Antidiabetic and Antihyperlipidemic Effects of Sulphurenic Acid, a Triterpenoid Compound from Antrodia camphorata, in Streptozotocin-Induced Diabetic Mice


Abstract
1. Introduction
2. Results
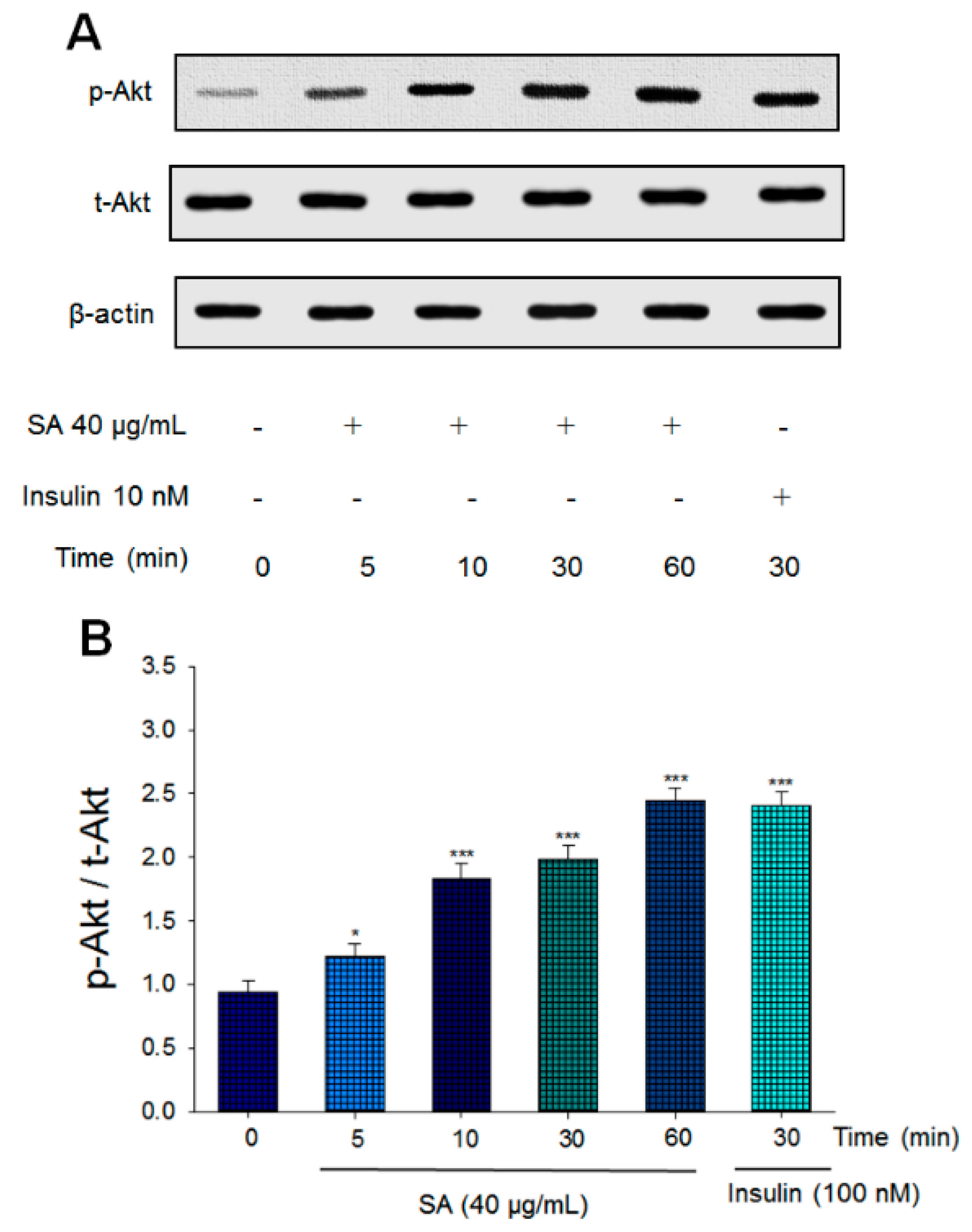
2.1. Expression Levels of Phosphorylated Akt in a Cell Line
2.2. Treatments in STZ-induced Diabetic Mice
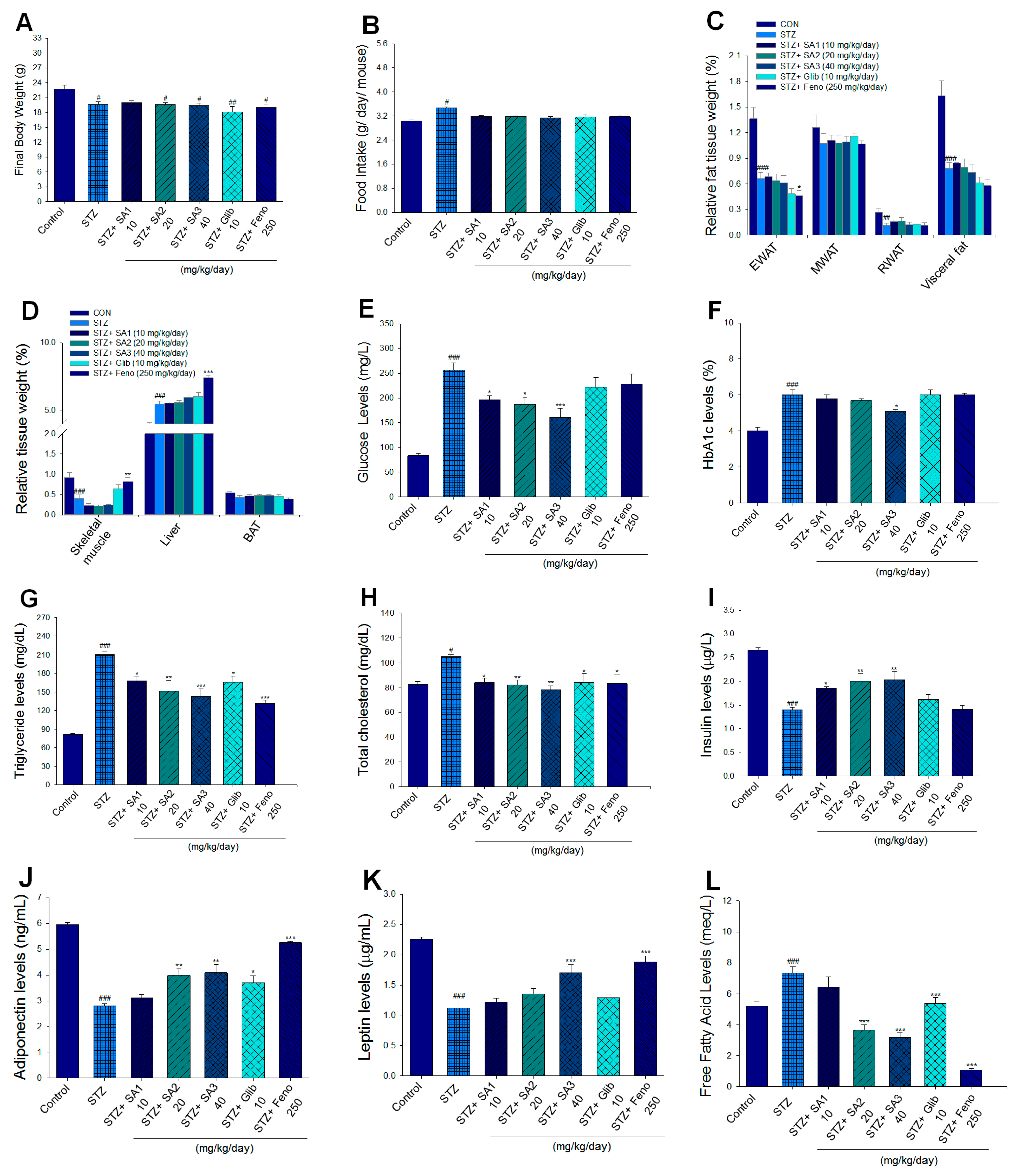
2.2.1. Body Weight, Food Intake, and Relative Tissue Weights
2.2.2. Blood Metabolic Parameters
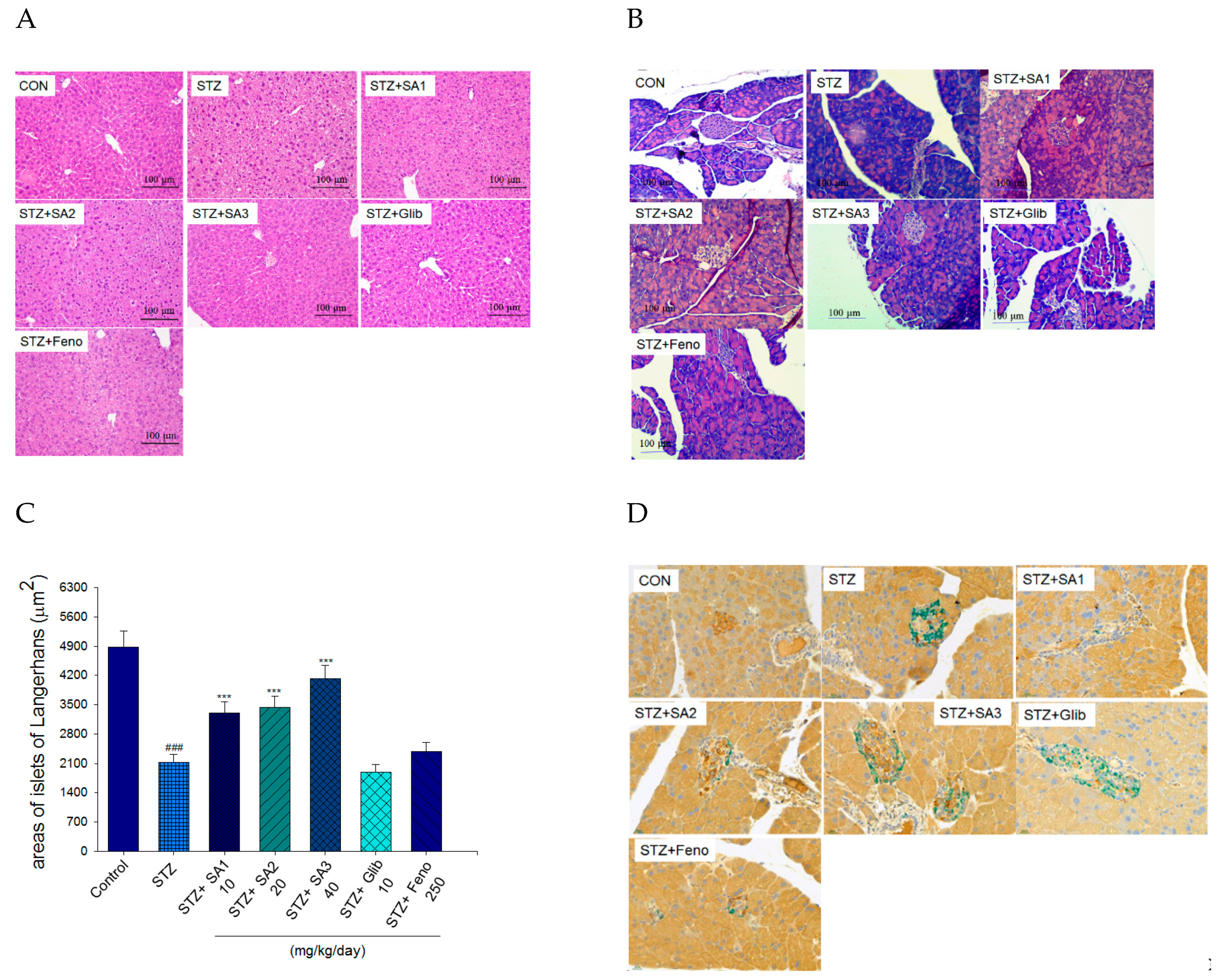
2.2.3. Histology
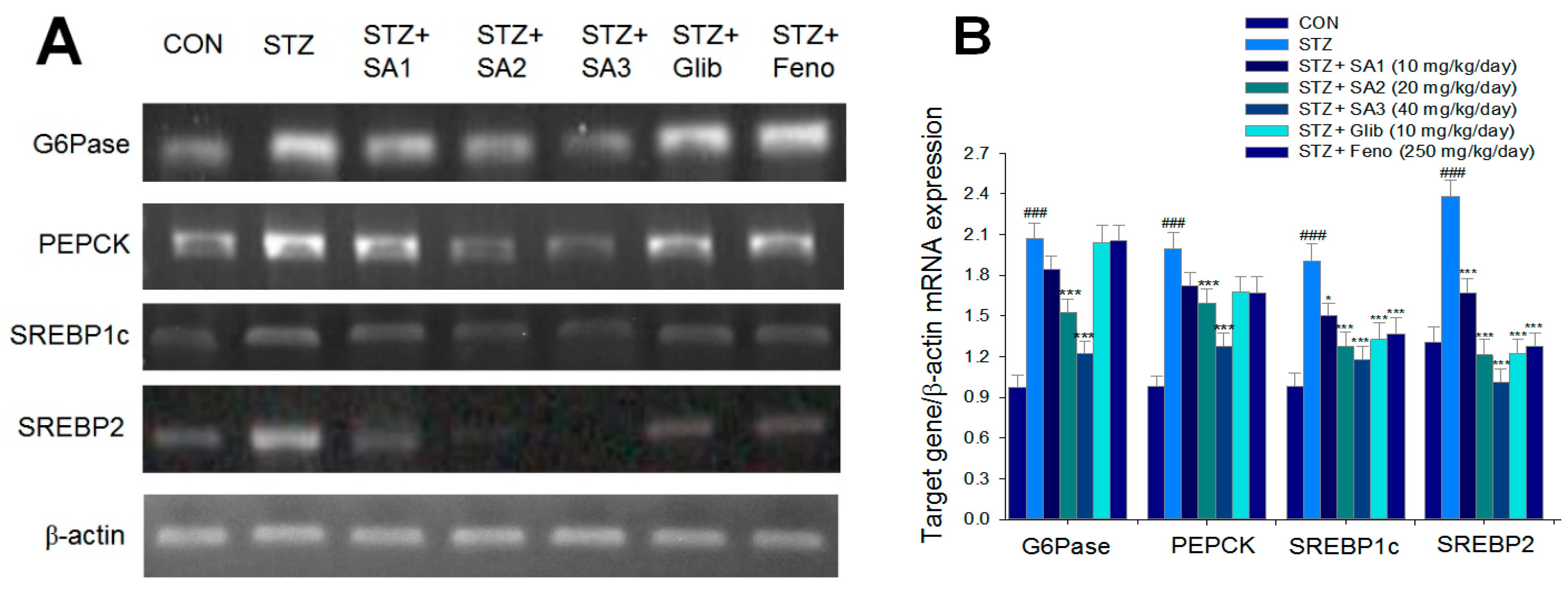
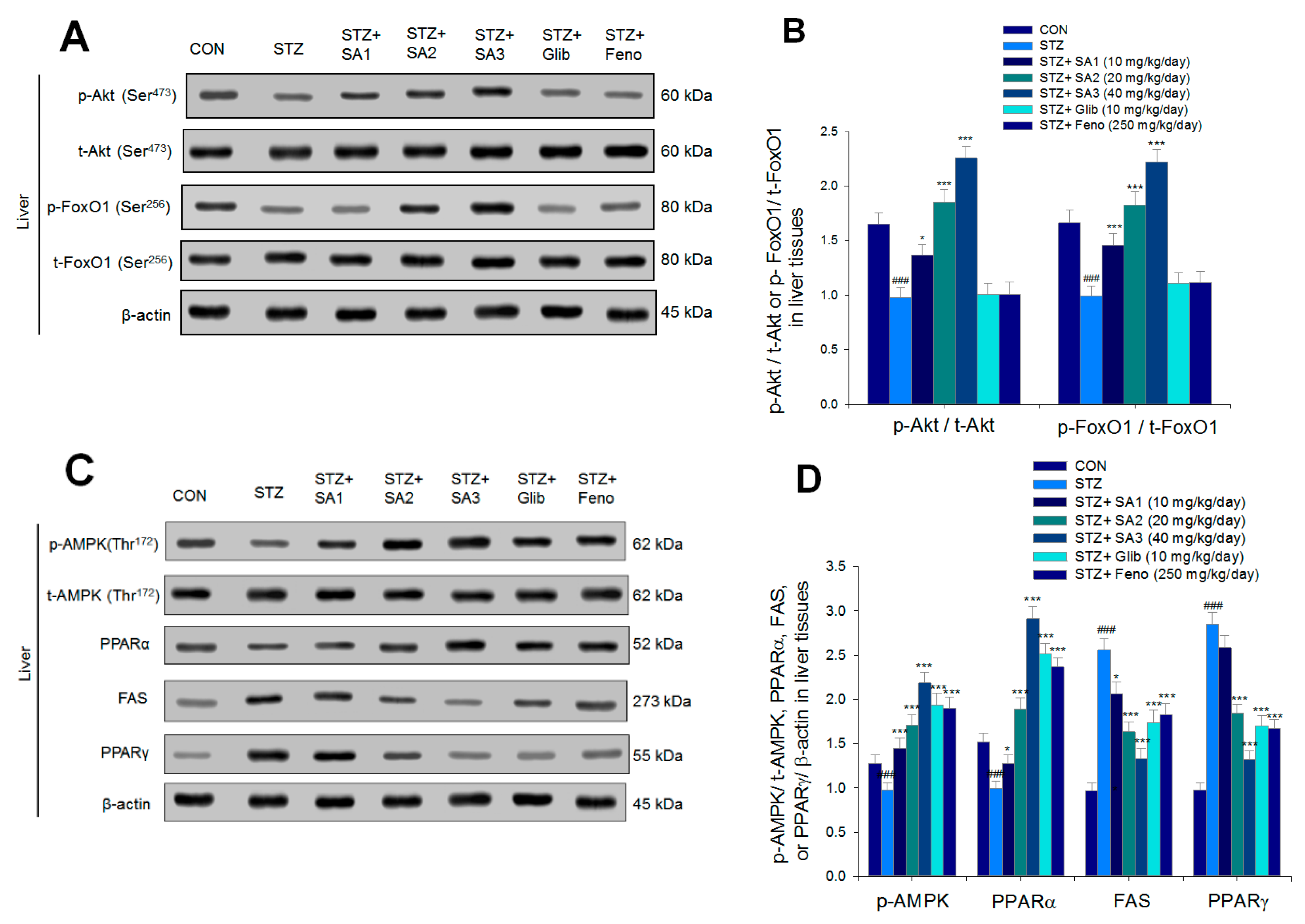
2.2.4. Hepatic Targeted Gene mRNA Levels
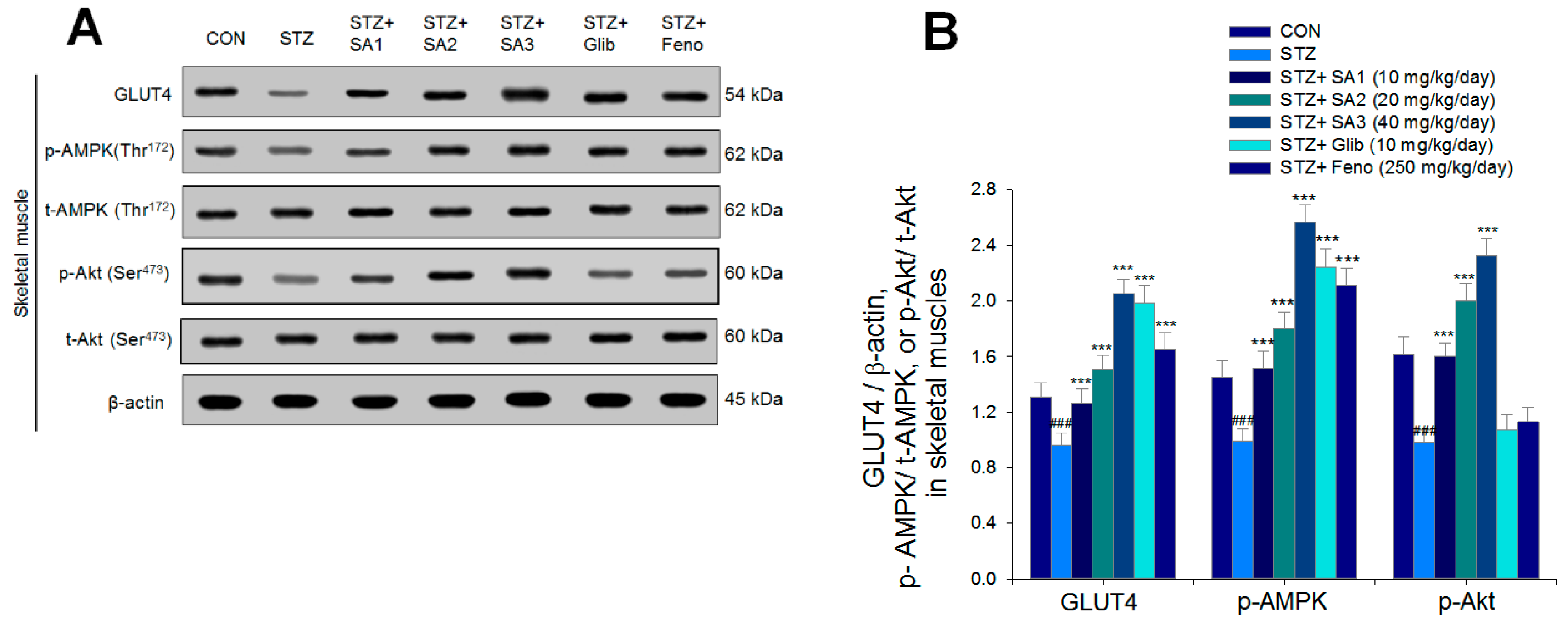
2.2.5. Targeted Gene Expression Levels in Different Tissues
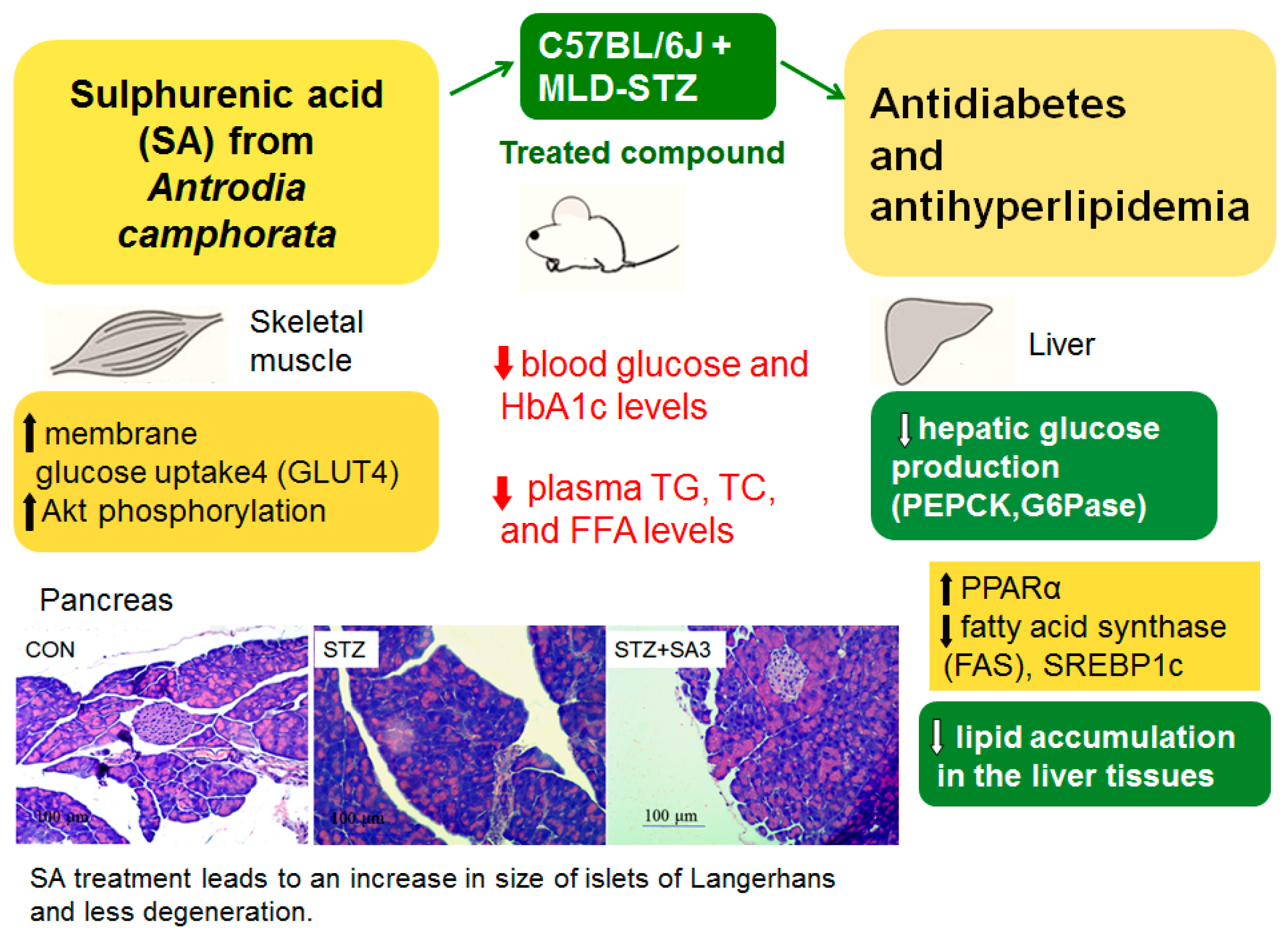
3. Discussion
4. Materials and Methods
4.1. Chemicals
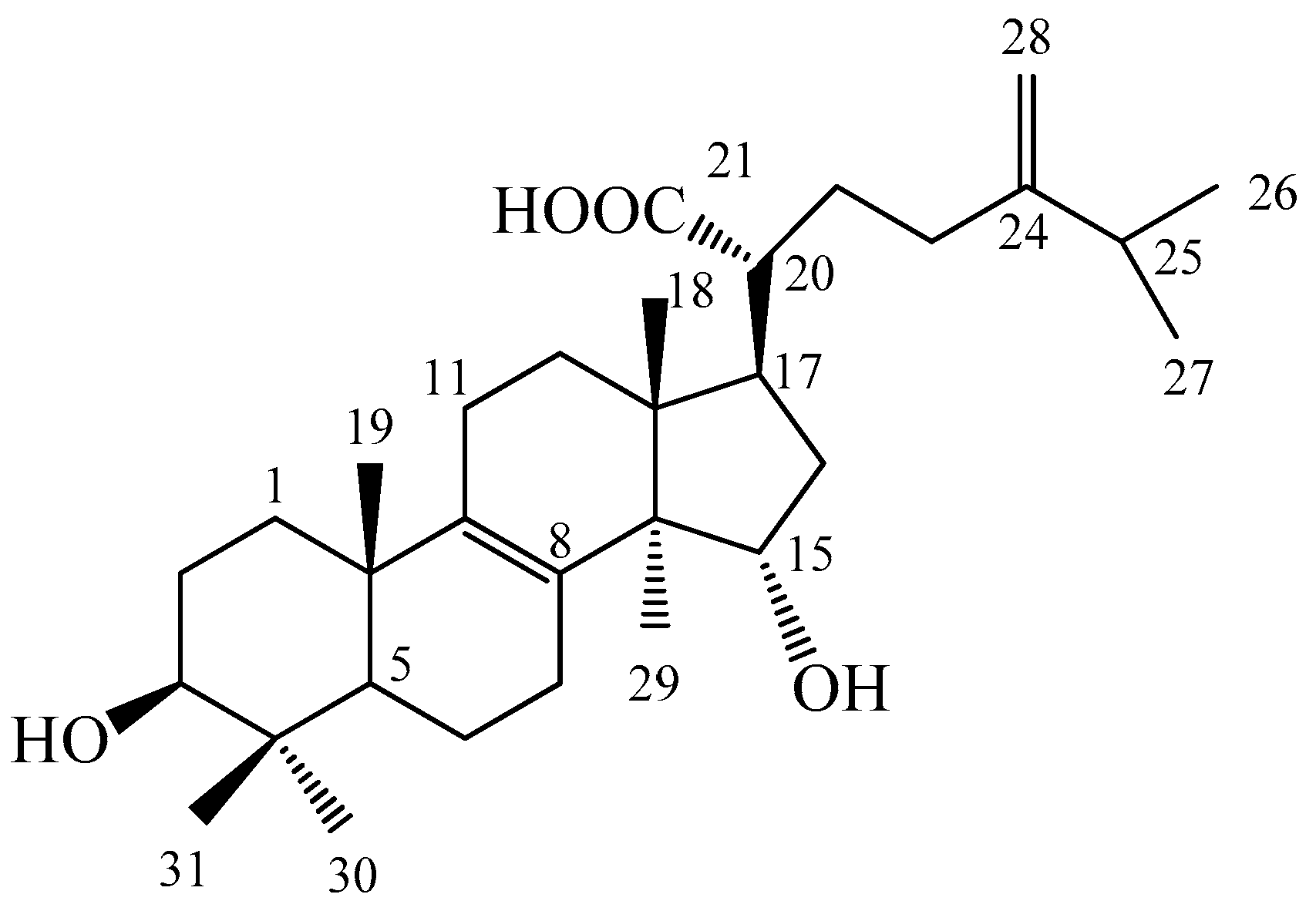
4.2. Isolation and Determination of the Active Compound
4.3. In Vitro Experiment
4.3.1. Cell Culture
4.3.2. Detection of Expression Levels of Phosphorylated Akt (Ser 473) in a Cell Line
4.4. Animals and Treatments
4.5. Analysis of Blood Glucose, Biochemical, Adipocytokine, and Glycated Hemoglobin (HbA1C) Levels
4.6. Histological Examination
4.7. Relative Quantification of mRNA and Western Blotting
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Chiang, J.L.; Kirkman, M.S.; Laffel, L.M.; Peters, A.L. Type 1 diabetes through the life span: A position statement of the American Diabetes Association. Diabetes Care 2014, 37, 2034–2054. [Google Scholar] [CrossRef] [PubMed]
- Daneman, D. Type 1 diabetes. Lancet 2006, 367, 847–858. [Google Scholar] [CrossRef]
- Geethangili, M.; Tzeng, Y.M. Review of pharmacological effects of Antrodia camphorata and its bioactive compounds. Evid-Based Complement. Altern. Med. 2011. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.J.; Deng, J.S.; Huang, S.S.; Lee, C.Y.; Hou, W.C.; Wang, S.Y.; Sung, P.J.; Kuo, Y.H. Hepatoprotective effects of eburicoic acid and dehydroeburicoic acid from Antrodia camphorata in a mouse model of acute hepatic injury. Food Chem. 2013, 141, 3020–3027. [Google Scholar] [CrossRef]
- Song, T.Y.; Yen, G.C. Protective effects of fermented filtrate from Antrodia camphorata in submerged culture against CCl4-induced hepatic toxicity in rats. J. Agric. Food Chem. 2003, 51, 1571–1577. [Google Scholar] [CrossRef] [PubMed]
- Kuo, Y.H.; Lin, C.H.; Shih, C.C. Ergostatrien-3β-ol from Antrodia camphorata inhibits diabetes and hyperlipidemia in high-fat-diet treated mice via regulation of hepatic related genes, glucose transporter 4, and AMP-Activated protein kinase phosphorylation. J. Agric. Food Chem. 2015, 63, 2479–2489. [Google Scholar] [CrossRef]
- Kuo, Y.H.; Lin, C.H.; Shih, C.C. Antidiabetic and antihyperlipidemic properties of a triterpenoid compound, dehydroeburicoic acid, from Antrodia camphorata in vitro and in streptozotocin-induced mice. J. Agric. Food Chem. 2015, 63, 10140–10151. [Google Scholar] [CrossRef]
- Kuo, Y.H.; Lin, C.H.; Shih, C.C. Dehydroeburicoic acid from Antrodia camphorata prevents the diabetic and dyslipidemic state via modulation of glucose transporter 4, peroxisome proliferator-activated receptor α expression and AMP-activated protein kinase phosphorylation in high-fat-fed mice. Int. J. Mol. Sci. 2016, 17, 872. [Google Scholar]
- Lin, C.H.; Kuo, Y.H.; Shih, C.C. Eburicoic acid, a triterpenoid compound from Antrodia camphorata, displays antidiabetic and antihyperlipidemic effects in palmitate-treated C2C12 myotubes and in high-fat diet-fed Mice. Int. J. Mol. Sci. 2017, 18, 2314. [Google Scholar] [CrossRef]
- Lin, C.H.; Kuo, Y.H.; Shih, C.C. Antidiabetic and hypolipidemic activities of eburicoic acid, a triterpenoid compound from Antrodia camphorata by regulation of Akt phosphorylation, gluconeogenesis, and PPARα in streptozotocin-induced diabetic mice. Rsc. Adv. 2018, 8, 20462–20476. [Google Scholar] [CrossRef]
- Kuo, Y.H.; Lin, C.H.; Shih, C.C.; Yang, C.S.; Antcin, K. A triterpenoid compound from Antrodia camphorata, displays antidiabetic and antihyperlipidemic effects via glucose transporter 4 and AMP-activated protein kinase phosphorylation in muscles. Evid-Based Complement. Altern. Med. 2016. [Google Scholar] [CrossRef] [PubMed]
- Tomlison, K.C.; Gardiner, S.M.; Hebden, R.A.; Bennett, T. Functional consequences of streptozotocin induced diabetes mellitus, with particular reference to the cardiovascular system. Pharmacol. Rev. 1992, 44, 103–150. [Google Scholar]
- Hayashi, K.; Kojima, R.; Ilo, M. Strain differences in the diabetogenic activity of streptozotocin in mice. Biol. Pharm. Bull. 2006, 29, 1110–1119. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Miao, J.; Zhang, Y. Protective effect of β-casomorphin-7 on type 1 diabetes rats induced with streptozotocin. Peptides 2010, 31, 1725–1729. [Google Scholar] [CrossRef] [PubMed]
- Elsner, M.; Guldbakke, B.; Tiedge, M.; Munday, R.; Lenzen, S. Relative importance of transport and alkylation for pancreatic beta-cell toxicity of streptozotocin. Diabetologia 2004, 43, 1528–1533. [Google Scholar] [CrossRef] [PubMed]
- Szkudelski, T. The mechanism of alloxan and streptozotocin action in β cells of the rat pancreas. Physiol. Res. 2001, 50, 537–546. [Google Scholar] [PubMed]
- Li, Z.; Karlsson, F.A.; Sandler, S. Islet loss and alpha cell expansion in type 1 diabetes induced by multiple low-dose streptozotocin administration in mice. J. Endocrinol. 2000, 165, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Mueckler, M. Facilitative glucose transporters. Eur. J. Biochem. 1994, 219, 713–725. [Google Scholar] [CrossRef]
- Bryant, N.J.; Govers, R.; James, D.E. Regulated transport of the glucose transporter GLUT4. Nature reviews. Mol. Cell. Biol. 2002, 3, 267–277. [Google Scholar] [CrossRef] [PubMed]
- DeFronzo, R.A.; Jocot, E.; Jequier, E.; Maeder, E.; Wahren, J.; Felber, J.R. The effect of insulin on the disposal of intravenous glucose. Diabetes 1981, 30, 1000–1007. [Google Scholar] [CrossRef]
- Ryder, J.W.; Yang, J.; Galuska, D.; Rincon, J.; Bjornholm, M.; Krook, A.; Lund, S.; Pedersen, O.; Wallberg-Henriksson, H.; Zierath, J.R.; et al. Use of a novel impermeable biotinylated photolabeling reagent to assess insulin- and hypoxia-stimulated cell surface GLUT4 content in skeletal muscle from type 2 diabetic patients. Diabetes 2000, 49, 647–654. [Google Scholar] [CrossRef] [PubMed]
- Minokoshi, Y.; Kahn, C.R.; Kahn, B.B. Tissue-specific ablation of the GLUT4 glucose transporter or the insulin receptor challenges assumptions about insulin and glucose homeostasis. J. Biol. Chem. 2003, 278, 33609–33612. [Google Scholar] [CrossRef] [PubMed]
- Lund, S.; Holman, G.D.; Schmitz, O.; Pedersen, O. Contraction stimulates translocation of glucose transporter GLUT4 in skeletal muscle through a mechanism distinct from that of insulin. Proc. Natl. Acad. Sci. USA 1995, 92, 5817–5821. [Google Scholar] [CrossRef] [PubMed]
- Welsh, G.I.; Hers, I.; Berwick, D.C.; Dell, G.; Wherlock, M.; Birkin, R.; Leney, S.; Tavsre, J.M. Role of protein kinase B in insulin-regulated glucose uptake. Biochem. Soc. Trans. 2005, 33, 346–349. [Google Scholar] [CrossRef] [PubMed]
- Joost, H.G.; Bell, G.I.; Best, J.D.; Birnbaum, M.J.; Charron, M.J.; Chen, Y.T.; Doege, H.; James, D.E.; Lodish, H.F.; Moley, K.H.; et al. Nomenclature of the GLUT4/SLC2A family of sugar/polyol transport facilitators. Am. J. Physiol. Endocrinol. Metab. 2002, 282, E974–E976. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Bryan, L.; Nichols, C.G.; Wechsler, S.W.; Clement, J.P.; Boyd, A.E.; Gonzalez, G.; Herrera-Sosa, H.; Nguy, K.; Bryan, J.; Nelson, D.A. Cloning the β cell high-affinity sulfonylurea receptor: A regulator of insulin secretion. Science 1995, 268, 423–426. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, D.B.; Hayes, G.R.; Lockwood, D.H. In vitro effects of sulfonylurea on glucose transport and translocation of glucose transporters in adipocytes from streptozocin-induced diabetic rats. American Diabetes Association. Diabetes 1989, 38, 205–211. [Google Scholar] [CrossRef]
- Blumenthal, S.A. Potentiation of the hepatic action of insulin by chlorpropamide. Diabetes 1977, 26, 485–489. [Google Scholar] [CrossRef]
- Stein, S.C.; Woods, A.; Jones, N.A.; Davison, M.D.; Carling, D. The regulation of AMP-activated protein kinase by phosphorylation. Biochem. J. 2000, 345, 437–443. [Google Scholar] [CrossRef]
- Staels, B.; Fruchart, J.C. Therapeutic roles of peroxisome proliferator-activated receptor agonists. Diabetes 2005, 54, 2460–2470. [Google Scholar] [CrossRef]
- Guignot, L.; Mithieux, G. Mechanisms by which insulin, associated or not with glucose, may inhibit hepatic glucose production in the rat. Am. J. Physiol. 1999, 227, 984–989. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, D.E.; Little, R.R.; Lorenz, R.A.; Malone, J.I.; Nathan, D.M.; Peterson, C.M. Tests of glycemia in diabetes. Diabetes Care 2004, 27, S91–S93. [Google Scholar] [CrossRef] [PubMed]
- Huebschmann, A.G.; Regensteiner, J.G.; Vlassara, H.; Reusch, J.E. Diabetes and advanced glycoxidation end products. Diabetes Care 2006, 29, 1420–1432. [Google Scholar] [CrossRef]
- Al-Shamaorry, L.; Al-Khazraji, S.M.; Twaiji, H.A. Hypoglycemic effect of Artemisia herba alba II. Effect of a valuable extract on some blood glucose parameters in diabetic animals. J. Ethnopharmacol. 1994, 43, 167–171. [Google Scholar] [CrossRef]
- Rajkumar, L.; Srinivasan, N.; Balasubramanian, K.; Govindarajulu, P. Increased degradation of dermal collagen in diabetic rats. J. Exp. Biol. 1991, 29, 1081–1083. [Google Scholar]
- Chatterja, M.N.; Shinde, R. Text Book of Medical Biochemisty, 5th ed.; Jaypee Brothers Medical Publishers: Panama City, Panama, 2002. [Google Scholar]
- Kitazawa, N.; Miura, T.; Kako, M.; Usami, M.; Tanigawa, K.; Ishida, H.; Seino, Y. Determination of Hemoglobin A1C in normal and diabetic mice: Neonatal streptozotocin-induced diabetic mice and KK-Ay mice. Biol. Pharm. Bull. 1996, 19, 1078–1079. [Google Scholar] [CrossRef][Green Version]
- Coskun, O.; Kanter, M.; Korkmaz, A.; Oter, S. Quercetin, a flavonoid antioxidant, prevents and protects streptozotocin-induced oxidative stress and beta-cell damage in rat pancreas. Pharmacol. Res. 2005, 51, 117–123. [Google Scholar] [CrossRef]
- Fukudome, D.; Matsuda, M.; Kawasaki, T.; Ago, Y.; Matsuda, T. The radical scavenger edaravone counteracts diabetes in multiple low-dose streptozotocin-treated mice. Eur. J. Pharmacol. 2008, 583, 164–169. [Google Scholar] [CrossRef]
- Kaneto, H.; Kajimoto, Y.; Miyagawa, J.; Matsuoka, T.; Fujitani, Y.; Umayahara, Y.; Hanafusa, T.; Matsuzawa, Y.; Yamasaki, Y.; Hori, M. Beneficial effects of antioxidants in diabetes: Possible protection of pancreatic β-cells against glucose toxicity. Diabetes 1999, 48, 2398–2406. [Google Scholar] [CrossRef]
- Cherrington, A.D.; Edgerton, D.S.; Ramnanan, C. The role of insulin in the regulation of PEPCK and gluconeogenesis in vivo. Us Endocrinol. 2009, 5, 34–39. [Google Scholar]
- Matsuzaki, H.; Daitoku, H.; Hatta, M.; Tanaka, K.; Fukamizu, A. Insulin-induced phosphorylation of FKHR (Foxo1) targets to proteasomal degradation. Proc. Natl. Acad. Sci. USA 2003, 100, 11285–11290. [Google Scholar] [CrossRef] [PubMed]
- Nakae, J.; Kitamura, T.; Silver, D.L.; Accili, D. The forkhead transcription factor Foxo1 (Fkhr) confers insulin sensitivity onto glucose-6-phospatase expression. J. Clin. Invest. 2001, 108, 1359–1367. [Google Scholar] [CrossRef] [PubMed]
- Oh, K.J.; Han, H.S.; Kim, M.J.; Koo, S.H. CREBP and FoxO1: Two transcription factors for the regulation of hepatic gluconeogenesis. Bmb. Rep. 2013, 46, 567–574. [Google Scholar] [CrossRef] [PubMed]
- Daisy, P.; Balasubramanian, K.; Manikkam, R.; Eliza, J.; Selvaraj, J.C. Insulin mimetic impact of catechin isolated from Cassia fistula on the glucose oxidation and molecular mechanisms of glucose uptake on streptozotocin induced diabetic wistar rats. Phytomedicine 2010, 17, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.S.; Yokozawa, T.; Oura, H. Improvement of hyperglycemia and hyperlipidemia in streptozotocin-diabetic rats by a methanolic extract of Prunus davidiana stems and its main component, pruning. Planta Med. 1991, 57, 208–211. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.R.; Dwivedi, S.K.; Swaruo, D. Hypoglycaemic and hypolipidemic effects of Cinnmomum tamala Nees leaves. Indianj. Exp. Biol. 1996, 34, 372–374. [Google Scholar]
- Wakil, S. Fatty acid synthase, a proficient multifunctional enzyme. Biochemistry 1989, 28, 4523–4530. [Google Scholar] [CrossRef] [PubMed]
- Shimano, H.; Yahagi, N.; Amemiya-Kudo, M.; Hasty, A.H.; Osuga, J.; Tamura, Y.; Shionoiri, F.; Iizuka, Y.; Ohashi, K.; Harada, K.; et al. Sterol regulatory element-binding protein-1 as a key transcription factor for nutritional induction of lipogenic enzyme genes. J. Biol. Chem. 1999, 274, 35832–35839. [Google Scholar] [CrossRef]
- Saladin, R.; Fajas, L.; Dana, S.; Halvorsen, Y.D.; Auwerx, J.; Briggs, M. Differential regulation of peroxisome proliferator activated receptor gamma1 (PPARgamma1) and PPARgamma2 messenger RNA expression in the early stages of adipogenesis. Cell Growth Differ. 1999, 10, 43–48. [Google Scholar]
- Yamauchi, T.; Kamon, J.; Minokoshi, Y.; Ito, Y.; Waki, H.; Uchida, S. Adiponectin stimulates glucose utilization and fatty-acid oxidation by activating AMP-activated protein kinase. Nat. Med. 2002, 8, 1288–1295. [Google Scholar] [CrossRef] [PubMed]
- Minokoshi, Y.; Kim, Y.B.; Peroni, O.D.; Fryer, L.G.; Muller, C.; Carling, D.; Kahn, B.B. Leptin stimulates fatty-acid oxidation by activating AMP-activated protein. Nature 2002, 415, 339–343. [Google Scholar] [CrossRef] [PubMed]
- Bourgeois, F.; Alexiu, A.; Lemonnier, D. Dietary-induced obesity: Effect of dietary fats on adipose tissue cellularity in mice. Br. J. Nutr. 1983, 49, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.I.; Chen, C.C.; Lin, T.W.; Tsai, Y.T.; Nam, M.K. A 90-day subchronic toxicological assessment of Antrodia cinnamomea in Sprague-Dawley rats. Food Chem. Toxicol. 2010, 49, 429–433. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, K.; Matsumoto, K.; Mine, C.; Bando, S.; Arihara, S. Five lanostane triterpenoids and three saponins from the fruit body of Laetiporus versisporus. Chem. Pharm. Bull. 2000, 48, 1418–1421. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.C.; Kuo, Y.C.; Huang, R.L.; Lin, L.C.; Don, M.J.; Chang, T.T.; Chou, C.J. New ergostane and lanostane from Antrodia camphorata. J. Chin. Med. 2003, 14, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.H.; Wu, J.B.; Jian, J.Y.; Shih, C.C. (−)-Epicatechin-3-O-β-D-allopyranoside from Davallia formosana prevents diabetes and dyslipidemia in streptozotocin-induced diabetic mice. PLoS ONE 2017, 12, e0173984. [Google Scholar] [CrossRef] [PubMed]
- Shih, C.C.; Chen, M.H.; Lin, C.H. Validation of the antidiabetic and hypolipidemic effects of Clitocybe nuda by assessment of glucose transporter 4 and gluconeogenesis and AMPK phosphorylation in streptozotocin-induced mice. Evid-Based Complement. Altern. Med. 2014, 2014, 705636. [Google Scholar] [CrossRef] [PubMed]
- Novikova, L.; Smirnova, I.V.; Rawal, S.; Dotson, A.L.; Benedict, S.H.; Stehno-Bittel, L. Variations in rodent models of type 1 diabetes: Islet morphology. J. Diabetes Res. 2013, 2013, 965832. [Google Scholar] [CrossRef]
- Klip, A.; Ramlal, T.; Young, D.A.; Holloszy, J.O. Insulin-induced translocation of glucose transporters in rat hindlimb muscles. Febs. Lett. 1987, 224, 224–230. [Google Scholar] [CrossRef]
- Shih, C.C.; Lin, C.H.; Lin, W.L.; Wu, J.B. Momordica charantia extract on insulin resistance and the skeletal muscle GLUT4 protein in fructose-fed rats. J. Ethnopharmacol. 2009, 123, 82–90. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Accession Number | Forward Primer and Reverse Primer | PCR Product (bp) | Annealing Temperature (°C) |
|---|---|---|---|---|
| Liver | ||||
| PEPCK | NM_011044.2 | F: CTACAACTTCGGCAAATACC R: TCCAGATACCTGTCGATCTC | 330 | 52 |
| G6Pase | NM_008061.3 | F: GAACAACTAAAGCCTCTGAAAC R: TTGCTCGATACATAAAACACTC | 350 | 50 |
| SREBP1c | NM_011480 | F: GGCTGTTGTCTACCATAAGC R: AGGAAGAAACGTGTCAAGAA | 219 | 50 |
| SREBP2 | AF289715.2 | F: ATATCATTGAAAAGCGCTAC R: ATTTTCAAGTCCACATCACT | 256 | 47 |
| β-actin | NM_007392 | F: TCTCCACCTTCCAGCAGATGT R: GCTCAGTAACAGTCCGCCTAGA | 99 | 55 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, C.-H.; Hsiao, L.-W.; Kuo, Y.-H.; Shih, C.-C. Antidiabetic and Antihyperlipidemic Effects of Sulphurenic Acid, a Triterpenoid Compound from Antrodia camphorata, in Streptozotocin-Induced Diabetic Mice. Int. J. Mol. Sci. 2019, 20, 4897. https://doi.org/10.3390/ijms20194897
Lin C-H, Hsiao L-W, Kuo Y-H, Shih C-C. Antidiabetic and Antihyperlipidemic Effects of Sulphurenic Acid, a Triterpenoid Compound from Antrodia camphorata, in Streptozotocin-Induced Diabetic Mice. International Journal of Molecular Sciences. 2019; 20(19):4897. https://doi.org/10.3390/ijms20194897
Chicago/Turabian StyleLin, Cheng-Hsiu, Li-Wei Hsiao, Yueh-Hsiung Kuo, and Chun-Ching Shih. 2019. "Antidiabetic and Antihyperlipidemic Effects of Sulphurenic Acid, a Triterpenoid Compound from Antrodia camphorata, in Streptozotocin-Induced Diabetic Mice" International Journal of Molecular Sciences 20, no. 19: 4897. https://doi.org/10.3390/ijms20194897
APA StyleLin, C.-H., Hsiao, L.-W., Kuo, Y.-H., & Shih, C.-C. (2019). Antidiabetic and Antihyperlipidemic Effects of Sulphurenic Acid, a Triterpenoid Compound from Antrodia camphorata, in Streptozotocin-Induced Diabetic Mice. International Journal of Molecular Sciences, 20(19), 4897. https://doi.org/10.3390/ijms20194897