Bisphenol S Impaired In Vitro Ovine Early Developmental Oocyte Competence

 ,
,
Abstract
:1. Introduction
2. Results
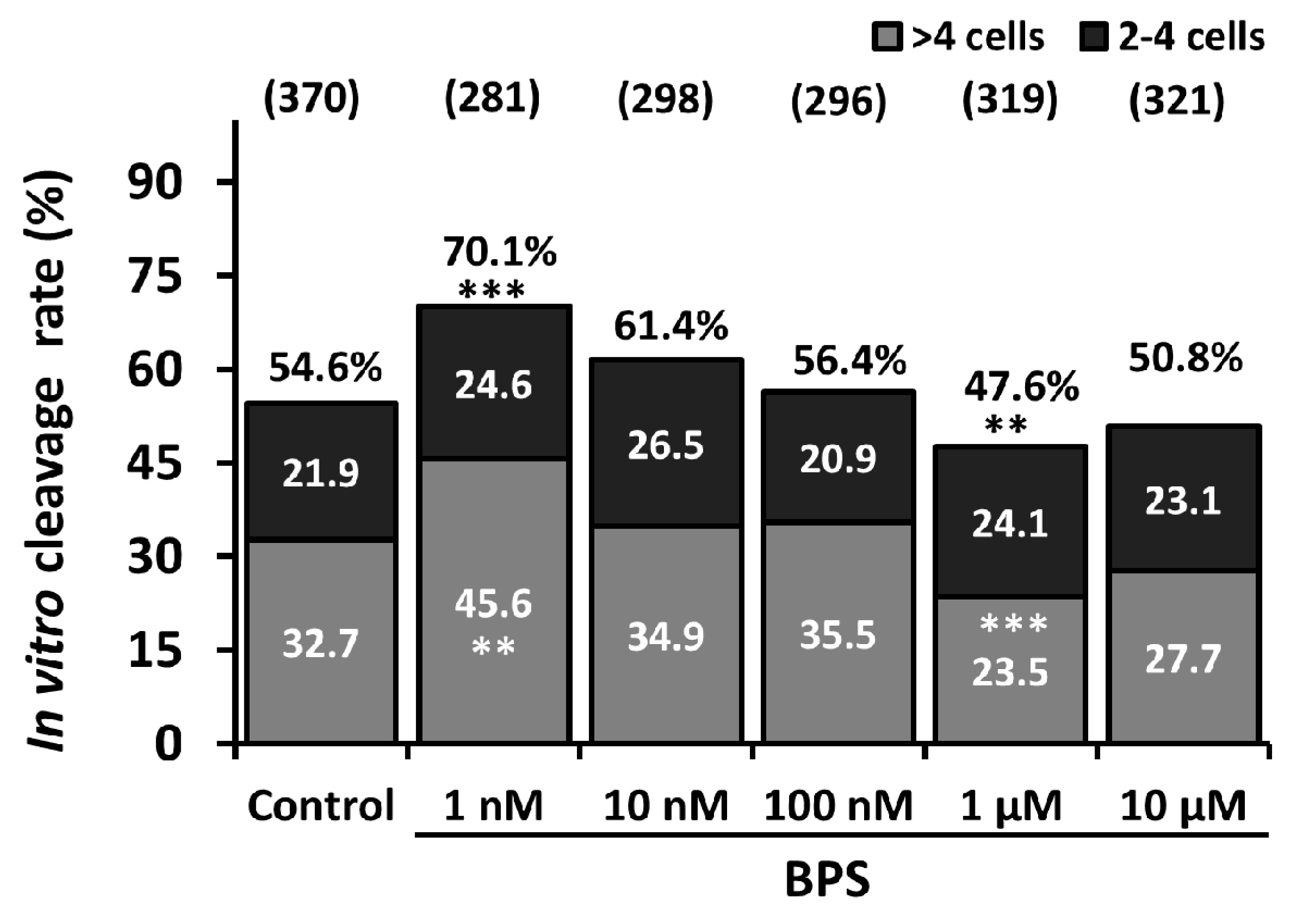
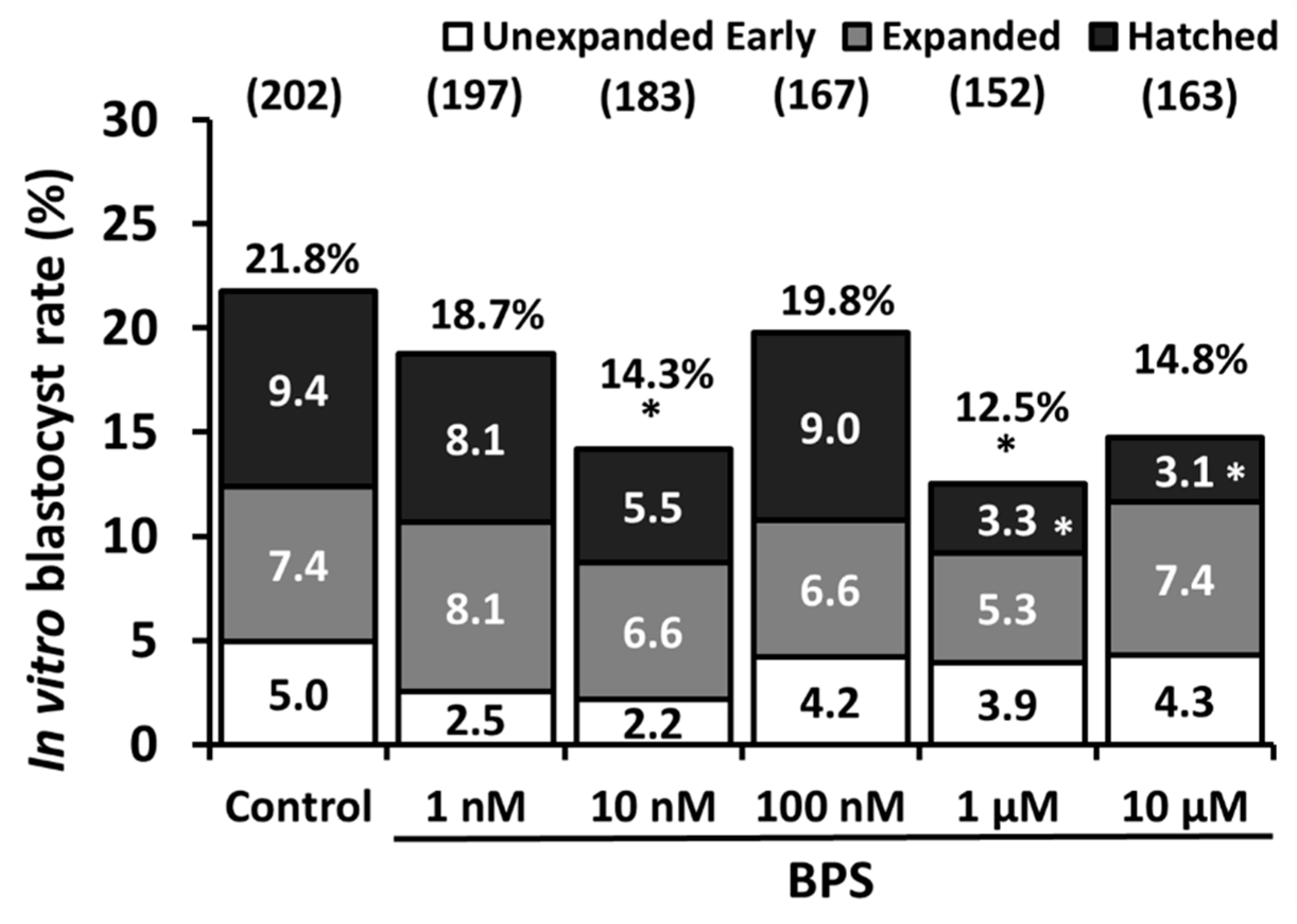
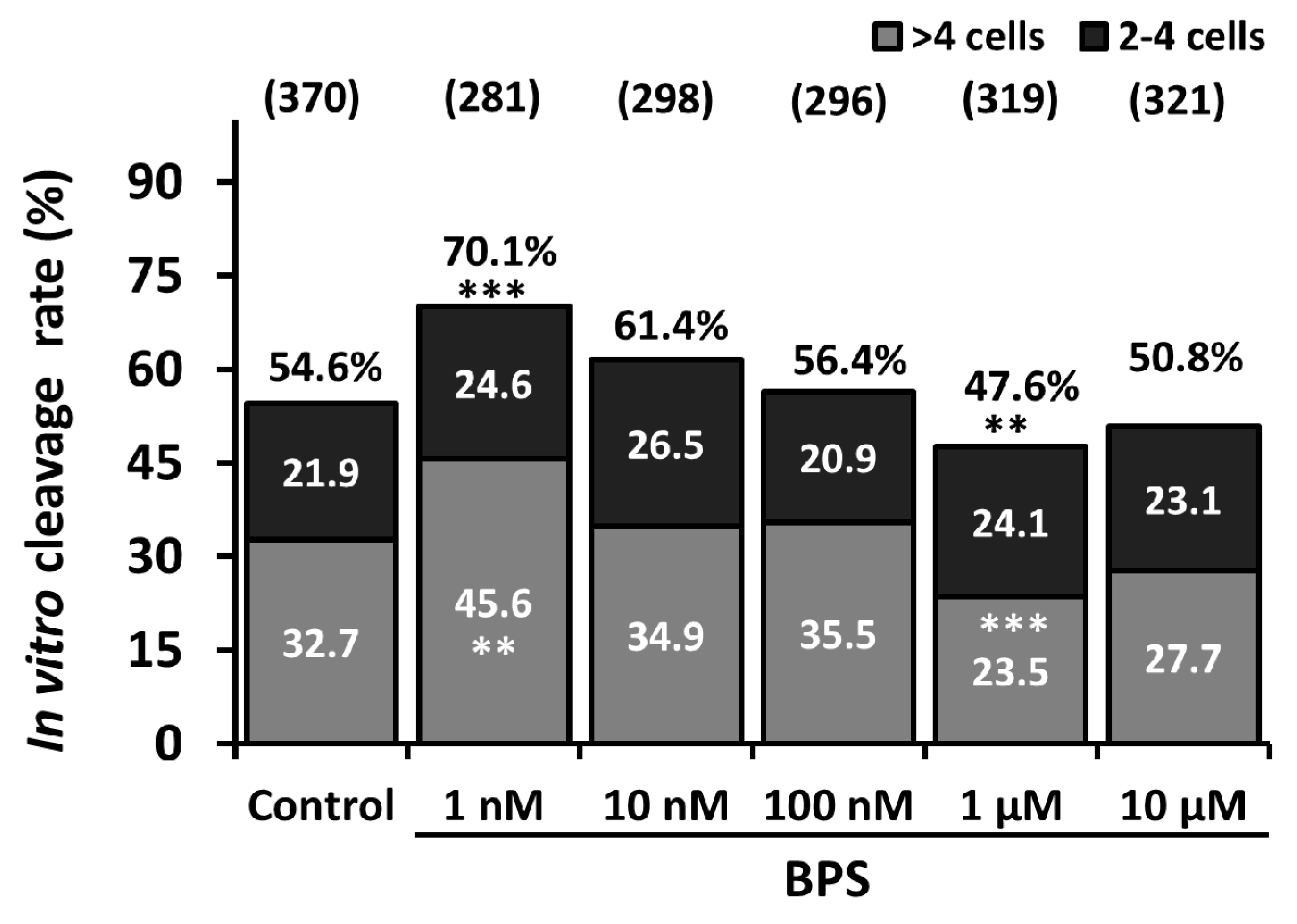
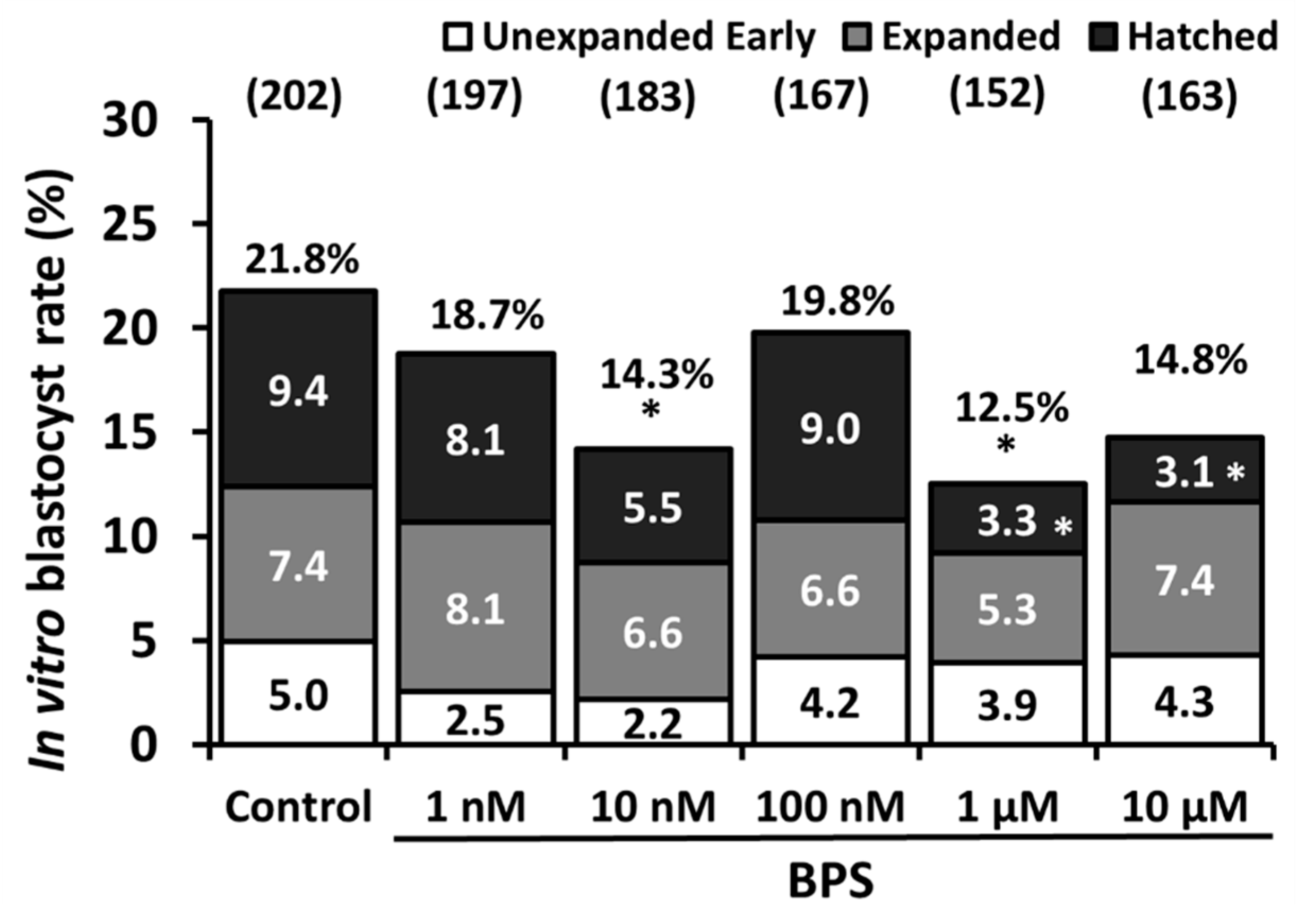
2.1. Low-Dose BPS Treatment During IVM and Effects on in vitro Embryo Development (IVD)
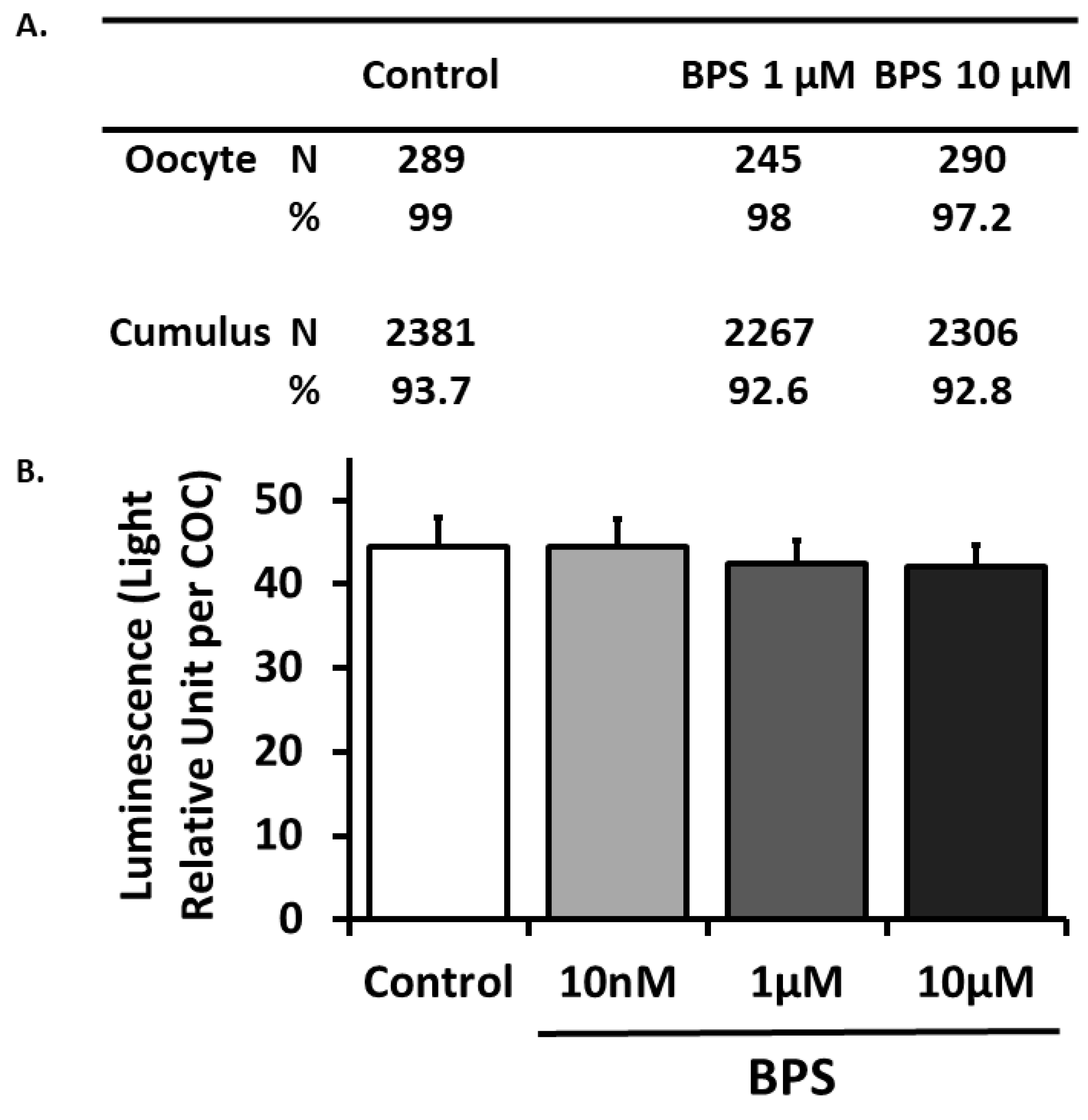
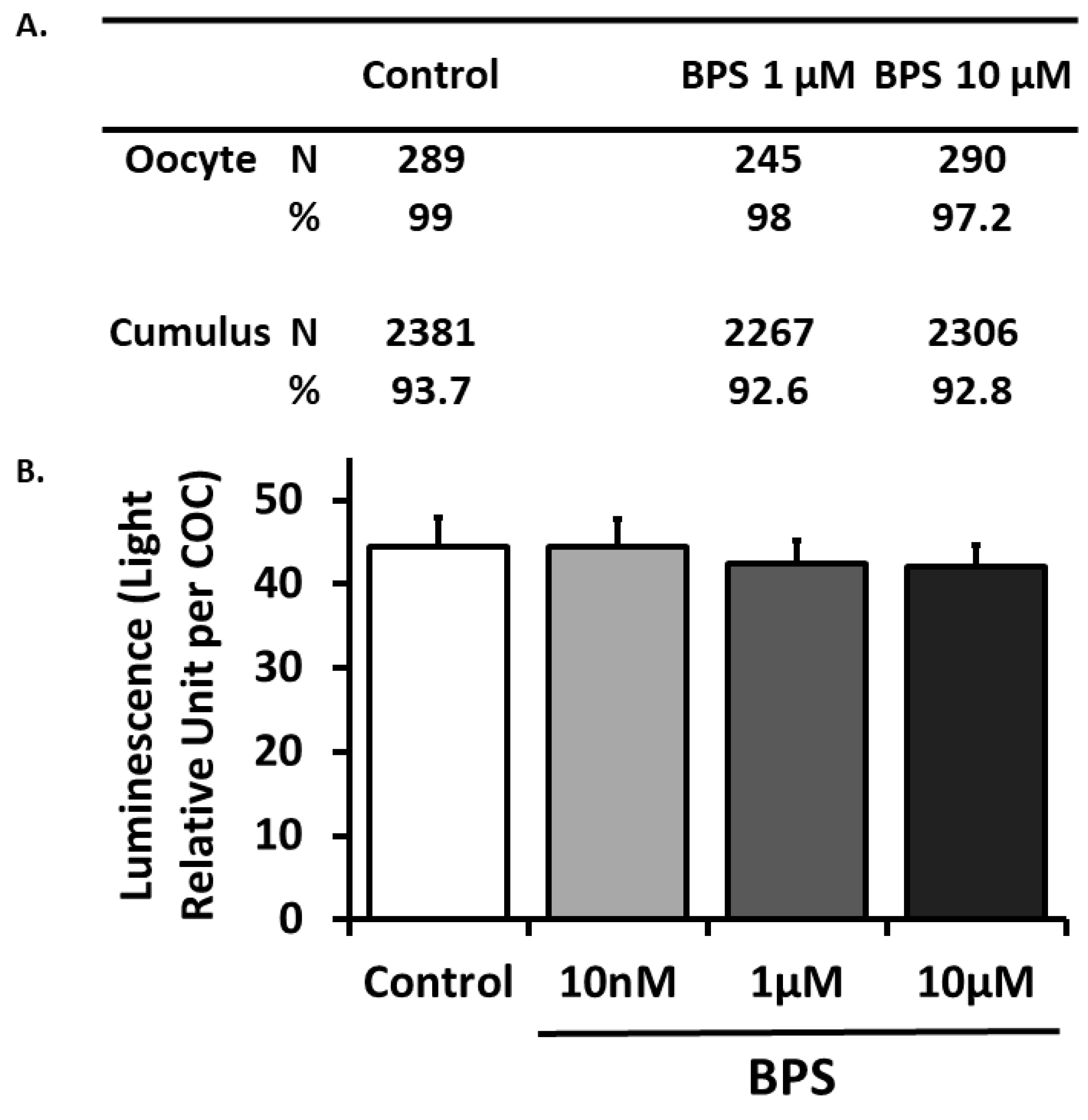
2.2. BPS Effect on COC Viability
2.3. BPS Effect on Oocyte Maturation
2.4. 24-h IVM BPS Treatment Effect on Day 6 Embryo Cell Number
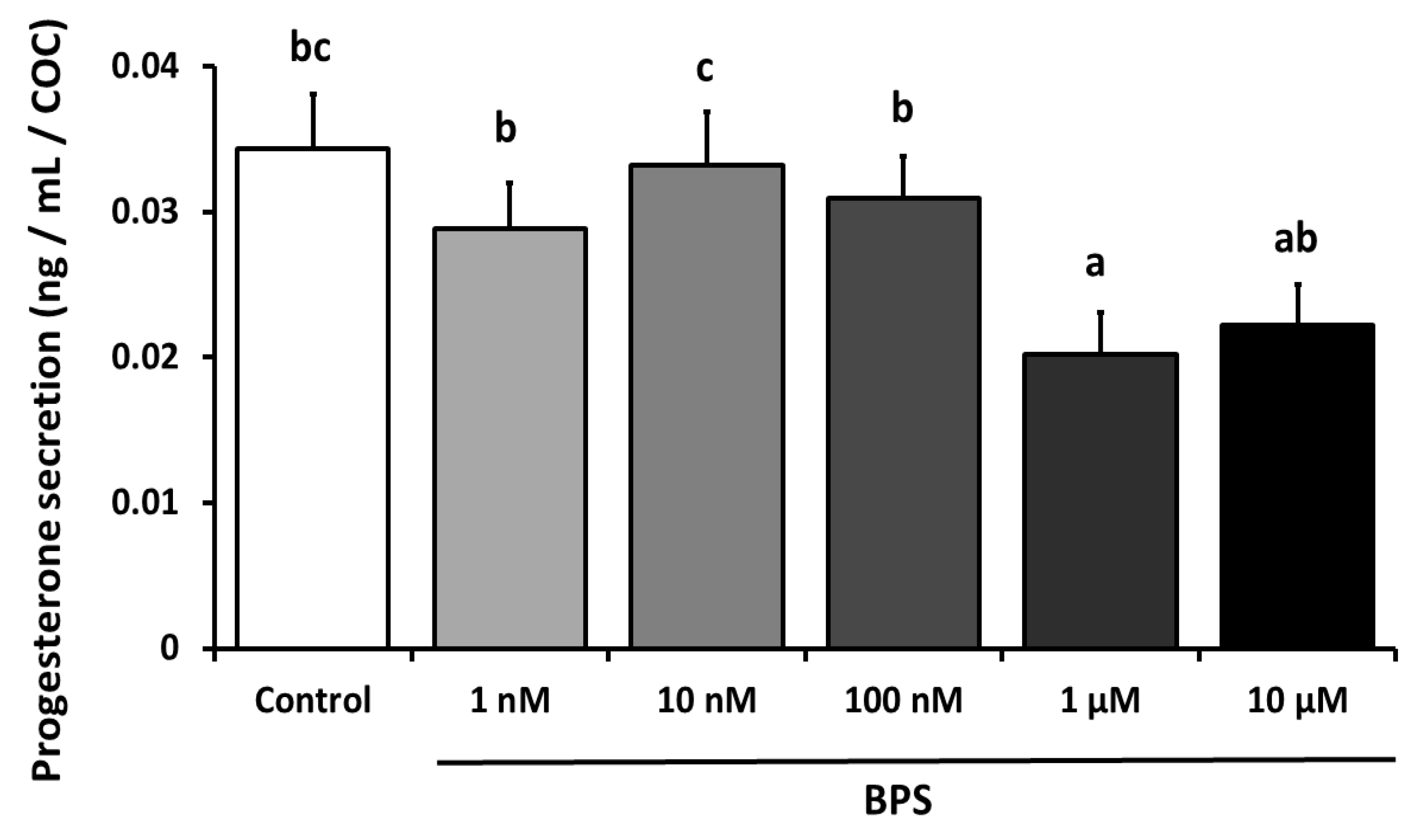
2.5. BPS Effect on COC Progesterone Secretion During 24-h IVM
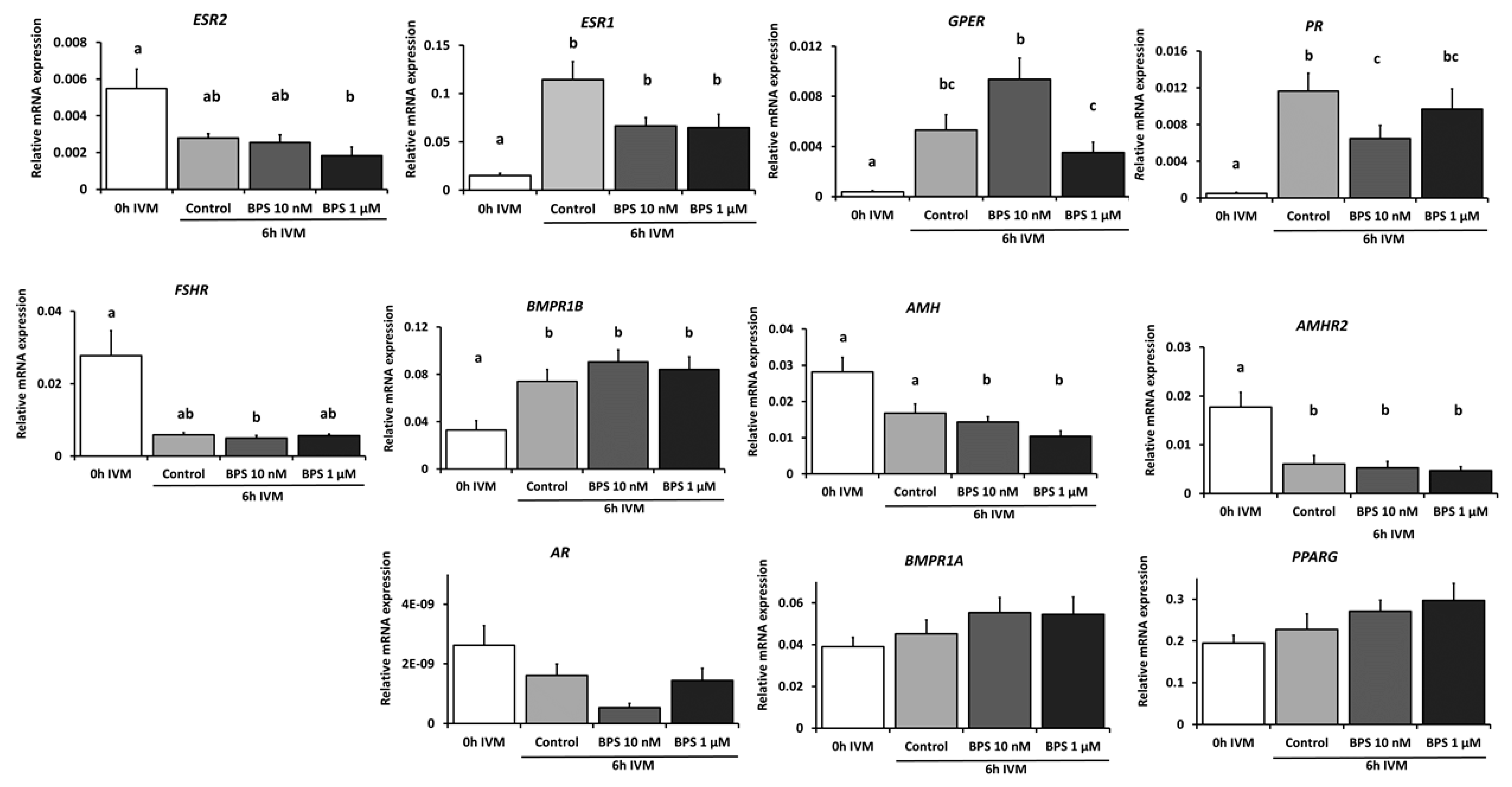
2.6. BPS Effect on Cumulus Cell mRNA Expression and on Oocyte MAPK3/1 Phosphorylation After 6 h IVM
3. Discussion
3.1. BPS Reduced Oocyte Nuclear Maturation
3.2. Low BPS Doses Impaired in vitro Ovine Early Developmental Oocyte Competence
3.3. BPS Reduced CC Progesterone Secretion
3.4. BPS Mechanism of Action
4. Materials and Methods
4.1. Ethics
4.2. Chemicals and Antibody
4.3. Biological Material
4.4. IVM
4.5. IVF and IVD
4.6. Analysis of Oocyte Nuclear Meiotic Maturation Stages
4.7. Cell Viability Analysis
4.8. Progesterone Assay
4.9. Gene Expression in CC
4.9.1. RNA Extraction and Reverse Transcription
4.9.2. qPCR Amplification
4.10. Western Blot Analysis
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Eladak, S.; Grisin, T.; Moison, D.; Guerquin, M.J.; N’Tumba-Byn, T.; Pozzi-Gaudin, S.; Benachi, A.; Livera, G.; Rouiller-Fabre, V.; Habert, R. A new chapter in the bisphenol A story: Bisphenol S and bisphenol F are not safe alternatives to this compound. Fertil. Steril. 2015, 103, 11–21. [Google Scholar] [CrossRef] [Green Version]
- Kang, J.H.; Kondo, F.; Katayama, Y. Human exposure to bisphenol A. Toxicology 2006, 226, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Kubwabo, C.; Kosarac, I.; Stewart, B.; Gauthier, B.; Lalonde, K.; Lalonde, P. Migration of bisphenol A from plastic baby bottles, baby bottle liners and reusable polycarbonate drinking bottles. Food Addit. Contam. 2009, 26, 928–937. [Google Scholar] [CrossRef] [PubMed]
- Calafat, A.M.; Kuklenyik, Z.; Reidy, J.A.; Caudill, S.P.; Ekong, J.; Needham, L.L. Urinary concentrations of bisphenol A and 4-nonylphenol in a human reference population. Environ. Health Perspect. 2005, 113, 391–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikezuki, Y.; Tsutsumi, O.; Takai, Y.; Kamei, Y.; Taketani, Y. Determination of bisphenol A concentrations in human biological fluids reveals significant early prenatal exposure. Hum. Reprod. 2002, 17, 2839–2841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez, M.F.; Arrebola, J.P.; Taoufiki, J.; Navalon, A.; Ballesteros, O.; Pulgar, R.; Vilchez, J.L.; Olea, N. Bisphenol-A and chlorinated derivatives in adipose tissue of women. Reprod. Toxicol. 2007, 24, 259–264. [Google Scholar] [CrossRef]
- Vandenberg, L.N.; Chahoud, I.; Heindel, J.J.; Padmanabhan, V.; Paumgartten, F.J.; Schoenfelder, G. Urinary, circulating, and tissue biomonitoring studies indicate widespread exposure to bisphenol A. Environ. Health Perspect. 2010, 118, 1055–1070. [Google Scholar] [CrossRef] [Green Version]
- Richter, C.A.; Birnbaum, L.S.; Farabollini, F.; Newbold, R.R.; Rubin, B.S.; Talsness, C.E.; Vandenbergh, J.G.; Walser-Kuntz, D.R.; vom Saal, F.S. In vivo effects of bisphenol A in laboratory rodent studies. Reprod. Toxicol. 2007, 24, 199–224. [Google Scholar] [CrossRef] [Green Version]
- Wetherill, Y.B.; Akingbemi, B.T.; Kanno, J.; McLachlan, J.A.; Nadal, A.; Sonnenschein, C.; Watson, C.S.; Zoeller, R.T.; Belcher, S.M. In vitro molecular mechanisms of bisphenol A action. Reprod. Toxicol. 2007, 24, 178–198. [Google Scholar] [CrossRef]
- Rochester, J.R. Bisphenol A and human health: A review of the literature. Reprod. Toxicol. 2013, 42, 132–155. [Google Scholar] [CrossRef]
- Lang, I.A.; Galloway, T.S.; Scarlett, A.; Henley, W.E.; Depledge, M.; Wallace, R.B.; Melzer, D. Association of urinary bisphenol A concentration with medical disorders and laboratory abnormalities in adults. JAMA 2008, 300, 1303–1310. [Google Scholar] [CrossRef] [PubMed]
- Grun, F.; Blumberg, B. Perturbed nuclear receptor signaling by environmental obesogens as emerging factors in the obesity crisis. Rev. Endocr. Metab. Disord. 2007, 8, 161–171. [Google Scholar] [CrossRef] [PubMed]
- Peretz, J.; Vrooman, L.; Ricke, W.A.; Hunt, P.A.; Ehrlich, S.; Hauser, R.; Padmanabhan, V.; Taylor, H.S.; Swan, S.H.; VandeVoort, C.A.; et al. Bisphenol A and reproductive health: Update of experimental and human evidence, 2007-2013. Environ. Health Perspect. 2014, 122, 775–786. [Google Scholar] [CrossRef] [PubMed]
- Nadal, A.; Fuentes, E.; Ripoll, C.; Villar-Pazos, S.; Castellano-Munoz, M.; Soriano, S.; Martinez-Pinna, J.; Quesada, I.; Alonso-Magdalena, P. Extranuclear-initiated estrogenic actions of endocrine disrupting chemicals: Is there toxicology beyond paracelsus? J. Steroid Biochem. Mol. Biol. 2018, 176, 16–22. [Google Scholar] [CrossRef]
- Kuiper, G.G.; Lemmen, J.G.; Carlsson, B.; Corton, J.C.; Safe, S.H.; Van Der Saag, P.T.; Van Der Burg, B.; Gustafsson, J.-A.K. Interaction of estrogenic chemicals and phytoestrogens with estrogen receptor β. Endocrinology 1998, 139, 4252–4263. [Google Scholar] [CrossRef]
- Machtinger, R.; Orvieto, R. Bisphenol A, oocyte maturation, implantation, and IVF outcome: Review of animal and human data. Reprod. Biomed. Online 2014, 29, 404–410. [Google Scholar] [CrossRef] [Green Version]
- Mok-Lin, E.; Ehrlich, S.; Williams, P.L.; Petrozza, J.; Wright, D.L.; Calafat, A.M.; Ye, X.; Hauser, R. Urinary bisphenol A concentrations and ovarian response among women undergoing IVF. Int. J. Androl. 2010, 33, 385–393. [Google Scholar] [CrossRef]
- Fujimoto, V.Y.; Kim, D.; vom Saal, F.S.; Lamb, J.D.; Taylor, J.A.; Bloom, M.S. Serum unconjugated bisphenol A concentrations in women may adversely influence oocyte quality during in vitro fertilization. Fertil. Steril. 2011, 95, 1816–1819. [Google Scholar] [CrossRef]
- Ehrlich, S.; Williams, P.L.; Missmer, S.A.; Flaws, J.A.; Ye, X.; Calafat, A.M.; Petrozza, J.C.; Wright, D.; Hauser, R. Urinary bisphenol A concentrations and early reproductive health outcomes among women undergoing IVF. Hum. Reprod. 2012, 27, 3583–3592. [Google Scholar] [CrossRef]
- Mlynarcikova, A.; Kolena, J.; Fickova, M.; Scsukova, S. Alterations in steroid hormone production by porcine ovarian granulosa cells caused by bisphenol A and bisphenol A dimethacrylate. Mol. Cell. Endocrinol. 2005, 244, 57–62. [Google Scholar] [CrossRef]
- Zhou, W.; Liu, J.; Liao, L.; Han, S.; Liu, J. Effect of bisphenol A on steroid hormone production in rat ovarian theca-interstitial and granulosa cells. Mol. Cell. Endocrinol. 2008, 283, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Grasselli, F.; Baratta, L.; Baioni, L.; Bussolati, S.; Ramoni, R.; Grolli, S.; Basini, G. Bisphenol A disrupts granulosa cell function. Domest. Anim. Endocrinol. 2010, 39, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Mlynarcikova, A.; Nagyova, E.; Fickova, M.; Scsukova, S. Effects of selected endocrine disruptors on meiotic maturation, cumulus expansion, synthesis of hyaluronan and progesterone by porcine oocyte-cumulus complexes. Toxicol. In Vitro 2009, 23, 371–377. [Google Scholar] [CrossRef] [PubMed]
- Machtinger, R.; Combelles, C.M.; Missmer, S.A.; Correia, K.F.; Williams, P.; Hauser, R.; Racowsky, C. Bisphenol-A and human oocyte maturation in vitro. Hum. Reprod. 2013, 28, 2735–2745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferris, J.; Favetta, L.A.; King, W.A. Bisphenol A Exposure during Oocyte Maturation in vitro Results in Spindle Abnormalities and Chromosome Misalignment in Bos taurus. Cytogenet. Genome Res. 2015, 145, 50–58. [Google Scholar] [CrossRef]
- EFSA Panel on Food Contact Materials; Enzymes, Flavourings and Processing Aids (CEF). Scientific Opinion on the risks to public health related to the presence of bisphenol A (BPA) in foodstuffs. EFSA J. 2015, 13. [Google Scholar] [CrossRef]
- Usman, A.; Ahmad, M. From BPA to its analogues: Is it a safe journey? Chemosphere 2016, 158, 131–142. [Google Scholar] [CrossRef]
- Liao, C.; Liu, F.; Alomirah, H.; Loi, V.D.; Mohd, M.A.; Moon, H.B.; Nakata, H.; Kannan, K. Bisphenol S in urine from the United States and seven Asian countries: Occurrence and human exposures. Environ. Sci. Technol. 2012, 46, 6860–6866. [Google Scholar] [CrossRef]
- Ji, K.; Hong, S.; Kho, Y.; Choi, K. Effects of bisphenol s exposure on endocrine functions and reproduction of zebrafish. Environ. Sci. Technol. 2013, 47, 8793–8800. [Google Scholar] [CrossRef]
- Rochester, J.R.; Bolden, A.L. Bisphenol S and F: A Systematic Review and Comparison of the Hormonal Activity of Bisphenol A Substitutes. Environ. Health Perspect. 2015, 123, 643–650. [Google Scholar] [CrossRef]
- Ullah, H.; Jahan, S.; Ain, Q.U.; Shaheen, G.; Ahsan, N. Effect of bisphenol S exposure on male reproductive system of rats: A histological and biochemical study. Chemosphere 2016, 152, 383–391. [Google Scholar] [CrossRef]
- Naderi, M.; Wong, M.Y.; Gholami, F. Developmental exposure of zebrafish (Danio rerio) to bisphenol-S impairs subsequent reproduction potential and hormonal balance in adults. Aquat. Toxicol. 2014, 148, 195–203. [Google Scholar] [CrossRef]
- Ahsan, N.; Ullah, H.; Ullah, W.; Jahan, S. Comparative effects of Bisphenol S and Bisphenol A on the development of female reproductive system in rats; a neonatal exposure study. Chemosphere 2018, 197, 336–343. [Google Scholar] [CrossRef]
- Nevoral, J.; Kolinko, Y.; Moravec, J.; Zalmanova, T.; Hoskova, K.; Prokesova, S.; Klein, P.; Ghaibour, K.; Hosek, P.; Stiavnicka, M.; et al. Long-term exposure to very low doses of bisphenol S affects female reproduction. Reproduction 2018, 156, 47–57. [Google Scholar] [CrossRef] [Green Version]
- Nourian, A.; Soleimanzadeh, A.; Jalali, A.S.; Najafi, G. Effects of bisphenol-S low concentrations on oxidative stress status and in vitro fertilization potential in mature female mice. Vet. Res. Forum 2017, 8, 341–345. [Google Scholar]
- Zalmanova, T.; Hoskova, K.; Nevoral, J.; Adamkova, K.; Kott, T.; Sulc, M.; Kotikova, Z.; Prokesova, S.; Jilek, F.; Kralickova, M.; et al. Bisphenol S negatively affects the meotic maturation of pig oocytes. Sci. Rep. 2017, 7, 485. [Google Scholar] [CrossRef]
- Campen, K.A.; Kucharczyk, K.M.; Bogin, B.; Ehrlich, J.M.; Combelles, C.M.H. Spindle abnormalities and chromosome misalignment in bovine oocytes after exposure to low doses of bisphenol A or bisphenol S. Hum. Reprod. 2018, 33, 895–904. [Google Scholar] [CrossRef]
- Corbel, T.; Gayrard, V.; Viguie, C.; Puel, S.; Lacroix, M.Z.; Toutain, P.L.; Picard-Hagen, N. Bisphenol A disposition in the sheep maternal-placental-fetal unit: Mechanisms determining fetal internal exposure. Biol. Reprod. 2013, 89, 11. [Google Scholar] [CrossRef] [Green Version]
- Grandin, F.C.; Lacroix, M.Z.; Gayrard, V.; Gauderat, G.; Mila, H.; Toutain, P.L.; Picard-Hagen, N. Bisphenol S instead of Bisphenol A: Toxicokinetic investigations in the ovine materno-feto-placental unit. Environ. Int. 2018, 120, 584–592. [Google Scholar] [CrossRef]
- Gingrich, J.; Pu, Y.; Ehrhardt, R.; Karthikraj, R.; Kannan, K.; Veiga-Lopez, A. Toxicokinetics of bisphenol A, bisphenol S, and bisphenol F in a pregnancy sheep model. Chemosphere 2019, 220, 185–194. [Google Scholar] [CrossRef]
- Scaramuzzi, R.J.; Baird, D.T.; Campbell, B.K.; Driancourt, M.A.; Dupont, J.; Fortune, J.E.; Gilchrist, R.B.; Martin, G.B.; McNatty, K.P.; McNeilly, A.S.; et al. Regulation of folliculogenesis and the determination of ovulation rate in ruminants. Reprod. Fertil. Dev. 2011, 23, 444–467. [Google Scholar] [CrossRef] [Green Version]
- Monniaux, D.; Clement, F.; Dalbies-Tran, R.; Estienne, A.; Fabre, S.; Mansanet, C.; Monget, P. The ovarian reserve of primordial follicles and the dynamic reserve of antral growing follicles: What is the link? Biol. Reprod. 2014, 90, 85. [Google Scholar] [CrossRef] [Green Version]
- Ye, X.; Wong, L.Y.; Kramer, J.; Zhou, X.; Jia, T.; Calafat, A.M. Urinary Concentrations of Bisphenol A and Three Other Bisphenols in Convenience Samples of U.S. Adults during 2000-2014. Environ. Sci. Technol. 2015, 49, 11834–11839. [Google Scholar] [CrossRef] [PubMed]
- Vandenberg, L.N.; Hunt, P.A.; Gore, A.C. Endocrine disruptors and the future of toxicology testing—lessons from CLARITY-BPA. Nat. Rev. Endocrinol. 2019, 15, 366–374. [Google Scholar] [CrossRef] [PubMed]
- Vandenberg, L.N.; Ehrlich, S.; Belcher, S.M.; Ben-Jonathan, N.; Dolinoy, D.C.; Hugo, E.R.; Hunt, P.A.; Newbold, R.R.; Rubin, B.S.; Saili, K.S.; et al. Low dose effects of bisphenol A. Endocr. Disruptors 2014, 1. [Google Scholar] [CrossRef]
- Campen, K.A.; McNatty, K.P.; Pitman, J.L. A protective role of cumulus cells after short-term exposure of rat cumulus cell-oocyte complexes to lifestyle or environmental contaminants. Reprod. Toxicol. 2017, 69, 19–33. [Google Scholar] [CrossRef]
- Carabatsos, M.J.; Sellitto, C.; Goodenough, D.A.; Albertini, D.F. Oocyte-Granulosa Cell Heterologous Gap Junctions Are Required for the Coordination of Nuclear and Cytoplasmic Meiotic Competence. Dev. Biol. 2000, 226, 167–179. [Google Scholar] [CrossRef] [Green Version]
- Can, A.; Semiz, O.; Cinar, O. Bisphenol-A induces cell cycle delay and alters centrosome and spindle microtubular organization in oocytes during meiosis. Mol. Hum. Reprod. 2005, 11, 389–396. [Google Scholar] [CrossRef]
- Eichenlaub-Ritter, U.; Vogt, E.; Cukurcam, S.; Sun, F.; Pacchierotti, F.; Parry, J. Exposure of mouse oocytes to bisphenol A causes meiotic arrest but not aneuploidy. Mutat. Res. 2008, 651, 82–92. [Google Scholar] [CrossRef]
- Lenie, S.; Cortvrindt, R.; Eichenlaub-Ritter, U.; Smitz, J. Continuous exposure to bisphenol A during in vitro follicular development induces meiotic abnormalities. Mutat. Res. 2008, 651, 71–81. [Google Scholar] [CrossRef]
- Wang, X.; Jiang, S.W.; Wang, L.; Sun, Y.; Xu, F.; He, H.; Wang, S.; Zhang, Z.; Pan, X. Interfering effects of bisphenol A on in vitro growth of preantral follicles and maturation of oocyes. Clin. Chim. Acta 2018, 485, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Han, J.; Duan, X.; Xiong, B.; Cui, X.S.; Kim, N.H.; Liu, H.L.; Sun, S.C. The toxic effects and possible mechanisms of Bisphenol A on oocyte maturation of porcine in vitro. Oncotarget 2016, 7, 32554–32565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chao, H.H.; Zhang, X.F.; Chen, B.; Pan, B.; Zhang, L.J.; Li, L.; Sun, X.F.; Shi, Q.H.; Shen, W. Bisphenol A exposure modifies methylation of imprinted genes in mouse oocytes via the estrogen receptor signaling pathway. Histochem. Cell Biol. 2012, 137, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Susiarjo, M.; Hassold, T.J.; Freeman, E.; Hunt, P.A. Bisphenol A exposure in utero disrupts early oogenesis in the mouse. PLoS Genet. 2007, 3, e5. [Google Scholar] [CrossRef]
- Ferris, J.; Mahboubi, K.; MacLusky, N.; King, W.A.; Favetta, L.A. BPA exposure during in vitro oocyte maturation results in dose-dependent alterations to embryo development rates, apoptosis rate, sex ratio and gene expression. Reprod. Toxicol. 2016, 59, 128–138. [Google Scholar] [CrossRef]
- Arancio, A.L.; Cole, K.D.; Dominguez, A.R.; Cohenour, E.R.; Kadie, J.; Maloney, W.C.; Cilliers, C.; Schuh, S.M. Bisphenol A, Bisphenol AF, di-n-butyl phthalate, and 17beta-estradiol have shared and unique dose-dependent effects on early embryo cleavage divisions and development in Xenopus laevis. Reprod. Toxicol. 2019, 84, 65–74. [Google Scholar] [CrossRef]
- Guo, J.; Zhao, M.H.; Shin, K.T.; Niu, Y.J.; Ahn, Y.D.; Kim, N.H.; Cui, X.S. The possible molecular mechanisms of bisphenol A action on porcine early embryonic development. Sci. Rep. 2017, 7, 8632. [Google Scholar] [CrossRef]
- Shen, J.; Kang, Q.; Mao, Y.; Yuan, M.; Le, F.; Yang, X.; Xu, X.; Jin, F. Urinary bisphenol A concentration is correlated with poorer oocyte retrieval and embryo implantation outcomes in patients with tubal factor infertility undergoing in vitro fertilisation. Ecotoxicol. Environ. Saf. 2020, 187, 109816. [Google Scholar] [CrossRef]
- Vandenberg, L.N.; Hauser, R.; Marcus, M.; Olea, N.; Welshons, W.V. Human exposure to bisphenol A (BPA). Reprod. Toxicol. 2007, 24, 139–177. [Google Scholar] [CrossRef]
- Thayer, K.A.; Taylor, K.W.; Garantziotis, S.; Schurman, S.H.; Kissling, G.E.; Hunt, D.; Herbert, B.; Church, R.; Jankowich, R.; Churchwell, M.I.; et al. Bisphenol A, Bisphenol S, and 4-Hydroxyphenyl 4-Isoprooxyphenylsulfone (BPSIP) in Urine and Blood of Cashiers. Environ. Health Perspect. 2016, 124, 437–444. [Google Scholar] [CrossRef]
- Berni, M.; Gigante, P.; Bussolati, S.; Grasselli, F.; Grolli, S.; Ramoni, R.; Basini, G. Bisphenol S, a Bisphenol A alternative, impairs swine ovarian and adipose cell functions. Domest. Anim. Endocrinol. 2019, 66, 48–56. [Google Scholar] [CrossRef]
- Campen, K.A.; Lavallee, M.; Combelles, C. The impact of bisphenol S on bovine granulosa and theca cells. Reprod. Domest. Anim. 2018, 53, 450–457. [Google Scholar] [CrossRef]
- Pogrmic-Majkic, K.; Samardzija Nenadov, D.; Fa, S.; Stanic, B.; Trninic Pjevic, A.; Andric, N. BPA activates EGFR and ERK1/2 through PPARgamma to increase expression of steroidogenic acute regulatory protein in human cumulus granulosa cells. Chemosphere 2019, 229, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Bujnakova Mlynarcikova, A.; Scsukova, S. Simultaneous effects of endocrine disruptor bisphenol A and flavonoid fisetin on progesterone production by granulosa cells. Environ. Toxicol. Pharm. 2018, 59, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Mansur, A.; Adir, M.; Yerushalmi, G.; Hourvitz, A.; Gitman, H.; Yung, Y.; Orvieto, R.; Machtinger, R. Does BPA alter steroid hormone synthesis in human granulosa cells in vitro? Hum. Reprod. 2016, 31, 1562–1569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banerjee, O.; Singh, S.; Prasad, S.K.; Bhattacharjee, A.; Banerjee, A.; Banerjee, A.; Saha, A.; Maji, B.K.; Mukherjee, S. Inhibition of catalase activity with 3-amino-1,2,4-triazole intensifies bisphenol A (BPA)-induced toxicity in granulosa cells of female albino rats. Toxicol. Ind. Health 2018, 34, 787–797. [Google Scholar] [CrossRef]
- Li, Q.; Davila, J.; Kannan, A.; Flaws, J.A.; Bagchi, M.K.; Bagchi, I.C. Chronic Exposure to Bisphenol A Affects Uterine Function During Early Pregnancy in Mice. Endocrinology 2016, 157, 1764–1774. [Google Scholar] [CrossRef] [Green Version]
- Fan, H.-Y.; Sun, Q.-Y. Involvement of mitogen-activated protein kinase cascade during oocyte maturation and fertilization in mammals. Biol. Reprod. 2004, 70, 535–547. [Google Scholar] [CrossRef]
- Gordo, A.C.; He, C.L.; Smith, S.; Fissore, R.A. Mitogen activated protein kinase plays a significant role in metaphase II arrest, spindle morphology, and maintenance of maturation promoting factor activity in bovine oocytes. Mol. Reprod. Dev. Inc. Gamete Res. 2001, 59, 106–114. [Google Scholar] [CrossRef]
- Liang, C.-G.; Su, Y.-Q.; Fan, H.-Y.; Schatten, H.; Sun, Q.-Y. Mechanisms regulating oocyte meiotic resumption: Roles of mitogen-activated protein kinase. Mol. Endocrinol. 2007, 21, 2037–2055. [Google Scholar] [CrossRef]
- Ptak, A.; Gregoraszczuk, E.L. Bisphenol A induces leptin receptor expression, creating more binding sites for leptin, and activates the JAK/Stat, MAPK/ERK and PI3K/Akt signalling pathways in human ovarian cancer cell. Toxicol. Lett. 2012, 210, 332–337. [Google Scholar] [CrossRef]
- Chu, P.W.; Yang, Z.J.; Huang, H.H.; Chang, A.A.; Cheng, Y.C.; Wu, G.J.; Lan, H.C. Low-dose bisphenol A activates the ERK signaling pathway and attenuates steroidogenic gene expression in human placental cells. Biol. Reprod. 2018, 98, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Melnick, R.; Lucier, G.; Wolfe, M.; Hall, R.; Stancel, G.; Prins, G.; Gallo, M.; Reuhl, K.; Ho, S.M.; Brown, T.; et al. Summary of the National Toxicology Program’s report of the endocrine disruptors low-dose peer review. Environ. Health Perspect. 2002, 110, 427–431. [Google Scholar] [CrossRef] [PubMed]
- Andrade, A.J.; Grande, S.W.; Talsness, C.E.; Gericke, C.; Grote, K.; Golombiewski, A.; Sterner-Kock, A.; Chahoud, I. A dose response study following in utero and lactational exposure to di-(2-ethylhexyl) phthalate (DEHP): Reproductive effects on adult male offspring rats. Toxicology 2006, 228, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Awada, Z.; Sleiman, F.; Mailhac, A.; Mouneimne, Y.; Tamim, H.; Zgheib, N.K. BPA exposure is associated with non-monotonic alteration in ESR1 promoter methylation in peripheral blood of men and shorter relative telomere length in peripheral blood of women. J. Expo. Sci. Environ. Epidemiol. 2019, 29, 118–128. [Google Scholar] [CrossRef] [PubMed]
- Villar-Pazos, S.; Martinez-Pinna, J.; Castellano-Munoz, M.; Alonso-Magdalena, P.; Marroqui, L.; Quesada, I.; Gustafsson, J.A.; Nadal, A. Molecular mechanisms involved in the non-monotonic effect of bisphenol-a on ca2+ entry in mouse pancreatic beta-cells. Sci. Rep. 2017, 7, 11770. [Google Scholar] [CrossRef]
- Tervit, H.R.; Whittingham, D.G.; Rowson, L.E.A. Successful Culture in Vitro of Sheep and Cattle Ova. Reproduction 1972, 30, 493–497. [Google Scholar] [CrossRef]
- Crocomo, L.F.; Marques Filho, W.C.; Ackermann, C.L.; Paschoal, D.M.; Guastali, M.D.; Dias Maziero, R.R.; Sudano, M.J.; Landim-Alvarenga Fda, C.; Bicudo, S.D. Time course of the meiotic arrest in sheep cumulus-oocyte complexes treated with roscovitine. Zygote 2016, 24, 310–318. [Google Scholar] [CrossRef]
- Dhali, A.; Javvaji, P.K.; Kolte, A.P.; Francis, J.R.; Roy, S.C.; Sejian, V. Temporal expression of cumulus cell marker genes during in vitro maturation and oocyte developmental competence. J. Assist. Reprod. Genet. 2017, 34, 1493–1500. [Google Scholar] [CrossRef]
- Canépa, S.; Lainé, A.-L.; Bluteau, A.; Fagu, C.; Flon, C.; Monniaux, D. Validation d’une méthode immunoenzymatique pour le dosage de la progestérone dans le plasma des ovins et des bovins. Cahier des Techniques de l’INRA 2008, 64, 19–30. [Google Scholar]
- Elis, S.; Oseikria, M.; Vitorino Carvalho, A.; Bertevello, P.S.; Corbin, E.; Teixeira-Gomes, A.P.; Lecardonnel, J.; Archilla, C.; Duranthon, V.; Labas, V.; et al. Docosahexaenoic acid mechanisms of action on the bovine oocyte-cumulus complex. J. Ovarian Res. 2017, 10, 74. [Google Scholar] [CrossRef] [PubMed]
- Maillard, V.; Desmarchais, A.; Durcin, M.; Uzbekova, S.; Elis, S. Docosahexaenoic acid (DHA) effects on proliferation and steroidogenesis of bovine granulosa cells. Reprod. Biol. Endocrinol. 2018, 16, 40. [Google Scholar] [CrossRef] [PubMed]
- Fox, J. Getting started with the R commander: A basic-statistics graphical user interface to R. J. Stat. Softw. 2005, 14, 1–42. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing (Version 0.97. 316); R Foundation for Statistical Computing: Vienna, Austria, 2015. [Google Scholar]
- Lenth, R.V. Least-Squares Means: TheRPackagelsmeans. J. Stat. Softw. 2016, 69. [Google Scholar] [CrossRef] [Green Version]
- Wheeler, B.; Torchiano, M. lmPerm: Permutation Tests for Linear Models; R Package Version 1; The R project for statistical computing. Available online: http://www.r-project.org/ (accessed on 1 February 2020).
- Konietschke, F.; Placzek, M.; Schaarschmidt, F.; Hothorn, L.A. Nparcomp: An R Software Package for Nonparametric Multiple Comparisons and Simultaneous Confidence Intervals. J. Stat. Softw. 2015, 64. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N Oocyte | Mature Oocyte % | |
|---|---|---|
| Control | 152 | 88.2% ± 2.6% |
| BPS 1 nM | 117 | 87.2% ± 3.1% |
| BPS 1 µM | 117 | 82.8% ± 3.5% |
| BPS 10 µM | 171 | 76.7% ± 3.2% * |
| N Embryo | Cell Number | |
|---|---|---|
| Control | 70 | 28.3 ± 2.9 |
| BPS 10 nM | 75 | 33.1 ± 3.4 |
| BPS 1 µM | 63 | 22.2 ± 2.3 |
| Abbreviation | Gene | Forward Primer | Reverse Primer | bp | E |
|---|---|---|---|---|---|
| FSHR | Follicle stimulating hormone receptor | 5′-GGGCCAAGTCAACTTACCACT-3′ | 5-TGCAAATTGGATGAAGGTCA-3′ | 144 | 1.89 |
| AMHR2 | Anti-Mullerian Hormone Receptor Type 2 | 5′-GAAAAAGGGCCTTGCTGAA-3′ | 5′-CAGGACTGCTCACCTTGCT-3′ | 113 | 1.83 |
| PPARG | Peroxisome proliferator-activated receptor gamma | 5′-ATGTCCTCAATGGGCTTCAC-3′ | 5′-GTGAAGTTCAACGCACTGGA-3′ | 113 | 1.87 |
| ESR1 | Estrogen receptor 1 | 5′-GGTTCCGTATGATGAATCT-3′ | 5′-CAAGGTGTCTGTGATCTT-3′ | 158 | 1.96 |
| ESR2 | Estrogen receptor 2 | 5′-GTCGGTTCTTATCTATGGTA-3′ | 5′-ACTATGGAGTCTGGTCAT-3′ | 114 | 1.99 |
| PR | Progesterone receptor | 5′-AGTCATCATTCTATTCATTATGC-3′ | 5′-TGGCTTCTTAGTCCTTCT-3′ | 142 | 1.98 |
| AR | Androgen receptor | 5′-CCTTCACCAATGTCAACT-3′ | 5′-ATCCACTGGAATAATGCTAA-3′ | 200 | 1.90 |
| GPER | G protein coupled estrogen receptor 1 | 5′-TCCCCGACCTGTACTTCATC-3′ | 5′-GAGGAAGAAGACGCTGCTGT-3′ | 167 | 1.98 |
| BMPR1A | Bone morphogenetic protein receptor type 1A | 5′-TGTCGGACCAACTTATGTAACC-3′ | 5′-TGAGCAAAGCCAGCCATCG-3′ | 100 | 1.92 |
| BMPR1B | Bone morphogenetic protein receptor type 1B | 5′-TCTTGAGGCAGGATTGTGAGC-3′ | 5′-GGTGGAGCAGTGACGAGTG-3′ | 77 | 1.93 |
| AMH | Anti-müllerian hormone | 5′-GTGGTGCTGCTGCTAAAGATG-3′ | 5′-TCGGACAGGCTGATGAGGAG-3′ | 104 | 1.88 |
| ACTB | Beta Actin | 5′-CCAGCACGATGAAGATCAAG-3′ | 5′-ACATCTGCTGGAAGGTGGAC-3′ | 102 | 1.97 |
| RPL19 | Ribosomal Protein L19 | 5′-CACAAGCTGAAGGCAGACAA-3′ | 5′-TGATGATTTCCTCCTTCTTGG-3′ | 129 | 1.94 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Desmarchais, A.; Téteau, O.; Papillier, P.; Jaubert, M.; Druart, X.; Binet, A.; Maillard, V.; Elis, S. Bisphenol S Impaired In Vitro Ovine Early Developmental Oocyte Competence. Int. J. Mol. Sci. 2020, 21, 1238. https://doi.org/10.3390/ijms21041238
Desmarchais A, Téteau O, Papillier P, Jaubert M, Druart X, Binet A, Maillard V, Elis S. Bisphenol S Impaired In Vitro Ovine Early Developmental Oocyte Competence. International Journal of Molecular Sciences. 2020; 21(4):1238. https://doi.org/10.3390/ijms21041238
Chicago/Turabian StyleDesmarchais, Alice, Ophélie Téteau, Pascal Papillier, Manon Jaubert, Xavier Druart, Aurélien Binet, Virginie Maillard, and Sebastien Elis. 2020. "Bisphenol S Impaired In Vitro Ovine Early Developmental Oocyte Competence" International Journal of Molecular Sciences 21, no. 4: 1238. https://doi.org/10.3390/ijms21041238
APA StyleDesmarchais, A., Téteau, O., Papillier, P., Jaubert, M., Druart, X., Binet, A., Maillard, V., & Elis, S. (2020). Bisphenol S Impaired In Vitro Ovine Early Developmental Oocyte Competence. International Journal of Molecular Sciences, 21(4), 1238. https://doi.org/10.3390/ijms21041238