Abstract
Malaria remains a leading cause of morbidity and mortality worldwide, with significant efforts directed towards developing effective vaccines to curb its impact. The One Health concept, recognizing the interconnectedness of human, animal, and environmental health, offers a comprehensive approach to understanding and combating this disease. This review examines the development of malaria vaccine through the lens of the One Health framework, highlighting the integration of human, vector, and environmental factors in the fight against malaria. We discuss the current landscape of malaria vaccine development, including the deployment of the RTS,S/AS01 vaccine and the progress of other candidates such as the R21/Matrix-M and PfSPZ vaccine. The challenges posed by the complex lifecycle of the malaria parasite, its genetic diversity, and the environmental factors influencing transmission are explored. This review also discusses emerging technologies and innovations that could enhance vaccine efficacy and delivery. Additionally, we consider ethical, social, and economic factors critical to the successful implementation of vaccination programs. In concluding, this review underscores the importance of adopting a One Health approach to malaria vaccine development, advocating for integrated efforts to address the multifaceted challenges of malaria control and eradication.
1. Introduction
Malaria persists as a formidable global health challenge, inflicting substantial morbidity and mortality, particularly in Sub-Saharan Africa and parts of Southeast Asia [1,2]. Despite concerted global efforts to control the disease, including the use of insecticide-treated bed nets (ITNs), antimalarial drugs, and vector control strategies, malaria caused an estimated 229 million cases and 409,000 deaths worldwide in 2019 [3]. According to the Global Burden of Disease Study 2019, the age-standardized incidence rate (ASR) of malaria globally decreased by an average of 0.80% annually from 1990 to 2019. However, a slight increase in the ASR was noted from 2015 to 2019, particularly in high-middle, middle, and low-middle sociodemographic index (SDI) regions. Central, Western, and Eastern Sub-Saharan Africa have consistently had the highest ASRs since 1990, with the incidence rate of children under five being the highest among all age groups [4]. In 2019, Sub-Saharan Africa accounted for 95% of all malaria cases and 96% of malaria deaths globally, with children under five comprising around 80% of these deaths. Furthermore, in Southeast Asia, specifically hypoendemic and mesoendemic areas, nearly one billion people are exposed to the risk of malaria [5]. The persistent high prevalence of malaria in those regions underscores the urgent need for effective prevention and control measures, particularly targeted at vulnerable populations such as young children and travelers to endemic areas [6]. The development of a vaccine against malaria represents a crucial frontier in the global fight against this disease. The quest for a malaria vaccine has been a long and complex journey, beginning in earnest in the 1960s with research focused on whole-organism vaccines using irradiated parasites [7]. This was followed by efforts targeting subunit vaccines in the 1980s, culminating in the development of the RTS,S/AS01 vaccine, the first to demonstrate significant efficacy in Phase III clinical trials [8]. Despite this progress, the RTS,S/AS01 vaccine’s partial protection and waning efficacy over time underscore the need for continued research and development of more effective malaria vaccines.
The One Health perspective is an interdisciplinary approach that recognizes the interconnectedness of humans, animals, and environmental health. It emphasizes the need for collaborative efforts across various sectors to effectively manage and prevent diseases that affect humans, animals, and ecosystems. This perspective has been particularly relevant in understanding and controlling zoonotic diseases, which are diseases transmitted from animals to humans. Although primarily associated with zoonotic diseases, the One Health approach is also crucial for nonzoonotic human malaria control. It provides a framework to integrate various control measures, such as vector management and environmental interventions, to reduce malaria transmission. Malaria transmission dynamics are influenced by a complex interplay of factors, including human behavior, vector ecology, parasite biology, and environmental changes, such as climate change and land use patterns [9]. For nonzoonotic human malaria, One Health framework highlights the importance of controlling the mosquito vectors and addressing environmental factors that contribute to their breeding and survival. Asymptomatic individuals, who harbor the malaria parasite without showing symptoms, play a critical role in malaria transmission. They serve as hidden reservoirs, continuously infecting mosquitoes that can then transmit the disease to other humans. Addressing this silent reservoir is vital for effective malaria control and requires widespread diagnostic testing and treatment strategies that include these asymptomatic carriers [10]. Vector control remains a cornerstone of malaria prevention. Traditional methods such as ITNs and indoor residual spraying (IRS) have been effective but face challenges like insecticide resistance. Newer strategies, including biological control and genetic manipulation of mosquitoes, are being explored to sustain and enhance vector control efforts [11]. Integrated vector management (IVM), which combines multiple control methods tailored to local conditions, is advocated to manage resistance and achieve more sustainable control of malaria vectors [12]. Recognizing these interdependencies, the One Health approach facilitates a comprehensive understanding of the disease ecology, potentially unveiling novel targets for vaccine development and strategies for vaccine deployment [13]. The rationale for applying the One Health concept to malaria vaccine development lies in the opportunity to leverage interdisciplinary research and collaborative efforts to address the multifaceted challenges posed by malaria. By integrating knowledge from fields such as veterinary medicine, environmental science, epidemiology, and public health, the One Health approach can inform the design of vaccines that are not only effective in diverse human populations but also sustainable in the face of changing environmental conditions and vector populations.
Despite its potential advantages, the application of the One Health framework to malaria vaccination has not been extensively discussed by previous researchers [14,15]. There is currently a notable gap in the literature concerning the integration of the One Health framework into malaria vaccination programs. Specifically, the current literature provides limited insights into how this integrated approach could be leveraged to optimize malaria vaccine deployment and outcomes. Thus, the aim of this review is to explore the current updates on malaria vaccination program from a One Health perspective. We examine the updates on malaria vaccine research, discuss the integration of One Health principles in addressing the challenges of vaccine development, and highlight innovative approaches that promise to enhance the efficacy and deployment of malaria vaccines. Through this lens, we underscore the potential of a One Health approach to catalyze breakthroughs in malaria vaccination, ultimately contributing to the global effort to eradicate this devastating disease.
2. The Malaria Parasite and Its Lifecycle: A One Health View
Malaria, a life-threatening disease caused by parasites of the genus Plasmodium, remains a significant global health challenge. The most prevalent and virulent species is Plasmodium falciparum, responsible for the majority of malaria-related deaths, particularly in Africa. Plasmodium vivax, though less deadly, contributes substantially to global malaria morbidity, with a wider geographical distribution, including parts of Asia and Latin America. Plasmodium malariae and Plasmodium ovale, although less common, also pose health risks, causing milder but often chronic infections that can persist for years without treatment [3,16,17,18,19].
The lifecycle of Plasmodium species (Figure 1) involves complex interactions between the human host, the Anopheles mosquito vector, and environmental reservoirs. Generally, the lifecycle begins when an infected mosquito transmits Plasmodium sporozoites through its saliva during a blood meal. The sporozoites enter the host’s bloodstream, reaching the liver, where they multiply asexually. The resulting merozoites re-enter the bloodstream and infect red blood cells, leading to the clinical symptoms of malaria. Some merozoites develop into sexual forms (gametocytes), which, when taken up by another mosquito during a blood meal, continue the cycle in the mosquito’s gut, eventually leading to the production of new sporozoites that migrate to the mosquito’s salivary glands, ready to infect a new host [20,21]. The lifecycles of Plasmodium vivax/ovale and Plasmodium falciparum share the basic stages of malaria infection, including the mosquito bite transmission of sporozoites, liver stage development, and blood stage infection. However, a significant difference lies in the liver stage. Plasmodium vivax and Plasmodium ovale have dormant liver stage forms called hypnozoites, which can reactivate months or even years after the initial infection, causing relapses of the disease without the need for another mosquito bite (Table 1). This capability for relapse is absent in Plasmodium falciparum, which does not form hypnozoites and, thus, leads to a single acute phase of illness following the liver stage unless reinfected by another mosquito bite. This difference has profound implications for treatment, as Plasmodium vivax and Plasmodium ovale infections require not only drugs to treat the blood stage of the infection but also an additional treatment with primaquine or other drugs effective against hypnozoites to prevent relapses [18,22].
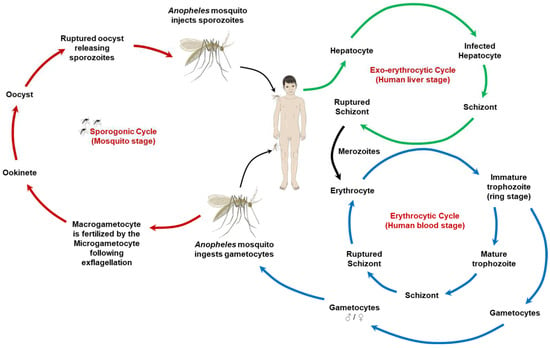
Figure 1.
The lifecycle of malaria parasites. This illustration depicts the complex lifecycle of malaria parasites, which involves both human and mosquito stages. It highlights the various developmental stages of Plasmodium species that cause malaria, transitioning between humans as the host and mosquitoes as the vector. The cycle starts with the infectious bite of a mosquito, releasing sporozoites into the human bloodstream, followed by liver cell invasion and multiplication. The subsequent release of merozoites into the bloodstream marks the beginning of the erythrocytic cycle, where further replication occurs within red blood cells. Some of these parasites develop into gametocytes, which can be taken up by mosquitoes during a blood meal, continuing the cycle in the mosquito host through sexual reproduction (i.e., the stage in which macrogametocyte is fertilized by the microgametocyte following exflagellation to form the zygote/ookinete) to produce new sporozoites, ready to infect another human host.

Table 1.
The lifecycle stages and key features of Plasmodium spp.
The environmental reservoirs, including the breeding habitats of Anopheles mosquitoes, are critical in the malaria lifecycle. Factors such as climate change, deforestation, and water management practices can significantly impact mosquito populations and malaria transmission dynamics [23,24]. The complex lifecycle of Plasmodium species poses significant challenges for vaccine development and disease control strategies. A successful vaccine must induce immunity capable of targeting multiple stages of the parasite’s lifecycle, potentially including the sporozoite stage to prevent infection, the merozoite stage to prevent blood-stage disease, and the gametocyte stage to reduce transmission. The variability in lifecycle stages between Plasmodium species further complicates vaccine development, necessitating species-specific or multispecies approaches [25]. Control strategies must also consider the One Health perspective, recognizing the interplay between human behavior, mosquito ecology, and environmental factors. Integrated approaches that combine vaccine deployment with vector control (e.g., ITNs, IRS), environmental management, and community engagement are essential for effective malaria control and elimination efforts [26,27].
3. Current Landscape of Malaria Vaccine Development
The RTS,S/AS01 vaccine, also known as Mosquirix™, represents the most advanced malaria vaccine to date. It is a recombinant protein-based vaccine designed to induce immunity against the malaria parasite Plasmodium falciparum. Its structure combines the circumsporozoite protein (CSP) of Plasmodium falciparum with the hepatitis B surface antigen (HBsAg), creating virus-like particles. This complex is known as RTS,S, where ‘R’ represents the CSP region containing T-cell epitopes, ‘T’ the HBsAg, ‘S’ the CSP central repeat region, and the additional ‘S’ the HBsAg used to form particles. The adjuvant system AS01, comprising MPL (monophosphoryl lipid A) and QS-21, is employed to enhance the immune response. AS01 triggers a potent adjuvant effect, stimulating both humoral and cellular immune responses, crucial for its protective effects against malaria. The vaccine’s construction aims to elicit a strong immune response against the CSP of Plasmodium falciparum, preventing the parasite from infecting liver cells and, thus, interrupting the lifecycle of the malaria parasite before it can cause disease symptoms [8,28]. This vaccine has undergone extensive Phase III clinical trials, showing a vaccine efficacy of about 36% in children aged 5–17 months over a four-year period [8]. Based on these results, the World Health Organization (WHO) initiated a pilot implementation in three African countries (Ghana, Kenya, and Malawi) in 2019 to further assess the vaccine’s real-world effectiveness and feasibility of delivery within existing health systems [29].
Mosquirix™ has been the subject of numerous studies over the last decade, focusing on its safety, efficacy, and immunogenicity in different populations, primarily in African children. The RTS,S/AS01 vaccine showed greater activity against malaria parasites matching the vaccine strain’s CSP allele, suggesting that its efficacy is influenced by the genetic diversity of the parasite. Vaccine efficacy was about 50% against matched infections in children aged 5 to 17 months [30]. Another RTS,S/AS01E study showed sustained efficacy of at least 15 months, demonstrating potential as a public health intervention against childhood malaria [31]. The vaccine showed an initial protective effect against clinical malaria, but efficacy waned over time, particularly in areas with higher exposure to malaria. This highlights the need for strategies to maintain vaccine effectiveness over time [32]. Immunologically, RTS,S/AS01E induces specific Th1 cell responses in young children, a key target population for the vaccine. The combination of anti-CS antibodies and CS-specific TNFα+ CD4+ T cells accounted for the protective effect [33]. RTS,S/AS01E has also been evaluated for integration into the Expanded Program on Immunization (EPI), showing favorable safety and immunogenicity profiles. This supports its potential use alongside other routine vaccinations in endemic regions [34].
In addition to RTS,S/AS01, other vaccine candidates are currently in clinical trials (Table 2). The R21/Matrix-M vaccine candidate has recently emerged as a promising alternative, demonstrating over 70% efficacy in Phase IIb trials when boosted with the Matrix-M adjuvant. It also targets the CSP but with a different formulation that may enhance its efficacy and durability [35]. In a groundbreaking phase 3 trial, the R21/Matrix-M malaria vaccine demonstrated high efficacy and tolerability in a large cohort of African children aged 5–36 months, across diverse malaria transmission environments. This trial, enrolling over 4800 participants, showcased a 75% and 68% efficacy at seasonal and standard transmission sites, respectively, for preventing the first clinical malaria episode within 12 months post-vaccination. Noteworthy was the vaccine’s safety profile, with only injection site pain and fever reported as common adverse effects, and no significant increase in serious adverse events compared to the control group. Furthermore, the vaccine’s efficacy was robust against multiple malaria episodes and showcased a substantial reduction in malaria cases per 1000 children-years. Interestingly, the efficacy correlated with vaccine-induced antibodies against the CSP, indicating an immune response mechanism that varied by age, with younger children showing higher efficacy [36].
Table 2.
Properties of current malaria vaccine candidates.
The PfSPZ vaccine, developed by Sanaria Inc., uses attenuated Plasmodium falciparum sporozoites harvested from mosquito salivary glands. This vaccine aims to induce immunity by exposing the body to a weakened form of the parasite. The construct involves cryopreserved, aseptic, purified sporozoites that are either irradiated to prevent replication or genetically engineered for attenuation. Immunogenicity studies have shown that the vaccine elicits strong antibody and T-cell responses, critical for protection against malaria [37]. Clinical trials have demonstrated varying levels of efficacy. For instance, in a study involving Tanzanian adults, the vaccine showed 65% efficacy against controlled human malaria infection (CHMI) at 24 weeks with a five-dose regimen, although the immune responses were lower compared to malaria-naïve subjects in the United States [38]. Another trial in Equatorial Guinea indicated that the vaccine was well tolerated and immunogenic, with 70% of vaccinees developing antibodies to the Plasmodium falciparum CSP (PfCSP), albeit at lower levels than in United States participants [39]. Another trial in Mali showed that the vaccine was well tolerated and provided significant protection against malaria, with a vaccine efficacy of 48% at six months post-vaccination [40]. Additionally, a study in Kenyan infants revealed that the vaccine induced significant antibody responses and provided 45.8% efficacy against clinical malaria three months after the last dose [41].
The focus on developing vaccines primarily against Plasmodium falciparum is due to several reasons, with the foremost being its status as the deadliest of the malaria parasites affecting humans. Plasmodium falciparum is responsible for the majority of severe malaria cases and deaths worldwide, particularly in Sub-Saharan Africa, where the burden of the disease is heaviest. This species is known for its ability to cause severe anemia, cerebral malaria, and other life-threatening complications, making it a priority for vaccine development efforts [3,16]. Another reason is the complexity of the malaria parasite’s lifecycle and the genetic diversity of Plasmodium falciparum. The parasite undergoes several stages in both the human host and the mosquito vector, which presents challenges in identifying effective vaccine targets. Furthermore, Plasmodium falciparum has a high level of genetic variability, especially in the antigens that could be targeted by vaccines. Such genetic and antigenic diversities of Plasmodium falciparum pose significant challenges to vaccine development. The parasite’s ability to evade immune responses through variation in its surface antigens, particularly the CSP and merozoite surface proteins, complicates the creation of a broadly effective vaccine. This diversity necessitates the consideration of multiple antigens or epitopes in vaccine design to achieve higher and more sustained efficacy across different geographic regions where genetic variants of the parasite may differ [42]. Modern vaccine efforts, such as those exploring multistage and multiantigen approaches, aim to overcome these hurdles by targeting several key points in the parasite’s lifecycle and incorporating a wider range of antigens to elicit a more robust and comprehensive immune response [43,44].
While there is ongoing research into vaccines against other malaria species such as Plasmodium vivax, these efforts are less advanced. Plasmodium vivax is more prevalent in parts of Asia and Latin America and, although generally considered less deadly than Plasmodium falciparum, it poses unique challenges for vaccine development, including its ability to cause relapsing infections due to dormant liver stages (hypnozoites). The development of vaccines against Plasmodium vivax needs to target hypnozoites and to address the genetic diversity of Plasmodium vivax parasites. One example of ongoing research into a Plasmodium vivax vaccine is the development of the vivax malaria vaccine candidate, VMP001, which targets the CSP of Plasmodium vivax. This vaccine candidate aims to induce an immune response capable of preventing the parasite from establishing infection in the liver. Early-stage clinical trials have been conducted to evaluate the safety, immunogenicity, and efficacy of VMP001 combined with an adjuvant (such as Adjuvant System AS01B) [45].
4. One Health Challenges in Malaria Vaccination Program
The effectiveness of malaria vaccines, particularly those targeting the Plasmodium falciparum parasite, is intricately linked to vector biology and control strategies. The Anopheles mosquitoes, which are the vectors for malaria, have complex lifecycles and behaviors that can significantly influence the transmission dynamics of malaria and, consequently, the efficacy of vaccines. Strategies to control these vectors, including the use of ITNs and IRS, have been successful in reducing malaria transmission rates in various settings. However, the emergence of insecticide resistance among Anopheles populations poses a significant challenge to these control measures and could potentially undermine the gains made by vaccination efforts. The integration of vaccine deployment with effective vector control measures is crucial to achieving sustained reductions in malaria transmission and enhancing vaccine efficacy [26].
Environmental changes, including climate change and deforestation, have a profound impact on malaria transmission dynamics. Climate change affects the distribution, density, and biting behavior of Anopheles mosquitoes by altering the environmental conditions necessary for their development and survival. Warmer temperatures can shorten the development time of the parasite within the mosquito, potentially increasing the transmission potential of malaria. Deforestation and land use changes, on the other hand, can lead to increased breeding sites for mosquitoes, thereby elevating the risk of malaria transmission. These environmental factors not only pose challenges to the current malaria control strategies but also complicate the development and deployment of malaria vaccines by altering exposure patterns and potentially changing the geographical distribution of malaria, necessitating adaptive strategies in vaccine implementation [9,46,47].
The discovery of zoonotic malaria species, such as Plasmodium knowlesi in Southeast Asia, that can be transmitted from wildlife reservoirs (primarily monkeys) to humans has raised concerns about the potential for cross-species transmission and its implications for malaria vaccination effort. These zoonotic species complicate the already challenging task of malaria control and elimination by introducing additional reservoirs of the parasite that are not targeted by current human-focused control measures or vaccines. The existence of wildlife reservoirs necessitates a One Health approach to malaria vaccine development, which considers the ecological and biological interactions between humans, animals, and the environment. Addressing the challenge of zoonotic malaria and wildlife reservoirs requires vaccines that are effective across different Plasmodium species and capable of preventing transmission from nonhuman hosts, alongside strategies to manage the vector populations and habitats in a way that minimizes human–mosquito–wildlife contact [48].
5. Integrating One Health in Malaria Vaccine Research and Development
The One Health concept emphasizes the interconnectedness of human health, animal health, and environmental health, advocating for an integrated approach to disease prevention and control. In the context of malaria vaccine research and development, this approach necessitates collaboration across diverse scientific disciplines including parasitology, veterinary science, environmental science, and public health. Such interdisciplinary research efforts are critical for addressing the complex lifecycle of the malaria parasite, which not only affects humans but also has stages in mosquito vectors and potential nonhuman primate hosts. Understanding the ecological dynamics of mosquito vectors, including their breeding habitats and patterns of interaction with humans, is essential for developing effective malaria control strategies and vaccines. This requires the expertise of environmental scientists, ecologists, and potentially computational biologists, alongside parasitologists who study the disease-causing organisms and public health professionals who implement disease control measures [9,49,50].
One illustrative example of a One Health strategy in malaria control is the deployment of the RTS,S/AS01 (Mosquirix™) vaccine in conjunction with established vector control measures such as ITNs and IRS. The pilot implementation programs initiated by the WHO in African countries such as Ghana, Kenya, and Malawi have demonstrated how combining vaccine deployment with vector control can reduce malaria incidence more effectively than either strategy alone [29]. This integrated approach aligns with One Health principles by addressing both the human and environmental factors contributing to malaria transmission [8]. Another study involves the use of environmental management practices to control mosquito breeding sites alongside community health education programs. For instance, environmental modifications such as the removal of standing water and the promotion of larval source management techniques have been implemented alongside health education campaigns to reduce malaria transmission. This multifaceted approach has been credited with contributing to significant reductions in malaria prevalence in the region [1,51].
The One Health approach also opens avenues for the development of novel vaccine technologies and delivery systems that consider the ecological and social determinants of health. For example, research into nanoparticle-based vaccines offers the potential for creating more stable and effective formulations that can be easily distributed and administered in diverse environmental conditions, addressing logistical challenges in malaria-endemic regions [52,53,54]. Additionally, the exploration of transgenic mosquito strategies, informed by ecological and genetic studies, presents an innovative method of controlling malaria transmission by releasing genetically modified mosquitoes that are resistant to parasite infection or have reduced lifespan, thus interrupting the transmission cycle [55,56,57,58].
6. Ethical, Social, and Economic Considerations in Malaria Vaccination Program
Ethical considerations are paramount in conducting vaccine trials and deploying vaccines, especially in vulnerable populations that are most affected by malaria. Informed consent, equitable access to interventions, and the potential for exploitation are critical concerns. Ethical frameworks emphasize the importance of community engagement, ensuring that participants understand the benefits and risks associated with vaccine trials. Moreover, post-trial access to successful vaccines for trial participants and broader ethical considerations about the allocation of vaccine resources in low-income settings highlight the need for fairness and justice in global health initiatives. The WHO’s guidelines on ethical issues in public health surveillance and research underscore these principles, advocating for transparency, respect, and equity in all aspects of vaccine development and deployment [59].
Social determinants of health, such as poverty, education, and housing conditions, significantly influence malaria risk and vaccine uptake. Populations in malaria-endemic regions often face barriers to accessing healthcare, including vaccines, due to socioeconomic constraints, lack of awareness, and sometimes cultural beliefs that may lead to vaccine hesitancy. Studies have shown that community perceptions and understanding of malaria and vaccines can significantly impact vaccination programs’ success. For instance, misinformation and distrust in healthcare interventions can hinder vaccine acceptance, underscoring the need for culturally sensitive health education and community involvement in vaccination campaigns [60,61].
Malaria imposes significant economic burdens on individuals, families, and countries, particularly in Sub-Saharan Africa [62]. The direct costs of treatment, coupled with indirect costs such as lost productivity and long-term health effects, exacerbate poverty and hinder economic development [63]. Vaccination programs represent a potentially cost-effective strategy to reduce the economic impact of malaria by preventing disease, reducing healthcare expenditures, and improving productivity. Economic evaluations of the RTS,S/AS01 vaccine, for example, suggest that it could be cost-effective in high-burden settings, depending on the vaccine’s price, coverage, and efficacy over time. However, the cost-effectiveness of malaria vaccines also depends on their integration with other malaria control measures, such as vector control and effective case management. Thus, economic analyses must consider the broader context of malaria control strategies and health system strengthening to ensure the sustainable implementation of vaccination programs [64,65,66].
7. Future Directions and Innovations in Malaria Eradication Efforts
The landscape of vaccine research is rapidly evolving, with emerging technologies offering promising new avenues for malaria vaccine development. mRNA vaccines, which have gained significant attention due to their role in COVID-19 vaccination efforts, present a novel platform for malaria vaccine development. These vaccines work by instructing cells to produce a protein that is part of the malaria parasite, triggering an immune response without exposing the recipient to the disease. This technology offers advantages in terms of speed of development and potential efficacy. Moreover, nanoparticle delivery systems are being explored for their ability to enhance immune responses and provide targeted delivery of vaccine components. Nanoparticles can encapsulate antigens or act as adjuvants, improving the stability and immunogenicity of vaccines [52,53]. These innovative approaches could overcome some of the challenges faced by traditional vaccine methods, such as the need for cold storage and the variability in immune responses among different populations.
Genetic engineering offers groundbreaking possibilities for malaria control, particularly through the modification of mosquito vectors to reduce their ability to transmit the disease. CRISPR-Cas9 gene editing technology has been at the forefront of this research, enabling scientists to make precise alterations in the mosquito genome. Strategies include gene drive systems that spread malaria resistance genes through mosquito populations or modifications that reduce mosquito lifespan, thus decreasing the chances of malaria transmission. These approaches aim to complement traditional control methods by targeting the vector’s ability to carry and spread the malaria parasite. While promising, the ecological and ethical implications of releasing genetically modified organisms into the wild necessitate careful consideration and robust regulatory frameworks [67,68,69,70].
The strategy of using Wolbachia, a genus of bacteria, to control mosquito populations represents a novel and promising approach in the fight against mosquito-borne diseases, including malaria. To date, the primary application of Wolbachia-based strategies has been targeting Aedes aegypti mosquitoes, which are vectors for dengue, Zika, chikungunya, and yellow fever, rather than Anopheles mosquitoes, the vectors for malaria. The efforts involving Wolbachia have aimed at either reducing mosquito populations or making them less capable of transmitting diseases. Wolbachia are intracellular bacteria found in many insects, but not naturally in all mosquito species, including Aedes aegypti. When these bacteria are introduced into Aedes mosquitoes, they can interfere with the mosquitoes’ ability to transmit viruses. Additionally, Wolbachia can affect the reproductive processes of their hosts through mechanisms such as cytoplasmic incompatibility, which can reduce mosquito populations over time. One of the most notable efforts in this area has been the World Mosquito Program’s use of Wolbachia-infected Aedes aegypti mosquitoes to fight dengue. By releasing these modified mosquitoes into the environment, the program aims to establish Wolbachia in wild mosquito populations, thereby reducing the incidence of dengue and other arboviral diseases. Field trials in countries such as Australia, Brazil, Indonesia, and Vietnam have shown promising results, with significant reductions in dengue incidence in areas where Wolbachia-infected mosquitoes were released [71]. While the use of Wolbachia has not been directly applied to control Anopheles mosquitoes and malaria transmission, the concept represents an innovative vector control strategy that could potentially be adapted for malaria in the future [72,73,74]. Research is ongoing to explore similar biological control methods for Anopheles mosquitoes, such as gene drive technologies, which could complement traditional malaria control strategies and future vaccine efforts.
Global health initiatives and partnerships play a crucial role in advancing a One Health approach to malaria vaccine development and deployment. Organizations such as the WHO, the Bill & Melinda Gates Foundation, and Gavi, the Vaccine Alliance, have been instrumental in fostering collaboration between governments, research institutions, and the private sector. These partnerships support the integration of human health, animal health, and environmental science perspectives, promoting interdisciplinary research and the sharing of resources and knowledge. Initiatives like the Malaria Vaccine Implementation Programme (MVIP) demonstrate the potential of global collaboration in assessing and enhancing the real-world impact of malaria vaccines [75,76]. Furthermore, the Global Health Innovative Technology (GHIT) Fund exemplifies how international partnerships can accelerate the development of new tools for malaria control, including vaccines, diagnostics, and treatments, by leveraging innovations from around the world [77].
8. Summary
The adoption of a One Health approach to the malaria vaccination program represents a pivotal shift in our strategy to combat this enduring global health challenge. This holistic perspective underscores the intricate connections between human health, animal health, and environmental ecosystems in the transmission and control of malaria. By integrating knowledge across disciplines, we can enhance the efficacy of vaccines, tailor control strategies to specific ecological and social contexts, and, ultimately, achieve more sustainable outcomes in malaria eradication efforts. The potential impact of current and future malaria vaccines on global health cannot be overstated. The RTS,S/AS01 vaccine, as the frontrunner, has already demonstrated the feasibility and benefits of vaccination as part of a broader malaria control strategy. Emerging vaccine technologies, including mRNA platforms and nanoparticle delivery systems, along with innovative approaches like genetic engineering for vector control, promise to further revolutionize our ability to prevent malaria infections. These advancements, informed by One Health principles, offer hope for vaccines that are more effective, accessible, and adaptable to changing environmental and epidemiological landscapes. In conclusion, embracing a One Health approach in malaria vaccination program is not just beneficial—it is imperative for addressing the complex interplay of factors that sustain malaria transmission. This call to action extends to researchers, policymakers, and communities worldwide to unite in a concerted effort to leverage the full spectrum of tools and knowledge available in our fight against malaria, moving closer to a world free from the burden of this disease.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- O’Meara, W.; Mangeni, J.; Steketee, R.; Greenwood, B. Changes in the Burden of Malaria in Sub-Saharan Africa. Lancet Infect. Dis. 2010, 10, 545–555. [Google Scholar] [CrossRef] [PubMed]
- Kamau, A.; Mogeni, P.; Okiro, E.A.; Snow, R.W.; Bejon, P. A Systematic Review of Changing Malaria Disease Burden in Sub-Saharan Africa since 2000: Comparing Model Predictions and Empirical Observations. BMC Med. 2020, 18, 94. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. World Malaria Report 2020: 20 Years of Global Progress and Challenges. 2020. Available online: https://www.who.int/publications/i/item/9789240015791 (accessed on 20 March 2024).
- Liu, Q.; Jing, W.; Kang, L.; Liu, J.; Liu, M. Trends of the Global, Regional and National Incidence of Malaria in 204 Countries from 1990 to 2019 and Implications for Malaria Prevention. J. Travel Med. 2021, 28, taab046. [Google Scholar] [CrossRef]
- Hay, S.I.; Guerra, C.A.; Tatem, A.J.; Noor, A.M.; Snow, R.W. The Global Distribution and Population at Risk of Malaria: Past, Present, and Future. Lancet Infect. Dis. 2004, 4, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Shi, D.; Wei, L.; Liang, H.; Yan, D.; Zhang, J.; Wang, Z. Trends of the Global, Regional and National Incidence, Mortality, and Disability-Adjusted Life Years of Malaria, 1990–2019: An Analysis of the Global Burden of Disease Study 2019. RMHP 2023, 16, 1187–1201. [Google Scholar] [CrossRef]
- Cohen, S. Development of a Malaria Vaccine. J. R. Soc. Med. 1978, 71, 476–478. [Google Scholar] [CrossRef]
- RTS,S Clinical Trials Partnership. Efficacy and Safety of RTS,S/AS01 Malaria Vaccine with or without a Booster Dose in Infants and Children in Africa: Final Results of a Phase 3, Individually Randomised, Controlled Trial. Lancet 2015, 386, 31–45. [Google Scholar] [CrossRef]
- MacDonald, A.J.; Mordecai, E.A. Amazon Deforestation Drives Malaria Transmission, and Malaria Burden Reduces Forest Clearing. Proc. Natl. Acad. Sci. USA 2019, 116, 22212–22218. [Google Scholar] [CrossRef]
- Prusty, D.; Gupta, N.; Upadhyay, A.; Dar, A.; Naik, B.; Kumar, N.; Prajapati, V.K. Asymptomatic Malaria Infection Prevailing Risks for Human Health and Malaria Elimination. Infect. Genet. Evol. 2021, 93, 104987. [Google Scholar] [CrossRef]
- Wilson, A.L.; Courtenay, O.; Kelly-Hope, L.A.; Scott, T.W.; Takken, W.; Torr, S.J.; Lindsay, S.W. The Importance of Vector Control for the Control and Elimination of Vector-Borne Diseases. PLoS Negl. Trop. Dis. 2020, 14, e0007831. [Google Scholar] [CrossRef]
- Benelli, G.; Beier, J.C. Current Vector Control Challenges in the Fight against Malaria. Acta Trop. 2017, 174, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Zinsstag, J.; Schelling, E.; Waltner-Toews, D.; Tanner, M. From “One Medicine” to “One Health” and Systemic Approaches to Health and Well-Being. Prev. Vet. Med. 2011, 101, 148–156. [Google Scholar] [CrossRef]
- Tsoumani, M.E.; Voyiatzaki, C.; Efstathiou, A. Malaria Vaccines: From the Past towards the mRNA Vaccine Era. Vaccines 2023, 11, 1452. [Google Scholar] [CrossRef]
- Lancet, T. Malaria Vaccines: A Test for Global Health. Lancet 2024, 403, 503. [Google Scholar] [CrossRef]
- Monroe, A.; Williams, N.A.; Ogoma, S.; Karema, C.; Okumu, F. Reflections on the 2021 World Malaria Report and the Future of Malaria Control. Malar. J. 2022, 21, 154. [Google Scholar] [CrossRef] [PubMed]
- Baird, J.K. Evidence and Implications of Mortality Associated with Acute Plasmodium Vivax Malaria. Clin. Microbiol. Rev. 2013, 26, 36–57. [Google Scholar] [CrossRef]
- Mueller, I.; Galinski, M.R.; Baird, J.K.; Carlton, J.M.; Kochar, D.K.; Alonso, P.L.; Portillo, H.A. del Key Gaps in the Knowledge of Plasmodium Vivax, a Neglected Human Malaria Parasite. Lancet Infect. Dis. 2009, 9, 555–566. [Google Scholar] [CrossRef] [PubMed]
- Collins, W.E.; Jeffery, G.M. Plasmodium Malariae: Parasite and Disease. Clin. Microbiol. Rev. 2007, 20, 579–592. [Google Scholar] [CrossRef] [PubMed]
- Crompton, P.D.; Moebius, J.; Portugal, S.; Waisberg, M.; Hart, G.; Garver, L.S.; Miller, L.H.; Barillas-Mury, C.; Pierce, S.K. Malaria Immunity in Man and Mosquito: Insights into Unsolved Mysteries of a Deadly Infectious Disease. Annu. Rev. Immunol. 2014, 32, 157–187. [Google Scholar] [CrossRef]
- CDC-Centers for Disease Control and Prevention CDC—Malaria—About Malaria—Biology. Available online: https://www.cdc.gov/malaria/about/biology/index.html (accessed on 10 February 2024).
- World Health Organization. Guidelines for the Treatment of Malaria, 3rd ed.; World Health Organization: Geneva, Switzerland, 2015; ISBN 978-92-4-154912-7. [Google Scholar]
- Paaijmans, K.P.; Read, A.F.; Thomas, M.B. Understanding the Link between Malaria Risk and Climate. Proc. Natl. Acad. Sci. USA 2009, 106, 13844–13849. [Google Scholar] [CrossRef]
- Chandra, G.; Mukherjee, D. Chapter 35—Effect of Climate Change on Mosquito Population and Changing Pattern of Some Diseases Transmitted by Them. In Advances in Animal Experimentation and Modeling; Sobti, R.C., Ed.; Academic Press: Cambridge, MA, USA, 2022; pp. 455–460. ISBN 978-0-323-90583-1. [Google Scholar]
- RTS,S Clinical Trials Partnership. Efficacy and Safety of the RTS,S/AS01 Malaria Vaccine during 18 Months after Vaccination: A Phase 3 Randomized, Controlled Trial in Children and Young Infants at 11 African Sites. PLoS Med. 2014, 11, e1001685. [Google Scholar] [CrossRef]
- Bhatt, S.; Weiss, D.J.; Cameron, E.; Bisanzio, D.; Mappin, B.; Dalrymple, U.; Battle, K.E.; Moyes, C.L.; Henry, A.; Eckhoff, P.A.; et al. The Effect of Malaria Control on Plasmodium Falciparum in Africa between 2000 and 2015. Nature 2015, 526, 207–211. [Google Scholar] [CrossRef]
- Anoopkumar, A.N.; Aneesh, E.M. A Critical Assessment of Mosquito Control and the Influence of Climate Change on Mosquito-Borne Disease Epidemics. Environ. Dev. Sustain. 2022, 24, 8900–8929. [Google Scholar] [CrossRef]
- Kester, K.E.; Cummings, J.F.; Ofori-Anyinam, O.; Ockenhouse, C.F.; Krzych, U.; Moris, P.; Schwenk, R.; Nielsen, R.A.; Debebe, Z.; Pinelis, E.; et al. Randomized, Double-Blind, Phase 2a Trial of Falciparum Malaria Vaccines RTS,S/AS01B and RTS,S/AS02A in Malaria-Naive Adults: Safety, Efficacy, and Immunologic Associates of Protection. J. Infect. Dis. 2009, 200, 337–346. [Google Scholar] [CrossRef]
- Adepoju, P. RTS,S Malaria Vaccine Pilots in Three African Countries. Lancet 2019, 393, 1685. [Google Scholar] [CrossRef] [PubMed]
- Neafsey, D.; Juraska, M.; Bedford, T.; Benkeser, D.; Valim, C.; Griggs, A.; Lievens, M.; Abdulla, S.; Adjei, S.; Agbenyega, T.; et al. Genetic Diversity and Protective Efficacy of the RTS,S/AS01 Malaria Vaccine. N. Engl. J. Med. 2015, 373, 2025–2037. [Google Scholar] [CrossRef] [PubMed]
- Olotu, A.; Lusingu, J.; Leach, A.; Lievens, M.; Vekemans, J.; Msham, S.; Lang, T.; Gould, J.; Dubois, M.; Jongert, E.; et al. Efficacy of RTS,S/AS01E Malaria Vaccine and Exploratory Analysis on Anti-Circumsporozoite Antibody Titres and Protection in Children Aged 5–17 Months in Kenya and Tanzania: A Randomised Controlled Trial. Lancet Infect. Dis. 2011, 11, 102–109. [Google Scholar] [CrossRef]
- Olotu, A.; Fegan, G.; Wambua, J.K.; Nyangweso, G.; Leach, A.; Lievens, M.; Kaslow, D.; Njuguna, P.; Marsh, K.; Bejon, P. Seven-Year Efficacy of RTS,S/AS01 Malaria Vaccine among Young African Children. N. Engl. J. Med. 2016, 374, 2519–2529. [Google Scholar] [CrossRef] [PubMed]
- Olotu, A.; Moris, P.; Mwacharo, J.; Vekemans, J.; Kimani, D.; Janssens, M.; Kai, O.; Jongert, E.; Lievens, M.; Leach, A.; et al. Circumsporozoite-Specific T Cell Responses in Children Vaccinated with RTS,S/AS01E and Protection against P Falciparum Clinical Malaria. PLoS ONE 2011, 6, e25786. [Google Scholar] [CrossRef]
- Agnandji, S.; Asante, K.P.; Lyimo, J.; Vekemans, J.; Soulanoudjingar, S.; Owusu, R.; Shomari, M.; Leach, A.; Fernandes, J.; Dosoo, D.; et al. Evaluation of the Safety and Immunogenicity of the RTS,S/AS01E Malaria Candidate Vaccine When Integrated in the Expanded Program of Immunization. J. Infect. Dis. 2010, 202, 1076–1087. [Google Scholar] [CrossRef]
- Datoo, M.S.; Natama, M.H.; Somé, A.; Traoré, O.; Rouamba, T.; Bellamy, D.; Yameogo, P.; Valia, D.; Tegneri, M.; Ouedraogo, F.; et al. Efficacy of a Low-Dose Candidate Malaria Vaccine, R21 in Adjuvant Matrix-M, with Seasonal Administration to Children in Burkina Faso: A Randomised Controlled Trial. Lancet 2021, 397, 1809–1818. [Google Scholar] [CrossRef]
- Datoo, M.S.; Dicko, A.; Tinto, H.; Ouédraogo, J.-B.; Hamaluba, M.; Olotu, A.; Beaumont, E.; Lopez, F.R.; Natama, H.M.; Weston, S.; et al. Safety and Efficacy of Malaria Vaccine Candidate R21/Matrix-M in African Children: A Multicentre, Double-Blind, Randomised, Phase 3 Trial. Lancet 2024, 403, 533–544. [Google Scholar] [CrossRef] [PubMed]
- Seder, R.A.; Chang, L.-J.; Enama, M.E.; Zephir, K.L.; Sarwar, U.N.; Gordon, I.J.; Holman, L.A.; James, E.R.; Billingsley, P.F.; Gunasekera, A.; et al. Protection against Malaria by Intravenous Immunization with a Nonreplicating Sporozoite Vaccine. Science 2013, 341, 1359–1365. [Google Scholar] [CrossRef]
- Jongo, S.A.; Shekalaghe, S.A.; Church, L.W.P.; Ruben, A.J.; Schindler, T.; Zenklusen, I.; Rutishauser, T.; Rothen, J.; Tumbo, A.; Mkindi, C.; et al. Safety, Immunogenicity, and Protective Efficacy against Controlled Human Malaria Infection of Plasmodium Falciparum Sporozoite Vaccine in Tanzanian Adults. Am. J. Trop. Med. Hyg. 2018, 99, 338–349. [Google Scholar] [CrossRef]
- Olotu, A.; Urbano, V.; Hamad, A.; Eka, M.; Chemba, M.; Nyakarungu, E.; Raso, J.; Eburi, E.; Mandumbi, D.O.; Hergott, D.; et al. Advancing Global Health through Development and Clinical Trials Partnerships: A Randomized, Placebo-Controlled, Double-Blind Assessment of Safety, Tolerability, and Immunogenicity of PfSPZ Vaccine for Malaria in Healthy Equatoguinean Men. Am. J. Trop. Med. Hyg. 2017, 98, 308–318. [Google Scholar] [CrossRef] [PubMed]
- Sissoko, M.S.; Healy, S.A.; Katile, A.; Omaswa, F.; Zaidi, I.; Gabriel, E.E.; Kamate, B.; Samake, Y.; Guindo, M.A.; Dolo, A.; et al. Safety and Efficacy of PfSPZ Vaccine against Plasmodium Falciparum via Direct Venous Inoculation in Healthy Malaria-Exposed Adults in Mali: A Randomised, Double-Blind Phase 1 Trial. Lancet Infect. Dis. 2017, 17, 498–509. [Google Scholar] [CrossRef]
- Oneko, M.; Steinhardt, L.C.; Yego, R.; Wiegand, R.E.; Swanson, P.A.; Kc, N.; Akach, D.; Sang, T.; Gutman, J.R.; Nzuu, E.L.; et al. Safety, Immunogenicity and Efficacy of PfSPZ Vaccine against Malaria in Infants in Western Kenya: A Double-Blind, Randomized, Placebo-Controlled Phase 2 Trial. Nat. Med. 2021, 27, 1636–1645. [Google Scholar] [CrossRef]
- Thera, M.A.; Doumbo, O.K.; Coulibaly, D.; Laurens, M.B.; Ouattara, A.; Kone, A.K.; Guindo, A.B.; Traore, K.; Traore, I.; Kouriba, B.; et al. A Field Trial to Assess a Blood-Stage Malaria Vaccine. N. Engl. J. Med. 2011, 365, 1004–1013. [Google Scholar] [CrossRef] [PubMed]
- Draper, S.J.; Sack, B.K.; King, C.R.; Nielsen, C.M.; Rayner, J.C.; Higgins, M.K.; Long, C.A.; Seder, R.A. Malaria Vaccines: Recent Advances and New Horizons. Cell Host Microbe 2018, 24, 43–56. [Google Scholar] [CrossRef]
- Arama, C.; Troye-Blomberg, M. The Path of Malaria Vaccine Development: Challenges and Perspectives. J. Intern. Med. 2014, 275, 456–466. [Google Scholar] [CrossRef]
- Bennett, J.W.; Yadava, A.; Tosh, D.; Sattabongkot, J.; Komisar, J.; Ware, L.A.; McCarthy, W.F.; Cowden, J.J.; Regules, J.; Spring, M.D.; et al. Phase 1/2a Trial of Plasmodium Vivax Malaria Vaccine Candidate VMP001/AS01B in Malaria-Naive Adults: Safety, Immunogenicity, and Efficacy. PLoS Negl. Trop. Dis. 2016, 10, e0004423. [Google Scholar] [CrossRef] [PubMed]
- Olson, S.H.; Gangnon, R.; Silveira, G.A.; Patz, J.A. Deforestation and Malaria in Mâncio Lima County, Brazil. Emerg. Infect. Dis. 2010, 16, 1108–1115. [Google Scholar] [CrossRef] [PubMed]
- Guerra, C.A.; Snow, R.W.; Hay, S.I. A Global Assessment of Closed Forests, Deforestation and Malaria Risk. Ann. Trop. Med. Parasitol. 2006, 100, 189–204. [Google Scholar] [CrossRef]
- Moyes, C.L.; Shearer, F.M.; Huang, Z.; Wiebe, A.; Gibson, H.S.; Nijman, V.; Mohd-Azlan, J.; Brodie, J.F.; Malaivijitnond, S.; Linkie, M.; et al. Predicting the Geographical Distributions of the Macaque Hosts and Mosquito Vectors of Plasmodium Knowlesi Malaria in Forested and Non-Forested Areas. Parasites Vectors 2016, 9, 242. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.L.; Henry, J.M.; Citron, D.T.; Ssebuliba, D.M.; Nsumba, J.N.; Sánchez, C.H.M.; Brady, O.J.; Guerra, C.A.; García, G.A.; Carter, A.R.; et al. Spatial Dynamics of Malaria Transmission. PLoS Comput. Biol. 2023, 19, e1010684. [Google Scholar] [CrossRef]
- Benelli, G.; Duggan, M.F. Management of Arthropod Vector Data—Social and Ecological Dynamics Facing the One Health Perspective. Acta Trop. 2018, 182, 80–91. [Google Scholar] [CrossRef] [PubMed]
- Taffese, H.S.; Hemming-Schroeder, E.; Koepfli, C.; Tesfaye, G.; Lee, M.; Kazura, J.; Yan, G.-Y.; Zhou, G.-F. Malaria Epidemiology and Interventions in Ethiopia from 2001 to 2016. Infect. Dis. Poverty 2018, 7, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Patra, S.; Singh, M.; Wasnik, K.; Pareek, D.; Gupta, P.S.; Mukherjee, S.; Paik, P. Polymeric Nanoparticle Based Diagnosis and Nanomedicine for Treatment and Development of Vaccines for Cerebral Malaria: A Review on Recent Advancement. ACS Appl. Bio Mater. 2021, 4, 7342–7365. [Google Scholar] [CrossRef]
- Burkhard, P.; Lanar, D.E. Malaria Vaccine Based on Self-Assembling Protein Nanoparticles. Expert Rev. Vaccines 2015, 14, 1525–1527. [Google Scholar] [CrossRef]
- Wu, Y.; Narum, D.L.; Fleury, S.; Jennings, G.; Yadava, A. Particle-Based Platforms for Malaria Vaccines. Vaccine 2015, 33, 7518–7524. [Google Scholar] [CrossRef]
- Dong, S.; Dong, Y.; Simões, M.L.; Dimopoulos, G. Mosquito Transgenesis for Malaria Control. Trends Parasitol. 2022, 38, 54–66. [Google Scholar] [CrossRef]
- Knols, B.G.J.; Bossin, H.C.; Mukabana, W.R.; Robinson, A.S. Transgenic Mosquitoes and the Fight against Malaria: Managing Technology Push in a Turbulent GMO World. Am. J. Trop. Med. Hyg. 2007, 77, 232–242. [Google Scholar] [CrossRef] [PubMed]
- Christophides, G.K. Transgenic Mosquitoes and Malaria Transmission. Cell. Microbiol. 2005, 7, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Tripet, F.; Aboagye-Antwi, F.; Hurd, H. Ecological Immunology of Mosquito–Malaria Interactions. Trends Parasitol. 2008, 24, 219–227. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. WHO Guidelines on Ethical Issues in Public Health Surveillance. In WHO Guidelines on Ethical Issues in Public Health Surveillance; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- MacLennan, C.A.; Saul, A. Vaccines against Poverty. Proc. Natl. Acad. Sci. USA 2014, 111, 12307–12312. [Google Scholar] [CrossRef]
- Ricci, F. Social Implications of Malaria and Their Relationships with Poverty. Mediterr. J. Hematol. Infect. Dis. 2012, 4, e2012048. [Google Scholar] [CrossRef] [PubMed]
- Chima, R.I.; Goodman, C.A.; Mills, A. The Economic Impact of Malaria in Africa: A Critical Review of the Evidence. Health Policy 2003, 63, 17–36. [Google Scholar] [CrossRef]
- Khuu, D.; Eberhard, M.L.; Bristow, B.N.; Javanbakht, M.; Ash, L.R.; Shafir, S.C.; Sorvillo, F.J. Economic Impact of Malaria-Related Hospitalizations in the United States, 2000–2014. J. Infect. Public Health 2019, 12, 424–433. [Google Scholar] [CrossRef]
- Penny, M.A.; Verity, R.; Bever, C.A.; Sauboin, C.; Galactionova, K.; Flasche, S.; White, M.T.; Wenger, E.A.; Velde, N.V.d.; Pemberton-Ross, P.; et al. Public Health Impact and Cost-Effectiveness of the RTS,S/AS01 Malaria Vaccine: A Systematic Comparison of Predictions from Four Mathematical Models. Lancet 2016, 387, 367–375. [Google Scholar] [CrossRef]
- Tediosi, F.; Maire, N.; Penny, M.; Studer, A.; Smith, T.A. Simulation of the Cost-Effectiveness of Malaria Vaccines. Malar. J. 2009, 8, 127. [Google Scholar] [CrossRef]
- Galactionova, K.; Tediosi, F.; Camponovo, F.; Smith, T.A.; Gething, P.W.; Penny, M.A. Country Specific Predictions of the Cost-Effectiveness of Malaria Vaccine RTS,S/AS01 in Endemic Africa. Vaccine 2017, 35, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Kyrou, K.; Hammond, A.M.; Galizi, R.; Kranjc, N.; Burt, A.; Beaghton, A.K.; Nolan, T.; Crisanti, A. A CRISPR–Cas9 Gene Drive Targeting Doublesex Causes Complete Population Suppression in Caged Anopheles Gambiae Mosquitoes. Nat. Biotechnol. 2018, 36, 1062–1066. [Google Scholar] [CrossRef]
- Zhang, C.; Xiao, B.; Jiang, Y.; Zhao, Y.; Li, Z.; Gao, H.; Ling, Y.; Wei, J.; Li, S.; Lu, M.; et al. Efficient Editing of Malaria Parasite Genome Using the CRISPR/Cas9 System. mBio 2014, 5, e01414-e14. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.C.; Fidock, D.A. CRISPR-Mediated Genome Editing of Plasmodium Falciparum Malaria Parasites. Genome Med. 2014, 6, 63. [Google Scholar] [CrossRef] [PubMed]
- Garrood, W.T.; Kranjc, N.; Petri, K.; Kim, D.Y.; Guo, J.A.; Hammond, A.M.; Morianou, I.; Pattanayak, V.; Joung, J.K.; Crisanti, A.; et al. Analysis of Off-Target Effects in CRISPR-Based Gene Drives in the Human Malaria Mosquito. Proc. Natl. Acad. Sci. USA 2021, 118, e2004838117. [Google Scholar] [CrossRef] [PubMed]
- Costa, G.B.; Smithyman, R.; O’Neill, S.L.; Moreira, L.A. How to Engage Communities on a Large Scale? Lessons from World Mosquito Program in Rio de Janeiro, Brazil. Gates Open Res. 2021, 4, 109. [Google Scholar] [CrossRef] [PubMed]
- Walker, T.; Moreira, L.A. Can Wolbachia Be Used to Control Malaria? Mem. Inst. Oswaldo Cruz 2011, 106, 212–217. [Google Scholar] [CrossRef] [PubMed]
- Gomes, F.M.; Barillas-Mury, C. Infection of Anopheline Mosquitoes with Wolbachia: Implications for Malaria Control. PLoS Pathog. 2018, 14, e1007333. [Google Scholar] [CrossRef] [PubMed]
- Hughes, G.L.; Rivero, A.; Rasgon, J.L. Wolbachia Can Enhance Plasmodium Infection in Mosquitoes: Implications for Malaria Control? PLoS Pathog. 2014, 10, e1004182. [Google Scholar] [CrossRef]
- Dimala, C.A.; Kika, B.T.; Kadia, B.M.; Blencowe, H. Current Challenges and Proposed Solutions to the Effective Implementation of the RTS, S/AS01 Malaria Vaccine Program in Sub-Saharan Africa: A Systematic Review. PLoS ONE 2018, 13, e0209744. [Google Scholar] [CrossRef]
- van der Graaf, R.; Macklin, R.; Rid, A.; Bhan, A.; Gefenas, E.; Greco, D.; Haerry, D.; Hurst, S.; London, A.J.; Saracci, R.; et al. Integrating Public Health Programs and Research after the Malaria Vaccine Implementation Program (MVIP): Recommendations for next Steps. Vaccine 2020, 38, 6975–6978. [Google Scholar] [CrossRef] [PubMed]
- Katsuno, K. Japan’s Innovation for Global Health—GHIT’s Catalytic Role. Parasitol. Int. 2021, 80, 102232. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).