrosaurine: A new ceratopsid dinosaur (Dinosauria: Ceratopsidae) from the Allison Member of the Menefee Formation (Upper Cretaceous, early Campanian), northwestern New Mexico, USA. PalZ 2021, 95, 291–335. [Google Scholar] [CrossRef]
- Rivera-Sylva, H.E.; Hedrick, B.P.; Dodson, P. A Centrosaurine (Dinosauria: Ceratopsia) from the Aguja Formation (Late Campanian) of Northern Coahuila, Mexico. PLoS ONE 2016, 11, e0150529. [Google Scholar] [CrossRef]
- Lambe, L.M. The Cretaceous Theropodous Dinosaur Gorgosaurus. Can. Dep. Mines Geol. Surv. Mem. 1917, 100, 1–84. [Google Scholar]
- Currie, P.J.; Evans, D.C. Cranial anatomy of new specimens of Saurornitholestes langstoni (Dinosauria, Theropoda, Dromaeosauridae) from the Dinosaur Park Formation (Campanian) of Alberta. Anat. Rec. 2020, 303, 691–715. [Google Scholar] [CrossRef]
- Prieto-Márquez, A. Skeletal morphology of Kritosaurus navajovius (Dinosauria: Hadrosauridae) from the Late Cretaceous of the North American south-west, with an evaluation of the phylogenetic systematics and biogeography of Kritosaurini. J. Syst. Palaeontol. 2014, 12, 133–175. [Google Scholar] [CrossRef]
- Lehman, T.M.; Wick, S.L.; Wagner, J.R. Hadrosaurian dinosaurs from the Maastrichtian Javelina Formation, Big Bend National Park, Texas. J. Paleontol. 2016, 90, 333–356. [Google Scholar] [CrossRef]
- Sullivan, R.M.; Jasinski, S.E.; Guenther, M.F.; Lucas, S.G. The first lambeosaurin (Dinosauria, Hadrosauridae, Lambeosaurinae) from the Upper Cretaceous Ojo Alamo Formation (Naashoibito Member), San Juan Basin, New Mexico. New Mex. Mus. Nat. Hist. Sci. Bull. 2011, 53, 405–417. [Google Scholar]
- Price, G.D.; Twitchett, R.J.; Wheeley, J.R.; Buono, G. Isotopic evidence for long term warmth in the Mesozoic. Sci. Rep. 2013, 3, 1438. [Google Scholar] [CrossRef]
- Baghai, N.L. Classification and Analysis of Palynomorphs of the Aguja Formation (Campanian), Big Bend National Park, Brewster County, Texas. GCAGS Trans. 1994, 44, 63–70. [Google Scholar]
- Medus, J.; Almeida-Leñero, L. Des Normapolles du Crétacé supérieur de Piedras Negras (Coahuila, Mexique). Geobios 1982, 15, 225–229. [Google Scholar] [CrossRef]
- Estrada-Ruiz, E.; Martínez-Cabrera, H.I.; Callejas-Moreno, J.; Upchurch, G.R., Jr. Cretaceous tropical rainforests from northern Mexico and its relationship with south-central North American floras. Polibotánica 2013, 41–61. [Google Scholar]
- Wick, S.L. A new assemblage of fossil leaves and wood from The Aguja Formation (Upper Campanian) of West Texas: An interfluvial community in a volcanic landscape. Cretac. Res. 2023, 142, 105366. [Google Scholar] [CrossRef]
- Knowlton, F.H. Flora of the Fruitland and Kirtland Formations; US Government Printing Office: Washington, DC, USA, 1916.
- Wing, S.L.; Boucher, L.D. Ecological aspects of the Cretaceous flowering plant radiation. Annu. Rev. Earth Planet. Sci. 1998, 26, 379–421. [Google Scholar] [CrossRef]
- Miller, I.M.; Johnson, K.R.; Kline, D.E.; Nichols, D.J.; Barclay, R.S. A Late Campanian Flora from the Kaiparowits. In At the Top of the Grand Staircase: The Late Cretaceous of Southern Utah; Titus, A., Loewen, M.A., Eds.; Indiana University Press: Bloomington, IN, USA; Indianapolis, IN, USA, 2013; pp. 107–131. [Google Scholar]
- Koppelhus, E.B. Paleobotany. In Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed; Currie, P.J., Koppelhus, E.B., Eds.; Indiana University Press: Bloomington, IN, USA, 2005; pp. 131–138. [Google Scholar]
- Freeman, B.G.; Strimas-Mackey, M.; Miller, E.T. Interspecific competition limits bird species’ ranges in tropical mountains. Science 2022, 377, 416–420. [Google Scholar] [CrossRef]
- Sampson, S.D.; Loewen, M.A. Tyrannosaurus rex from the Upper Cretaceous (Maastrichtian) North Horn Formation of Utah: Biogeographic and paleoecologic implications. J. Vertebr. Paleontol. 2005, 25, 469–472. [Google Scholar] [CrossRef]
- Tokaryk, T.T.; Cumbaa, S.L.; Storer, J.E. Early Late Cretaceous birds from Saskatchewan, Canada: The oldest diverse avifauna known from North America. J. Vertebr. Paleontol. 1997, 17, 172–176. [Google Scholar] [CrossRef]
- Jasinski, S.E.; Sullivan, R.M.; Lucas, S.G. Taxonomic composition of the Alamo Wash local fauna from the Upper Cretaceous Ojo Alamo Formation (Naashoibito Member), San Juan Basin, New Mexico. New Mex. Mus. Nat. Hist. Sci. Bull. 2011, 53, 216–265. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Femur, maximum transverse diameter of shaft | 69.2 |
| Femur, maximum transverse diameter of distal end | 124.1 |
| Tibia, maximum anteroposterior width of proximal end | 93.6 * |
| Tibia, maximum transverse width of proximal end | 79.0 |
| Tibia, maximum transverse width of distal end | 115.0 |
| Fibula, maximum anteroposterior width of proximal end | 107.0 |
| Fibula, maximum mediolateral width of proximal end | 37.3 |
| Dorsal vertebra, maximum anteroposterior length of centrum | 90.0 |
| Dorsal vertebra, maximum dorsoventral height of anterior face | 88.7 |
| Dorsal vertebra, maximum width of anterior face | 71.7 * |
| Dorsal vertebra, maximum dorsoventral height of posterior face | 74.8 |
| Dorsal vertebra, maximum width of posterior face | 74.9 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rivera-Sylva, H.E.; Longrich, N.R. A New Tyrant Dinosaur from the Late Campanian of Mexico Reveals a Tribe of Southern Tyrannosaurs. Foss. Stud. 2024, 2, 245-272. https://doi.org/10.3390/fossils2040012
Rivera-Sylva HE, Longrich NR. A New Tyrant Dinosaur from the Late Campanian of Mexico Reveals a Tribe of Southern Tyrannosaurs. Fossil Studies. 2024; 2(4):245-272. https://doi.org/10.3390/fossils2040012
Chicago/Turabian StyleRivera-Sylva, Héctor E., and Nicholas R. Longrich. 2024. "A New Tyrant Dinosaur from the Late Campanian of Mexico Reveals a Tribe of Southern Tyrannosaurs" Fossil Studies 2, no. 4: 245-272. https://doi.org/10.3390/fossils2040012
APA StyleRivera-Sylva, H. E., & Longrich, N. R. (2024). A New Tyrant Dinosaur from the Late Campanian of Mexico Reveals a Tribe of Southern Tyrannosaurs. Fossil Studies, 2(4), 245-272. https://doi.org/10.3390/fossils2040012