Role of Maternal Dietary Protein and Amino Acids on Fetal Programming, Early Neonatal Development, and Lactation in Swine

 ,
, 


Abstract
:Simple Summary
Abstract
1. Introduction
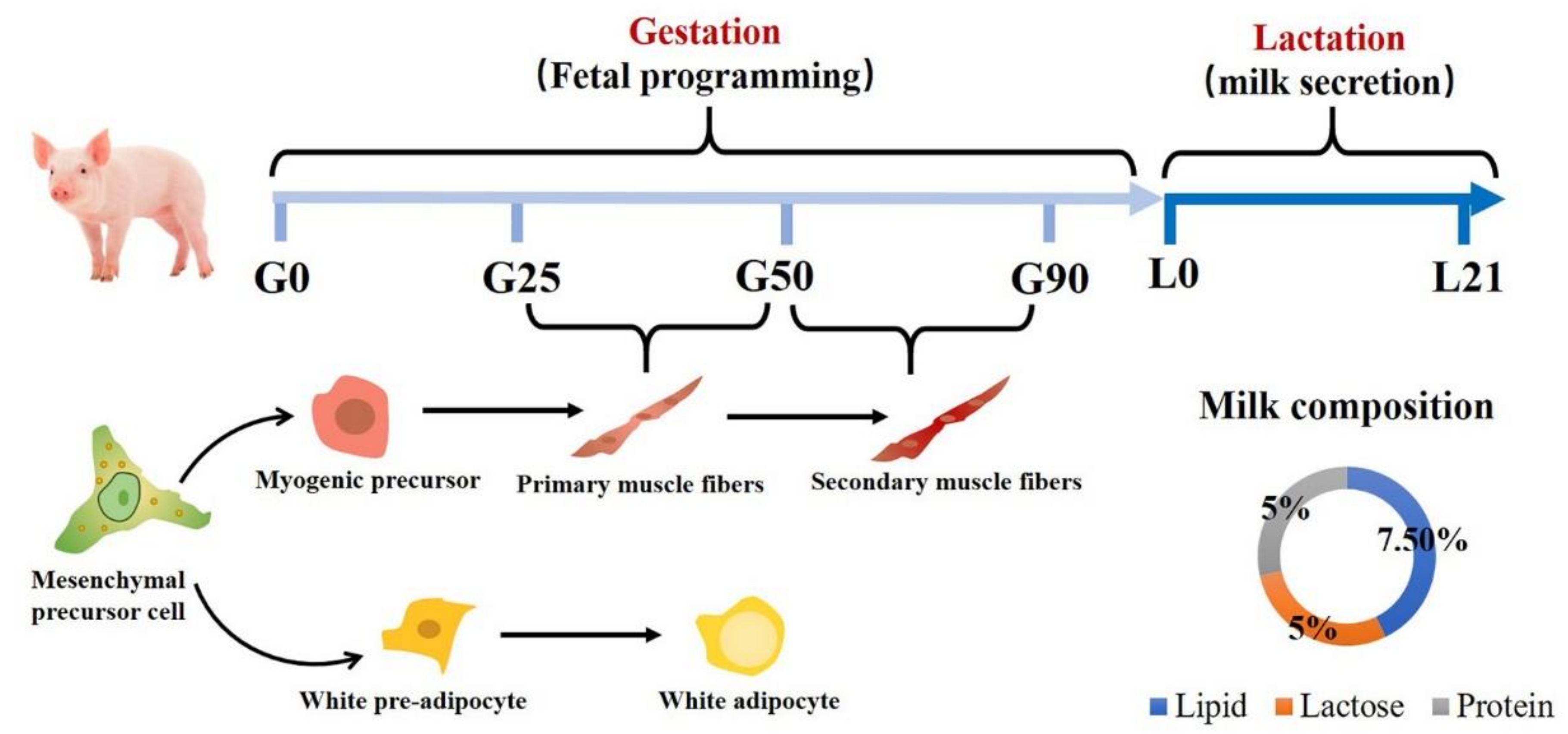
2. Muscle and Adipose Tissue Formation of Porcine Fetus
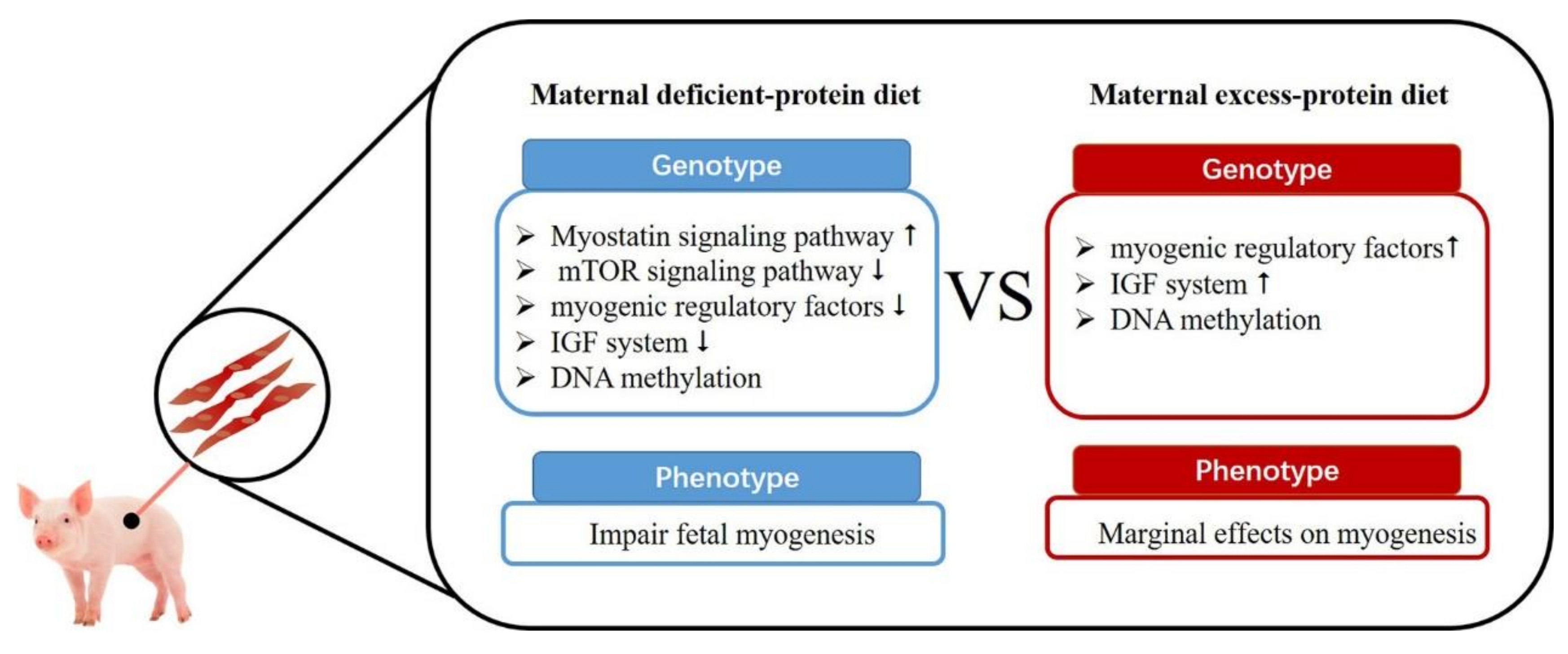
3. Maternal Protein Level on Fetal Skeletal Muscle Development
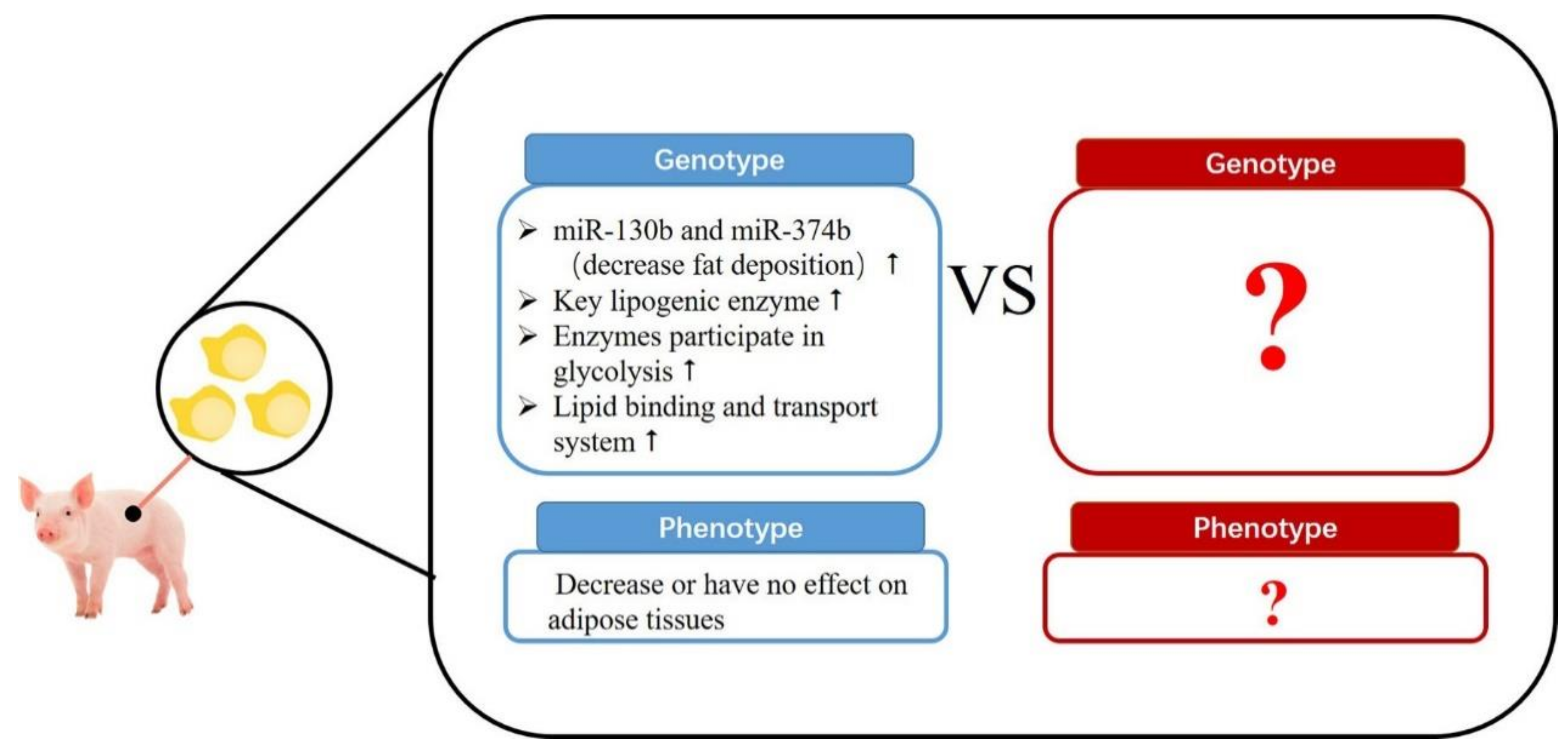
4. Maternal Protein Level on Adipose Tissue Development
5. Maternal Protein Level on Sow Lactation
6. Functional Amino Acids and Mammary Gland Development and Lactation
7. Conclusions and Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Belkacemi, L.; Nelson, D.M.; Desai, M.; Ross, M.G. Maternal undernutrition influences placental-fetal development. Biol. Reprod. 2010, 83, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Godfrey, K.M.; Barker, D.J. Fetal programming and adult health. Public Health Nutr. 2001, 4, 611–624. [Google Scholar] [CrossRef] [PubMed]
- Senger, P.L. Pathways to Pregnancy and Parturition; Current Conceptions, Inc.: Pullman, WA, USA, 2004. [Google Scholar]
- Abbott, D.H.; Padmanabhan, V.; Dumesic, D.A. Contributions of androgen and estrogen to fetal programming of ovarian dysfunction. Reprod. Biol. Endocrinol. 2006, 4, 17. [Google Scholar] [CrossRef] [PubMed]
- Wigmore, P.; Stickland, N. Muscle development in large and small pig fetuses. J. Anat. 1983, 137, 235. [Google Scholar] [PubMed]
- Swatland, H.; Cassens, R. Muscle growth: The problem of muscle fibers with an intrafascicular termination. J. Anim. Sci. 1972, 35, 336–344. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Chen, F.; Zhang, Y.; Lv, Y.; Heng, J.; Min, T.; Li, L.; Guan, W. Recent progress of porcine milk components and mammary gland function. J. Anim. Sci. Biotechnol. 2018, 9, 77. [Google Scholar] [CrossRef] [PubMed]
- Uruakpa, F.; Ismond, M.; Akobundu, E. Colostrum and its benefits: A review. Nutr. Res. 2002, 22, 755–767. [Google Scholar] [CrossRef]
- Blum, J.W.; Hammon, H. Colostrum effects on the gastrointestinal tract, and on nutritional, endocrine and metabolic parameters in neonatal calves. Livest. Prod. Sci. 2000, 66, 151–159. [Google Scholar] [CrossRef]
- Playford, R.J.; Macdonald, C.E.; Johnson, W.S. Colostrum and milk-derived peptide growth factors for the treatment of gastrointestinal disorders. Am. J. Clin. Nutr. 2000, 72, 5–14. [Google Scholar] [CrossRef] [Green Version]
- Burrin, D.G.; Shulman, R.J.; Reeds, P.J.; Davis, T.A.; Gravitt, K.R. Porcine colostrum and milk stimulate visceral organ and skeletal muscle protein synthesis in neonatal piglets. J. Nutr. 1992, 122, 1205–1213. [Google Scholar] [CrossRef]
- Wu, G.; Pond, W.G.; Ott, T.; Bazer, F.W. Maternal dietary protein deficiency decreases amino acid concentrations in fetal plasma and allantoic fluid of pigs. J. Nutr. 1998, 128, 894–902. [Google Scholar] [CrossRef] [PubMed]
- Du, M.; Tong, J.; Zhao, J.; Underwood, K.; Zhu, M.; Ford, S.; Nathanielsz, P. Fetal programming of skeletal muscle development in ruminant animals. J. Anim. Sci. 2010, 88, E51–E60. [Google Scholar] [CrossRef] [PubMed]
- Manikkam, M.; Crespi, E.J.; Doop, D.D.; Herkimer, C.; Lee, J.S.; Yu, S.; Brown, M.B.; Foster, D.L.; Padmanabhan, V. Fetal programming: Prenatal testosterone excess leads to fetal growth retardation and postnatal catch-up growth in sheep. Endocrinology 2004, 145, 790–798. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.; Wu, Z.; Dai, Z.; Wang, X.; Li, J.; Wang, B.; Wu, G. Fetal and neonatal programming of postnatal growth and feed efficiency in swine. J. Anim. Sci. Biotechnol. 2017, 8, 42. [Google Scholar] [CrossRef]
- Karunaratne, J.; Ashton, C.; Stickland, N. Fetal programming of fat and collagen in porcine skeletal muscles. J. Anat. 2005, 207, 763–768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dwyer, C.M.; Stickland, N.C.; Fletcher, J.M. The influence of maternal nutrition on muscle fiber number development in the porcine fetus and on subsequent postnatal growth. J. Anim. Sci. 1994, 72, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Rehfeldt, C.; Kuhn, G. Consequences of birth weight for postnatal growth performance and carcass quality in pigs as related to myogenesis. J. Anim. Sci. 2006, 84, E113–E123. [Google Scholar] [CrossRef]
- Sordella, R.; Jiang, W.; Chen, G.-C.; Curto, M.; Settleman, J. Modulation of Rho GTPase signaling regulates a switch between adipogenesis and myogenesis. Cell 2003, 113, 147–158. [Google Scholar] [CrossRef]
- Tong, J.F.; Yan, X.; Zhu, M.J.; Ford, S.P.; Nathanielsz, P.W.; Du, M. Maternal obesity downregulates myogenesis and β-catenin signaling in fetal skeletal muscle. Am. J. Physiol.-Endocrinol. Metab. 2009, 296, E917–E924. [Google Scholar] [CrossRef] [Green Version]
- Kablar, B.; Krastel, K.; Tajbakhsh, S.; Rudnicki, M.A. Myf5 and MyoD activation define independent myogenic compartments during embryonic development. Dev. Biol. 2003, 258, 307–318. [Google Scholar] [CrossRef] [Green Version]
- Hauser, N.; Mourot, J.; De Clercq, L.; Genart, C.; Remacle, C. The cellularity of developing adipose tissues in Pietrain and Meishan pigs. Reprod. Nutr. Dev. 1997, 37, 617–625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osgerby, J.; Wathes, D.; Howard, D.; Gadd, T. The effect of maternal undernutrition on ovine fetal growth. J. Endocrinol. 2002, 173, 131–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, M.-J.; Ford, S.P.; Nathanielsz, P.W.; Du, M. Effect of maternal nutrient restriction in sheep on the development of fetal skeletal muscle. Biol. Reprod. 2004, 71, 1968–1973. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Li, P.; Wang, J.; Li, X.; Gao, H.; Yin, Y.; Hou, Y.; Wu, G. Catabolism of nutritionally essential amino acids in developing porcine enterocytes. Amino Acids 2009, 37, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Rehfeldt, C.; Lang, I.; Görs, S.; Hennig, U.; Kalbe, C.; Stabenow, B.; Brüssow, K.-P.; Pfuhl, R.; Bellmann, O.; Nürnberg, G. Limited and excess dietary protein during gestation affects growth and compositional traits in gilts and impairs offspring fetal growth. J. Anim. Sci. 2011, 89, 329–341. [Google Scholar] [CrossRef]
- Rehfeldt, C.; Stabenow, B.; Pfuhl, R.; Block, J.; Nürnberg, G.; Otten, W.; Metges, C.; Kalbe, C. Effects of limited and excess protein intakes of pregnant gilts on carcass quality and cellular properties of skeletal muscle and subcutaneous adipose tissue in fattening pigs. J. Anim. Sci. 2012, 90, 184–196. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Pan, S.; Li, X.; Sun, Q.; Yang, X.; Zhao, R. Maternal low-protein diet affects myostatin signaling and protein synthesis in skeletal muscle of offspring piglets at weaning stage. Eur. J. Nutr. 2015, 54, 971–979. [Google Scholar] [CrossRef] [PubMed]
- Miller, L.; Garwood, V.; Judge, M. Factors affecting porcine muscle fiber type, diameter and number. J. Anim. Sci. 1975, 41, 66–77. [Google Scholar] [CrossRef]
- Rehfeldt, C.; Lefaucheur, L.; Block, J.; Stabenow, B.; Pfuhl, R.; Otten, W.; Metges, C.C.; Kalbe, C. Limited and excess protein intake of pregnant gilts differently affects body composition and cellularity of skeletal muscle and subcutaneous adipose tissue of newborn and weanling piglets. Eur. J. Nutr. 2012, 51, 151–165. [Google Scholar] [CrossRef]
- Mallinson, J.E.; Sculley, D.V.; Craigon, J.; Plant, R.; Langley-Evans, S.C.; Brameld, J.M. Fetal exposure to a maternal low-protein diet during mid-gestation results in muscle-specific effects on fibre type composition in young rats. Br. J. Nutr. 2007, 98, 292–299. [Google Scholar] [CrossRef]
- Kalbe, C.; Lösel, D.; Block, J.; Lefaucheur, L.; Brüssow, K.-P.; Bellmann, O.; Pfuhl, R.; Puppe, B.; Otten, W.; Metges, C. Moderate high or low maternal protein diets change gene expression but not the phenotype of skeletal muscle from porcine fetuses. Domest. Anim. Endocrinol. 2017, 58, 63–75. [Google Scholar] [CrossRef] [PubMed]
- Pan, S.; Jia, Y.; Yang, X.; Cai, D.; Liu, Z.; Song, H.; Zhao, R. Amino acid starvation-induced autophagy is involved in reduced subcutaneous fat deposition in weaning piglets derived from sows fed low-protein diet during gestation and lactation. Eur. J. Nutr. 2018, 57, 991–1001. [Google Scholar] [CrossRef] [PubMed]
- Altmann, S.; Murani, E.; Schwerin, M.; Metges, C.C.; Wimmers, K.; Ponsuksili, S. Maternal dietary protein restriction and excess affects offspring gene expression and methylation of non-SMC subunits of condensin I in liver and skeletal muscle. Epigenetics 2012, 7, 239–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Wang, J.; Li, R.; Yang, X.; Sun, Q.; Albrecht, E.; Zhao, R. Maternal dietary protein affects transcriptional regulation of myostatin gene distinctively at weaning and finishing stages in skeletal muscle of Meishan pigs. Epigenetics 2011, 6, 899–907. [Google Scholar] [CrossRef] [Green Version]
- McPherron, A.C.; Lawler, A.M.; Lee, S.-J. Regulation of skeletal muscle mass in mice by a new TGF-p superfamily member. Nature 1997, 387, 83. [Google Scholar] [CrossRef] [PubMed]
- Ríos, R.; Carneiro, I.; Arce, V.M.; Devesa, J. Myostatin is an inhibitor of myogenic differentiation. Am. J. Physiol.-Cell Physiol. 2002, 282, C993–C999. [Google Scholar]
- Jia, Y.; Gao, G.; Song, H.; Cai, D.; Yang, X.; Zhao, R. Low-protein diet fed to crossbred sows during pregnancy and lactation enhances myostatin gene expression through epigenetic regulation in skeletal muscle of weaning piglets. Eur. J. Nutr. 2016, 55, 1307–1314. [Google Scholar] [CrossRef]
- Ropka-Molik, K.; Eckert, R.; Piórkowska, K. The expression pattern of myogenic regulatory factors MyoD, Myf6 and Pax7 in postnatal porcine skeletal muscles. Gene Expr. Patterns 2011, 11, 79–83. [Google Scholar] [CrossRef]
- Cabeço, L.C.; Budri, P.E.; Baroni, M.; Castan, E.P.; Carani, F.R.; de Souza, P.A.T.; Boer, P.A.; Matheus, S.M.M.; Dal-Pai-Silva, M. Maternal protein restriction induce skeletal muscle changes without altering the MRFs MyoD and myogenin expression in offspring. J. Mol. Histol. 2012, 43, 461–471. [Google Scholar] [CrossRef]
- Florini, J.R.; Ewton, D.Z.; Coolican, S.A. Growth hormone and the insulin-like growth factor system in myogenesis. Endocr. Rev. 1996, 17, 481–517. [Google Scholar]
- Perry, V.; Norman, S.; Daniel, R.; Owens, P.; Grant, P.; Doogan, V. Insulin-like growth factor levels during pregnancy in the cow are affected by protein supplementation in the maternal diet. Anim. Reprod. Sci. 2002, 72, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Brameld, J.; Mostyn, A.; Dandrea, J.; Stephenson, T.; Dawson, J.; Buttery, P.; Symonds, M. Maternal nutrition alters the expression of insulin-like growth factors in fetal sheep liver and skeletal muscle. J. Endocrinol. 2000, 167, 429–437. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.-Z.; Sun, J.-J.; Zhang, L.-Z.; Li, C.-J.; Womack, J.E.; Li, Z.-J.; Lan, X.-Y.; Lei, C.-Z.; Zhang, C.-L.; Zhao, X. Genome-wide DNA methylation profiles and their relationships with mRNA and the microRNA transcriptome in bovine muscle tissue (Bos taurine). Sci. Rep. 2014, 4, 6546. [Google Scholar] [CrossRef] [PubMed]
- Kulis, M.; Esteller, M. DNA methylation and cancer. Adv. Genet. 2010, 70, 27–56. [Google Scholar] [PubMed]
- Lillycrop, K.A.; Slater-Jefferies, J.L.; Hanson, M.A.; Godfrey, K.M.; Jackson, A.A.; Burdge, G.C. Induction of altered epigenetic regulation of the hepatic glucocorticoid receptor in the offspring of rats fed a protein-restricted diet during pregnancy suggests that reduced DNA methyltransferase-1 expression is involved in impaired DNA methylation and changes in histone modifications. Br. J. Nutr. 2007, 97, 1064–1073. [Google Scholar] [PubMed]
- Lan, X.; Cretney, E.C.; Kropp, J.; Khateeb, K.; Berg, M.; Peñagaricano, F.; Magness, R.; Radunz, A.; Khatib, H. Maternal diet during pregnancy induces gene expression and DNA methylation changes in fetal tissues in sheep. Front. Genet. 2013, 4, 49. [Google Scholar] [CrossRef] [PubMed]
- Budge, H.; Gnanalingham, M.G.; Gardner, D.S.; Mostyn, A.; Stephenson, T.; Symonds, M.E. Maternal nutritional programming of fetal adipose tissue development: Long-term consequences for later obesity. Birth Defects Res. C Embryo Today Rev. 2005, 75, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Zambrano, E.; Bautista, C.; Deas, M.; Martinez-Samayoa, P.; Gonzalez-Zamorano, M.; Ledesma, H.; Morales, J.; Larrea, F.; Nathanielsz, P. A low maternal protein diet during pregnancy and lactation has sex-and window of exposure-specific effects on offspring growth and food intake, glucose metabolism and serum leptin in the rat. J. Physiol. 2006, 571, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Pan, S.; Zheng, Y.; Zhao, R.; Yang, X. MicroRNA-130b and microRNA-374b mediate the effect of maternal dietary protein on offspring lipid metabolism in Meishan pigs. Br. J. Nutr. 2013, 109, 1731–1738. [Google Scholar] [CrossRef]
- Sarr, O.; Louveau, I.; Kalbe, C.; Metges, C.; Rehfeldt, C.; Gondret, F. Prenatal exposure to maternal low or high protein diets induces modest changes in the adipose tissue proteome of newborn piglets. J. Anim. Sci. 2010, 88, 1626–1641. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Ren, L.; Chen, L.; Zhang, X.; Cheng, M.; Li, W.; Zhang, Y.; Gao, S. Differential expression of lipid metabolism related genes in porcine muscle tissue leading to different intramuscular fat deposition. Lipids 2009, 44, 1029. [Google Scholar] [CrossRef] [PubMed]
- Bellinger, L.; Lilley, C.; Langley-Evans, S.C. Prenatal exposure to a maternal low-protein diet programmes a preference for high-fat foods in the young adult rat. Br. J. Nutr. 2004, 92, 513–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellinger, L.; Langley-Evans, S.C. Fetal programming of appetite by exposure to a maternal low-protein diet in the rat. Clin. Sci. 2005, 109, 413–420. [Google Scholar] [CrossRef]
- Himms-Hagen, J. Obesity may be due to a malfunctioning of brown fat. Can. Med. Assoc. J. 1979, 121, 1361. [Google Scholar] [PubMed]
- Symonds, M.E.; Pearce, S.; Bispham, J.; Gardner, D.S.; Stephenson, T. Timing of nutrient restriction and programming of fetal adipose tissue development. Proc. Nutr. Soc. 2004, 63, 397–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bispham, J.; Gardner, D.; Gnanalingham, M.; Stephenson, T.; Symonds, M.; Budge, H. Maternal nutritional programming of fetal adipose tissue development: Differential effects on messenger ribonucleic acid abundance for uncoupling proteins and peroxisome proliferator-activated and prolactin receptors. Endocrinology 2005, 146, 3943–3949. [Google Scholar] [CrossRef]
- Dauncey, M.; Wooding, F.; Ingram, D. Evidence for the presence of brown adipose tissue in the pig. Res. Vet. Sci. 1981, 31, 76–81. [Google Scholar] [CrossRef]
- Trayhurn, P.; Temple, N.J.; Aerde, J.V. Evidence from immunoblotting studies on uncoupling protein that brown adipose tissue is not present in the domestic pig. Can. J. Physiol. Pharmacol. 1989, 67, 1480–1485. [Google Scholar] [CrossRef]
- Averette, L.A.; Odle, J.; Monaco, M.H.; Donovan, S.M. Dietary fat during pregnancy and lactation increases milk fat and insulin-like growth factor I concentrations and improves neonatal growth rates in swine. J. Nutr. 1999, 129, 2123–2129. [Google Scholar] [CrossRef]
- Højgaard, C.; Bruun, T.; Theil, P. Impact of Dietary Protein on Yield and Composition of Sow Milk. J. Anim. Sci. 2018, 96, 170. [Google Scholar] [CrossRef]
- Strathe, A.V.; Bruun, T.S.; Geertsen, N.; Zerrahn, J.-E.; Hansen, C.F. Increased dietary protein levels during lactation improved sow and litter performance. Anim. Feed Sci. Technol. 2017, 232, 169–181. [Google Scholar] [CrossRef]
- Huber, L.; de Lange, C.; Krogh, U.; Chamberlin, D.; Trottier, N. Impact of feeding reduced crude protein diets to lactating sows on nitrogen utilization. J. Anim. Sci. 2015, 93, 5254–5264. [Google Scholar] [CrossRef] [PubMed]
- Manjarin, R.; Zamora, V.; Wu, G.; Steibel, J.; Kirkwood, R.; Taylor, N.; Wils-Plotz, E.; Trifilo, K.; Trottier, N. Effect of amino acids supply in reduced crude protein diets on performance, efficiency of mammary uptake, and transporter gene expression in lactating sows. J. Anim. Sci. 2012, 90, 3088–3100. [Google Scholar] [CrossRef] [PubMed]
- Wu, G. Amino acids: Metabolism, functions, and nutrition. Amino Acids 2009, 37, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Guan, X.; Pettigrew, J.; Ku, P.; Ames, N.; Bequette, B.; Trottier, N. Dietary protein concentration affects plasma arteriovenous difference of amino acids across the porcine mammary gland. J. Anim. Sci. 2004, 82, 2953–2963. [Google Scholar] [CrossRef] [PubMed]
- Jang, Y.; Jang, S.; Kim, D.; Oh, H.; Kim, Y. Effects of dietary protein levels for gestating gilts on reproductive performance, blood metabolites and milk composition. Asian-Australas. J. Anim. Sci. 2014, 27, 83. [Google Scholar] [CrossRef] [PubMed]
- Renaudeau, D.; Noblet, J. Effects of exposure to high ambient temperature and dietary protein level on sow milk production and performance of piglets. J. Anim. Sci. 2001, 79, 1540–1548. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Hurley, W.; Han, I.; Easter, R. Changes in tissue composition associated with mammary gland growth during lactation in sows. J. Anim. Sci. 1999, 77, 2510–2516. [Google Scholar] [CrossRef] [PubMed]
- Trottier, N.; Shipley, C.; Easter, R. Plasma amino acid uptake by the mammary gland of the lactating sow. J. Anim. Sci. 1997, 75, 1266–1278. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.W.; Wu, G. Regulatory role for amino acids in mammary gland growth and milk synthesis. Amino Acids 2009, 37, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.W.; McPherson, R.L.; Wu, G. Dietary arginine supplementation enhances the growth of milk-fed young pigs. J. Nutr. 2004, 134, 625–630. [Google Scholar] [CrossRef] [PubMed]
- Mateo, R.; Wu, G.; Moon, H.; Carroll, J.; Kim, S. Effects of dietary arginine supplementation during gestation and lactation on the performance of lactating primiparous sows and nursing piglets. J. Anim. Sci. 2008, 86, 827–835. [Google Scholar] [CrossRef] [PubMed]
- O’Quinn, P.; Knabe, D.; Wu, G. Arginine catabolism in lactating porcine mammary tissue. J. Anim. Sci. 2002, 80, 467–474. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Hu, S.; Bannai, M.; Wu, G. L-Arginine regulates protein turnover in porcine mammary epithelial cells to enhance milk protein synthesis. Amino Acids 2018, 50, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Holanda, D.; Marcolla, C.; Guimarães, S.; Neves, M.; Hausman, G.; Duarte, M.; Abreu, M.; Saraiva, A. Dietary L-arginine supplementation increased mammary gland vascularity of lactating sows. Animal 2018, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Krogh, U.; Oksbjerg, N.; Purup, S.; Ramaekers, P.; Theil, P.K. Colostrum and milk production in multiparous sows fed supplementary arginine during gestation and lactation. J. Anim. Sci. 2016, 94, 22–25. [Google Scholar] [CrossRef]
- Krogh, U.; Oksbjerg, N.; Storm, A.C.; Feyera, T.; Theil, P.K. Mammary nutrient uptake in multiparous sows fed supplementary arginine during gestation and lactation. J. Anim. Sci. 2017, 95, 2517–2532. [Google Scholar] [CrossRef]
- Bass, B.; Bradley, C.; Johnson, Z.; Zier-Rush, C.; Boyd, R.; Usry, J.; Maxwell, C.; Frank, J. Influence of dietary L-arginine supplementation of sows during late pregnancy on piglet birth weight and sow and litter performance during lactation. J. Anim. Sci. 2017, 95, 248–256. [Google Scholar] [Green Version]
- Li, P.; Knabe, D.A.; Kim, S.W.; Lynch, C.J.; Hutson, S.M.; Wu, G. Lactating porcine mammary tissue catabolizes branched-chain amino acids for glutamine and aspartate synthesis. J. Nutr. 2006, 139, 1502–1509. [Google Scholar] [CrossRef]
- Appuhamy, J.R.N.; Knoebel, N.A.; Nayananjalie, W.D.; Escobar, J.; Hanigan, M.D. Isoleucine and Leucine Independently Regulate mTOR Signaling and Protein Synthesis in MAC-T Cells and Bovine Mammary Tissue Slices, 2. J. Nutr. 2012, 142, 484–491. [Google Scholar] [CrossRef]
- Lei, J.; Feng, D.; Zhang, Y.; Zhao, F.-Q.; Wu, Z.; San Gabriel, A.; Fujishima, Y.; Uneyama, H.; Wu, G. Nutritional and regulatory role of branched-chain amino acids in lactation. Front. Biosci. 2012, 17, 2725–2739. [Google Scholar] [CrossRef]
- Rezaei, R. Nutritional and Regulatory Roles for Branched-Chain Amino Acids in Milk Production by Lactating Sows. Ph.D. Thesis, Texas A&M University, College Station, TX, USA, 2015. [Google Scholar]
- Nissen, S.; Faidley, T.D.; Zimmerman, D.R.; Izard, R.; Fisher, C.T. Colostral milk fat percentage and pig performance are enhanced by feeding the leucine metabolite β-hydroxy-β-methyl butyrate to sows. J. Anim. Sci. 1994, 72, 2331–2337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flummer, C.; Theil, P.K. Effect of β-hydroxy β-methyl butyrate supplementation of sows in late gestation and lactation on sow production of colostrum and milk and piglet performance. J. Anim. Sci. 2012, 90, 372–374. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Zeng, Z.; Xu, X.; Tian, Q.; Ma, X.; Long, S.; Piao, M.; Cheng, Z.; Piao, X. Effects of the standardized ileal digestible valine: Lysine ratio on performance, milk composition and plasma indices of lactating sows. Anim. Sci. J. 2017, 88, 1082–1092. [Google Scholar] [CrossRef] [PubMed]
- Strathe, A.V.; Bruun, T.S.; Zerrahn, J.-E.; Tauson, A.-H.; Hansen, C.F. The effect of increasing the dietary valine-to-lysine ratio on sow metabolism, milk production, and litter growth. J. Anim. Sci. 2016, 94, 155–164. [Google Scholar] [CrossRef]
- Richert, B.; Goodband, R.; Tokach, M.; Nelssen, J. Increasing valine, isoleucine, and total branched-chain amino acids for lactating sows. J. Anim. Sci. 1997, 75, 2117–2128. [Google Scholar] [CrossRef]
- Rousselow, D.; Speer, V.; Haught, D. Leucine requirement of the lactating sow. J. Anim. Sci. 1979, 49, 498–506. [Google Scholar] [CrossRef]
- Dunshea, F.R.; Bauman, D.E.; Nugent, E.A.; Kerton, D.J.; King, R.H.; McCauley, I. Hyperinsulinaemia, supplemental protein and branched-chain amino acids when combined can increase milk protein yield in lactating sows. Br. J. Nutr. 2005, 93, 325–332. [Google Scholar] [CrossRef] [Green Version]
- Appuhamy, J.; Knapp, J.; Becvar, O.; Escobar, J.; Hanigan, M. Effects of jugular-infused lysine, methionine, and branched-chain amino acids on milk protein synthesis in high-producing dairy cows. J. Dairy Sci. 2011, 94, 1952–1960. [Google Scholar] [CrossRef]
- Moser, S.; Tokach, M.; Dritz, S.; Goodband, R.; Nelssen, J.; Loughmiller, J. The effects of branched-chain amino acids on sow and litter performance. J. Anim. Sci. 2000, 78, 658–667. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Animals | Experimental Design | Results | Conclusion | References |
|---|---|---|---|---|
| 14 primiparous pure-bred Meishan gilt sbody weight: 36.1 ± 1.8 kg | Digestible energy 13 MJ/kg Control treatment: 12.2% crude protein (CP) (gestation) and 14% CP (lactation) Low protein treatment: 6.1% CP (gestation) and 6.9% CP (lactation) | Muscle protein synthesis: p70S6K and eIF4E ↓ Negative regulator of skeletal muscle development and growth ↑ Muscle weight: Longissimus dorsi muscles ↓ | Maternal low protein diet throughout gestation and lactation causes retardation in muscle hypertrophy and protein synthesis | [28] |
| 16 Landrace × Yorkshire crossbred sows in the second parity | Digestible energy:13 MJ/kg (gestation),14.39 MJ/kg (lactation) Control treatment: 15% (CP, gestation) and 18% CP (lactation) Low protein treatment: 7.5% (CP, gestation) and 9% CP (lactation) | Back fat thickness ↓ Restricted amino acid response (AAR) pathway: CHOP, IRE1α, PERK, ATF-6, XBP-1 and Bip, ATF4, and EIF2α↑ Autophagy-related genes: ATG7 and LC3 ↑ | Maternal low protein diet throughout gestation and lactation causes offspring reduced adipogenesis and increased lipolysis | [33] |
| 56 German Landrace gilts | Metabolic energy: 13.7 MJ/kg Adequate protein treatment: 12.1% CP High protein treatment: 30% CP; Low protein treatment, 6.5% CP | Total number of myofibers ↔ Myogenic regulatory factors in low protein treatment: MYOG, MRF4, IGF1, IGF1R ↓ Myogenic regulatory factors in high protein treatment: MYOD, MYOG, IGF1R, and IGFBP5 ↑ | Moderate high or low maternal protein diets change gene expression but not the phenotype of skeletal muscle from porcine fetuses | [32] |
| 47 pure German Landrace gilts | Metabolic energy: 13.7 ME/kg Adequate protein treatment: 12.1% CP High protein treatment: 30% CP; Low protein treatment, 6.5% CP | Percentage of muscle tissue in in HP in high protein treatment ↑ Primary and secondary myofibers in low protein diet treatment ↓ Subcutaneous adipose tissue mass in low protein diet treatment and high protein diet treatment ↓ | Both limited and excess protein supply retards fetal growth, but only limited protein supply impairs myogenesis, persistently restricts muscle growth potential, and favors obesity at infancy. | [30] |
| 24 primiparous German Landrace sows | Metabolic energy: 13.7 ME/kg Adequate protein treatment: 12.1% CP High protein treatment: 30% CP; Low protein treatment, 6.5% CP | Candidate genes of nutrient- dependent pre- and postnatal development: muscular expression of NCAPD2 (LP ↓), NCAPG (↔), NCAPH (LP ↓) Key genes of methionine metabolism: both HP and LP diet significant influence DNMT1 (HP ↑), DNMT3a (LP ↓) and MAT2B (HP ↑) | Maternal protein supply regulate condensin I subunit gene expression by methylation process and in turn may affect cell division in skeletal muscle tissue | [34] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, S.; Heng, J.; Song, H.; Zhang, Y.; Lin, X.; Tian, M.; Chen, F.; Guan, W. Role of Maternal Dietary Protein and Amino Acids on Fetal Programming, Early Neonatal Development, and Lactation in Swine. Animals 2019, 9, 19. https://doi.org/10.3390/ani9010019
Zhang S, Heng J, Song H, Zhang Y, Lin X, Tian M, Chen F, Guan W. Role of Maternal Dietary Protein and Amino Acids on Fetal Programming, Early Neonatal Development, and Lactation in Swine. Animals. 2019; 9(1):19. https://doi.org/10.3390/ani9010019
Chicago/Turabian StyleZhang, Shihai, Jinghui Heng, Hanqing Song, Yufeng Zhang, Xiaofeng Lin, Min Tian, Fang Chen, and Wutai Guan. 2019. "Role of Maternal Dietary Protein and Amino Acids on Fetal Programming, Early Neonatal Development, and Lactation in Swine" Animals 9, no. 1: 19. https://doi.org/10.3390/ani9010019
APA StyleZhang, S., Heng, J., Song, H., Zhang, Y., Lin, X., Tian, M., Chen, F., & Guan, W. (2019). Role of Maternal Dietary Protein and Amino Acids on Fetal Programming, Early Neonatal Development, and Lactation in Swine. Animals, 9(1), 19. https://doi.org/10.3390/ani9010019