Antibody to Heat Shock Protein 70 (HSP70) Inhibits Human T-Cell Lymphoptropic Virus Type I (HTLV-I) Production by Transformed Rabbit T-Cell Lines

{kind=link}
{kind=link}
Abstract
:1. Introduction
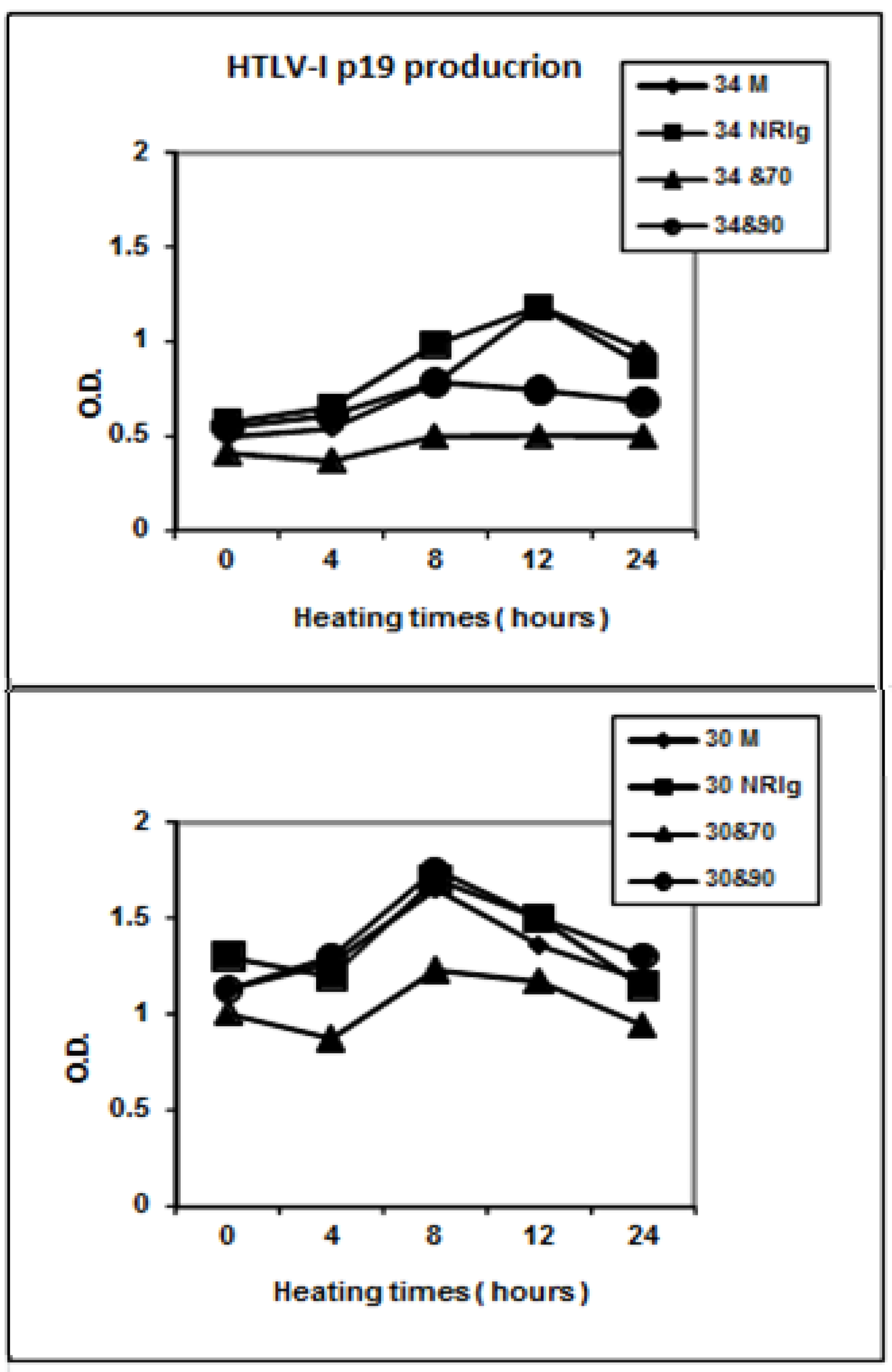
2. Results and Discussion
2.1. Expression of HSP on Cell Surface and Response to Heat Shock Treatment


2.2. Role of Anti-HSP 70 Antibodies on Virus Production
3. Experimental Section
3.1. Cell Lines and Treatment
3.2. Antibodies to Heat Shock Proteins
3.3. Enzyme-Linked Immunosorbent Assay (ELISA)
3.4. Western Blotting
3.5. Statistical Analysis
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Hsu, P.L.; Hsu, S.M. Abundance of heat shock proteins (hsp89, hsp60, and hsp27) in malignant cells of hodgkin’s disease. Canc. Res. 1998, 58, 5507–5513. [Google Scholar]
- Tamura, Y.; Tsuboi, N.; Sato, N.; Kikuchi, K. 70 kDa Heat shock cognate protein is a transformation-associated antigen and a possible target for the host’s anti-tumor immunity. J. Immunol. 1993, 151, 5516–5524. [Google Scholar]
- Ciocca, D.R; Calderwood, S.K. Heat shock proteins in cancer: Diagnostic, prognostic, predictive, and treatment implications. Cell Stress Chaperones 2005, 10, 86–103. [Google Scholar] [CrossRef]
- Martin, C.A.; Carsons, S.E.; Kowalewski, R.; Bernstein, D.; Valentino, M.; Santiago-Schwarz, F. Aberrant extracellular and dendritic cell (DC) surface expression of heat shock protein (hsp) 70 in the rheumatoid Joint: Possible mechanisms of hsp/DC-mediated cross-priming. J. Immunol. 2003, 171, 5736–5742. [Google Scholar]
- Macejak, D.G.; Luftig, R.B. Association of HSP70 with the adenovirus type 5 fiber protein in infected HEp-2 cells. Virology 1991, 180, 120–125. [Google Scholar] [CrossRef]
- Macejak, D.G.; Sarnow, P. Association of Heat Shock Protein 70 with Enterovirus Capsid Precursor P1 in Infected Human Cells. J. Virol. 1992, 66, 1520–1527. [Google Scholar]
- Poiesz, B.J.; Ruscetti, F.W.; Gazdar, A.F.; Bunn, P.A.; Mina, J.D.; Gallo, R.C. Detection and isolation of type C retrovirus particles from fresh and cultered lyphocytes of a patient with cutaneous T-cell lymphoma. Proc. Natl. Acad. Sci. USA 1980, 77, 7415–7419. [Google Scholar]
- Seto, A.; Kawanishi, M.; Matsuda, S.; Ogawa, K.; Eguchi, T.; Miyoshi, I. Induction of preleukemic stage of adult T cell leukemia like disease in rabbits. Jpn. J. Cancer Res. 1987, 78, 1150–1155. [Google Scholar]
- Sawasdikosol, S.; Hague, B.H.; Zhao, T.M.; Bowers, F.S.; Simpson, M.R.; Robinson, M.A.; Kindt, T.J. Selection of rabbit CD4-, CD8-T cell receptor-gamma/delta cells by in vitro transformation with human T lymphotropic virus type I. J. Exp. Med. 1993, 178, 1337–1345. [Google Scholar] [CrossRef]
- Simpson, R.M.; Zhao, T.M.; Hubbard, B.S.; Sawasdikosol, S.; Kindt, T.J. Experimental acute adult T cell leukemia lymphoma is associated with thymic atrophy in human T cell leukemia virus type I infection. Lab. Investig. 1996, 74, 696–710. [Google Scholar]
- Leno, M.; Simpson, R.M.; Bowers, F.S.; Kindt, T.J. Human T lymphocyte virus 1 from a leukemogenic cell line mediates in vivo and in vitro lymphocyte apoptosis. J. Exp. Med. 1995, 181, 1575–1580. [Google Scholar] [CrossRef]
- Zhao, T.M.; Robinson, M.A.; Sawasdikosol, S.; Simpson, R.M.; Kindt, T.J. Variation in HTLV-I sequences from rabbit cell lines with diverse in vivo effects. Virology 1993, 195, 271–274. [Google Scholar] [CrossRef]
- Zhao, T.M.; Robinson, M.A.; Bowers, F.S.; Kindt, T.J. Characterization of an infectious molecular clone of human T-cell leukemia virus type I. J. Virol. 1995, 69, 2024–2030. [Google Scholar]
- Andrews, J.M.; Oglesbee, M.J.; Trevino, A.V.; Guyot, D.J.; Newbound, G.C.; Lairmore, M.D. Enhanced human T-cell lymphotropic virus type I expression following induction of the cellular stress response. Virology 1995, 208, 816–820. [Google Scholar] [CrossRef]
- Andrews, J.M.; Newbound, G.C.; Oglesbee, M.; Brady, J.N.; Lairmore, M.D. The cellular stress response enhances human T-cell lymphotropic virus type 1 basal gene expression through the core promoter region of the long terminal repeat. J. Virol. 1997, 71, 741–745. [Google Scholar]
- Andrews, J.M.; Oglesbee, M.; Lairmore, M.D. The effect of the cellular stress response on human T-lymphotropic virus type I envelope protein expression. J. Gen. Virol. 1998, 79, 2905–2908. [Google Scholar]
- Chouchane, L.; Bowers, F.S.; Sawasdikosol, S.; Simpson, R.M.; Kindt, T.J. Heat-shock proteins expressed on the surface of human T cell leukemia virus type I-infected cell lines induce autoantibodies in rabbits. J. Infect. Dis. 1994, 169, 253–259. [Google Scholar] [CrossRef]
- Fang, D.; Haraguchi, Y.; Jinno, A.; Soda, Y.; Shimizu, N.; Hoshino, H. Heat shock cognate protein 70 is a cell fusion-enhancing factor but not an entry factor for human T-cell lymphotropic virus type I. Biochem. Biophys. Res. Commun. 1999, 2, 357–363. [Google Scholar]
- Hwang, Y.J.; Lee, S.P.; Kim, S.Y.; Choi, Y.H.; Kim, M.J.; Lee, C.H.; Lee, J.Y.; Kim, D.Y. Expression of heat shock protein 60 kDa is upregulated in cervical cancer. YonseiMed. J. 2009, 50, 399–406. [Google Scholar]
- Oglesbee, M.J.; Liu, Z.; Kenny, H.; Brooks, C.L. The highly inducible members of the 70 kDa family of heat shock proteins increase canine distemper virus polymerase activity. J. Gen. Virol. 1996, 77, 2125–2135. [Google Scholar] [CrossRef]
- Hirayama, E.; Atagi, H.; Hiraki, A.; ankim, J. Heat shock protein 70 is related to thermal inhibition of nuclear export of the influenza virus ribonucleoprotein complex. J. Virol. 2004, 78, 1263–1270. [Google Scholar] [CrossRef]
- Kocsis, J.; Prohászka, Z.; Bíró, A.; Füst, G.; Bánhegyi, D. Elevated levels of antibodies against 70 kDa heat shock proteins in the sera of patients with HIV infection. J. Med. Virol. 2003, 71, 480–482. [Google Scholar] [CrossRef]
- Mahana, W.; Samaan, A.; Kindt, T.J.; Simpson, M.R. Evidence for humoral and cellular reactivity against keratin and thyroglobulin in HTLV-I Infected Rabbits. Autoimmunity 2000, 32, 57–65. [Google Scholar] [CrossRef]
- Molvarec, A.; Derzsy, Z.; Kocsis, J.; Boze, T.; Nagy, B.; Balogh, K.; Makó, V.; Cervenak, L.; Mézes, M.; Karádi, I. et. al. Circulating anti-heat-shock-protein antibodies in normal pregnancy and preeclampsia. Cell Stress Chaperones 2009, 14, 491–498. [Google Scholar] [CrossRef]
- Ungar-Waron, H.; Brenner, J.; Paz, R.; Moalem, U.; Trainin, Z. Gamma delta T-lymphocytes and anti-heat shock protein reactivity in bovine leukaemia virus infected cattle. Vet. Immunol. Immunopathol. 1996, 2, 79–87. [Google Scholar]
- Torronteguy, C.; Frasson, A.; Zerwes, F.; Winnikov, E.; da Silva, V.D.; Ménoret, A.; Bonorino, C. Inducible heat shock protein 70 expression as a potential predictive marker of metastasis in breast tumors. Cell Stress Chaperones 2006, 11, 34–43. [Google Scholar] [CrossRef]
- Prud’homme, G.J.; Vanier, L.E. Cyclosporine, tolerance, and autoimmunity. Clin. Immunol. Immunopathol. 1993, 66, 185–192. [Google Scholar] [CrossRef]
- Iwakura, Y.; Saijo, S.; Kioka, Y.; Nakayama-Yamada, J.; Itagaki, K.; Tosu, M.; Asano, M.; Kanai, Y.; Kakimoto, K. Autoimmunity induction by human T cell leukemia virus type 1 in transgenic mice that develop chronic inflammatory arthropathy resembling rheumatoid arthritis in humans. J. Immunol. 1995, 155, 1588–1598. [Google Scholar]
- Williams, K.J.; Landgraf, B.E.; Whiting, N.L.; Zurlo, J. Correlation between the induction of heat shock protein 70 and enhanced viral reactivation in mammalian cells treated with ultraviolet light and heat shock. Cancer Res. 1989, 49, 2735–2742. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Fallouh, H.; Mahana, W. Antibody to Heat Shock Protein 70 (HSP70) Inhibits Human T-Cell Lymphoptropic Virus Type I (HTLV-I) Production by Transformed Rabbit T-Cell Lines. Toxins 2012, 4, 768-777. https://doi.org/10.3390/toxins4100768
Fallouh H, Mahana W. Antibody to Heat Shock Protein 70 (HSP70) Inhibits Human T-Cell Lymphoptropic Virus Type I (HTLV-I) Production by Transformed Rabbit T-Cell Lines. Toxins. 2012; 4(10):768-777. https://doi.org/10.3390/toxins4100768
Chicago/Turabian StyleFallouh, Hanan, and Wahib Mahana. 2012. "Antibody to Heat Shock Protein 70 (HSP70) Inhibits Human T-Cell Lymphoptropic Virus Type I (HTLV-I) Production by Transformed Rabbit T-Cell Lines" Toxins 4, no. 10: 768-777. https://doi.org/10.3390/toxins4100768
APA StyleFallouh, H., & Mahana, W. (2012). Antibody to Heat Shock Protein 70 (HSP70) Inhibits Human T-Cell Lymphoptropic Virus Type I (HTLV-I) Production by Transformed Rabbit T-Cell Lines. Toxins, 4(10), 768-777. https://doi.org/10.3390/toxins4100768