Exploring the Potential of Mesenchymal Stem Cell-Based Therapy in Mouse Models of Vascular Cognitive Impairment

 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
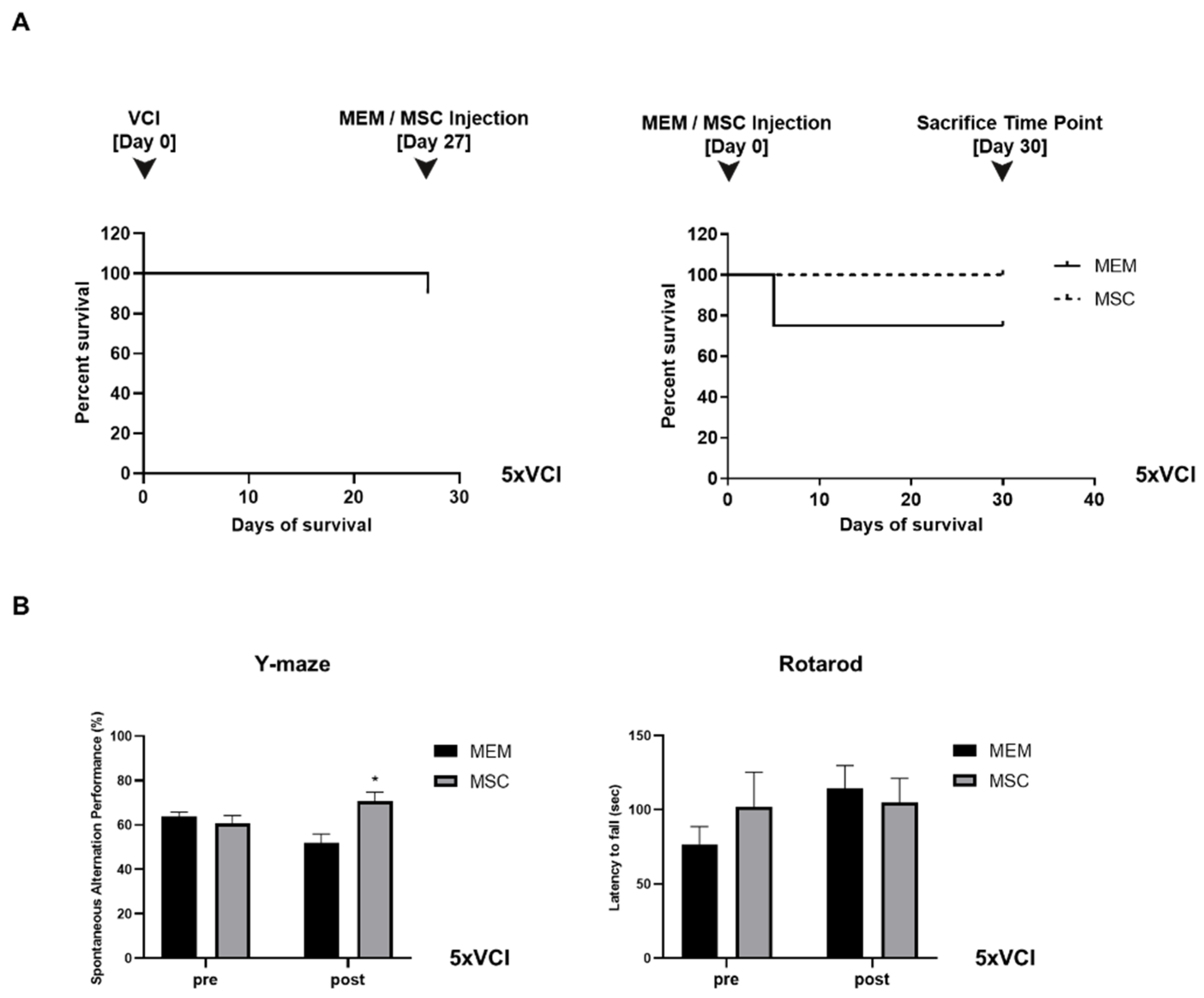
2.1. Experiment #1: Unexpectedly, the Survival of C57VCI Mice Was Decreased Following Transplantation of Human MSCs
2.2. Experiment #1: Spatial Working Memory and Motor Coordination Were Not Affected in C57VCI Mice after Transplantation of Human MSCs
2.3. Experiment #1: C57VCI Mice Display Heterogeneity in Pathological Manifestation
2.4. Experiment #1: Cell Aggregates Are Identified from Several C57VCI Mice that Received MSC Injections
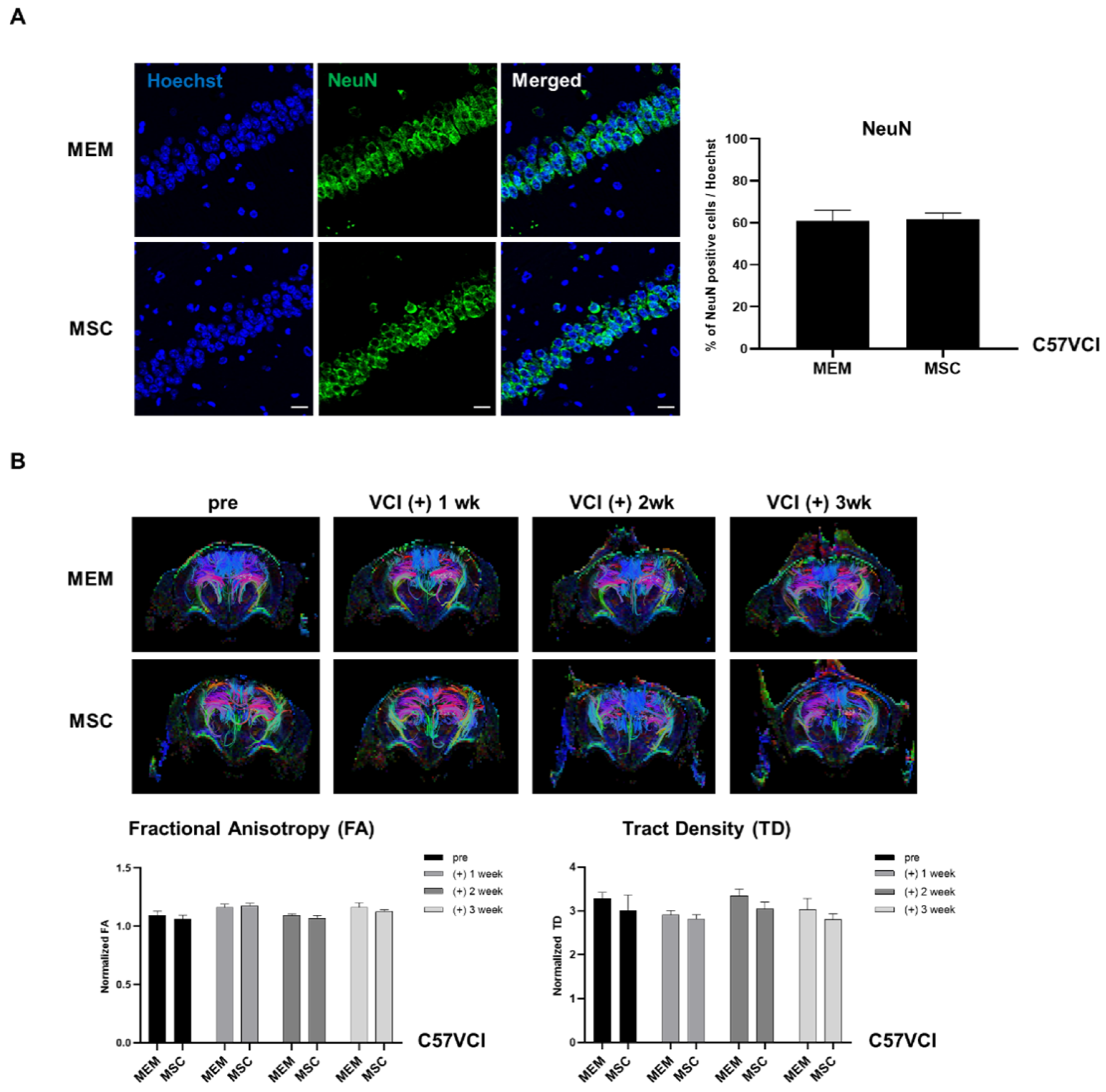
2.5. Experiment #1: Neuronal Density and White Matter Tracts of C57VCI Mice Are Unaffected after MSC Injection
2.6. Experiment #2: Mortality Was Not Observed from the MSC-Injected 5xVCI Mice
2.7. Spatial Working Memory is Enhanced Following Injections of MSCs in 5xVCI Mice
2.8. 5xVCI Mice Display Subtle Changes in Disease Progression
2.9. Amyloid Beta Levels of 5xVCI Mice Are Altered in the Thalamus Following MSC Injection
3. Discussion
4. Materials and Methods
4.1. Ethical Statement
4.2. Experimental Animals and Study Design
4.3. VCI Surgery
4.4. Culture and Preparation of Human MSCs
4.5. Transplantation of MSCs into the Lateral Ventricle of Mice
4.6. Behavior
4.7. MR Imaging and Fiber Tractography
4.8. Immunohistochemistry
4.9. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Jiwa, N.S.; Garrard, P.; Hainsworth, A.H. Experimental models of vascular dementia and vascular cognitive impairment: A systematic review. J. Neurochem. 2010, 115, 814–828. [Google Scholar] [CrossRef]
- Attems, J.; Jellinger, K.A. The overlap between vascular disease and Alzheimer’s disease—Lessons from pathology. BMC Med. 2014, 12, 206. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Kim, S.H.; Kim, G.H.; Seo, S.W.; Park, H.K.; Oh, S.J.; Kim, J.S.; Cheong, H.K.; Na, D.L. Identification of pure subcortical vascular dementia using 11C-Pittsburgh compound B. Neurology 2011, 77, 18–25. [Google Scholar] [CrossRef]
- Iadecola, C. The pathobiology of vascular dementia. Neuron 2013, 80, 844–866. [Google Scholar] [CrossRef] [Green Version]
- Meguro, K.; Dodge, H.H. Vascular Mild Cognitive Impairment: Identifying Disease in Community-Dwelling Older Adults, Reducing Risk Factors, and Providing Support. The Osaki-Tajiri and Kurihara Projects. J. Alzheimers Dis. 2019, 70 (Suppl. 1), S293–S302. [Google Scholar] [CrossRef] [Green Version]
- Iadecola, C.; Duering, M.; Hachinski, V.; Joutel, A.; Pendlebury, S.T.; Schneider, J.A.; Dichgans, M. Vascular Cognitive Impairment and Dementia: JACC Scientific Expert Panel. J. Am. Coll. Cardiol. 2019, 73, 3326–3344. [Google Scholar] [CrossRef] [PubMed]
- Gooch, J.; Wilcock, D.M. Animal Models of Vascular Cognitive Impairment and Dementia (VCID). Cell. Mol. Neurobiol. 2016, 36, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Madigan, J.B.; Wilcock, D.M.; Hainsworth, A.H. Vascular Contributions to Cognitive Impairment and Dementia: Topical Review of Animal Models. Stroke 2016, 47, 1953–1959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Kimura-Ohba, S.; Thompson, J.; Rosenberg, G.A. Rodent Models of Vascular Cognitive Impairment. Transl. Stroke Res. 2016, 7, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Hort, J.; Valis, M.; Kuca, K.; Angelucci, F. Vascular Cognitive Impairment: Information from Animal Models on the Pathogenic Mechanisms of Cognitive Deficits. Int. J. Mol. Sci. 2019, 20, 2405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McBean, D.E.; Kelly, P.A. Rodent models of global cerebral ischemia: A comparison of two-vessel occlusion and four-vessel occlusion. Gen. Pharmacol. 1998, 30, 431–434. [Google Scholar] [CrossRef]
- Pontarelli, F.; Ofengeim, D.; Zukin, R.S.; Jonas, E.A. Mouse Transient Global Ischemia Two-Vessel Occlusion Model. Bio Protoc. 2012, 2, e262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.Y.; Chen, R.J.; Lee, G.A. Two-vessel Occlusion Mouse Model of Cerebral Ischemia-reperfusion. J. Vis. Exp. 2019, 145, e59078. [Google Scholar] [CrossRef] [PubMed]
- Hattori, Y.; Enmi, J.; Kitamura, A.; Yamamoto, Y.; Saito, S.; Takahashi, Y.; Iguchi, S.; Tsuji, M.; Yamahara, K.; Nagatsuka, K.; et al. A novel mouse model of subcortical infarcts with dementia. J. Neurosci. 2015, 35, 3915–3928. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.K.; Kim, H.; Yang, J.; Kim, J.; Son, J.P.; Jang, H.; Na, D.L. Heterogeneous Disease Progression in a Mouse Model of Vascular Cognitive Impairment. Int. J. Mol. Sci. 2020, 21, 2820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strickland, S. Blood will out: Vascular contributions to Alzheimer’s disease. J. Clin. Investig. 2018, 128, 556–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, R.E.; Nikolic, K.; Ramsay, R.R. One for All? Hitting Multiple Alzheimer’s Disease Targets with One Drug. Front. Neurosci. 2016, 10, 177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benek, O.; Korabecny, J.; Soukup, O. A Perspective on Multi-target Drugs for Alzheimer’s Disease. Trends Pharmacol. Sci. 2020, 41, 434–445. [Google Scholar] [CrossRef]
- Weekman, E.M.; Sudduth, T.L.; Caverly, C.N.; Kopper, T.J.; Phillips, O.W.; Powell, D.K.; Wilcock, D.M. Reduced Efficacy of Anti-Abeta Immunotherapy in a Mouse Model of Amyloid Deposition and Vascular Cognitive Impairment Comorbidity. J. Neurosci. 2016, 36, 9896–9907. [Google Scholar] [CrossRef] [Green Version]
- Sun, M.K. Potential Therapeutics for Vascular Cognitive Impairment and Dementia. Curr. Neuropharmacol. 2018, 16, 1036–1044. [Google Scholar] [CrossRef]
- Kim, J.Y.; Kim, D.H.; Kim, J.H.; Lee, D.; Jeon, H.B.; Kwon, S.J.; Kim, S.M.; Yoo, Y.J.; Lee, E.H.; Choi, S.J.; et al. Soluble intracellular adhesion molecule-1 secreted by human umbilical cord blood-derived mesenchymal stem cell reduces amyloid-beta plaques. Cell Death Differ. 2012, 19, 680–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.E.; Lee, J.; Chang, E.H.; Kim, J.H.; Sung, J.H.; Na, D.L.; Chang, J.W. Activin A secreted by human mesenchymal stem cells induces neuronal development and neurite outgrowth in an in vitro model of Alzheimer’s disease: Neurogenesis induced by MSCs via activin A. Arch. Pharm. Res. 2016, 39, 1171–1179. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.K.; Park, S.E.; Kwon, S.J.; Shim, S.; Byeon, Y.; Kim, J.H.; Na, D.L.; Chang, J.W. Agouti Related Peptide Secreted Via Human Mesenchymal Stem Cells Upregulates Proteasome Activity in an Alzheimer’s Disease Model. Sci. Rep. 2017, 7, 39340. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.K.; Na, D.L.; Chang, J.W. Killing two birds with one stone: The multifunctional roles of mesenchymal stem cells in the treatment of neurodegenerative and muscle diseases. Histol. Histopathol. 2018, 33, 629–638. [Google Scholar] [PubMed]
- Marrazzo, P.; Crupi, A.N.; Alviano, F.; Teodori, L.; Bonsi, L. Exploring the roles of MSCs in infections: Focus on bacterial diseases. J. Mol. Med. 2019, 97, 437–450. [Google Scholar] [CrossRef]
- Kim, H.J.; Seo, S.W.; Chang, J.W.; Lee, J.I.; Kim, C.H.; Chin, J.; Choi, S.J.; Kwon, H.; Yun, H.J.; Lee, J.M.; et al. Stereotactic brain injection of human umbilical cord blood mesenchymal stem cells in patients with Alzheimer’s disease dementia: A phase 1 clinical trial. Alzheimers Dement. 2015, 1, 95–102. [Google Scholar] [CrossRef]
- Park, S.E.; Lee, N.K.; Na, D.L.; Chang, J.W. Optimal mesenchymal stem cell delivery routes to enhance neurogenesis for the treatment of Alzheimer’s disease: Optimal MSCs delivery routes for the treatment of AD. Histol. Histopathol. 2018, 33, 533–541. [Google Scholar]
- Jiang, W.; Gong, L.; Liu, F.; Mu, J. Stem cells and vascular dementia: From basic science to the clinic. Cell Tissue Bank. 2020. [Google Scholar] [CrossRef]
- Schneider, J.; Wilson, R.; Bienias, J.; Evans, D.; Bennett, D. Cerebral infarctions and the likelihood of dementia from Alzheimer disease pathology. Neurology 2004, 62, 1148–1155. [Google Scholar] [CrossRef]
- Esiri, M.M.; Nagy, Z.; Smith, M.Z.; Barnetson, L.; Smith, A.D. Cerebrovascular disease and threshold for dementia in the early stages of Alzheimer’s disease. Lancet 1999, 354, 919–920. [Google Scholar] [CrossRef]
- Schneider, J.A.; Boyle, P.A.; Arvanitakis, Z.; Bienias, J.L.; Bennett, D.A. Subcortical infarcts, Alzheimer’s disease pathology, and memory function in older persons. Ann. Neurol. 2007, 62, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Power, M.C.; Mormino, E.; Soldan, A.; James, B.D.; Yu, L.; Armstrong, N.M.; Bangen, K.J.; Delano-Wood, L.; Lamar, M.; Lim, Y.Y.; et al. Combined neuropathological pathways account for age-related risk of dementia. Ann. Neurol. 2018, 84, 10–22. [Google Scholar] [CrossRef] [PubMed]
- Strozyk, D.; Dickson, D.W.; Lipton, R.B.; Katz, M.; Derby, C.A.; Lee, S.; Wang, C.; Verghese, J. Contribution of vascular pathology to the clinical expression of dementia. Neurobiol. Aging 2010, 31, 1710–1720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalaria, R.N.; Erkinjuntti, T. Small vessel disease and subcortical vascular dementia. J. Clin. Neurol. 2006, 2, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Hattori, Y.; Enmi, J.; Iguchi, S.; Saito, S.; Yamamoto, Y.; Tsuji, M.; Nagatsuka, K.; Kalaria, R.N.; Iida, H.; Ihara, M. Gradual Carotid Artery Stenosis in Mice Closely Replicates Hypoperfusive Vascular Dementia in Humans. J. Am. Heart Assoc. 2016, 5, e002757. [Google Scholar] [CrossRef] [Green Version]
- Russell, D.G.; Cardona, P.J.; Kim, M.J.; Allain, S.; Altare, F. Foamy macrophages and the progression of the human tuberculosis granuloma. Nat. Immunol. 2009, 10, 943–948. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, R.; Nishimura, T.; Ochiai, T.; Nakada, S.; Nagatani, M.; Ogasawara, H. Availability of a microglia and macrophage marker, iba-1, for differential diagnosis of spontaneous malignant reticuloses from astrocytomas in rats. J. Toxicol. Pathol. 2013, 26, 55–60. [Google Scholar] [CrossRef] [Green Version]
- Duncombe, J.; Kitamura, A.; Hase, Y.; Ihara, M.; Kalaria, R.N.; Horsburgh, K. Chronic cerebral hypoperfusion: A key mechanism leading to vascular cognitive impairment and dementia. Closing the translational gap between rodent models and human vascular cognitive impairment and dementia. Clin. Sci. 2017, 131, 2451–2468. [Google Scholar] [CrossRef] [Green Version]
- Sarti, C.; Pantoni, L.; Bartolini, L.; Inzitari, D. Persistent impairment of gait performances and working memory after bilateral common carotid artery occlusion in the adult Wistar rat. Behav. Brain Res. 2002, 136, 13–20. [Google Scholar] [CrossRef]
- Matchynski-Franks, J.J.; Pappas, C.; Rossignol, J.; Reinke, T.; Fink, K.; Crane, A.; Twite, A.; Lowrance, S.A.; Song, C.; Dunbar, G.L. Mesenchymal Stem Cells as Treatment for Behavioral Deficits and Neuropathology in the 5xFAD Mouse Model of Alzheimer’s Disease. Cell Transplant. 2016, 25, 687–703. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.S.; Lee, N.K.; Yoo, D.; Lee, J.; Choi, S.J.; Oh, W.; Chang, J.W.; Na, D.L. Lowering the concentration affects the migration and viability of intracerebroventricular-delivered human mesenchymal stem cells. Biochem. Biophys. Res. Commun. 2017, 493, 751–757. [Google Scholar] [CrossRef] [PubMed]
- Giles, A.J.; Hutchinson, M.N.D.; Sonnemann, H.M.; Jung, J.; Fecci, P.E.; Ratnam, N.M.; Zhang, W.; Song, H.; Bailey, R.; Davis, D.; et al. Dexamethasone-induced immunosuppression: Mechanisms and implications for immunotherapy. J. Immunother. Cancer 2018, 6, 51. [Google Scholar] [CrossRef] [PubMed]
- Pardridge, W.M. CSF, blood-brain barrier, and brain drug delivery. Expert Opin. Drug Deliv. 2016, 13, 963–975. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.W.; Lee, N.K.; Yang, J.H.; Son, H.J.; Bang, S.I.; Chang, J.W.; Na, D.L. A Comparison of Immune Responses Exerted Following Syngeneic, Allogeneic, and Xenogeneic Transplantation of Mesenchymal Stem Cells into the Mouse Brain. Int. J. Mol. Sci. 2020, 21, 3052. [Google Scholar] [CrossRef]
- Chang, S.H.; Park, C.G. Allogeneic ADSCs induce CD8 T cell-mediated cytotoxicity and faster cell death after exposure to xenogeneic serum or proinflammatory cytokines. Exp. Mol. Med. 2019, 51, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Washida, K.; Hattori, Y.; Ihara, M. Animal Models of Chronic Cerebral Hypoperfusion: From Mouse to Primate. Int. J. Mol. Sci. 2019, 20, 6176. [Google Scholar] [CrossRef] [Green Version]
- Coltman, R.; Spain, A.; Tsenkina, Y.; Fowler, J.H.; Smith, J.; Scullion, G.; Allerhand, M.; Scott, F.; Kalaria, R.N.; Ihara, M.; et al. Selective white matter pathology induces a specific impairment in spatial working memory. Neurobiol. Aging 2011, 32, 2324.e7–2324.e12. [Google Scholar] [CrossRef] [Green Version]
- Zarow, C.; Vinters, H.V.; Ellis, W.G.; Weiner, M.W.; Mungas, D.; White, L.; Chui, H.C. Correlates of hippocampal neuron number in Alzheimer’s disease and ischemic vascular dementia. Ann. Neurol. 2005, 57, 896–903. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Gu, J.H.; Dai, C.L.; Liu, Q.; Iqbal, K.; Liu, F.; Gong, C.X. Chronic cerebral hypoperfusion causes decrease of O-GlcNAcylation, hyperphosphorylation of tau and behavioral deficits in mice. Front. Aging Neurosci. 2014, 6, 10. [Google Scholar] [CrossRef]
- Ahn, S.Y.; Chang, Y.S.; Sung, D.K.; Sung, S.I.; Yoo, H.S.; Im, G.H.; Choi, S.J.; Park, W.S. Optimal Route for Mesenchymal Stem Cells Transplantation after Severe Intraventricular Hemorrhage in Newborn Rats. PLoS ONE 2015, 10, e0132919. [Google Scholar] [CrossRef]
- Murakami, Y.; Zhao, Q.; Harada, K.; Tohda, M.; Watanabe, H.; Matsumoto, K. Choto-san, a Kampo formula, improves chronic cerebral hypoperfusion-induced spatial learning deficit via stimulation of muscarinic M1 receptor. Pharmacol. Biochem. Behav. 2005, 81, 616–625. [Google Scholar] [CrossRef] [PubMed]
- Cechetti, F.; Pagnussat, A.S.; Worm, P.V.; Elsner, V.R.; Ben, J.; da Costa, M.S.; Mestriner, R.; Weis, S.N.; Netto, C.A. Chronic brain hypoperfusion causes early glial activation and neuronal death, and subsequent long-term memory impairment. Brain Res. Bull. 2012, 87, 109–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pencea, V.; Bingaman, K.D.; Wiegand, S.J.; Luskin, M.B. Infusion of brain-derived neurotrophic factor into the lateral ventricle of the adult rat leads to new neurons in the parenchyma of the striatum, septum, thalamus, and hypothalamus. J. Neurosci. 2001, 21, 6706–6717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, Q.; Hei, Y.; Luo, Q.; Tian, Y.; Yang, J.; Li, J.; Wei, L.; Liu, W. BMSCs transplantation improves cognitive impairment via up-regulation of hippocampal GABAergic system in a rat model of chronic cerebral hypoperfusion. Neuroscience 2015, 311, 464–473. [Google Scholar] [CrossRef]
- Ma, N.; Cheng, H.; Lu, M.; Liu, Q.; Chen, X.; Yin, G.; Zhu, H.; Zhang, L.; Meng, X.; Tang, Y.; et al. Magnetic resonance imaging with superparamagnetic iron oxide fails to track the long-term fate of mesenchymal stem cells transplanted into heart. Sci. Rep. 2015, 5, 9058. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.E.; Ha, C.W.; Jung, M.; Jin, H.J.; Lee, M.; Song, H.; Choi, S.; Oh, W.; Yang, Y.S. Mesenchymal stem/progenitor cells developed in cultures from UC blood. Cytotherapy 2004, 6, 476–486. [Google Scholar] [CrossRef]
- Lee, N.K.; Kim, H.S.; Yoo, D.; Hwang, J.W.; Choi, S.J.; Oh, W.; Chang, J.W.; Na, D.L. Magnetic Resonance Imaging of Ferumoxytol-Labeled Human Mesenchymal Stem Cells in the Mouse Brain. Stem Cell Rev. 2016. [Google Scholar] [CrossRef] [Green Version]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Kim, C.; Kwak, M.J.; Cho, S.Y.; Ko, A.R.; Rheey, J.; Kwon, J.Y.; Chung, Y.; Jin, D.K. Decreased performance in IDUA knockout mouse mimic limitations of joint function and locomotion in patients with Hurler syndrome. Orphanet J. Rare Dis. 2015, 10, 121. [Google Scholar] [CrossRef] [Green Version]
- Lee, N.K.; Yang, J.; Chang, E.H.; Park, S.E.; Lee, J.; Choi, S.J.; Oh, W.; Chang, J.W.; Na, D.L. Intra-Arterially Delivered Mesenchymal Stem Cells Are Not Detected in the Brain Parenchyma in an Alzheimer’s Disease Mouse Model. PLoS ONE 2016, 11, e0155912. [Google Scholar] [CrossRef] [Green Version]









© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, N.K.; Kim, H.; Chang, J.W.; Jang, H.; Kim, H.; Yang, J.; Kim, J.; Son, J.P.; Na, D.L. Exploring the Potential of Mesenchymal Stem Cell-Based Therapy in Mouse Models of Vascular Cognitive Impairment. Int. J. Mol. Sci. 2020, 21, 5524. https://doi.org/10.3390/ijms21155524
Lee NK, Kim H, Chang JW, Jang H, Kim H, Yang J, Kim J, Son JP, Na DL. Exploring the Potential of Mesenchymal Stem Cell-Based Therapy in Mouse Models of Vascular Cognitive Impairment. International Journal of Molecular Sciences. 2020; 21(15):5524. https://doi.org/10.3390/ijms21155524
Chicago/Turabian StyleLee, Na Kyung, Hyeongseop Kim, Jong Wook Chang, Hyemin Jang, Hunnyun Kim, Jehoon Yang, Jeyun Kim, Jeong Pyo Son, and Duk L. Na. 2020. "Exploring the Potential of Mesenchymal Stem Cell-Based Therapy in Mouse Models of Vascular Cognitive Impairment" International Journal of Molecular Sciences 21, no. 15: 5524. https://doi.org/10.3390/ijms21155524
APA StyleLee, N. K., Kim, H., Chang, J. W., Jang, H., Kim, H., Yang, J., Kim, J., Son, J. P., & Na, D. L. (2020). Exploring the Potential of Mesenchymal Stem Cell-Based Therapy in Mouse Models of Vascular Cognitive Impairment. International Journal of Molecular Sciences, 21(15), 5524. https://doi.org/10.3390/ijms21155524