Resveratrol Modulates SIRT1 and DNMT Functions and Restores LINE-1 Methylation Levels in ARPE-19 Cells under Oxidative Stress and Inflammation

 ,
,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Oxidative Stress and Inflammatory Conditions Affect Cell Viability in ARPE-19 Cells
2.2. Oxidative Stress Affects DNMT and SIRT1 Functions and LINE-1 Methylation in ARPE-19
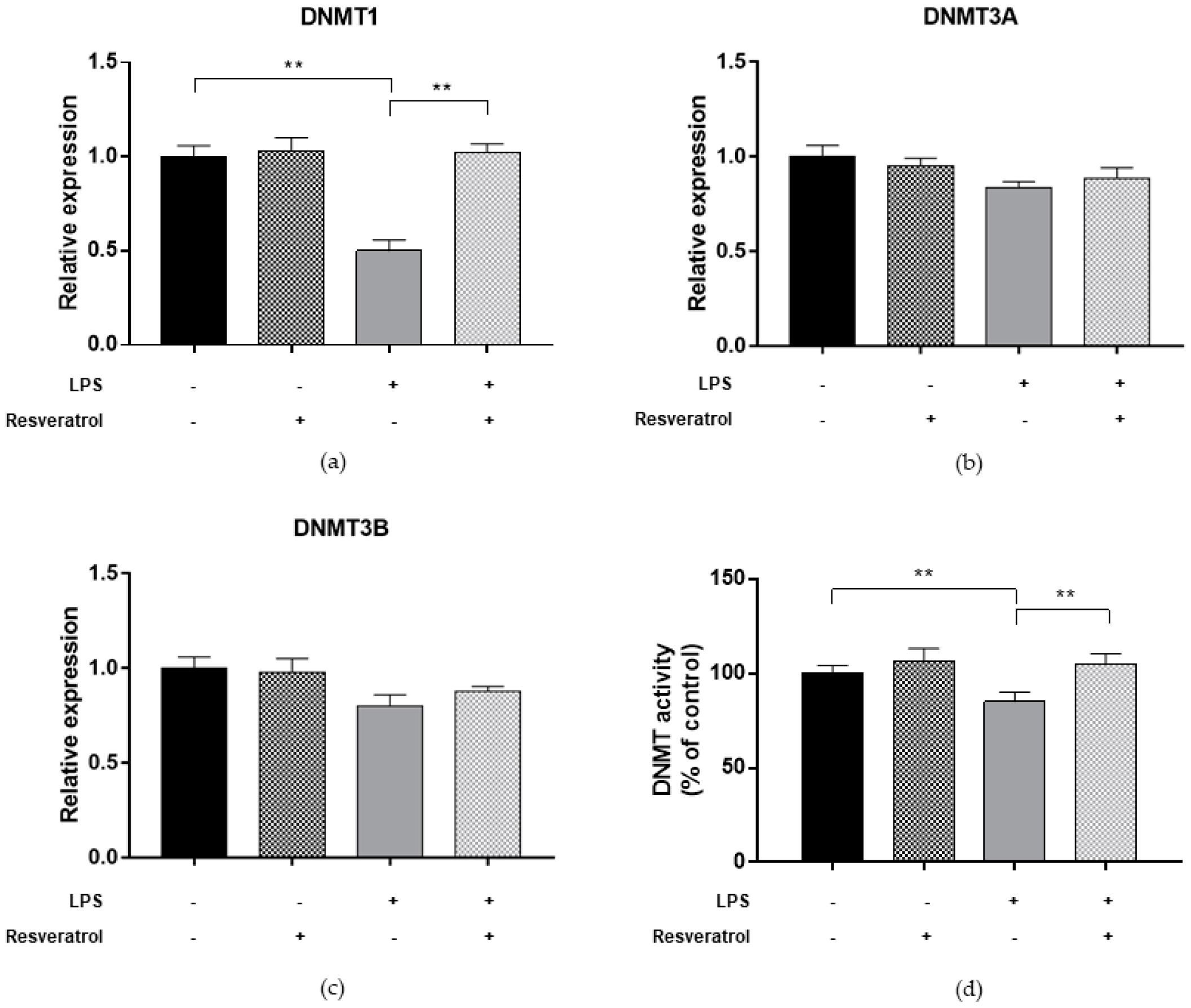
2.3. Inflammatory Condition Affects DNMTs and SIRT1 Functions and LINE-1 Methylation in ARPE-19
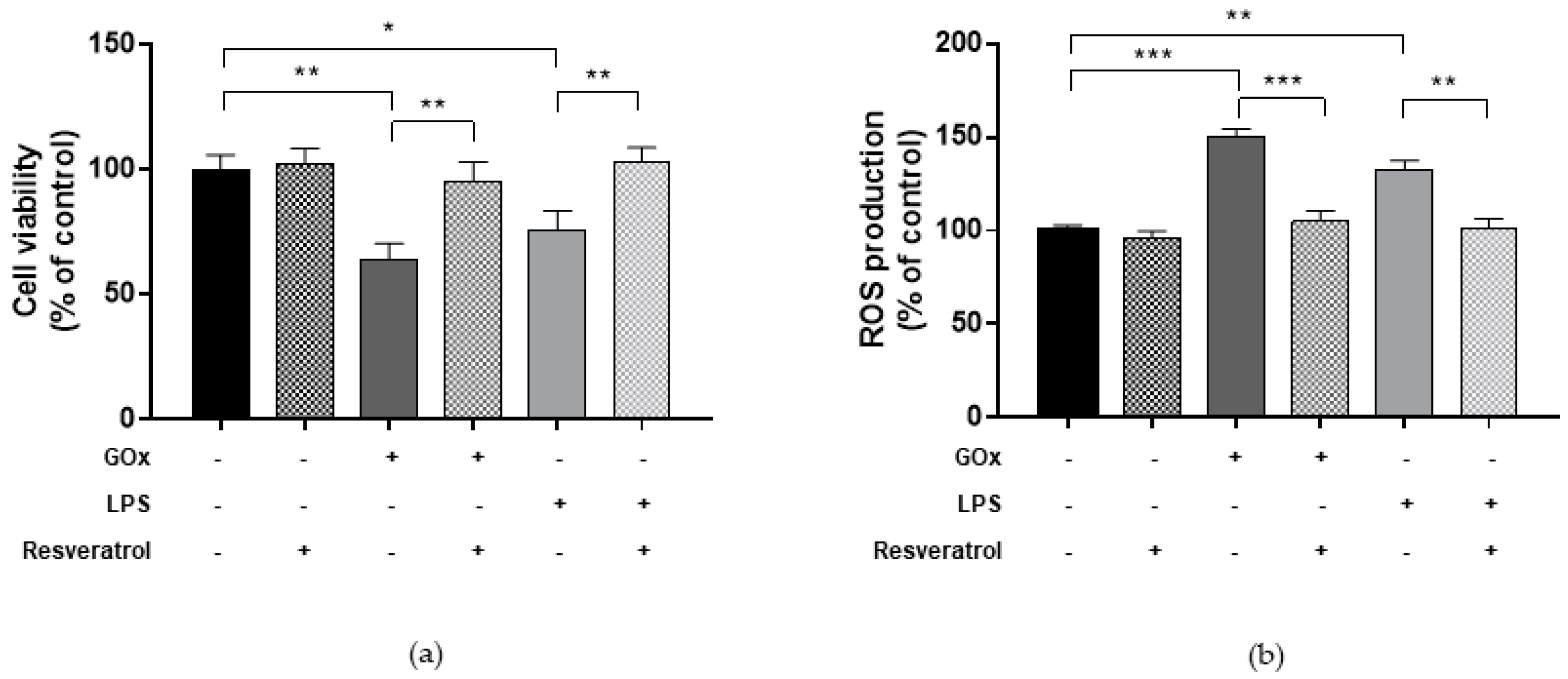
2.4. Resveratrol Ameliorates Viability and ROS Production in Cells under Oxidative and Inflammatory Conditions
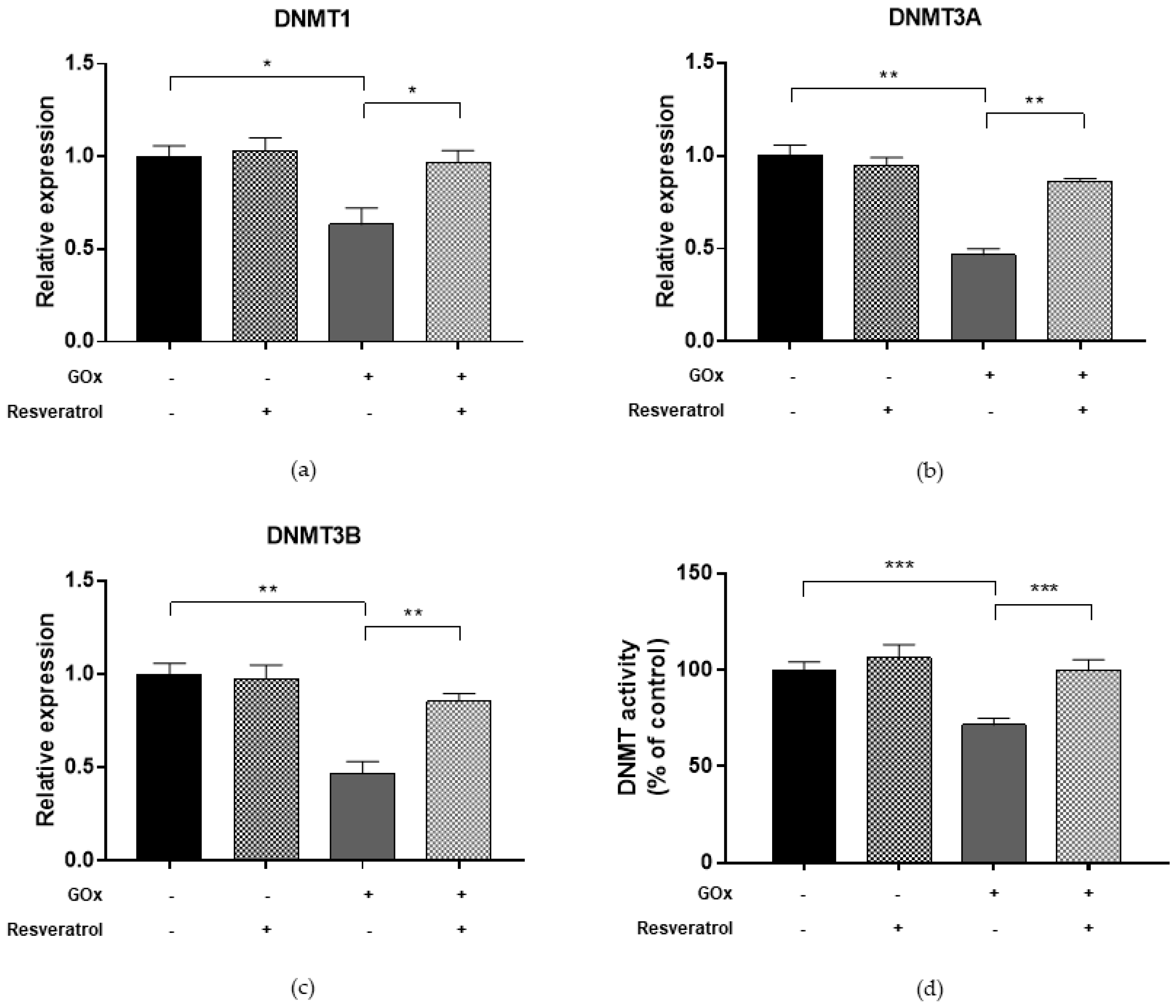
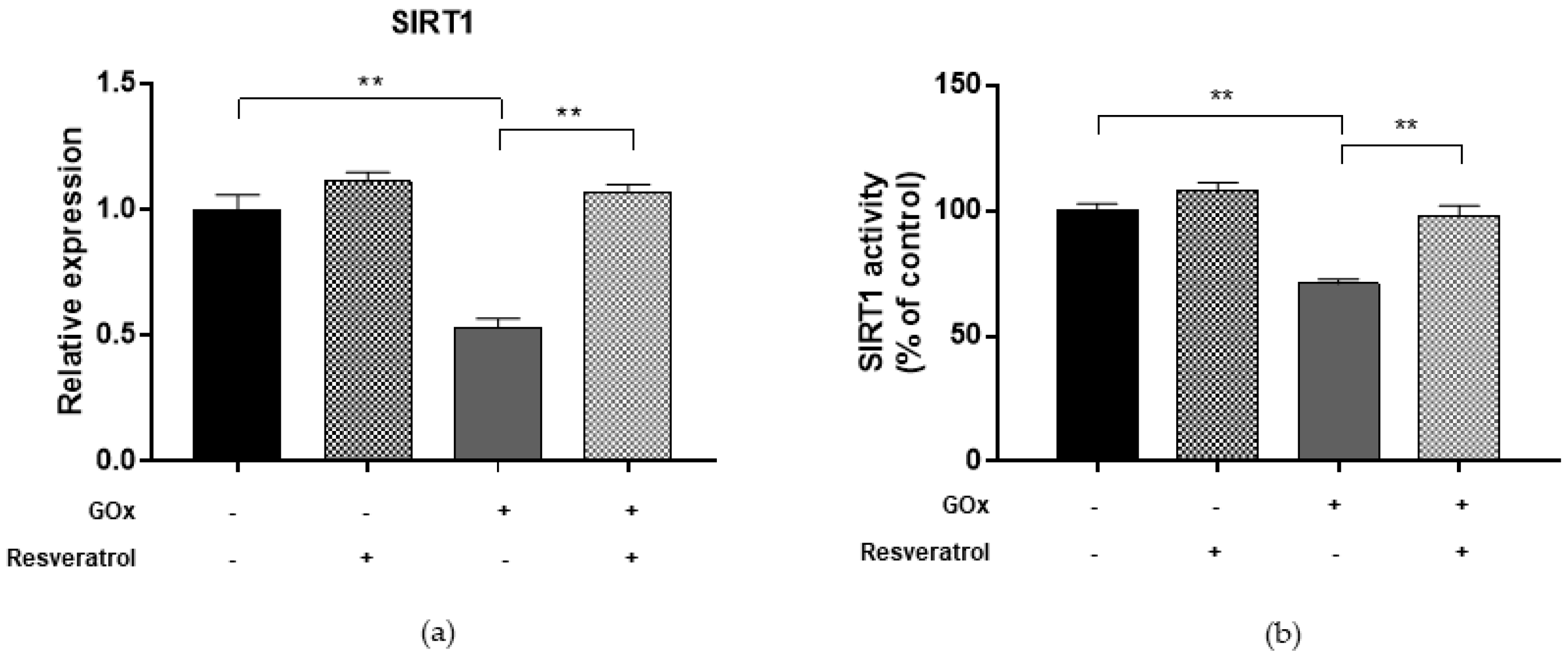
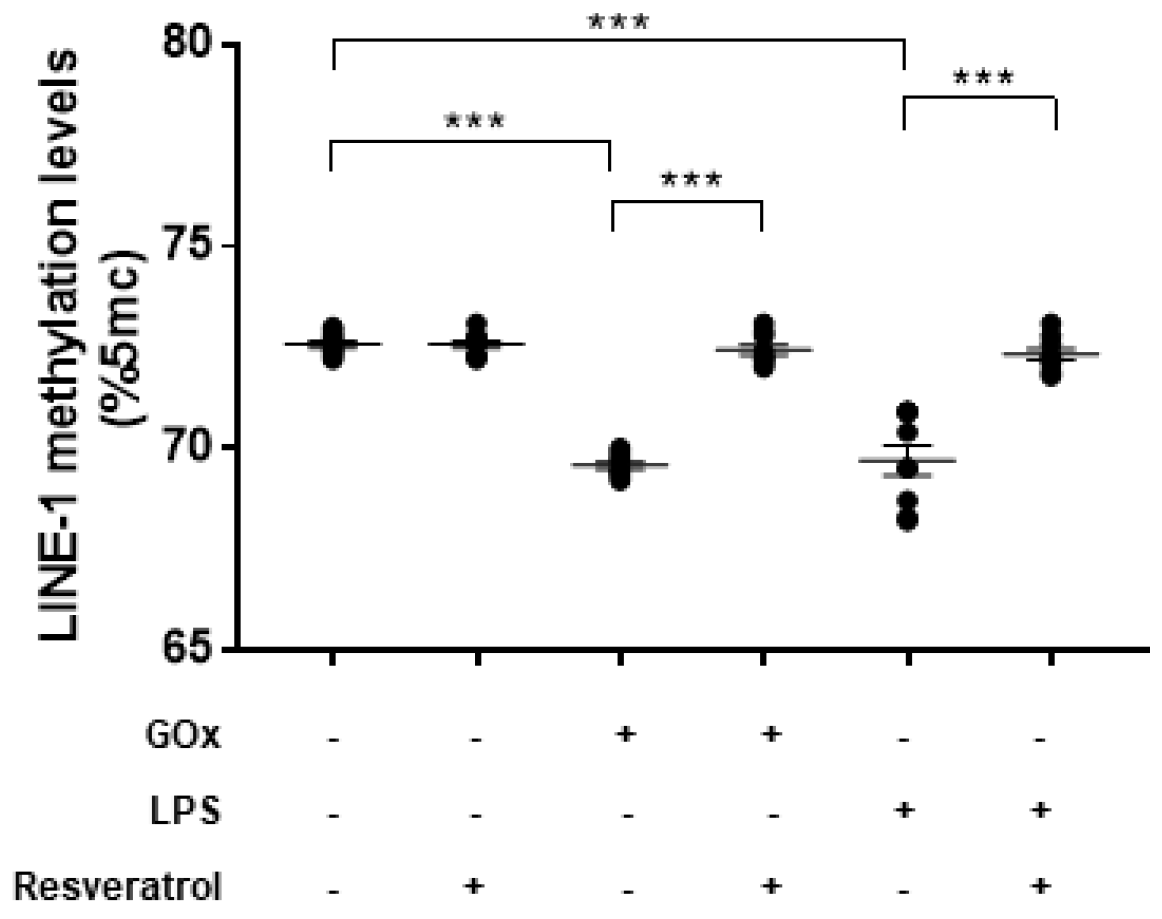
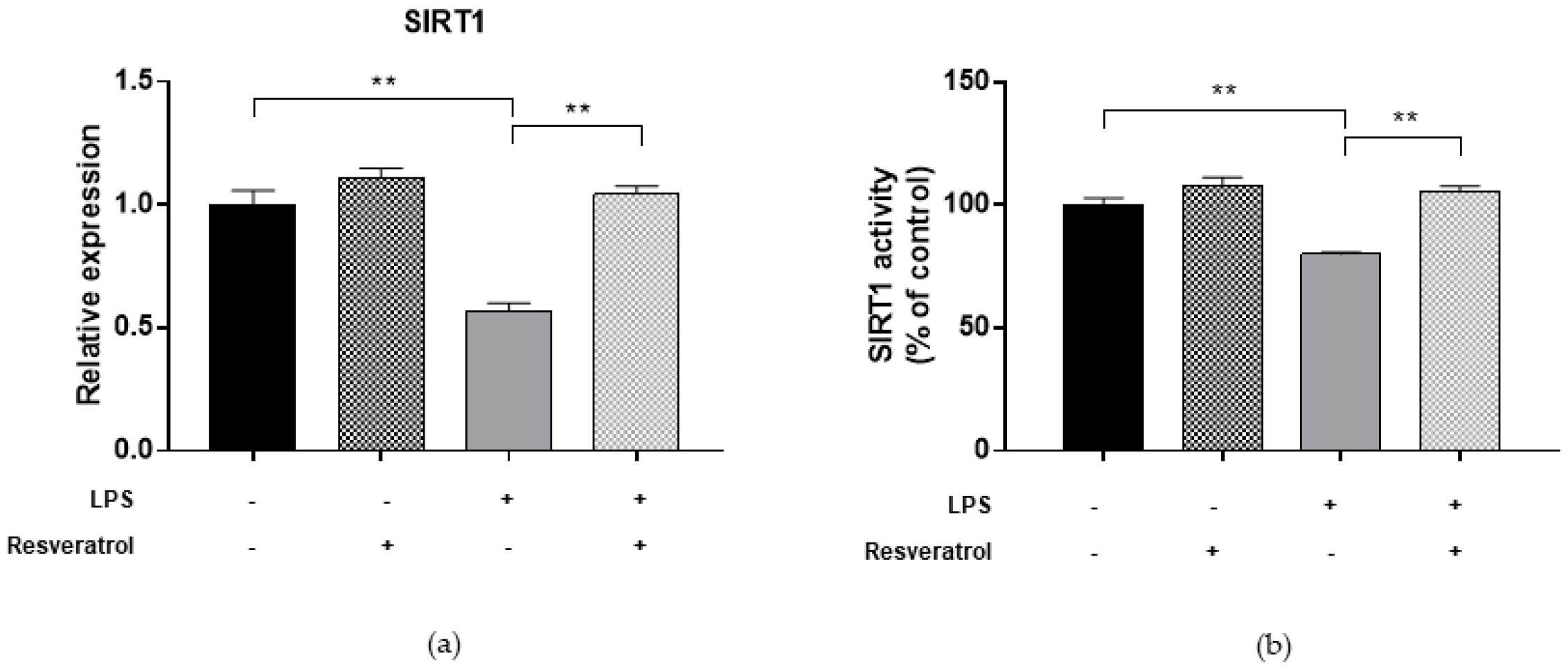
2.5. Resveratrol Restores DNMT and SIRT1 Functions and LINE-1 Methylation in Cells under Oxidative and Inflammatory Conditions
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Treatments
4.2. Determination of Cell Viability
4.3. Determination of ROS
4.4. Nuclear Protein Extraction
4.5. DNMT Activity Quantification
4.6. SIRT1 Activity Quantification
4.7. Quantitative Real-Time Polymerase Chain Reaction (qPCR)
4.8. LINE-1 Methylation Analysis
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AMD | Age-related macular degeneration |
| Aβ | Amyloid beta |
| ARPE-19 | Human retinal pigment epithelial |
| CNV | Choroidal neovascularization |
| DCFDA | 2′,7′–dichlorofluorescin diacetate |
| DMEM | Dulbecco’s Modified Eagle’s medium |
| DNMTs | DNA methyltransferases |
| GOx | Gluocose oxidase |
| GSTM1 | Glutathione S-transferase isoforms mu1 |
| GSTM5 | Glutathione S-transferase isoforms mu5 |
| IL17RC | Interleukin 17 receptor C |
| IL1β | Interleukin 1 beta |
| LINE-1 | Long interspersed nuclear element-1 |
| LPS | Lipopolysaccharid |
| MTT | Thiazolyl blue tetrazolium bromide |
| NF-κB | Nuclear factor-kappa B |
| qPCR | Quantitative real-time polymerase chain reaction |
| ROS | Reactive oxygen species |
| RPE | Retinal pigment epithelium |
| TNFα, | Tumor necrosis factor alpha |
| SIRT1 | Sirtuin 1 |
References
- Rudnicka, A.R.; Jarrar, Z.; Wormald, R.; Cook, D.G.; Fletcher, A.; Owen, C.G. Age and gender variations in age-related macular degeneration prevalence in populations of european ancestry: A meta-analysis. Ophthalmology 2012, 119, 571–580. [Google Scholar] [CrossRef] [PubMed]
- Jager, R.D.; Mieler, W.F.; Miller, J.W. Age-related macular degeneration. N. Engl. J. Med. 2008, 358, 2606–2617. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.; Patel, M.; Chan, C.C. Molecular pathology of age-related macular degeneration. Prog. Retin. Eye Res. 2009, 28, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferris, F.L.; Davis, M.D.; Clemons, T.E.; Lee, L.Y.; Chew, E.Y.; Lindblad, A.S.; Milton, R.C.; Bressler, S.B.; Klein, R. Age-Related Eye Disease Study (AREDS) Research Group. A simplified severity scale for age-related macular degeneration: Areds report no. 18. Arch. Ophthalmol. 2005, 123, 1570–1574. [Google Scholar] [PubMed]
- Colijn, J.M.; Buitendijk, G.H.S.; Prokofyeva, E.; Alves, D.; Cachulo, M.L.; Khawaja, A.P.; Cougnard-Gregoire, A.; Merle, B.M.J.; Korb, C.; Erke, M.G.; et al. Prevalence of age-related macular degeneration in europe: The past and the future. Ophthalmology 2017, 124, 1753–1763. [Google Scholar] [CrossRef] [PubMed]
- Tomany, S.C.; Cruickshanks, K.J.; Klein, R.; Klein, B.E.; Knudtson, M.D. Sunlight and the 10-year incidence of age-related maculopathy: The beaver dam eye study. Arch. Ophthalmol. 2004, 122, 750–757. [Google Scholar] [CrossRef] [PubMed]
- Thornton, J.; Edwards, R.; Mitchell, P.; Harrison, R.A.; Buchan, I.; Kelly, S.P. Smoking and age-related macular degeneration: A review of association. Eye 2005, 19, 935–944. [Google Scholar] [CrossRef] [PubMed]
- Seddon, J.M.; Ajani, U.A.; Sperduto, R.D.; Hiller, R.; Blair, N.; Burton, T.C.; Farber, M.D.; Gragoudas, E.S.; Haller, J.; Miller, D.T. Dietary carotenoids, vitamins a, c, and e, and advanced age-related macular degeneration. Eye disease case-control study group. JAMA 1994, 272, 1413–1420. [Google Scholar] [CrossRef] [PubMed]
- Tomany, S.C.; Klein, R.; Klein, B.E. Beaver Dam Eye Study. The relationship between iris color, hair color, and skin sun sensitivity and the 10-year incidence of age-related maculopathy: The beaver dam eye study. Ophthalmology 2003, 110, 1526–1533. [Google Scholar] [CrossRef]
- Seddon, J.M.; Rosner, B.; Sperduto, R.D.; Yannuzzi, L.; Haller, J.A.; Blair, N.P.; Willett, W. Dietary fat and risk for advanced age-related macular degeneration. Arch. Ophthalmol. 2001, 119, 1191–1199. [Google Scholar] [CrossRef] [PubMed]
- Fritsche, L.G.; Chen, W.; Schu, M.; Yaspan, B.L.; Yu, Y.; Thorleifsson, G.; Zack, D.J.; Arakawa, S.; Cipriani, V.; Ripke, S.; et al. Seven new loci associated with age-related macular degeneration. Nat. Genet. 2013, 45, 433–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fritsche, L.G.; Igl, W.; Bailey, J.N.; Grassmann, F.; Sengupta, S.; Bragg-Gresham, J.L.; Burdon, K.P.; Hebbring, S.J.; Wen, C.; Gorski, M.; et al. A large genome-wide association study of age-related macular degeneration highlights contributions of rare and common variants. Nat. Genet. 2016, 48, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Barchitta, M.; Maugeri, A. Association between vascular endothelial growth factor polymorphisms and age-related macular degeneration: An updated meta-analysis. Dis. Markers 2016, 8486406. [Google Scholar] [CrossRef] [PubMed]
- Maugeri, A.; Barchitta, M.; Agodi, A. The association between complement factor h rs1061170 polymorphism and age-related macular degeneration: A comprehensive meta-analysis stratified by stage of disease and ethnicity. Acta Ophthalmol. 2018, in press. [Google Scholar]
- Seddon, J.M.; George, S.; Rosner, B.; Klein, M.L. Cfh gene variant, y402h, and smoking, body mass index, environmental associations with advanced age-related macular degeneration. Hum. Hered. 2006, 61, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Rhee, S.G.; Bae, Y.S.; Lee, S.R.; Kwon, J. Hydrogen peroxide: A key messenger that modulates protein phosphorylation through cysteine oxidation. Sci. STKE 2000, 2000. [Google Scholar] [CrossRef] [PubMed]
- Finkel, T. Signal transduction by reactive oxygen species. J. Cell Biol. 2011, 194, 7–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eno, C.O.; Zhao, G.; Venkatanarayan, A.; Wang, B.; Flores, E.R.; Li, C. Noxa couples lysosomal membrane permeabilization and apoptosis during oxidative stress. Free Radic. Biol. Med. 2013, 65, 26–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.N.; Jung, H.Y.; Eum, W.S.; Kim, D.W.; Shin, M.J.; Ahn, E.H.; Kim, S.J.; Lee, C.H.; Yong, J.I.; Ryu, E.J.; et al. Neuroprotective effects of pep-1-carbonyl reductase 1 against oxidative-stress-induced ischemic neuronal cell damage. Free Radic. Biol. Med. 2014, 69, 181–196. [Google Scholar] [CrossRef] [PubMed]
- Medeiros, R.; Rodrigues, G.B.; Figueiredo, C.P.; Rodrigues, E.B.; Grumman, A.; Menezes-de-Lima, O.; Passos, G.F.; Calixto, J.B. Molecular mechanisms of topical anti-inflammatory effects of lipoxin a(4) in endotoxin-induced uveitis. Mol. Pharmacol. 2008, 74, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Lu, H.; Hu, X.; Chen, W.; Xu, Y.; Wang, J. Expression of tlr4-myd88 and NF-κB in the iris during endotoxin-induced uveitis. Mediat. Inflamm. 2010, 2010, 748218. [Google Scholar] [CrossRef] [PubMed]
- Girol, A.P.; Mimura, K.K.; Drewes, C.C.; Bolonheis, S.M.; Solito, E.; Farsky, S.H.; Gil, C.D.; Oliani, S.M. Anti-inflammatory mechanisms of the annexin a1 protein and its mimetic peptide ac2-26 in models of ocular inflammation in vivo and in vitro. J. Immunol. 2013, 190, 5689–5701. [Google Scholar] [CrossRef] [PubMed]
- Paimela, T.; Ryhänen, T.; Mannermaa, E.; Ojala, J.; Kalesnykas, G.; Salminen, A.; Kaarniranta, K. The effect of 17beta-estradiol on il-6 secretion and nf-kappab DNA-binding activity in human retinal pigment epithelial cells. Immunol. Lett. 2007, 110, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Leung, K.W.; Barnstable, C.J.; Tombran-Tink, J. Bacterial endotoxin activates retinal pigment epithelial cells and induces their degeneration through il-6 and il-8 autocrine signaling. Mol. Immunol. 2009, 46, 1374–1386. [Google Scholar] [CrossRef] [PubMed]
- Saccà, S.C.; Roszkowska, A.M.; Izzotti, A. Environmental light and endogenous antioxidants as the main determinants of non-cancer ocular diseases. Mutat. Res. 2013, 752, 153–171. [Google Scholar] [CrossRef] [PubMed]
- Sacca, S.C.; Bolognesi, C.; Battistella, A.; Bagnis, A.; Izzotti, A. Gene-environment interactions in ocular diseases. Mutat. Res. 2009, 667, 98–117. [Google Scholar] [CrossRef] [PubMed]
- Wolffe, A.P.; Guschin, D. Review: Chromatin structural features and targets that regulate transcription. J. Struct. Biol. 2000, 129, 102–122. [Google Scholar] [CrossRef] [PubMed]
- Franchini, D.M.; Schmitz, K.M.; Petersen-Mahrt, S.K. 5-methylcytosine DNA demethylation: More than losing a methyl group. Annu. Rev. Genet. 2012, 46, 419–441. [Google Scholar] [CrossRef] [PubMed]
- Clapier, C.R.; Cairns, B.R. The biology of chromatin remodeling complexes. Annu. Rev. Biochem. 2009, 78, 273–304. [Google Scholar] [CrossRef] [PubMed]
- Auclair, G.; Weber, M. Mechanisms of DNA methylation and demethylation in mammals. Biochimie 2012, 94, 2202–2211. [Google Scholar] [CrossRef] [PubMed]
- Fabris, S.; Ronchetti, D.; Agnelli, L.; Baldini, L.; Morabito, F.; Bicciato, S.; Basso, D.; Todoerti, K.; Lombardi, L.; Lambertenghi-Deliliers, G.; et al. Transcriptional features of multiple myeloma patients with chromosome 1q gain. Leukemia 2007, 21, 1113–1116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carreira, P.E.; Richardson, S.R.; Faulkner, G.J. L1 retrotransposons, cancer stem cells and oncogenesis. FEBS J. 2014, 281, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Rodić, N.; Burns, K.H. Long interspersed element-1 (line-1): Passenger or driver in human neoplasms? PLoS Genet. 2013, 9, e1003402. [Google Scholar] [CrossRef] [PubMed]
- Barchitta, M.; Quattrocchi, A.; Maugeri, A.; Vinciguerra, M.; Agodi, A. Line-1 hypomethylation in blood and tissue samples as an epigenetic marker for cancer risk: A systematic review and meta-analysis. PLoS ONE 2014, 9, e109478. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.; Oka, S.; Lee, D.Y.; Park, J.; Aponte, A.M.; Jung, Y.S.; Bitterman, J.; Zhai, P.; He, Y.; Kooshapur, H.; et al. Sirt1 carboxyl-domain is an atp-repressible domain that is transferrable to other proteins. Nat. Commun. 2017, 8, 15560. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.; Yuan, Z.; Ling, H.; Fukasawa, K.; Robertson, K.; Olashaw, N.; Koomen, J.; Chen, J.; Lane, W.S.; Seto, E. Sirt1 deacetylates the DNA methyltransferase 1 (dnmt1) protein and alters its activities. Mol. Cell. Biol. 2011, 31, 4720–4734. [Google Scholar] [CrossRef] [PubMed]
- Howitz, K.T.; Bitterman, K.J.; Cohen, H.Y.; Lamming, D.W.; Lavu, S.; Wood, J.G.; Zipkin, R.E.; Chung, P.; Kisielewski, A.; Zhang, L.L.; et al. Small molecule activators of sirtuins extend saccharomyces cerevisiae lifespan. Nature 2003, 425, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Jang, M.; Cai, L.; Udeani, G.O.; Slowing, K.V.; Thomas, C.F.; Beecher, C.W.; Fong, H.H.; Farnsworth, N.R.; Kinghorn, A.D.; Mehta, R.G.; et al. Cancer chemopreventive activity of resveratrol, a natural product derived from grapes. Science 1997, 275, 218–220. [Google Scholar] [CrossRef] [PubMed]
- Lançon, A.; Frazzi, R.; Latruffe, N. Anti-oxidant, anti-inflammatory and anti-angiogenic properties of resveratrol in ocular diseases. Molecules 2016, 21, 304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iloki-Assanga, S.B.; Lewis-Luján, L.M.; Fernández-Angulo, D.; Gil-Salido, A.A.; Lara-Espinoza, C.L.; Rubio-Pino, J.L. Retino-protective effect of bucida buceras against oxidative stress induced by h2o2 in human retinal pigment epithelial cells line. BMC Complement. Altern. Med. 2015, 15, 254. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.M.; Huang, C.H.; Li, H.J.; Hsiao, C.Y.; Su, C.C.; Lee, P.L.; Hung, C.F. Protective effects of resveratrol against uva-induced damage in arpe19 cells. Int. J. Mol. Sci. 2015, 16, 5789–5802. [Google Scholar] [CrossRef] [PubMed]
- Shaw, P.X.; Stiles, T.; Douglas, C.; Ho, D.; Fan, W.; Du, H.; Xiao, X. Oxidative stress, innate immunity, and age-related macular degeneration. AIMS Mol. Sci. 2016, 3, 196–221. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Li, P.; Fang, L.; Zhu, H.; Xu, L.; Cheng, H.; Zhang, J.; Li, F.; Feng, Y.; Li, Y.; et al. Negative regulation of dnmt3a de novo DNA methylation by frequently overexpressed UHRF family proteins as a mechanism for widespread DNA hypomethylation in cancer. Cell Discov. 2016, 2, 16007. [Google Scholar] [CrossRef] [PubMed]
- Maugeri, A.; Mazzone, M.G.; Giuliano, F.; Vinciguerra, M.; Basile, G.; Barchitta, M.; Agodi, A. Curcumin modulates DNA methyltransferases functions in a cellular model of diabetic retinopathy. Oxid. Med. Cell. Longev. 2018, 2018, 5407482. [Google Scholar] [CrossRef]
- Hunter, A.; Spechler, P.A.; Cwanger, A.; Song, Y.; Zhang, Z.; Ying, G.S.; Hunter, A.K.; Dezoeten, E.; Dunaief, J.L. DNA methylation is associated with altered gene expression in amd. Investig. Ophthalmol. Vis. Sci. 2012, 53, 2089–2105. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Liu, B.; Tuo, J.; Shen, D.; Chen, P.; Li, Z.; Liu, X.; Ni, J.; Dagur, P.; Sen, H.N.; et al. Hypomethylation of the il17rc promoter associates with age-related macular degeneration. Cell Rep. 2012, 2, 1151–1158. [Google Scholar] [CrossRef] [PubMed]
- Oliver, V.F.; Franchina, M.; Jaffe, A.E.; Branham, K.E.; Othman, M.; Heckenlively, J.R.; Swaroop, A.; Campochiaro, B.; Vote, B.J.; Craig, J.E.; et al. Hypomethylation of the il17rc promoter in peripheral blood leukocytes is not a hallmark of age-related macular degeneration. Cell Rep. 2013, 5, 1527–1535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, C.H.; Cherng, J.Y.; Chiou, G.Y.; Chen, Y.C.; Chien, C.H.; Kao, C.L.; Chang, Y.L.; Chien, Y.; Chen, L.K.; Liu, J.H.; et al. Delivery of oct4 and sirt1 with cationic polyurethanes-short branch pei to aged retinal pigment epithelium. Biomaterials 2011, 32, 9077–9088. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Liu, C.; Wang, F.; Wang, H. Sirt1 negatively regulates amyloid-beta-induced inflammation via the NF-κB pathway. Braz. J. Med. Biol. Res. 2013, 46, 659–669. [Google Scholar] [CrossRef] [PubMed]
- Kauppinen, A.; Suuronen, T.; Ojala, J.; Kaarniranta, K.; Salminen, A. Antagonistic crosstalk between NF-κB and sirt1 in the regulation of inflammation and metabolic disorders. Cell. Signal. 2013, 25, 1939–1948. [Google Scholar] [CrossRef] [PubMed]
- Maloney, S.C.; Antecka, E.; Granner, T.; Fernandes, B.; Lim, L.A.; Orellana, M.E.; Burnier, M.N. Expression of sirt1 in choroidal neovascular membranes. Retina 2013, 33, 862–866. [Google Scholar] [CrossRef] [PubMed]
- Nagineni, C.N.; Raju, R.; Nagineni, K.K.; Kommineni, V.K.; Cherukuri, A.; Kutty, R.K.; Hooks, J.J.; Detrick, B. Resveratrol suppresses expression of vegf by human retinal pigment epithelial cells: Potential nutraceutical for age-related macular degeneration. Aging Dis. 2014, 5, 88–100. [Google Scholar] [PubMed]
- Balaiya, S.; Murthy, R.K.; Chalam, K.V. Resveratrol inhibits proliferation of hypoxic choroidal vascular endothelial cells. Mol. Vis. 2013, 19, 2385–2392. [Google Scholar] [PubMed]
- Zhao, P.; Sui, B.D.; Liu, N.; Lv, Y.J.; Zheng, C.X.; Lu, Y.B.; Huang, W.T.; Zhou, C.H.; Chen, J.; Pang, D.L.; et al. Anti-aging pharmacology in cutaneous wound healing: Effects of metformin, resveratrol, and rapamycin by local application. Aging Cell 2017, 16, 1083–1093. [Google Scholar] [CrossRef] [PubMed]
- Yurdagul, A.; Kleinedler, J.J.; McInnis, M.C.; Khandelwal, A.R.; Spence, A.L.; Orr, A.W.; Dugas, T.R. Resveratrol promotes endothelial cell wound healing under laminar shear stress through an estrogen receptor-α-dependent pathway. Am. J. Physiol. Heart Circ. Physiol. 2014, 306, H797–806. [Google Scholar] [CrossRef] [PubMed]
- Tsai, T.Y.; Chen, T.C.; Wang, I.J.; Yeh, C.Y.; Su, M.J.; Chen, R.H.; Tsai, T.H.; Hu, F.R. The effect of resveratrol on protecting corneal epithelial cells from cytotoxicity caused by moxifloxacin and benzalkonium chloride. Investig. Ophthalmol. Vis. Sci. 2015, 56, 1575–1584. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative pcr and the 2(-delta delta c(t)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Agodi, A.; Barchitta, M.; Quattrocchi, A.; Maugeri, A.; Canto, C.; Marchese, A.E.; Vinciguerra, M. Low fruit consumption and folate deficiency are associated with line-1 hypomethylation in women of a cancer-free population. Genes Nutr. 2015, 10, 480. [Google Scholar] [CrossRef] [PubMed]
- Barchitta, M.; Quattrocchi, A.; Maugeri, A.; Canto, C.; La Rosa, N.; Cantarella, M.A.; Spampinato, G.; Scalisi, A.; Agodi, A. Line-1 hypermethylation in white blood cell DNA is associated with high-grade cervical intraepithelial neoplasia. BMC Cancer 2017, 17, 601. [Google Scholar] [CrossRef] [PubMed]






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maugeri, A.; Barchitta, M.; Mazzone, M.G.; Giuliano, F.; Basile, G.; Agodi, A. Resveratrol Modulates SIRT1 and DNMT Functions and Restores LINE-1 Methylation Levels in ARPE-19 Cells under Oxidative Stress and Inflammation. Int. J. Mol. Sci. 2018, 19, 2118. https://doi.org/10.3390/ijms19072118
Maugeri A, Barchitta M, Mazzone MG, Giuliano F, Basile G, Agodi A. Resveratrol Modulates SIRT1 and DNMT Functions and Restores LINE-1 Methylation Levels in ARPE-19 Cells under Oxidative Stress and Inflammation. International Journal of Molecular Sciences. 2018; 19(7):2118. https://doi.org/10.3390/ijms19072118
Chicago/Turabian StyleMaugeri, Andrea, Martina Barchitta, Maria Grazia Mazzone, Francesco Giuliano, Guido Basile, and Antonella Agodi. 2018. "Resveratrol Modulates SIRT1 and DNMT Functions and Restores LINE-1 Methylation Levels in ARPE-19 Cells under Oxidative Stress and Inflammation" International Journal of Molecular Sciences 19, no. 7: 2118. https://doi.org/10.3390/ijms19072118
APA StyleMaugeri, A., Barchitta, M., Mazzone, M. G., Giuliano, F., Basile, G., & Agodi, A. (2018). Resveratrol Modulates SIRT1 and DNMT Functions and Restores LINE-1 Methylation Levels in ARPE-19 Cells under Oxidative Stress and Inflammation. International Journal of Molecular Sciences, 19(7), 2118. https://doi.org/10.3390/ijms19072118