Trophic Patterns of Bighead Carp and Silver Carp Follow the Seasonality of Resource Availability

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
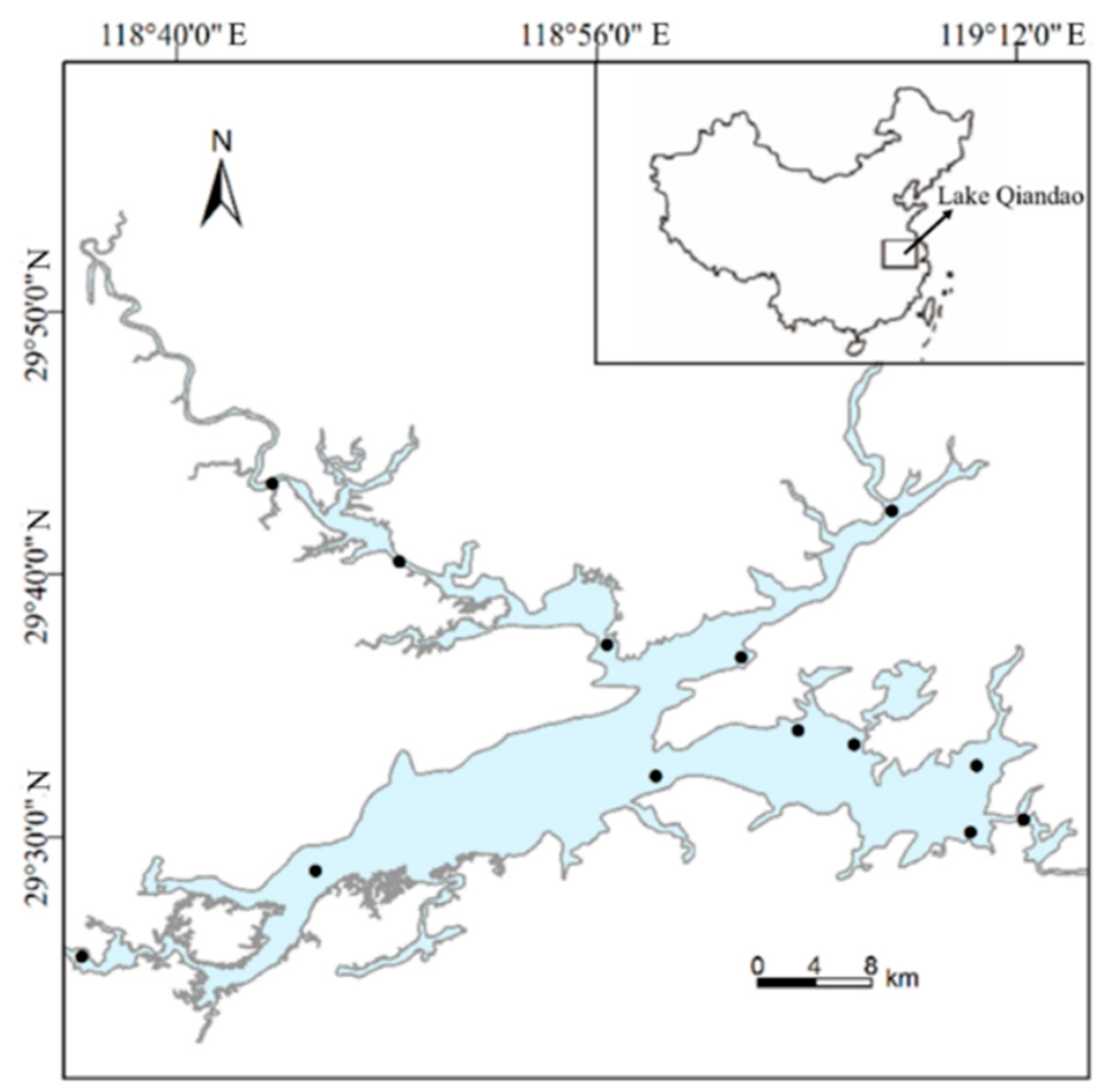
2.1. Study Area
2.2. Sample Processing and Treatment
2.3. Stable Isotope Analyses
2.4. Trophic Level Estimates and Trophic Niche Width
2.5. Structural Equation Model
2.6. Statistical Analyses
3. Result
3.1. The Seasonal Variations of Trophic Patterns of Filter-Feeding Fishes, Water Parameters and Densities of Plankton
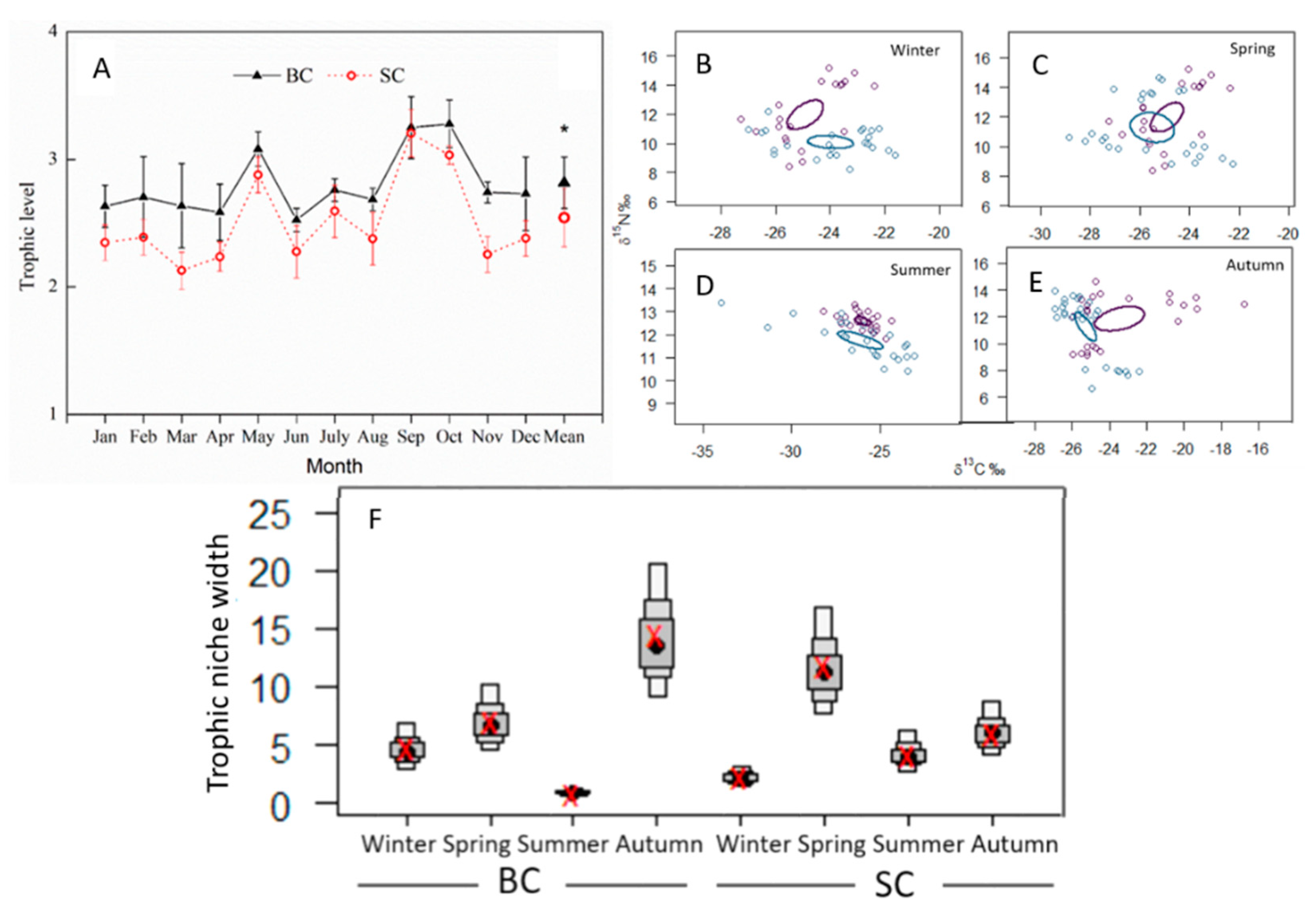
3.1.1. The Seasonal Variations of Trophic Patterns of Silver Carp and Bighead Carp
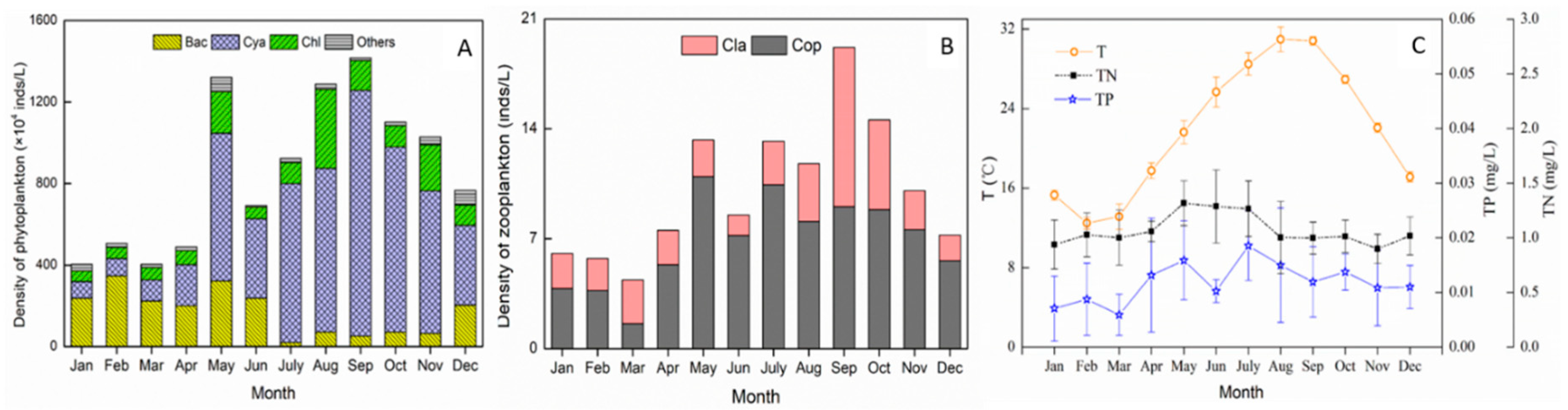
3.1.2. Seasonal Variations of Abiotic and Biological Parameters
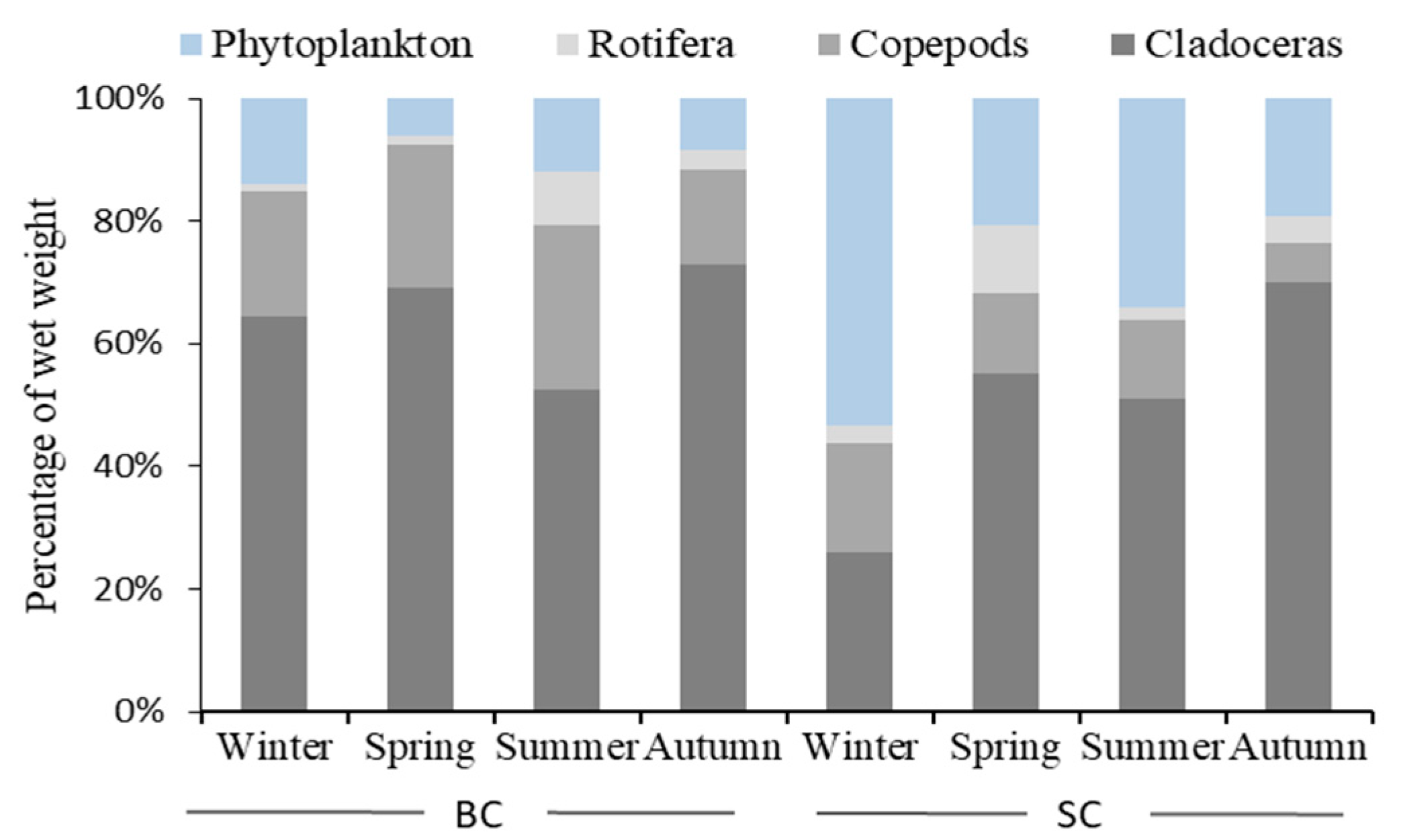
3.2. Dietary Compositions of Silver Carp and Bighead Carp Based on Gut Contents and Stable Isotopic Analysis
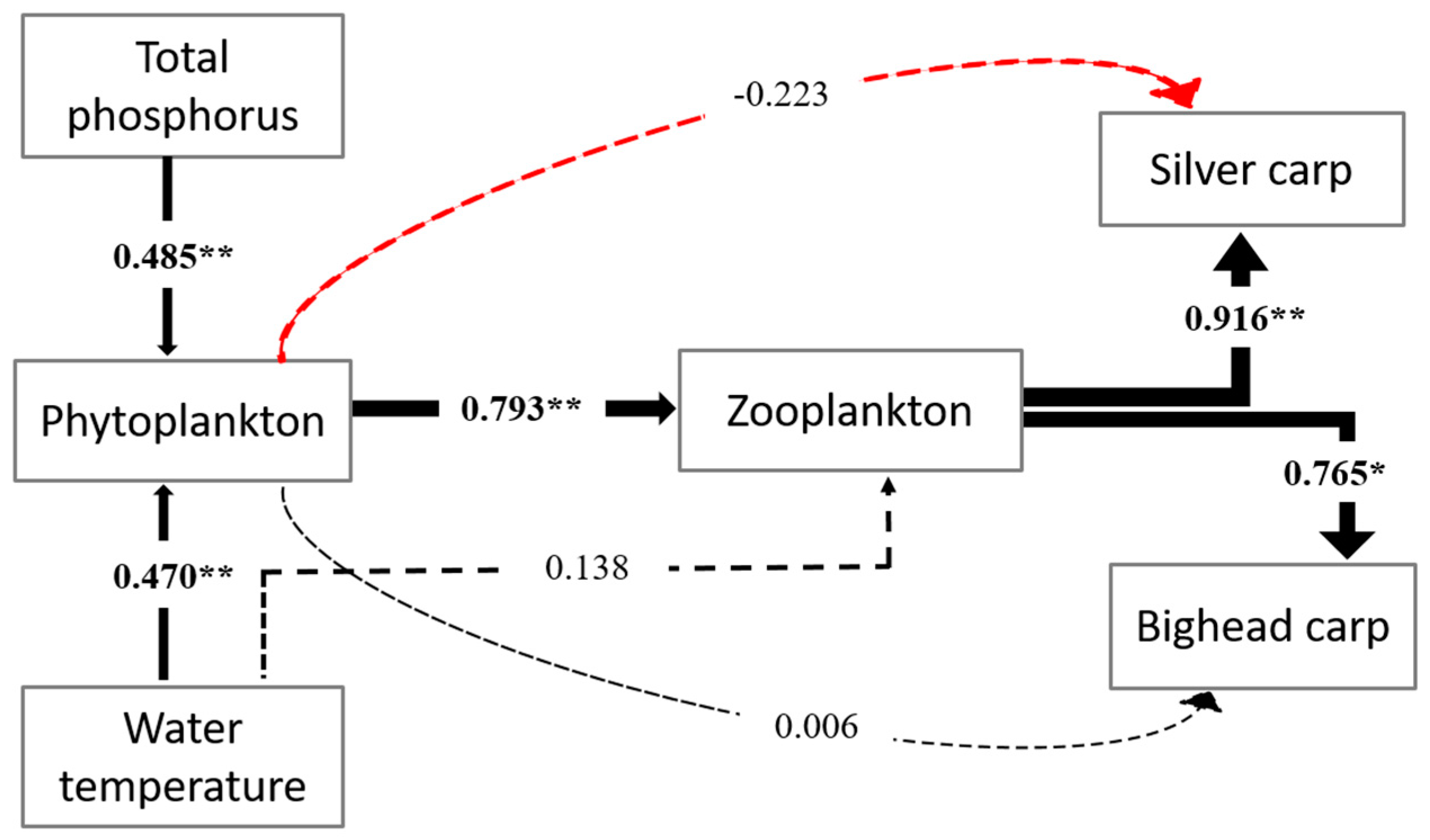
3.3. The Factors Affecting the Seasonal Variation of Trophic Levels of Fish
4. Discussion
4.1. Seasonal Trophic Patterns of the Bighead Carp and Silver Carp
4.2. The Effects of Bottom-Up and Top-Down Control on Trophic Levels of Filter-Feeding Fish
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Elton, C. Animal Ecology; Macmillan: New York, NY, USA, 1927. [Google Scholar]
- Zhou, Q.; Xie, P.; Xu, J.; Ke, Z.X.; Guo, L.G.; Cao, T. Seasonal variations in stable isotope ratios of two biomanipulation fishes and seston in a large pen culture in hypereutrophic Meiliang Bay, Lake Taihu. Ecol. Eng. 2009, 35, 1603–1609. [Google Scholar] [CrossRef]
- Hampton, S.E.; Galloway, A.W.; Powers, S.M.; Ozersky, T.; Woo, K.H.; Batt, R.D.; Labou, S.G.; O’Reilly, C.M.; Sharma, S.; Lottig, N.R. Ecology under lake ice. Ecol. Lett. 2017, 20, 98–111. [Google Scholar] [CrossRef] [PubMed]
- McMeans, B.C.; McCann, K.S.; Humphries, M.; Rooney, N.; Fisk, A.T. Food Web Structure in Temporally-Forced Ecosystems. Trends Ecol. Evol. 2015, 30, 662–672. [Google Scholar] [CrossRef] [PubMed]
- Sha, Y.C.; Su, G.H.; Zhang, P.Y.; Zhang, H.; Xu, J. Diverse dietary strategy of lake anchovy Coilia ectenes taihuensis in lakes with different trophic status. J. Ichthyol. 2015, 55, 866–873. [Google Scholar] [CrossRef]
- Kondoh, M. Foraging Adaptation and the Relationship Between Food-Web Complexity and Stability. Science 2003, 299, 1388–1391. [Google Scholar] [CrossRef] [PubMed]
- McCann, K.S.; Rasmussen, J.B.; Umbanhowar, J. The dynamics of spatially coupled food webs. Ecol. Lett. 2005, 8, 513–523. [Google Scholar] [CrossRef]
- Yao, X.; Huang, G.; Xie, P.; Xu, J. Trophic niche differences between coexisting omnivores silver carp and bighead carp in a pelagic food web. Ecol. Res. 2016, 31, 831–839. [Google Scholar] [CrossRef]
- Vander Zanden, M.J.; Shuter, B.J.; Lester, N.P.; Rasmussen, J.B. Within- and among-population variation in the trophic position of a pelagic predator, lake trout (Salvelinus namaycush). Can. J. Fish. Aquat. Sci. 2000, 57, 725–731. [Google Scholar] [CrossRef]
- Post, D.M.; Takimoto, G. Proximate structural mechanisms for variation in food-chain length. Oikos 2007, 116, 775–782. [Google Scholar] [CrossRef]
- Tunney, T.D.; McCann, K.S.; Lester, N.P.; Shuter, B.J. Food web expansion and contraction in response to changing environmental conditions. Nat. Commun. 2012, 3, 1105. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Guo, L.G.; Zhang, H.; Xu, J.; Hu, H.M.; Xue, T.; Luo, C.Q.; Yi, C.L.; Hu, Y.F.; De Silva, K.; et al. Spatial Variation in Trophic Structure of Dominant Fish Species in Lake Dongting, China during Dry Season. Water 2018, 10, 602. [Google Scholar] [CrossRef]
- Akin, S.; Winemiller, K.O. Seasonal variation in food web composition and structure in a temperate tidal estuary. Estuaries Coasts 2006, 29, 552–567. [Google Scholar] [CrossRef]
- Zhou, Q.; Xie, P.; Xu, J.; Ke, Z.; Guo, L. Growth and food availability of silver and bighead carps: Evidence from stable isotope and gut content analysis. Aquac. Res. 2009, 40, 1616–1625. [Google Scholar] [CrossRef]
- Ruiz-Cooley, R.I.; Gerrodette, T.; Fiedler, P.C.; Chivers, S.J.; Danil, K.; Ballance, L.T. Temporal variation in pelagic food chain length in response to environmental change. Sci. Adv. 2017, 3, e1701140. [Google Scholar] [CrossRef]
- Hellmann, C.; Wissel, B.; Winkelmann, C. Omnivores as seasonally important predators in a stream food web. Freshw. Sci. 2013, 32, 548–562. [Google Scholar] [CrossRef]
- Nakano, S.; Murakami, M. Reciprocal subsidies: Dynamic interdependence between terrestrial and aquatic food webs. Proc. Natl. Acad. Sci. USA 2001, 98, 166–170. [Google Scholar] [CrossRef] [Green Version]
- Soykan, C.U.; Sabo, J.L. Spatiotemporal food web dynamics along a desert riparian-upland transition. Ecography 2009, 32, 354–368. [Google Scholar] [CrossRef]
- Xie, P. Gut contents of silver carp, Hypophthalmichthys molitrix, and the disruption of a centric diatom, Cyclotella, on passage through the esophagus and intestine. Aquaculture 1999, 180, 295–305. [Google Scholar] [CrossRef]
- Yi, C.; Guo, L.; Ni, L.; Luo, C. Silver carp exhibited an enhanced ability of biomanipulation to control cyanobacteria bloom compared to bighead carp in hypereutrophic Lake Taihu mesocosms. Ecol. Eng. 2016, 89, 7–13. [Google Scholar] [CrossRef]
- Li, Y.; Xie, P.; Zhang, J.; Tao, M.; Deng, X. Effects of filter-feeding planktivorous fish and cyanobacteria on structuring the zooplankton community in the eastern plain lakes of China. Ecol. Eng. 2017, 99, 238–245. [Google Scholar] [CrossRef]
- McMeans, B.C.; Kadoya, T.; Pool, T.K.; Holtgrieve, G.W.; Lek, S.; Kong, H.; Winemiller, K.; Elliott, V.; Rooney, N.; Laffaille, P.; et al. Consumer trophic positions respond variably to seasonally fluctuating environments. Ecology 2019, 100, e02570. [Google Scholar] [CrossRef]
- Post, D.M. Using stable isotopes to estimate trophic position: Models, methods, and assumptions. Ecology 2002, 83, 703–718. [Google Scholar] [CrossRef]
- Xu, J.; Zhang, H.; Cai, Y.; García Molinos, J.; Zhang, M. Optimal response to habitat linkage of local fish diversity and mean trophic level. Limnol. Oceanogr. 2016, 61, 1438–1448. [Google Scholar] [CrossRef] [Green Version]
- Xie, P.; Liu, J. Practical success of biomanipulation using filter-feeding Fish to control cyanobacteria blooms: A synthesis of decades of research and application in a subtropical hypereutrophic lake. Sci. World J. 2001, 1, 337–356. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhang, Y.; Jeppesen, E.; Murphy, K.R.; Shi, K.; Liu, M.; Liu, X.; Zhu, G. Inflow rate-driven changes in the composition and dynamics of chromophoric dissolved organic matter in a large drinking water lake. Water Res. 2016, 100, 211–221. [Google Scholar] [CrossRef]
- Zhai, X.Y.; Zhang, Y.Y.; Wang, X.L.; Xia, J.; Liang, T. Non-point source pollution modelling using Soil and Water Assessment Tool and its parameter sensitivity analysis in Xin’anjiang catchment, China. Hydrol. Process 2014, 28, 1627–1640. [Google Scholar] [CrossRef]
- Sooknah, R.D.; Wilkie, A.C.J.E.E. Nutrient removal by floating aquatic macrophytes cultured in anaerobically digested flushed dairy manure wastewater. Ecol. Eng. 2004, 22, 27–42. [Google Scholar] [CrossRef]
- Yu, J.L.; Liu, Z.W.; He, H.; Zhen, W.; Guan, B.H.; Chen, F.Z.; Li, K.Y.; Zhong, P.; Teixeira-de Mello, F.; Jeppesen, E. Submerged macrophytes facilitate dominance of omnivorous fish in a subtropical shallow lake: Implications for lake restoration. Hydrobiologia 2016, 775, 97–107. [Google Scholar] [CrossRef]
- Wedchaparn, O.; Zhao, L.; Fan, Y.; He, D.; Zhang, H.; Ivan, L.N.; Liu, Q.; Ayisi, C.L. Comparison of the trophic niches between two planktivorous fishes in two large lakes using stable isotope analysis. Biochem. Syst. Ecol. 2016, 68, 148–155. [Google Scholar] [CrossRef]
- Huang, X.; Chen, W.; Cai, Q. Survey, Observation and Analysis of Lake Ecology; Standard Methods for Observation and Analysis in Chinese Ecosystem Research Network, Series V.; Standards Press of China: Beijing, China, 1999. [Google Scholar]
- Vitál, Z.; Specziár, A.; Mozsár, A.; Takács, P.; Boros, G. Applicability of gill raker filtrates and foregut contents in the diet assessment of filter-feeding Asian carps. Fundam. Appl. Limnol. 2015, 187, 79–86. [Google Scholar] [CrossRef] [Green Version]
- Xie, P. Gut contents of bighead carp (Aristichthys nobilis) and the processing and digestion of algal cells in the alimentary canal. Aquaculture 2001, 195, 149–161. [Google Scholar] [CrossRef]
- O’Reilly, C.M.; Hecky, R.E.; Cohen, A.S.; Plisnier, P.D. Interpreting stable isotopes in food webs: Recognizing the role of time averaging at different trophic levels. Limnol. Oceanogr. 2002, 47, 306–309. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Wen, Z.; Gong, Z.; Zhang, M.; Xie, P.; Hansson, L.A. Seasonal Trophic Niche Shift and Cascading Effect of a Generalist Predator Fish. PLoS ONE 2012, 7, e49691. [Google Scholar] [CrossRef] [PubMed]
- Jackson, A.L.; Inger, R.; Parnell, A.C.; Bearhop, S. Comparing isotopic niche widths among and within communities: SIBER - Stable Isotope Bayesian Ellipses in R. J. Anim. Ecol. 2011, 80, 595–602. [Google Scholar] [CrossRef] [PubMed]
- Bollen, K.A.; Stine, R.A. Bootstrapping Goodness-of-Fit Measures in Structural Equation Models. Sociol. Methods Res. 1992, 21, 205–229. [Google Scholar] [CrossRef]
- Arbuckle, J.L. IBM SPSS Amos 20 User’s Guide; IBM Corp: New York, NY, USA, 2011. [Google Scholar]
- Parnell, A.C.; Inger, R.; Bearhop, S.; Jackson, A.L. Source partitioning using stable isotopes: Coping with too much variation. PLoS ONE 2010, 5, e9672. [Google Scholar] [CrossRef] [PubMed]
- Macarthur, R.H.; Pianka, E.R. On optimal use of patchy environment. Am. Nat. 1966, 100, 603–609. [Google Scholar] [CrossRef]
- Svanback, R.; Bolnick, D.I. Intraspecific competition affects the strength of individual specialization: An optimal diet theory method. Evol. Ecol. Res. 2005, 7, 993–1012. [Google Scholar]
- Hansen, A.; Ghosal, R.; Caprio, J.; Claus, A.W.; Sorensen, P.W. Anatomical and physiological studies of bigheaded carps demonstrate that the epibranchial organ functions as a pharyngeal taste organ. J. Exp. Biol 2014, 217, 3945–3954. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Xie, P. Studies on the Food Web Structure of Lake Donghu Using Stable Carbon and Nitrogen Isotope Ratios. J. Freshw. Ecol. 2004, 19, 645–650. [Google Scholar] [CrossRef]
- Song, G. Feeding Habits of Silver Carp and Bighead Carp in Qiandaohu Lake; Shanghai Ocean University: Shanghai, China, 2013. [Google Scholar]
- Beaugrand, G.; Brander, K.M.; Lindley, J.A.; Souissi, S.; Reid, P.C. Plankton effect on cod recruitment in the North Sea. Nature 2003, 426, 661–664. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, S.S.; Suzuki, K.; Kadoya, T.; Nakagawa, M.; Takamura, N. Bottom-up linkages between primary production, zooplankton, and fish in a shallow, hypereutrophic lake. Ecology 2018, 99, 2025–2036. [Google Scholar] [CrossRef] [PubMed]
- Sherwood, G.D.; Pazzia, I.; Moeser, A.; Hontela, A.; Rasmussen, J.B. Shifting gears: Enzymatic evidence for the energetic advantage of switching diet in wild-living fish. Can. J. Fish. Aquat. Sci. 2002, 59, 229–241. [Google Scholar] [CrossRef]
- Leigh, C.; Burford, M.A.; Sheldon, F.; Bunn, S.E. Dynamic stability in dry season food webs within tropical floodplain rivers. Mar. Freshw. Res. 2010, 61, 357–368. [Google Scholar] [CrossRef] [Green Version]
- Pettit, N.E.; Naiman, R.J.; Warfe, D.M.; Jardine, T.D.; Douglas, M.M.; Bunn, S.E.; Davies, P.M. Productivity and Connectivity in Tropical Riverscapes of Northern Australia: Ecological Insights for Management. Ecosystems 2017, 20, 492–514. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bighead Carp | |||||
|---|---|---|---|---|---|
| Season | Winter | Spring | Summer | Autumn | |
| Silver carp | Winter | 1 | <0.001 ** | 0.179 | 0.002 ** |
| Spring | <0.001 ** | 1 | 0.002 ** | 0.208 | |
| Summer | 0.003 ** | 0.042 * | 1 | 0.030 * | |
| Autumn | <0.001 ** | 0.886 | 0.016 * | 1 | |
| Fish Species | Food Resource | Contribution Proportions (%) | |||
|---|---|---|---|---|---|
| Winter | Spring | Summer | Autumn | ||
| Bighead carp | PP | 42.07 ± 2.92 | 32.71± 23.71 | 47.88 ± 2.21 | 26.42 ± 11.47 |
| ZP | 57.93 ± 2.92 | 67.29 ± 23.71 | 52.11 ± 2.21 | 73.57 ± 11.47 | |
| Silver carp | PP | 52.68 ± 7.78 | 37.52± 20.52 | 56.98 ± 3.51 | 32.95 ± 5.20 |
| ZP | 47.32 ± 7.78 | 62.48 ± 20.52 | 43.01 ± 3.51 | 67.05 ± 5.20 | |
| Trophic Level | Density of Plankton | ||||
|---|---|---|---|---|---|
| BC | SC | PP | ZP | ||
| Density of phytoplankton | Bac | 0.356 | 0.413 | 0.088 | 0.033 * |
| Cya | 0.005 ** | 0.003 ** | <0.001 ** | <0.001 ** | |
| Chl | 0.654 | 0.784 | 0.008 ** | 0.174 | |
| PP | 0.013 * | 0.014 * | 1 | <0.001 ** | |
| Density of zooplankton | Cop | 0.064 | 0.051 | <0.01 ** | <0.001 ** |
| Cla | 0.006 ** | 0.002** | 0.038 * | 0.002 ** | |
| ZP | 0.003 * | 0.001** | <0.001 ** | 1 | |
| Water parameters | TP | 0.296 | 0.238 | 0.021 * | 0.019 * |
| TN | 0.919 | 0.614 | 0.609 | 0.539 | |
| T | 0.184 | 0.118 | 0.001 ** | <0.001 ** | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, J.; Chen, J.; Deng, X.; Wu, Z.; Yu, Z.; Xu, J.; Su, H.; Liu, J.; Wang, L.; Wu, Y.; et al. Trophic Patterns of Bighead Carp and Silver Carp Follow the Seasonality of Resource Availability. Water 2019, 11, 1429. https://doi.org/10.3390/w11071429
Yu J, Chen J, Deng X, Wu Z, Yu Z, Xu J, Su H, Liu J, Wang L, Wu Y, et al. Trophic Patterns of Bighead Carp and Silver Carp Follow the Seasonality of Resource Availability. Water. 2019; 11(7):1429. https://doi.org/10.3390/w11071429
Chicago/Turabian StyleYu, Jia, Jun Chen, Xuwei Deng, Zhixu Wu, Zuoming Yu, Jun Xu, Haojie Su, Jiarui Liu, Li Wang, Yao Wu, and et al. 2019. "Trophic Patterns of Bighead Carp and Silver Carp Follow the Seasonality of Resource Availability" Water 11, no. 7: 1429. https://doi.org/10.3390/w11071429
APA StyleYu, J., Chen, J., Deng, X., Wu, Z., Yu, Z., Xu, J., Su, H., Liu, J., Wang, L., Wu, Y., & Xie, P. (2019). Trophic Patterns of Bighead Carp and Silver Carp Follow the Seasonality of Resource Availability. Water, 11(7), 1429. https://doi.org/10.3390/w11071429