Effects of Very Low Calorie Ketogenic Diet on the Orexinergic System, Visceral Adipose Tissue, and ROS Production



 ,
,  ,
, 
 , ,
, ,  ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Study Protocol
2.3. Orexin-A Assay
2.4. Dietary Intervention
2.5. Cell Culture and Cell Proliferation Assay
2.6. ROS Production Assay
2.7. d-ROM Test
2.8. Statistical Analysis
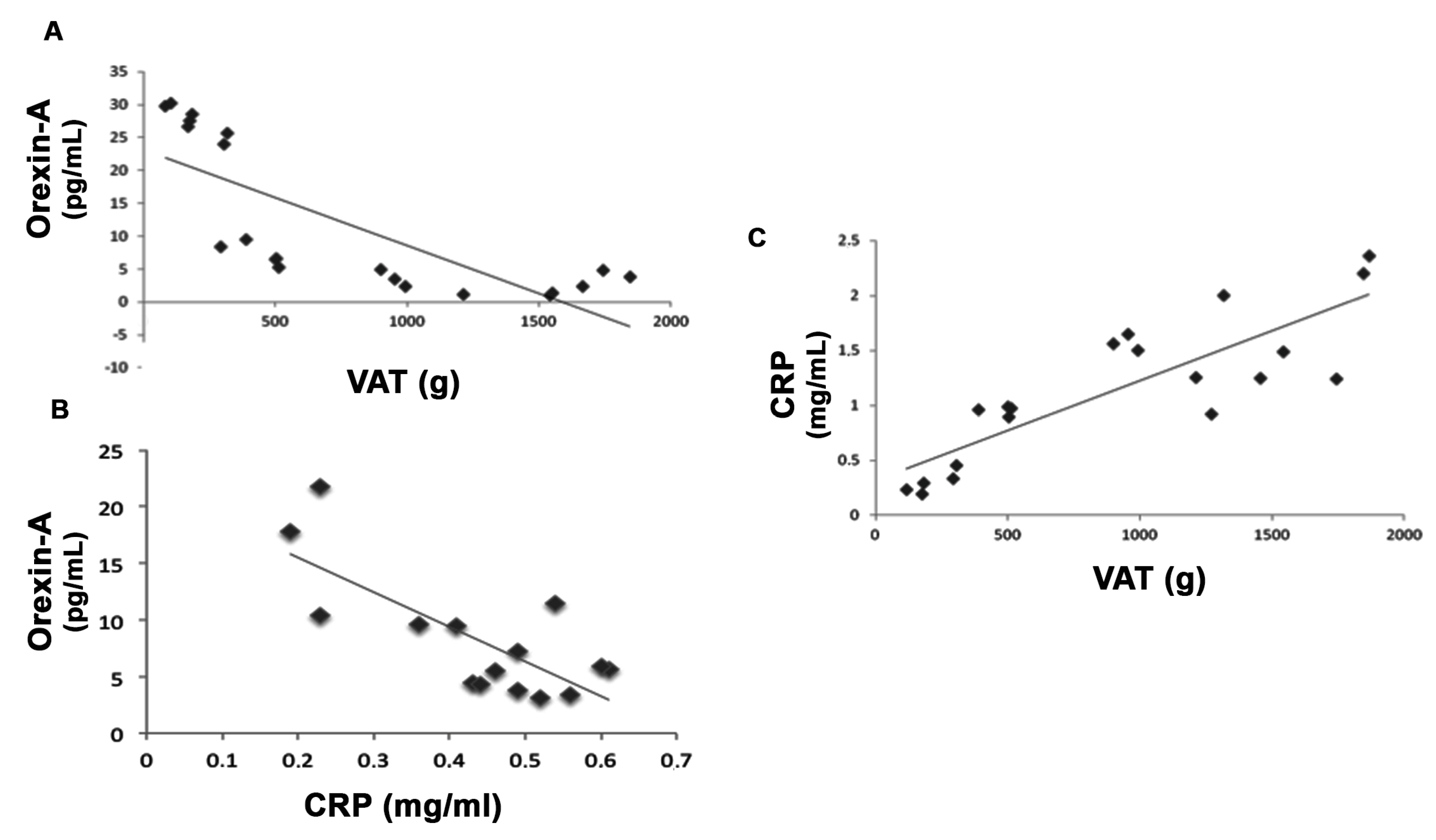
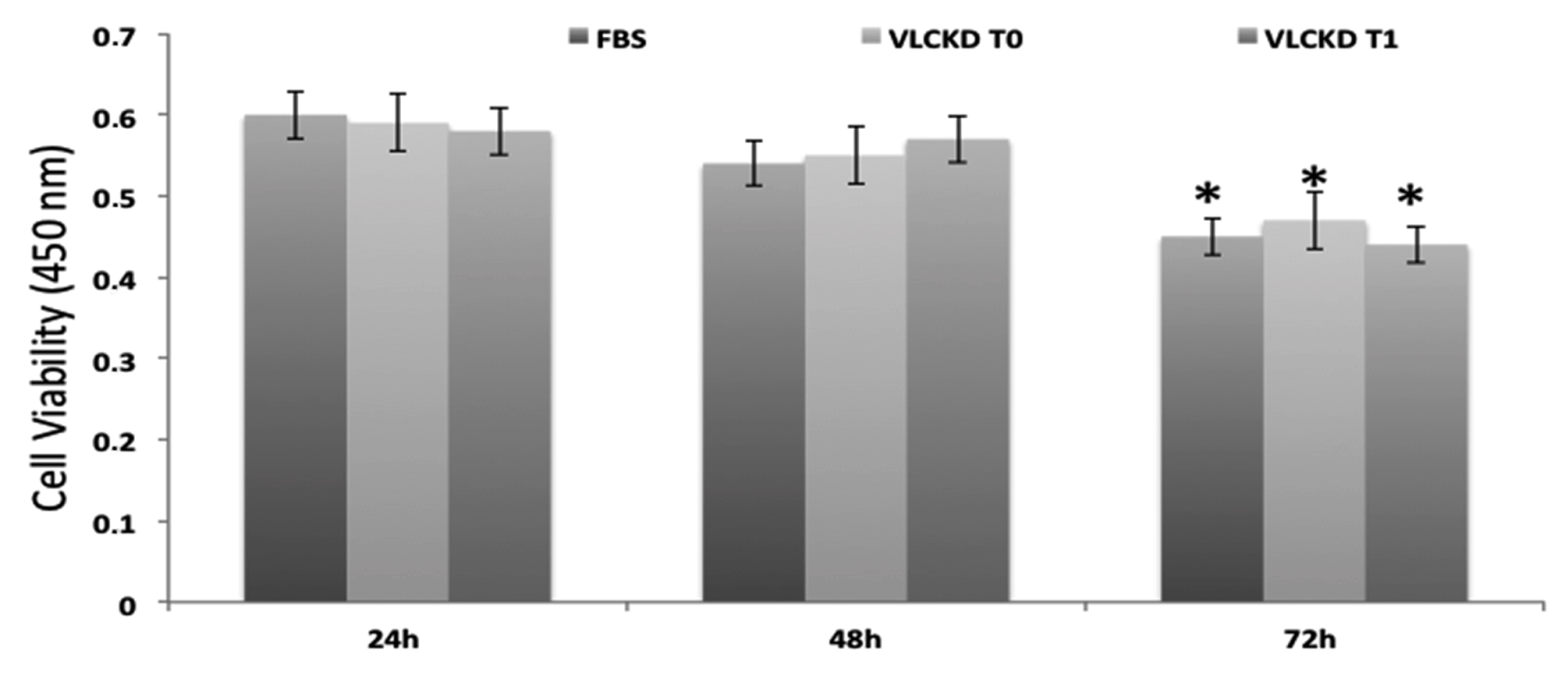
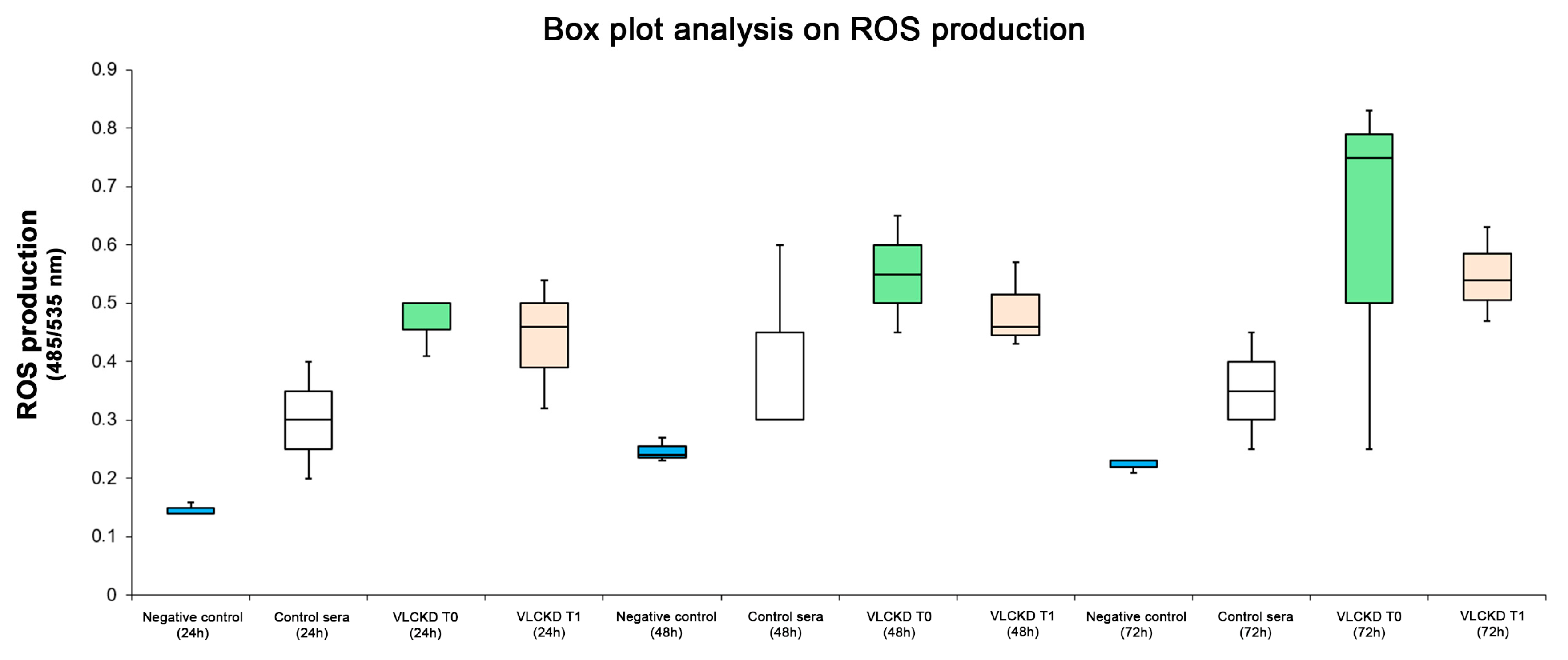
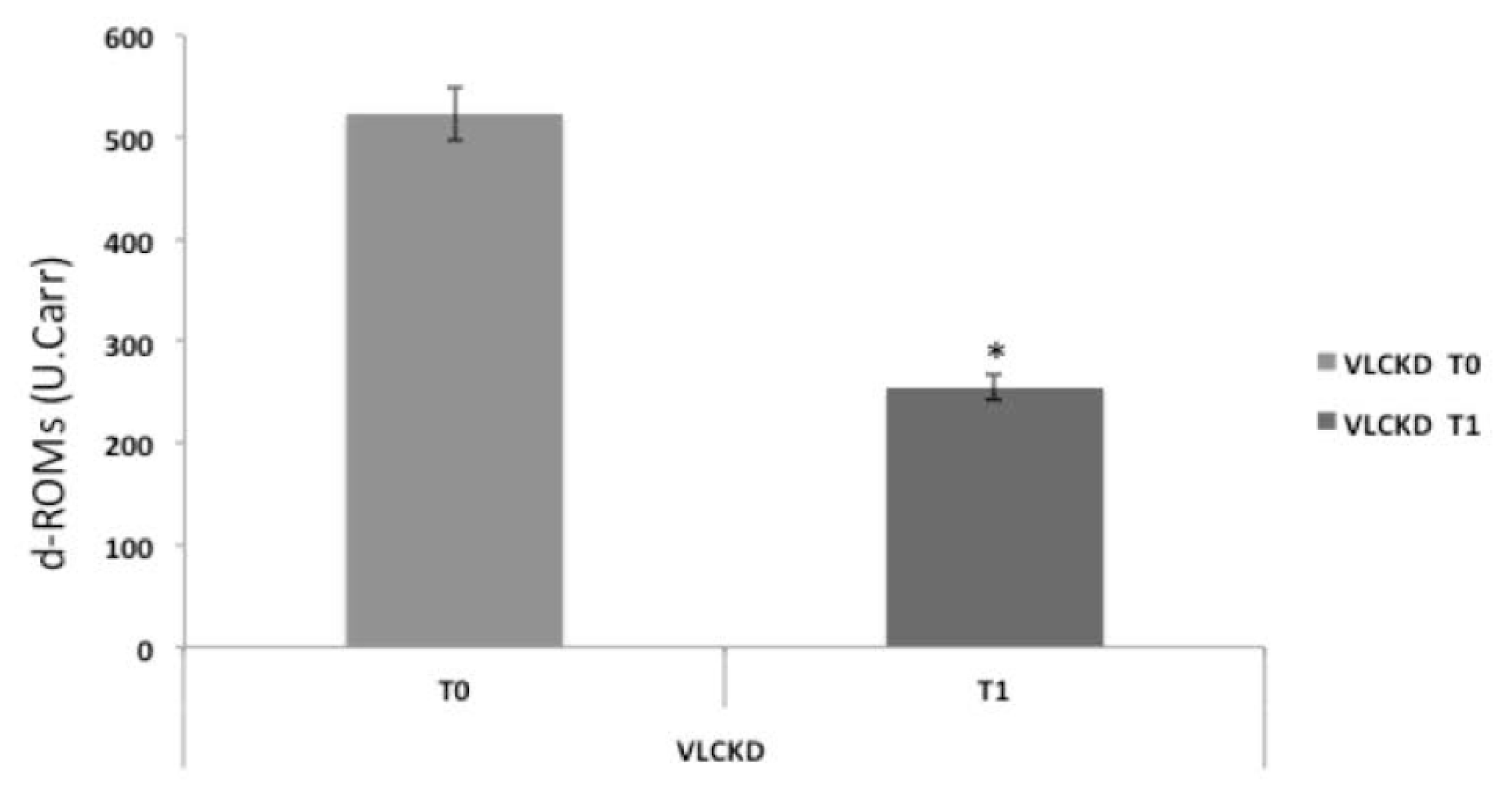
3. Results
3.1. Anthropometric and Biochemical Parameters of VLCKD Obese Patients
3.2. MTT Test
3.3. ROS Production
3.4. Serum Reactive Oxygen Metabolites Test
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bray, G.A.; Kim, K.K.; Wilding, J.P.H. Obesity: A chronic relapsing progressive disease process. A position statement of the World Obesity Federation. Obes. Rev. 2017, 18, 715–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joint WHO/FAO Expert Consultation. World Health Organization: Diet., Nutrition and the Prevention of Chronic Diseases; WHO Technical Report Series 916; WHO: Geneva, Switzerland, 2003. [Google Scholar]
- Polito, R.; Nigro, E.; Messina, A.; Monaco, M.L.; Monda, V.; Scudiero, O.; Cibelli, G.; Valenzano, A.; Picciocchi, E.; Zammit, C.; et al. Adiponectin and Orexin-A as a Potential Immunity Link Between Adipose Tissue and Central Nervous System. Front. Physiol. 2018, 9, 982. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, M.M. Subcutaneous and visceral adipose tissue: Structural and functional differences. Obes. Rev. 2010, 11, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Alexopoulos, N.; Katritsis, D.; Raggi, P. Visceral adipose tissue as a source of inflammation and promoter of atherosclerosis. Atherosclerosis 2014, 233, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Bi, X.; Seabolt, L.; Shibao, C.; Buchowski, M.; Kang, H.; Keil, C.D.; Tyree, R.; Silver, H.J. Dxa-measured visceral adipose tissue predicts impaired glucose tolerance and metabolic syndrome in obese Caucasian and African-American women. Eur. J. Clin. Nutr. 2015, 69, 329–336. [Google Scholar] [CrossRef] [Green Version]
- Hauner, H. Secretory factors from human adipose tissue and their functional role. Proc. Nutr. Soc. 2005, 64, 163–169. [Google Scholar] [CrossRef] [Green Version]
- Halberg, N.; Wernstedt-Asterholm, I.; Scherer, P.E. The adipocyte as an endocrine cell. Endocrinol. Metab. Clin. N. Am. 2008, 37, 753–768. [Google Scholar] [CrossRef] [Green Version]
- Messina, A.; Monda, V.; Sessa, F.; Valenzano, A.; Salerno, M.; Bitetti, I.; Precenzano, F.; Marotta, R.; Lavano, F.; Lavano, S.M.; et al. Sympathetic, Metabolic Adaptations, and Oxidative Stress in Autism Spectrum Disorders: How Far from Physiology? Front. Physiol. 2018, 9, 261. [Google Scholar] [CrossRef]
- Roccella, M.; Marotta, R.; Operto, F.F.; Smirni, D.; Precenzano, F.; Bitetti, I.; Messina, G.; Sessa, F.; Di Mizio, G.; Loreto, C.; et al. NREM Sleep Instability in Pediatric Migraine without Aura. Front. Neurol. 2019, 10, 932. [Google Scholar] [CrossRef] [Green Version]
- Tsujino, N.; Sakurai, T. Orexin/hypocretin: A neuropeptide at the interface of sleep, energy homeostasis, and reward system. Pharmacol. Rev. 2009, 61, 162–176. [Google Scholar] [CrossRef] [Green Version]
- Messina, G.; Dalia, C.; Tafuri, D.; Monda, V.; Palmieri, F.; Dato, A.; Russo, A.; De Blasio, S.; Messina, A.; De Luca, V.; et al. Orexin-A controls sympathetic activity and eating behavior. Front. Psychol. 2014, 5, 997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alam, M.N.; Kumar, S.; Bashir, T.; Suntsova, N.; Methippara, M.M.; Szymusiak, R. GABA-mediated control of hypocretin- but not melanin-concentrating hormone-immunoreactive neurones during sleep in rats. J. Physiol. 2005, 563, 569–582. [Google Scholar] [CrossRef] [PubMed]
- Adam, J.A.; Menheere, P.P.C.A.; van Dielen, F.M.H.; Soeters, P.B.; Buurman, W.A.; Greve, J.W.M. Decreased plasma orexin-A levels in obese individuals. Int. J. Obes. 2002, 26, 274–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Digby, J.E.; Chen, J.; Tang, J.Y.; Lehnert, H.; Matthews, R.N.; Randeva, H.S. Orexin receptor expression in human adipose tissue: Effects of orexin-A and orexin-B. J. Endocrinol. 2006, 191, 129–136. [Google Scholar] [CrossRef]
- Blais, A.; Drouin, G.; Chaumontet, C.; Voisin, T.; Couvelard, A.; Even, P.C. Impact of Orexin-A Treatment on Food Intake, Energy Metabolism and Body Weight in Mice. PLoS ONE 2017, 12, e0169908. [Google Scholar] [CrossRef] [Green Version]
- Kok, S.W.; Overeem, S.; Visscher, T.L.S.; Lammers, G.J.; Seidell, J.C.; Pijl, H. Hypocretin deficiency in narcoleptic humans is associated with abdominal obesity. Obes. Res. 2003, 11, 1147–1154. [Google Scholar] [CrossRef] [Green Version]
- Salminen, A.; Kaaeniranta, A.; Kauppinen, A. Crosstalk between Oxidative Stress and SIRT1: Impact on the Aging Process. Int. J. Mol. Sci. 2013, 14, 3834–3859. [Google Scholar] [CrossRef] [Green Version]
- Boison, D. New insights into the mechanisms of the ketogenic diet. Curr. Opin. Neurol. 2017, 30, 187–192. [Google Scholar] [CrossRef] [Green Version]
- Adam-Perrot, A.; Clifton, P.; Brouns, F. Low carbohydrate diets: Nutritional and physiological aspects. Obes. Rev. 2006, 7, 49–58. [Google Scholar] [CrossRef]
- Micklesfield, L.K.; Goedecke, J.H.; Punyanitya, M.; Wilson, K.E.; Kelly, T.L. Dual-energy X-ray performs as well as clinical computed tomography for the measurement of visceral fat. Obesity 2012, 20, 1109–1114. [Google Scholar] [CrossRef]
- SCOOP-VLCD Task 7.3 Reports on Tasks for Scientific Cooperation; Collection of Data on Products Intended for Use in Very-Lowcalorie- Diets; Report Brussels European Commission: Brussels, Belgium, 2002.
- Cornelli, U.; Terranova, R.; Luca, S.; Cornelli, M.; Alberti, A. Bioavailability and antioxidant activity of some food supplements in men and women using the D-Roms test as a marker of oxidative stress. J. Nutr. 2001, 12, 3208–3211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno, B.; Crujeiras, A.B.; Bellido, D.; Sajoux, I.; Casanueva, F.F. Obesity treatment by very low-calorie-ketogenic diet at two years: Reduction in visceral fat and on the burden of disease. Endocrine 2016, 3, 681–690. [Google Scholar] [CrossRef] [PubMed]
- Farb, M.G.; Gokce, N. Visceral adiposopathy: A vascular perspective. Horm. Mol. Biol. Clin. Investig. 2015, 2, 125–136. [Google Scholar] [CrossRef] [PubMed]
- Magnuson, A.M.; Fouts, J.K.; Regan, D.P.; Booth, A.D.; Dow, S.W.; Foster, M.T. Adipose Tissue Extrinsic Factor: Obesity-Induced Inflammation and the Role of the Visceral Lymph Node. Physiol. Behav. 2018, 190, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Abraham, P.A.; Attipoe, S.; Kazman, J.B.; Zeno, S.A.; Poth, M.; Deuster, P.A. Role of plasma adiponectin/C-reactive protein ratio in obesity and type 2 diabetes among African Americans. Afr. Health Sci. 2017, 99, 107. [Google Scholar] [CrossRef] [Green Version]
- Moro, T.; Tinsley, G.; Bianco, A.; Marcolin, G.; Pacelli, Q.F.; Battaglia, G. Effects of eight weeks of time-restricted feeding (16/8) on basal metabolism, maximal strength, body composition, inflammation, and cardiovascular risk factors in resistance-trained males. J. Transl. Med. 2016, 13, 290. [Google Scholar] [CrossRef]
- Kosinski, C.; Jornayvaz, F.R. Effects of Ketogenic Diets on Cardiovascular Risk Factors: Evidence from Animal and Human Studies. Nutrients 2017, 19, 517. [Google Scholar] [CrossRef]
- Fung, T.T.; van Dam, R.M.; Hankinson, S.E.; Stampfer, M.; Willett, W.C.; Hu, F.B. Low-carbohydrate diets and all-cause and cause-specific mortality: Two cohort studies. Ann. Intern. Med. 2010, 153, 289–298. [Google Scholar] [CrossRef]
- Scott, S.N.; Anderson, L.; Morton, J.P.; Wagenmakers, A.J.M.; Riddell, M.C. Carbohydrate Restriction in Type 1 Diabetes: A Realistic Therapy for Improved Glycaemic Control and Athletic Performance? Nutrients 2019, 7, 1022. [Google Scholar] [CrossRef] [Green Version]
- Bueno, N.B.; de Melo, I.S.; de Oliveira, S.L.; da Rocha, A. Very-low-carbohydrate ketogenic diet v.low-fat diet for long-term weight loss: A meta-analysis of randomised controlled trials. Br. J. Nutr. 2013, 110, 1178–1187. [Google Scholar] [CrossRef] [Green Version]
- Kwiterovich, P.O., Jr.; Vining, E.P.; Pyzik, P.; Skolasky, R., Jr.; Freeman, J.M. Effect of a High-Fat Ketogenic Diet on Plasma Levels of Lipids, Lipoproteins, and Apolipoproteins in Children. JAMA 2003, 290, 912–920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puchowicz, M.A.; Zechel, J.; Valerio, J.; Emancipator, D.; Xu, K.; Pundik, S.; LaManna, J.C.; Lust, D. Neuroprotection in Diet Induced Ketotic Rat Brain Following Focal Ischemia. J. Cereb. Blood Flow Metab. 2008, 12, 1907–1916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shiuchi, T.; Haque, M.S.; Okamoto, S.; Inoue, T.; Kageyama, H.; Lee, S. Hypothalamic orexin stimulates feeding-associated glucose utilization in skeletal muscle via sympathetic nervous system. Cell Metab. 2009, 10, 466–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chieffi, S.; Carotenuto, M.; Monda, V.; Valenzano, A.; Villano, I.; Precenzano, F. Orexin System: The Key for a Healthy Life. Front. Physiol. 2017, 8, 357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coborn, J.E.; Deporter, D.P.; Mavanji, V.; Sinton, C.M.; Kotz, C.M.; Billington, C.J. Role of orexin-a in the ventrolateral preoptic area on components of total energy expenditure. Int. J. Obes. 2017, 41, 1256–1262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, R.H.; Alexopoulos, H.; Jensen, L.T.; Fugger, L.; Burdakov, D. Adaptive sugar sensors in hypothalamic feeding circuits. Proc. Natl. Acad. Sci. USA 2008, 105, 11975–11980. [Google Scholar] [CrossRef] [Green Version]
- Furukawa, S.; Fujita, T.; Shimabujuro, M. Increased oxidative stress in obesity and its impact on metabolic syndrome. J. Clin. Investig. 2004, 114, 1752–1761. [Google Scholar] [CrossRef]
- Gletsu Miller, N.; Hansen, J.M.; Jones, D.P.; Go, Y.M.; Torres, W.E.; Ziegler, T.R.; Lin, E. Loss of Total and Visceral Adipose Tissue Mass Predicts Decreases in Oxidative Stress After Weight Loss Surgery. Obesity 2009, 3, 439–446. [Google Scholar] [CrossRef]
- Cancello, R.; Henegar, C.; Viguerie, N. Reduction of macrophage infiltration and chemoattractant gene expression changes in white adipose tissue of morbidly obese subjects after surgery-induced weight loss. Diabetes 2005, 54, 2277–2286. [Google Scholar] [CrossRef] [Green Version]
- Clement, K.; Viguerie, N.; Poitou, C. Weight loss regulates inflammation-related genes in white adipose tissue of obese subjects. FASEB J. 2004, 18, 1657–1669. [Google Scholar] [CrossRef] [Green Version]
- Ziegler, D.R.; Ribeiro, L.C.; Hagenn, M. Ketogenic diet increases glutathione peroxidase activity in rat hippocampus. Neurochem. Res. 2003, 28, 1793–1797. [Google Scholar] [CrossRef] [PubMed]
- Venugopal, R.; Jaiswal, A.K. Nrf1 and Nrf2 positively and c-Fos and Fra1 negatively regulate the human antioxidant response element-mediated expression of NAD(P)H:quinone oxidoreductase1 gene. Proc. Natl. Acad. Sci. USA 1996, 93, 14960–14965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greco, T.; Glenn, T.; Hoyda, D.; Prins, L.M. Ketogenic diet decreases oxidative stress and improves mitochondrial respiratory complex activity. J. Cereb. Blood Flow Metab. 2016, 36, 1603–1613. [Google Scholar] [CrossRef] [PubMed]
- Jornayvaz, F.R.; Jurczak, M.J.; Lee, H.-Y.; Birkenfeld, A.L.; Frederick, D.W.; Zhang, D.; Zhang, X.M.; Samuel, V.T.; Shulman, G.I. A high-fat, ketogenic diet causes hepatic insulin resistance in mice, despite increasing energy expenditure and preventing weight gain. Am. J. Physiol. Endocrinol. Metab. 2010, 299, E808–E815. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | VLCKD Obese Subjects | Statistical Analysis | |
|---|---|---|---|
| T0 | T1 | p-Value | |
| Age | 48 ± 8.2 | ns | |
| Height (m) | 1.67 ± 0.11 | ns | |
| Weight (kg) | 91.33 ± 17.11 | 78.73 ± 13.36 | <0.001 |
| BMI (kg/m2) | 32.19 ± 4.78 | 27.76 ± 3.62 | <0.001 |
| TOTAL CHOLESTEROL (mg/dL) | 220.13 ± 50.77 | 173.91 ± 32.93 | <0.05 |
| HDL (mg/dL) | 55.13 ± 11.14 | 47.76 ± 9.14 | ns |
| LDL (mg/dL) | 141.83 ± 36.48 | 107.57 ± 27.72 | <0.05 |
| TRIGLYCERIDES (mg/dL) | 135.54 ± 125.27 | 83.25 ± 26.14 | <0.05 |
| TOTAL BILIRUBIN (mg/dL) | 0.61 ± 0.22 | 0.68 ± 0.27 | ns |
| DIRECT BILIRUBIN (mg/dL) | 0.13 ± 0.18 | 0.16 ± 0.09 | ns |
| INDIRECT BILIRUBIN (mg/dL) | 0.48 ± 0.33 | 0.52 ± 0.21 | ns |
| HEMOGLOBIN (g/dL) | 14.13 ± 1.33 | 13.83 ± 0.94 | ns |
| GLYCATED HEMOGLOBIN (Hba1c) (%) | 5.65 ± 0.39 | 5.38 ± 0.33 | ns |
| INSULINEMIA (uUl/mL) | 10.53 ± 7.18 | 5.37 ± 3.79 | <0.05 |
| URIC ACID (mg/dL) | 4.86 ± 1.01 | 5.27 ± 1.13 | ns |
| TOTAL PROTEINS (g/dL) | 7.30 ± 0.4 | 7.13 ± 0.4 | ns |
| SERUM GLUTAMIC OXALOACETIC TRANSAMINASE (U/L) | 21.27 ± 5.98 | 23.31 ± 11.47 | <0.05 |
| SERUM GLUTAMIC PYRUVIC TRANSAMINASE (U/L) | 26.51 ± 14.89 | 26.06 ± 16.27 | <0.05 |
| GAMMA GLUTAMYL TRANSFERASE (U/L) | 31.19 ± 19.88 | 15.31 ± 5.41 | <0.05 |
| AZOTEMIA (mg/dL) | 35.35 ± 8.43 | 34.68 ± 9.16 | ns |
| CALCEMIA (mg/dL) | 9.57 ± 0.33 | 9.72 ± 0.35 | ns |
| SODIUM (mmol/L) | 139.19 ± 2.48 | 139.18 ± 2 | ns |
| C-REACTIVE PROTEIN (mg/mL) | 0.89 ± 0.1 | 0.48 ± 0.07 | <0.05 |
| Orexin-A (pg/mL) | 9.91 ± 0.27 | 16.24 ± 1.33 | <0.001 |
| Parameters | VLCKD Obese Subjects | Statistical Analysis | |
|---|---|---|---|
| T0 | T1 | p-Value | |
| Visceral Adipose Tissue (VAT) (g) | 1541.55 ± 141.63 | 927.79 ± 104.92 | <0.001 |
| Fat Mass (FM) (g) | 39,208.77± 1432.55 | 27377.0 ± 1217.48 | <0.001 |
| FFM (g) | 48,789.57± 1712.36 | 48093.68 ± 1670.65 | ns |
| BMD | 1225.57 ± 21.23 | 1229.31 ± 21.46 | ns |
| Parameters | VLCKD Obese Subjects | Statistical Analysis | |
|---|---|---|---|
| T0 | T1 | p-Value | |
| SYSTOLIC ARTERIAL PRESSURE | 130.6 | 117. 5 | <0.001 |
| DIASTOLIC ARTERIAL PRESSURE | 82. 8 | 75.8 | <0.001 |
| CARDIAC FREQUENCY PRE 6MWT | 77.7 | 74.5 | <0.001 |
| CARDIAC FREQUENCY POST 6MWT | 142.6 | 138.4 | <0.001 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valenzano, A.; Polito, R.; Trimigno, V.; Di Palma, A.; Moscatelli, F.; Corso, G.; Sessa, F.; Salerno, M.; Montana, A.; Di Nunno, N.; et al. Effects of Very Low Calorie Ketogenic Diet on the Orexinergic System, Visceral Adipose Tissue, and ROS Production. Antioxidants 2019, 8, 643. https://doi.org/10.3390/antiox8120643
Valenzano A, Polito R, Trimigno V, Di Palma A, Moscatelli F, Corso G, Sessa F, Salerno M, Montana A, Di Nunno N, et al. Effects of Very Low Calorie Ketogenic Diet on the Orexinergic System, Visceral Adipose Tissue, and ROS Production. Antioxidants. 2019; 8(12):643. https://doi.org/10.3390/antiox8120643
Chicago/Turabian StyleValenzano, Anna, Rita Polito, Valentina Trimigno, Antonella Di Palma, Fiorenzo Moscatelli, Gaetano Corso, Francesco Sessa, Monica Salerno, Angelo Montana, Nunzio Di Nunno, and et al. 2019. "Effects of Very Low Calorie Ketogenic Diet on the Orexinergic System, Visceral Adipose Tissue, and ROS Production" Antioxidants 8, no. 12: 643. https://doi.org/10.3390/antiox8120643
APA StyleValenzano, A., Polito, R., Trimigno, V., Di Palma, A., Moscatelli, F., Corso, G., Sessa, F., Salerno, M., Montana, A., Di Nunno, N., Astuto, M., Daniele, A., Carotenuto, M., Messina, G., Cibelli, G., & Monda, V. (2019). Effects of Very Low Calorie Ketogenic Diet on the Orexinergic System, Visceral Adipose Tissue, and ROS Production. Antioxidants, 8(12), 643. https://doi.org/10.3390/antiox8120643