Physiological and Nutraceutical Quality of Green and Red Pigmented Lettuce in Response to NaCl Concentration in Two Successive Harvests

 ,
,  ,
, 
 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Growth Conditions, Lettuce Cultivars, Experimental Design and Salt Application
2.2. Sample Preparation
2.3. Leaf Area, Fresh Yield and Leaf Biomass Determination
2.4. Physiological Parameters: SPAD Index and Leaf Gas Exchange
2.5. Macro and Micro Mineral Content Analysis
2.6. Bioactive Compounds and Antioxidant Activity Assays Analysis
2.7. Statistics
3. Results
3.1. Biometric Measurements
3.2. Leaf Mineral Profile
3.3. SPAD Index and Leaf Gas Exchange
3.4. Qualitative Characteristics
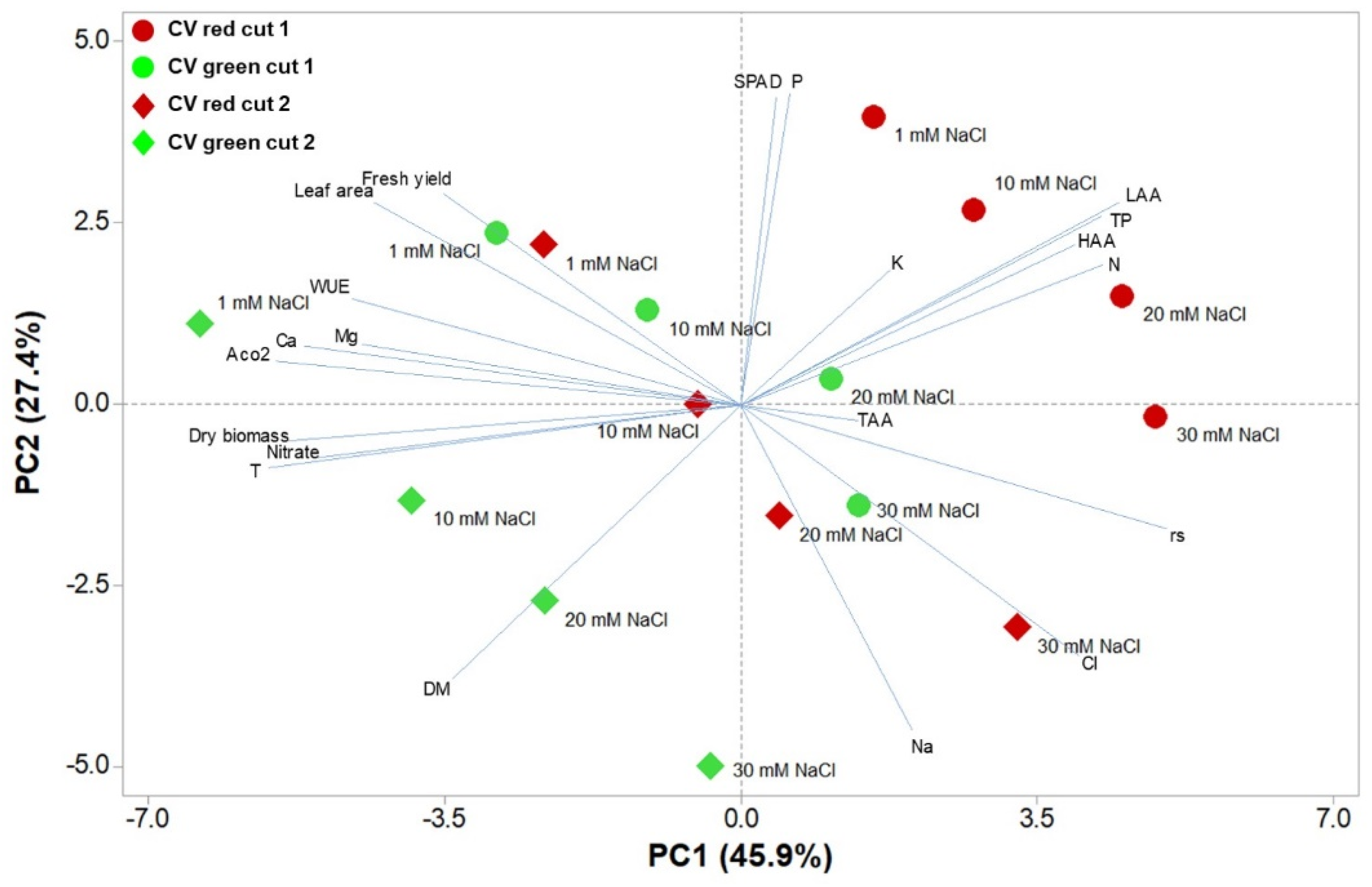
3.5. Principal Component Analysis (PCA)
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sofo, A.; Lundegårdh, B.; Mårtensson, A.; Manfra, M.; Pepe, G.; Sommella, E.; De Nisco, M.; Tenore, G.C.; Campiglia, P.; Scopa, A. Different agronomic and fertilization systems affect polyphenolic profile, antioxidant capacity and mineral composition of lettuce. Sci. Hortic. 2016, 204, 106–115. [Google Scholar] [CrossRef]
- Alvino, A.; Barbieri, G. Vegetables of temperate climates: Leafy vegetables. In The Encyclopedia of Food and Health; Oxford Academic Press: Oxford, UK, 2016; pp. 393–400. [Google Scholar] [CrossRef]
- Pepe, G.; Sommella, E.; Manfra, M.; De Nisco, M.; Tenore, G.C.; Scopa, A.; Sofo, A.; Marzocco, S.; Adesso, S.; Novellino, T.; et al. Evaluation of anti-inflammatory activity and fast UHPLC-DAD-IT-TOF profiling of polyphenolic compounds extracted from green lettuce (Lactuca sativa L.; var. Maravilla de Verano). Food Chem. 2015, 167, 153–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Hidalgo, S.; Artés-Hernández, F.; Gómez, P.A.; Fernández, J.A.; Artés, F. Quality of fresh-cut baby spinach grown under a floating trays system as affected by nitrogen fertilisation and innovative packaging treatments. J. Sci. Food Agric. 2010, 90, 1089–1097. [Google Scholar] [CrossRef] [PubMed]
- Gil, M.I.; Garrido, Y. Chapter 21.1—Leafy vegetables: Baby leaves. In Controlled and Modified Atmospheres for Fresh and Fresh-Cut Produce; Gil, M.I., Beaudry, R., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 527–536. [Google Scholar] [CrossRef]
- Massaglia, S.; Merlino, V.M.; Borra, D.; Bargetto, A.; Sottile, F.; Peano, C. Consumer attitudes and preference exploration towards fresh-cut salads using best-worst scaling and latent class analysis. Foods 2019, 8, 568. [Google Scholar] [CrossRef] [Green Version]
- Llorach, R.; Martínez-Sánchez, A.; Tomás-Barberán, F.; Gil, M.; Ferreres, F. Characterisation of polyphenols and antioxidant properties of five lettuce varieties and escarole. Food Chem. 2008, 108, 1028–1038. [Google Scholar] [CrossRef]
- Mulabagal, V.; Ngouajio, M.; Nair, A.; Zhang, Y.; Gottumukkala, A.; Nair, M. In vitro evaluation of red and green lettuce (Lactuca sativa) for functional food properties. Food Chem. 2010, 118, 300–306. [Google Scholar] [CrossRef]
- Baslam, M.; Morales, F.; Garmendia, I.; Goicoechea, N. Nutritional quality of outer and inner leaves of green and red pigmented lettuces (Lactuca sativa L.) consumed as salads. Sci. Hortic. 2013, 151, 103–111. [Google Scholar] [CrossRef]
- Kim, M.J.; Moon, Y.; Tou, J.C.; Mou, B.; Waterland, N.L. Nutritional value, bioactive compounds and health benefits of lettuce (Lactuca sativa L.). J. Food Compos. Anal. 2016, 49, 19–34. [Google Scholar] [CrossRef]
- Durazzo, A.; Azzini, E.; Lazzé, M.; Raguzzini, A.; Pizzala, R.; Maiani, G.; Palomba, L. Antioxidants in Italian head lettuce (Lactuca sativa var. capitataL.) grown in organic and conventional systems under greenhouse conditions. J. Food Biochem. 2013, 38, 56–61. [Google Scholar] [CrossRef]
- Ribas-Agustí, A.; Gratacós-Cubarsí, M.; Sárraga, C.; García-Regueiro, J.-A.; Castellari, M. Analysis of eleven phenolic compounds including novel p-coumaroyl derivatives in lettuce (Lactuca sativa L.) by ultra-high-performance liquid chromatography with photodiode array and mass spectrometry detection. Phytochem. Anal. 2011, 22, 555–563. [Google Scholar] [CrossRef]
- Shahidi, F.; Ambigaipalan, P. Phenolics and polyphenolics in foods, beverages and spices: Antioxidant activity and health effects—A review. J. Funct. Foods 2015, 18, 820–897. [Google Scholar] [CrossRef]
- Złotek, U.; Świeca, M.; Jakubczyk, A. Effect of abiotic elicitation on main health-promoting compounds, antioxidant activity and commercial quality of butter lettuce (Lactuca sativa L.). Food Chem. 2014, 148, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-J.; Fonseca, J.M.; Choi, J.-H.; Kubota, C.; Kwon, D.Y. Salt in irrigation water affects the nutritional and visual properties of romaine lettuce (Lactuca sativa L.). J. Agric. Food Chem. 2008, 56, 3772–3776. [Google Scholar] [CrossRef] [PubMed]
- Annunziata, M.G.; Attico, A.; Woodrow, P.; Oliva, M.A.; Fuggi, A.; Carillo, P. An improved fluorimetric HPLC method for quantifying tocopherols in Brassica rapa L. subsp. sylvestris after harvest. J. Food Compos. Anal. 2012, 27, 145–150. [Google Scholar] [CrossRef]
- Sarno, V.; Manzo, R. Italian companies’ attitude towards GM crops. Nutr. Food Sci. 2016, 46, 685–694. [Google Scholar] [CrossRef]
- Rouphael, Y.; Kyriacou, M.C.; Carillo, P.; Pizzolongo, F.; Romano, R.; Sifola, M.I. Chemical eustress elicits tailored responses and enhances the functional quality of novel food perilla frutescens. Molecules 2019, 24, 185. [Google Scholar] [CrossRef] [Green Version]
- Rouphael, Y.; Kyriacou, M.C. Enhancing quality of fresh vegetables through salinity eustress and biofortification applications facilitated by soilless cultivation. Front. Plant Sci. 2018, 9, 9. [Google Scholar] [CrossRef]
- Pérez-López, U.; Miranda-Apodaca, J.; Muñoz-Rueda, A.; Mena-Petite, A. Lettuce production and antioxidant capacity are differentially modified by salt stress and light intensity under ambient and elevated CO2. J. Plant Physiol. 2013, 170, 1517–1525. [Google Scholar] [CrossRef]
- Pérez-López, U.; Miranda-Apodaca, J.; Lacuesta, M.; Mena-Petite, A.; Muñoz-Rueda, A. Growth and nutritional quality improvement in two differently pigmented lettuce cultivars grown under elevated CO2 and/or salinity. Sci. Hortic. 2015, 195, 56–66. [Google Scholar] [CrossRef]
- Akula, R.; Ravishankar, G.A. Influence of abiotic stress signals on secondary metabolites in plants. Plant Signal. Behav. 2011, 6, 1720–1731. [Google Scholar] [CrossRef] [PubMed]
- Woodrow, P.; Ciarmiello, L.F.; Annunziata, M.G.; Pacifico, S.; Iannuzzi, F.; Mirto, A.; D‘Amelia, L.; Dell’Aversana, E.; Piccolella, S.; Fuggi, A.; et al. Durum wheat seedling responses to simultaneous high light and salinity involve a fine reconfiguration of amino acids and carbohydrate metabolism. Physiol. Plant. 2017, 159, 290–312. [Google Scholar] [CrossRef] [PubMed]
- Lucini, L.; Borgognone, D.; Rouphael, Y.; Cardarelli, M.; Bernardi, J.; Colla, G. Mild Potassium chloride stress alters the mineral composition, hormone network, and phenolic profile in artichoke leaves. Front. Plant Sci. 2016, 7, 948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kyriacou, M.C.; Rouphael, Y. Towards a new definition of quality for fresh fruits and vegetables. Sci. Hortic. 2018, 234, 463–469. [Google Scholar] [CrossRef]
- Giordano, M.; El-Nakhel, C.; Pannico, A.; Kyriacou, M.C.; Stazi, S.R.; De Pascale, S.; Rouphael, Y. Iron biofortification of red and green pigmented lettuce in closed soilless cultivation impacts crop performance and modulates mineral and bioactive composition. Agronomy 2019, 9, 290. [Google Scholar] [CrossRef] [Green Version]
- Petropoulos, S.A.; Levizou, E.; Ntatsi, G.; Fernandes, Â.; Petrotos, K.; Akoumianakis, K.; Barros, L.; Ferreira, I.C.F.R. Salinity effect on nutritional value, chemical composition and bioactive compounds content of Cichorium spinosum L. Food Chem. 2017, 214, 129–136. [Google Scholar] [CrossRef] [Green Version]
- Neocleous, D.; Koukounaras, A.; Siomos, A.S.; Vasilakakis, M. Assessing the salinity effects on mineral composition and nutritional quality of green and red “Baby” lettuce. J. Food Qual. 2014, 37, 1–8. [Google Scholar] [CrossRef]
- Sarker, U.; Islam, M.T.; Oba, S. Salinity stress accelerates nutrients, dietary fiber, minerals, phytochemicals and antioxidant activity in Amaranthus tricolor leaves. PLoS ONE 2018, 13, e0206388. [Google Scholar] [CrossRef] [Green Version]
- Sarker, U.; Oba, S. Salinity stress enhances color parameters, bioactive leaf pigments, vitamins, polyphenols, flavonoids and antioxidant activity in selected Amaranthus leafy vegetables. J. Sci. Food Agric. 2019, 99, 2275–2284. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Augmentation of leaf color parameters, pigments, vitamins, phenolic acids, flavonoids and antioxidant activity in selected Amaranthus tricolor under salinity stress. Sci. Rep. 2018, 8, 12349. [Google Scholar] [CrossRef] [Green Version]
- Colla, G.; Rouphael, Y.; Cardarelli, M.; Svecova, E.; Rea, E.; Lucini, L. Effects of saline stress on mineral composition, phenolic acids and flavonoids in leaves of artichoke and cardoon genotypes grown in floating system. J. Sci. Food Agric. 2013, 93, 1119–1127. [Google Scholar] [CrossRef]
- Rouphael, Y.; Cardarelli, M.; Lucini, L.; Rea, E.; Colla, G. Nutrient solution concentration affects growth, mineral composition, phenolic acids, and flavonoids in leaves of artichoke and cardoon. HortScience 2012, 47, 1424–1429. [Google Scholar] [CrossRef] [Green Version]
- Tomasi, N.; Pinton, R.; Costa, L.D.; Cortella, G.; Terzano, R.; Mimmo, T.; Scampicchio, M.; Cesco, S. New ‘solutions’ for floating cultivation system of ready-to-eat salad: A review. Trends Food Sci. Technol. 2015, 46, 267–276. [Google Scholar] [CrossRef]
- Borgognone, D.; Rouphael, Y.; Cardarelli, M.; Lucini, L.; Colla, G. Changes in biomass, mineral composition, and quality of cardoon in response to NO3−: Cl− ratio and nitrate deprivation from the nutrient solution. Front. Plant Sci. 2016, 7, 978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbieri, G.; Bottino, A.; Di Stasio, E.; Vallone, S.; Maggio, A. Proline and light as quality enhancers of rocket (Eruca sativa Miller) grown under saline conditions. Sci. Hortic. 2011, 128, 393–400. [Google Scholar] [CrossRef]
- Nicoletto, C.; Santagata, S.; Bona, S.; Sambo, P. Influence of cut number on qualitative traits in different cultivars of sweet basil. Ind. Crops Prod. 2013, 44, 465–472. [Google Scholar] [CrossRef]
- Sarker, U.; Hossain, M.M.; Oba, S. Nutritional and antioxidant components and antioxidant capacity in green morph Amaranthus leafy vegetable. Sci. Rep. 2020, 10, 1336. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G.; Graziani, G.; Ritieni, A.; Cardarelli, M.; De Pascale, S. Phenolic composition, antioxidant activity and mineral profile in two seed-propagated artichoke cultivars as affected by microbial inoculants and planting time. Food Chem. 2017, 234, 10–19. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S.; Daramy, M.A. Nutrients, minerals, antioxidant pigments and phytochemicals, and antioxidant capacity of the leaves of stem amaranth. Sci. Rep. 2020, 10, 3892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarker, U.; Oba, S. Nutrients, minerals, pigments, phytochemicals, and radical scavenging activity in Amaranthus blitum leafy vegetables. Sci. Rep. 2020, 10, 3868. [Google Scholar] [CrossRef] [Green Version]
- Fogliano, V.; Verde, V.; Randazzo, G.; Ritieni, A. Method for measuring antioxidant activity and its application to monitoring the antioxidant capacity of wines. J. Agric. Food Chem. 1999, 47, 1035–1040. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Antioxidant constituents of three selected red and green color Amaranthus leafy vegetable. Sci. Rep. 2019, 9, 18233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciarmiello, L.F.; Piccirillo, P.; Carillo, P.; De Luca, A.; Woodrow, P. Determination of the genetic relatedness of fig (Ficus carica L.) accessions using RAPD fingerprint and their agro-morphological characterization. S. Afr. J. Bot. 2015, 97, 40–47. [Google Scholar] [CrossRef]
- El-Nakhel, C.; Giordano, M.; Pannico, A.; Carillo, P.; Fusco, G.M.; De Pascale, S.; Rouphael, Y. Cultivar-specific performance and qualitative descriptors for butterhead salanova lettuce produced in closed soilless cultivation as a candidate salad crop for human life support in space. Life 2019, 9, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carillo, P.; Cirillo, C.; De Micco, V.; Arena, C.; De Pascale, S.; Rouphael, Y. Morpho-anatomical, physiological and biochemical adaptive responses to saline water of Bougainvillea spectabilis Willd. trained to different canopy shapes. Agric. Water Manag. 2019, 212, 12–22. [Google Scholar] [CrossRef]
- Annunziata, M.G.; Ciarmiello, L.F.; Woodrow, P.; Maximova, E.; Fuggi, A.; Carillo, P. Durum wheat roots adapt to salinity remodeling the cellular content of nitrogen metabolites and sucrose. Front. Plant Sci. 2017, 7, 515. [Google Scholar] [CrossRef] [Green Version]
- Shabala, S.; Cuin, T.A. Potassium transport and plant salt tolerance. Physiol. Plant. 2008, 133, 651–669. [Google Scholar] [CrossRef]
- Rouphael, Y.; Raimondi, G.; Lucini, L.; Carillo, P.; Kyriacou, M.C.; Colla, G.; Cirillo, V.; Pannico, A.; El-Nakhel, C.; De Pascale, S. Physiological and metabolic responses triggered by omeprazole improve tomato plant tolerance to NaCl stress. Front. Plant Sci. 2018, 9, 249. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Zheng, Q.; Shen, Q.; Guo, S. The critical role of potassium in plant stress response. Int. J. Mol. Sci. 2013, 14, 7370–7390. [Google Scholar] [CrossRef] [Green Version]
- Cuin, T.A.; Tian, Y.; Betts, S.A.; Chalmandrier, R.; Shabala, S. Ionic relations and osmotic adjustment in durum and bread wheat under saline conditions. Funct. Plant Biol. 2009, 36, 1110–1119. [Google Scholar] [CrossRef]
- Grieve, C.M.; Grattan, S. Mineral nutrient acquisition and response by plants grown in saline environments. In Handbook of Plant and Crop Stress; Marcel Dekker, Inc.: New York, NY, USA, 1999; pp. 203–229. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Catalase, superoxide dismutase and ascorbate-glutathione cycle enzymes confer drought tolerance of Amaranthus tricolor. Sci. Rep. 2018, 8, 16496. [Google Scholar] [CrossRef] [Green Version]
- Sarker, U.; Oba, S. Drought stress effects on growth, ROS markers, compatible solutes, phenolics, flavonoids, and antioxidant activity in amaranthus tricolor. Appl. Biochem. Biotechnol. 2018, 186, 999–1016. [Google Scholar] [CrossRef] [PubMed]
- Sarker, U.; Oba, S. Drought stress enhances nutritional and bioactive compounds, phenolic acids and antioxidant capacity of Amaranthus leafy vegetable. BMC Plant Biol. 2018, 18, 258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarker, U.; Oba, S. Response of nutrients, minerals, antioxidant leaf pigments, vitamins, polyphenol, flavonoid and antioxidant activity in selected vegetable amaranth under four soil water content. Food Chem. 2018, 252, 72–83. [Google Scholar] [CrossRef] [PubMed]
- Ridolfi, M.; Roupsard, O.; Garrec, J.; Dreyer, E. Effects of a calcium deficiency on stomatal conductance and photosynthetic activity of Quercus robur seedlings grown on nutrient solution. Ann. For. Sci. 1996, 53, 325–335. [Google Scholar] [CrossRef]
- Tränkner, M.; Tavakol, E.; Jákli, B. Functioning of potassium and magnesium in photosynthesis, photosynthate translocation and photoprotection. Physiol. Plant. 2018, 163, 414–431. [Google Scholar] [CrossRef] [Green Version]
- Cakmak, I.; Kirkby, E.A. Role of magnesium in carbon partitioning and alleviating photooxidative damage. Physiol. Plant 2008, 133, 692–704. [Google Scholar] [CrossRef]
- Rouphael, Y.; Kyriacou, M.C.; Petropoulos, S.A.; De Pascale, S.; Colla, G. Improving vegetable quality in controlled environments. Sci. Hortic. 2018, 234, 275–289. [Google Scholar] [CrossRef]
- Ferchichi, S.; Hessini, K.; Dell’Aversana, E.; D’Amelia, L.; Woodrow, P.; Ciarmiello, L.F.; Fuggi, A.; Carillo, P. Hordeum vulgare and Hordeum maritimum respond to extended salinity stress displaying different temporal accumulation pattern of metabolites. Funct. Plant Biol. 2018, 45, 1096–1109. [Google Scholar] [CrossRef]
- Song, Y.; Nakajima, T.; Xu, D.; Homma, K.; Kokubun, M. Genotypic variation in salinity tolerance and its association with nodulation and nitrogen uptake in soybean. Plant Prod. Sci. 2017, 20, 490–498. [Google Scholar] [CrossRef]
- Gharibzahedi, S.M.T.; Jafari, S.M. The importance of minerals in human nutrition: Bioavailability, food fortification, processing effects and nanoencapsulation. Trends in Food Sci. Technol. 2017, 62, 119–132. [Google Scholar] [CrossRef]
- Corrado, G.; Chiaiese, P.; Lucini, L.; Miras-Moreno, B.; Colla, G.; Rouphael, Y. Successive harvests affect yield, quality and metabolic profile of sweet basil (Ocimum basilicum L.). Agronomy 2020, 10, 830. [Google Scholar] [CrossRef]
- Van Oosten, M.J.; Dell’Aversana, E.; Ruggiero, A.; Cirillo, V.; Gibon, Y.; Woodrow, P.; Maggio, A.; Carillo, P. Omeprazole treatment enhances nitrogen use efficiency through increased nitrogen uptake and assimilation in corn. Front. Plant Sci. 2019, 10, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carillo, P.; Mastrolonardo, G.; Nacca, F.; Fuggi, A. Nitrate reductase in durum wheat seedlings as affected by nitrate nutrition and salinity. Funct. Plant Biol. 2005, 32, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Lillo, C.; Smith, L.H.; Nimmo, H.G.; Wilkins, M.B. Rhythms in magnesium ion inhibition and hysteretic properties of nitrate reductase in the CAM plant Bryophyllum fedtschenkoi. Physiol. Plant. 1996, 98, 140–146. [Google Scholar] [CrossRef]
- Carillo, P.; Woodrow, P.; Raimondi, G.; El-Nakhel, C.; Pannico, A.; Kyriacou, M.C.; Colla, G.; Mori, M.; Giordano, M.; De Pascale, S.; et al. Omeprazole promotes chloride exclusion and induces salt tolerance in greenhouse basil. Agronomy 2019, 9, 355. [Google Scholar] [CrossRef] [Green Version]
- Touraine, B.; Clarkson, D.T.; Muller, B. Regulation of nitrate uptake at the whole plant level. In A Whole Plant Perspective on Carbon-Nitrogen Interactions; SPB Academic Publishing: The Hague, The Netherlands, 1994; pp. 11–30. [Google Scholar]


{kind=link}
| Source of Variance | Leaf Area | Fresh Yield | Dry Biomass | Leaf Dry Matter |
|---|---|---|---|---|
| (cm2 plant−1) | (kg m−2) | (g m−2) | (%) | |
| Cultivar (CV) | ||||
| Green | 92.10 ± 2.55 | 5.59 ± 0.14 a | 271.7 ± 6.61 a | 4.89 ± 0.10 a |
| Red | 88.37 ± 2.02 | 4.98 ± 0.13 b | 214.3 ± 4.96 b | 4.33 ± 0.07 b |
| Salinity (S) | ||||
| 1 mM NaCl | 99.58 ± 3.16 a | 5.82 ± 0.22 a | 261.7 ± 14.6 a | 4.48 ± 0.13 b |
| 10 mM NaCl | 94.92 ± 2.67 a | 5.39 ± 0.16 b | 247.2 ± 10.2 ab | 4.58 ± 0.12 b |
| 20 mM NaCl | 88.25 ± 1.69 b | 5.20 ± 0.19 b | 235.1 ± 10.3 b | 4.53 ± 0.16 b |
| 30 mM NaCl | 78.18 ± 1.40 c | 4.73 ± 0.15 c | 228.1 ± 10.1 b | 4.83 ± 0.16 a |
| Cut (C) | ||||
| Cut 1 | 89.20 ± 1.80 | 5.49 ± 0.14 a | 234.9 ± 7.15 b | 4.27 ± 0.06 b |
| Cut 2 | 91.27 ± 2.74 | 5.08 ± 0.15 b | 251.1 ± 9.12 a | 4.94 ± 0.10 a |
| CV × S | ||||
| Green × 1 mM NaCl | 101.65 ± 5.37 | 6.15 ± 0.34 | 297.0 ± 17.6 | 4.83 ± 0.13 |
| Green × 10 mM NaCl | 98.66 ± 4.55 | 5.72 ± 0.17 | 272.2 ± 9.05 | 4.77 ± 0.15 |
| Green × 20 mM NaCl | 89.82 ± 1.71 | 5.47 ± 0.30 | 261.0 ± 12.5 | 4.81 ± 0.25 |
| Green × 30 mM NaCl | 78.28 ± 0.91 | 5.02 ± 0.12 | 256.8 ± 7.61 | 5.14 ± 0.26 |
| Red × 1 mM NaCl | 97.51 ± 3.68 | 5.48 ± 0.21 | 226.4 ± 11.3 | 4.13 ± 0.10 |
| Red × 10 mM NaCl | 91.19 ± 2.25 | 5.06 ± 0.21 | 222.2 ± 11.3 | 4.40 ± 0.18 |
| Red × 20 mM NaCl | 86.68 ± 2.94 | 4.93 ± 0.20 | 209.2 ± 6.66 | 4.26 ± 0.15 |
| Red × 30 mM NaCl | 78.08 ± 2.79 | 4.43 ± 0.22 | 199.4 ± 7.92 | 4.51 ± 0.11 |
| S × C | ||||
| 1 mM NaCl × Cut 1 | 94.94 ± 2.95 | 5.86 ± 0.34 | 250.8 ± 19.7 | 4.26 ± 0.15 |
| 10 mM NaCl × Cut 1 | 92.51 ± 4.24 | 5.57 ± 0.19 | 236.5 ± 12.1 | 4.24 ± 0.10 |
| 20 mM NaCl × Cut 1 | 88.37 ± 3.00 | 5.49 ± 0.33 | 228.9 ± 15.4 | 4.16 ± 0.09 |
| 30 mM NaCl × Cut 1 | 80.98 ± 1.35 | 5.03 ± 0.13 | 223.4 ± 9.31 | 4.43 ± 0.10 |
| 1 mM NaCl × Cut 2 | 104.22 ± 5.17 | 5.77 ± 0.30 | 272.5 ± 22.4 | 4.70 ± 0.18 |
| 10 mM NaCl × Cut 2 | 97.34 ± 3.32 | 5.22 ± 0.26 | 257.8 ± 16.4 | 4.93 ± 0.10 |
| 20 mM NaCl × Cut 2 | 88.14 ± 1.87 | 4.91 ± 0.10 | 241.3 ± 14.7 | 4.90 ± 0.22 |
| 30 mM NaCl × Cut 2 | 75.38 ± 1.91 | 4.43 ± 0.21 | 232.8 ± 18.9 | 5.22 ± 0.22 |
| CV × C | ||||
| Green × Cut 1 | 89.09 ± 3.00 | 5.81 ± 0.20 | 260.6 ± 8.31 | 4.49 ± 0.05 b |
| Green × Cut 2 | 95.11 ± 4.06 | 5.37 ± 0.19 | 282.8 ± 9.54 | 5.28 ± 0.11 a |
| Red × Cut 1 | 89.31 ± 2.15 | 5.17 ± 0.14 | 209.2 ± 4.98 | 4.06 ± 0.05 c |
| Red × Cut 2 | 87.43 ± 3.50 | 4.78 ± 0.20 | 219.4 ± 8.58 | 4.60 ± 0.07 b |
| Significance | ||||
| Cultivar (CV) | ns | *** | *** | *** |
| Salinity (S) | *** | *** | ** | *** |
| Cut (C) | ns | ** | * | *** |
| CV × S | ns | ns | ns | ns |
| S × C | ns | ns | ns | ns |
| CV × C | ns | ns | ns | * |
| CV × S × C | ns | ns | ns | ns |
| Source of Variance | N | P | K | Ca | Mg | Na | Cl |
|---|---|---|---|---|---|---|---|
| (g kg−1 dw) | (g kg−1 dw) | (g kg−1 dw) | (g kg−1 dw) | (g kg−1 dw) | (g kg−1 dw) | (g kg−1 dw) | |
| Cultivar (CV) | |||||||
| Green | 40.51 ± 0.32 b | 13.80 ± 0.37 b | 38.11 ± 2.13 b | 7.68 ± 0.61 a | 3.02 ± 0.21 | 14.89 ± 1.78 | 22.82 ± 2.05 b |
| Red | 41.54 ± 0.52 a | 14.63 ± 0.24 a | 59.61 ± 0.94 a | 6.75 ± 0.37 b | 3.02 ± 0.17 | 13.23 ± 1.67 | 26.54 ± 2.05 a |
| Salinity (S) | |||||||
| 1 mM NaCl | 40.72 ± 0.52 | 15.27 ± 0.34 a | 53.02 ± 2.78 a | 9.56 ± 0.63 a | 3.87 ± 0.32 a | 2.98 ± 0.29 d | 10.00 ± 0.88 d |
| 10 mM NaCl | 40.93 ± 0.52 | 14.56 ± 0.34 ab | 52.31 ± 2.48 a | 8.10 ± 0.52 b | 3.18 ± 0.19 b | 11.91 ± 1.05 c | 23.43 ± 0.88 c |
| 20 mM NaCl | 40.94 ± 0.92 | 14.14 ± 0.43 b | 45.91 ± 4.69 b | 6.02 ± 0.55 c | 2.60 ± 0.21 c | 18.31 ± 1.34 b | 30.01 ± 0.94 b |
| 30 mM NaCl | 41.53 ± 0.49 | 12.90 ± 0.46 c | 44.21 ± 4.83 b | 5.18 ± 0.39 d | 2.42 ± 0.10 c | 23.03 ± 1.24 a | 35.28 ± 1.20 a |
| Cut (C) | |||||||
| Cut 1 | 42.16 ± 0.31 a | 15.00 ± 0.28 a | 47.83 ± 3.60 b | 5.89 ± 0.43 b | 2.49 ± 0.13 b | 11.47 ± 1.25 b | 24.09 ± 1.71 b |
| Cut 2 | 39.90 ± 0.43 b | 13.43 ± 0.29 b | 49.90 ± 1.56 a | 8.54 ± 0.43 a | 3.54 ± 0.18 a | 16.64 ± 1.96 a | 25.27 ± 2.40 a |
| CV × S | |||||||
| Green × 1 mM NaCl | 39.55 ± 0.66 | 15.19 ± 0.58 | 44.48 ± 2.05 b | 10.74 ± 0.73 a | 4.12 ± 0.39 a | 3.15 ± 0.52 e | 8.45 ± 0.70 |
| Green × 10 mM NaCl | 40.72 ± 0.54 | 13.99 ± 0.51 | 45.26 ± 1.69 b | 9.25 ± 0.48 ab | 3.36 ± 0.24 abc | 13.55 ± 1.17 cd | 21.33 ± 0.90 |
| Green × 20 mM NaCl | 41.01 ± 0.67 | 14.01 ± 0.75 | 32.32 ± 4.43 c | 5.77 ± 1.00 d | 2.29 ± 0.37 d | 18.82 ± 2.18 b | 27.99 ± 1.42 |
| Green × 30 mM NaCl | 40.76 ± 0.68 | 12.02 ± 0.61 | 30.38 ± 4.44 c | 4.96 ± 0.69 d | 2.29 ± 0.14 d | 24.03 ± 1.99 a | 33.51 ± 2.03 |
| Red × 1 mM NaCl | 41.88 ± 0.46 | 15.34 ± 0.40 | 61.57 ± 0.85 a | 8.38 ± 0.82 bc | 3.62 ± 0.52 ab | 2.80 ± 0.29 e | 11.54 ± 1.39 |
| Red × 10 mM NaCl | 41.13 ± 0.93 | 15.13 ± 0.36 | 59.36 ± 2.08 a | 6.95 ± 0.66 cd | 3.00 ± 0.30 bcd | 10.27 ± 1.55 d | 25.53 ± 0.92 |
| Red × 20 mM NaCl | 40.86 ± 1.80 | 14.26 ± 0.47 | 59.49 ± 1.78 a | 6.26 ± 0.55 d | 2.90 ± 0.15 bcd | 17.80 ± 1.76 bc | 32.04 ± 0.52 |
| Red × 30 mM NaCl | 42.29 ± 0.60 | 13.77 ± 0.51 | 58.03 ± 2.57 a | 5.40 ± 0.43 d | 2.55 ± 0.13 cd | 22.04 ± 1.55 ab | 37.05 ± 0.96 |
| S × C | |||||||
| 1 mM NaCl × Cut 1 | 41.36 ± 0.64 | 16.15 ± 0.28 | 52.91 ± 4.70 | 7.95 ± 0.70 bc | 2.92 ± 0.26 c | 3.56 ± 0.38 f | 12.12 ± 1.13 e |
| 10 mM NaCl × Cut 1 | 41.91 ± 0.47 | 15.26 ± 0.22 | 53.78 ± 4.36 | 6.96 ± 0.69 cd | 2.62 ± 0.15 cd | 9.06 ± 1.08 e | 23.68 ± 1.66 d |
| 20 mM NaCl × Cut 1 | 42.99 ± 0.75 | 14.91 ± 0.53 | 42.82 ± 9.09 | 4.32 ± 0.35 e | 2.14 ± 0.32 d | 14.07 ± 0.67 d | 28.33 ± 1.51 c |
| 30 mM NaCl × Cut 1 | 42.38 ± 0.53 | 13.66 ± 0.58 | 41.80 ± 9.47 | 4.32 ± 0.57 e | 2.28 ± 0.17 d | 19.19 ± 0.38 c | 32.22 ± 1.51 b |
| 1 mM NaCl × Cut 2 | 40.07 ± 0.78 | 14.38 ± 0.33 | 53.14 ± 3.47 | 11.16 ± 0.50 a | 4.82 ± 0.12 a | 2.39 ± 0.29 f | 7.87 ± 0.53 f |
| 10 mM NaCl × Cut 2 | 39.94 ± 0.75 | 13.87 ± 0.52 | 50.84 ± 2.68 | 9.24 ± 0.46 b | 3.74 ± 0.09 b | 14.75 ± 0.66 d | 23.18 ± 0.81 d |
| 20 mM NaCl × Cut 2 | 38.89 ± 1.20 | 13.36 ± 0.52 | 48.99 ± 3.18 | 7.71 ± 0.23 c | 3.05 ± 0.10 c | 22.55 ± 0.56 b | 31.69 ± 0.70 b |
| 30 mM NaCl × Cut 2 | 40.68 ± 0.70 | 12.13 ± 0.60 | 46.62 ± 3.27 | 6.05 ± 0.23 d | 2.56 ± 0.08 cd | 26.88 ± 0.85 a | 38.34 ± 0.53 a |
| CV × C | |||||||
| Green × Cut 1 | 41.43 ± 0.34 | 15.01 ± 0.40 a | 32.84 ± 3.53 d | 6.19 ± 0.82 | 2.48 ± 0.23 | 12.27 ± 1.70 | 21.20 ± 2.24 |
| Green × Cut 2 | 39.60 ± 0.40 | 12.59 ± 0.40 b | 43.39 ± 1.19 c | 9.17 ± 0.69 | 3.55 ± 0.29 | 17.51 ± 3.01 | 24.43 ± 3.48 |
| Red × Cut 1 | 42.89 ± 0.43 | 14.98 ± 0.40 a | 62.81 ± 0.88 a | 5.58 ± 0.32 | 2.50 ± 0.11 | 10.67 ± 1.88 | 26.98 ± 2.40 |
| Red × Cut 2 | 40.20 ± 0.78 | 14.28 ± 0.26 a | 56.41 ± 1.01 b | 7.92 ± 0.48 | 3.54 ± 0.23 | 15.78 ± 2.63 | 26.10 ± 3.43 |
| Significance | |||||||
| Cultivar (CV) | * | ** | *** | *** | ns | ns | *** |
| Salinity (S) | ns | *** | *** | *** | *** | *** | *** |
| Cut (C) | *** | *** | * | *** | *** | *** | * |
| CV × S | ns | ns | *** | *** | *** | * | ns |
| S × C | ns | ns | ns | ** | *** | *** | *** |
| CV × C | ns | *** | *** | ns | ns | ns | ns |
| CV × S × C | ns | ns | *** | ** | ns | ns | ns |
| Source of Variance | SPAD Index | ACO2 | rs | T | WUEi |
|---|---|---|---|---|---|
| (μmol CO2 m−2 s−1) | (m2 s−1 mol−1) | (mol H2O m−2 s−1) | (μmol CO2 mol−1 H2O) | ||
| Cultivar (CV) | |||||
| Green | 16.53 ± 0.30 b | 7.82 ± 0.25 a | 3.96 ± 0.21 b | 6.41 ± 0.16 a | 1.24 ± 0.04 a |
| Red | 18.66 ± 0.43 a | 5.20 ± 0.21 b | 4.83 ± 0.26 a | 5.43 ± 0.12 b | 0.95 ± 0.03 b |
| Salinity (S) | |||||
| 1 mM NaCl | 19.34 ± 0.51 a | 7.97 ± 0.38 a | 3.32 ± 0.22 c | 6.33 ± 0.27 a | 1.29 ± 0.07 a |
| 10 mM NaCl | 18.14 ± 0.55 b | 6.88 ± 0.37 b | 3.96 ± 0.25 bc | 6.03 ± 0.20 a | 1.14 ± 0.05 b |
| 20 mM NaCl | 17.31 ± 0.43 b | 6.04 ± 0.38 c | 4.61 ± 0.26 b | 6.01 ± 0.20 a | 0.99 ± 0.04 c |
| 30 mM NaCl | 15.60 ± 0.39 c | 5.15 ± 0.33 d | 5.83 ± 0.35 a | 5.30 ± 0.17 b | 0.97 ± 0.05 c |
| Cut (C) | |||||
| Cut 1 | 18.14 ± 0.44 a | 6.19 ± 0.31 b | 4.56 ± 0.21 a | 5.47 ± 0.12 b | 1.11 ± 0.05 |
| Cut 2 | 17.06 ± 0.40 b | 6.83 ± 0.27 a | 4.20 ± 0.26 b | 6.37 ± 0.17 a | 1.08 ± 0.04 |
| CV × S | |||||
| Green × 1 mM NaCl | 18.05 ± 0.41 | 9.53 ± 0.24 | 3.12 ± 0.38 d | 6.78 ± 0.46 | 1.47 ± 0.09 |
| Green × 10 mM NaCl | 16.83 ± 0.47 | 8.00 ± 0.49 | 3.65 ± 0.44 cd | 6.46 ± 0.28 | 1.26 ± 0.08 |
| Green × 20 mM NaCl | 16.19 ± 0.27 | 7.31 ± 0.40 | 4.13 ± 0.31 bcd | 6.43 ± 0.26 | 1.14 ± 0.04 |
| Green × 30 mM NaCl | 15.07 ± 0.53 | 6.46 ± 0.35 | 4.92 ± 0.33 bc | 5.97 ± 0.18 | 1.10 ± 0.08 |
| Red × 1 mM NaCl | 20.64 ± 0.54 | 6.41 ± 0.33 | 3.48 ± 0.26 d | 5.87 ± 0.24 | 1.10 ± 0.06 |
| Red × 10 mM NaCl | 19.46 ± 0.64 | 5.76 ± 0.33 | 4.27 ± 0.15 bcd | 5.60 ± 0.22 | 1.03 ± 0.05 |
| Red × 20 mM NaCl | 18.42 ± 0.50 | 4.76 ± 0.37 | 5.38 ± 0.34 b | 5.60 ± 0.26 | 0.83 ± 0.05 |
| Red × 30 mM NaCl | 16.12 ± 0.52 | 3.85 ± 0.18 | 7.25 ± 1.30 a | 4.64 ± 0.09 | 0.83 ± 0.04 |
| S × C | |||||
| 1 mM NaCl × Cut 1 | 19.44 ± 0.94 | 7.75 ± 0.57 a | 3.51 ± 0.18 | 5.95 ± 0.28 | 1.30 ± 0.07 |
| 10 mM NaCl × Cut 1 | 19.00 ± 0.80 | 6.56 ± 0.59 bc | 4.08 ± 0.16 | 5.71 ± 0.25 | 1.16 ± 0.10 |
| 20 mM NaCl × Cut 1 | 18.08 ± 0.63 | 5.07 ± 0.50 d | 5.38 ± 0.38 | 5.23 ± 0.16 | 0.95 ± 0.08 |
| 30 mM NaCl × Cut 1 | 16.03 ± 0.47 | 5.38 ± 0.57 cd | 6.04 ± 0.47 | 4.99 ± 0.18 | 1.06 ± 0.09 |
| 1 mM NaCl × Cut 2 | 19.25 ± 0.50 | 8.19 ± 0.52 a | 3.09 ± 0.45 | 6.71 ± 0.46 | 1.27 ± 0.12 |
| 10 mM NaCl × Cut 2 | 17.29 ± 0.63 | 7.20 ± 0.46 ab | 3.84 ± 0.58 | 6.35 ± 0.28 | 1.13 ± 0.05 |
| 20 mM NaCl × Cut 2 | 16.54 ± 0.44 | 7.00 ± 0.41 ab | 4.14 ± 0.58 | 6.79 ± 0.18 | 1.03 ± 0.04 |
| 30 mM NaCl × Cut 2 | 15.16 ± 0.60 | 4.93 ± 0.36 d | 5.93 ± 0.55 | 5.62 ± 0.27 | 0.88 ± 0.05 |
| CV × C | |||||
| Green × Cut 1 | 16.90 ± 0.26 | 7.71 ± 0.34 | 4.22 ± 0.20 | 5.97 ± 0.17 | 1.30 ± 0.06 |
| Green × Cut 2 | 16.17 ± 0.54 | 7.94 ± 0.36 | 3.69 ± 0.37 | 6.85 ± 0.24 | 1.18 ± 0.06 |
| Red × Cut 1 | 19.37 ± 0.67 | 4.67 ± 0.28 | 5.39 ± 0.40 | 4.97 ± 0.09 | 0.93 ± 0.04 |
| Red × Cut 2 | 17.95 ± 0.48 | 5.73 ± 0.27 | 4.81 ± 0.35 | 5.89 ± 0.19 | 0.97 ± 0.04 |
| Significance | |||||
| Cultivar (CV) | *** | *** | *** | *** | *** |
| Salinity (S) | *** | *** | *** | *** | *** |
| Cut (C) | *** | ** | ** | *** | ns |
| CV × S | ns | ns | * | ns | ns |
| S × C | ns | ** | ns | ns | ns |
| CV × C | ns | ns | ns | ns | ns |
| CV × S × C | ns | ns | ns | ns | ns |
| Source of Variance | Nitrate | LAA | HAA | TP | TAA |
|---|---|---|---|---|---|
| (mg kg−1 fw) | (mmol Trolox 100g−1 dw) | (mmol Ascorbic ac. eq. kg−1 dw) | (mg eq. Gallic acid g−1 dw) | (mg 100g−1 fw) | |
| Cultivar (CV) | |||||
| Green | 2019 ± 93.2 | 5.05 ± 0.22 b | 1.44 ± 0.04 | 51.10 ± 1.99 b | 21.72 ± 1.24 b |
| Red | 1881 ± 67.4 | 6.73 ± 0.30 a | 1.49 ± 0.03 | 66.50 ± 2.89 a | 30.61 ± 2.54 a |
| Salinity (S) | |||||
| 1 mM NaCl | 2125 ± 142 a | 5.81 ± 0.48 | 1.43 ± 0.05 | 58.88 ± 4.19 | 20.39 ± 1.32 c |
| 10 mM NaCl | 2048 ± 120 a | 6.07 ± 0.48 | 1.44 ± 0.04 | 58.45 ± 5.59 | 27.28 ± 2.59 b |
| 20 mM NaCl | 1925 ± 91.4 a | 5.82 ± 0.38 | 1.51 ± 0.05 | 59.88 ± 3.88 | 35.54 ± 3.91 a |
| 30 mM NaCl | 1703 ± 69.3 b | 5.85 ± 0.47 | 1.47 ± 0.06 | 57.98 ± 2.99 | 21.44 ± 1.89 c |
| Cut (C) | |||||
| Cut 1 | 1679 ± 39.5 b | 6.81 ± 0.25 a | 1.58 ± 0.02 a | 67.10 ± 2.80 a | 24.43 ± 1.81 b |
| Cut 2 | 2221 ± 75.3 a | 4.97 ± 0.25 b | 1.34 ± 0.03 b | 50.50 ± 1.90 b | 27.89 ± 2.48 a |
| CV × S | |||||
| Green × 1 mM NaCl | 2265 ± 253 | 4.71 ± 0.51 | 1.41 ± 0.08 | 54.11 ± 2.81 bcd | 21.10 ± 2.02 c |
| Green × 10 mM NaCl | 2154 ± 185 | 5.08 ± 0.48 | 1.45 ± 0.05 | 46.26 ± 5.09 d | 22.32 ± 3.00 c |
| Green × 20 mM NaCl | 1957 ± 115 | 5.25 ± 0.52 | 1.47 ± 0.08 | 51.07 ± 4.18 cd | 23.72 ± 1.82 c |
| Green × 30 mM NaCl | 1700 ± 108 | 5.16 ± 0.34 | 1.43 ± 0.09 | 52.97 ± 3.75 bcd | 19.74 ± 3.19 c |
| Red × 1 mM NaCl | 1985 ± 132 | 6.91 ± 0.54 | 1.45 ± 0.06 | 63.66 ± 7.76 ab | 19.67 ± 1.84 c |
| Red × 10 mM NaCl | 1943 ± 156 | 7.05 ± 0.6399 | 1.44 ± 0.07 | 70.65 ± 7.21 a | 32.24 ± 3.28 b |
| Red × 20 mM NaCl | 1892 ± 152 | 6.39 ± 0.49 | 1.54 ± 0.08 | 68.69 ± 4.22 a | 47.36 ± 2.86 a |
| Red × 30 mM NaCl | 1705 ± 97.7 | 6.55 ± 0.81 | 1.52 ± 0.08 | 63.00 ± 3.91 ab | 23.14 ± 2.10 c |
| S × C | |||||
| 1 mM NaCl × Cut 1 | 1806 ± 66.2 | 6.75 ± 0.59 | 1.56 ± 0.03 | 68.56 ± 5.91 ab | 19.02 ± 1.04 c |
| 10 mM NaCl × Cut 1 | 1733 ± 91.8 | 6.87 ± 0.48 | 1.53 ± 0.06 | 70.96 ± 7.13 a | 21.15 ± 2.53 c |
| 20 mM NaCl × Cut 1 | 1682 ± 66.6 | 6.76 ± 0.38 | 1.63 ± 0.05 | 66.46 ± 5.54 ab | 33.75 ± 4.69 ab |
| 30 mM NaCl × Cut 1 | 1494 ± 28.2 | 6.85 ± 0.64 | 1.62 ± 0.04 | 62.42 ± 4.51 abc | 23.81 ± 2.32 bc |
| 1 mM NaCl × Cut 2 | 2443 ± 210 | 4.87 ± 0.57 | 1.30 ± 0.05 | 49.21 ± 2.23 cd | 21.75 ± 2.41 c |
| 10 mM NaCl × Cut 2 | 2364 ± 122 | 5.27 ± 0.72 | 1.36 ± 0.02 | 45.95 ± 4.90 d | 33.41 ± 2.86 ab |
| 20 mM NaCl × Cut 2 | 2167 ± 94.0 | 4.87 ± 0.38 | 1.39 ± 0.07 | 53.31 ± 4.29 bcd | 37.33 ± 6.65 a |
| 30 mM NaCl × Cut 2 | 1912 ± 52.9 | 4.86 ± 0.40 | 1.32 ± 0.07 | 53.55 ± 3.35 bcd | 19.08 ± 2.84 c |
| CV × C | |||||
| Green × Cut 1 | 1737 ± 52.9 | 5.88 ± 0.20 | 1.56 ± 0.03 | 56.95 ± 2.07 b | 21.42 ± 1.54 c |
| Green × Cut 2 | 2301 ± 138 | 4.21 ± 0.20 | 1.32 ± 0.04 | 45.25 ± 2.45 c | 22.03 ± 2.00 c |
| Red × Cut 1 | 1620 ± 55.8 | 7.73 ± 0.25 | 1.61 ± 0.04 | 77.24 ± 3.14 a | 27.45 ± 3.11 b |
| Red × Cut 2 | 2142 ± 59.1 | 5.72 ± 0.35 | 1.36 ± 0.03 | 55.76 ± 2.03 b | 33.76 ± 3.93 a |
| Significance | |||||
| Cultivar (CV) | ns | *** | ns | *** | *** |
| Salinity (S) | ** | ns | ns | ns | *** |
| Cut (C) | *** | *** | *** | *** | * |
| CV × S | ns | ns | ns | * | *** |
| S × C | ns | ns | ns | * | *** |
| CV × C | ns | ns | ns | * | * |
| CV × S × C | ns | ns | ns | ns | ns |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carillo, P.; Giordano, M.; Raimondi, G.; Napolitano, F.; Di Stasio, E.; Kyriacou, M.C.; Sifola, M.I.; Rouphael, Y. Physiological and Nutraceutical Quality of Green and Red Pigmented Lettuce in Response to NaCl Concentration in Two Successive Harvests. Agronomy 2020, 10, 1358. https://doi.org/10.3390/agronomy10091358
Carillo P, Giordano M, Raimondi G, Napolitano F, Di Stasio E, Kyriacou MC, Sifola MI, Rouphael Y. Physiological and Nutraceutical Quality of Green and Red Pigmented Lettuce in Response to NaCl Concentration in Two Successive Harvests. Agronomy. 2020; 10(9):1358. https://doi.org/10.3390/agronomy10091358
Chicago/Turabian StyleCarillo, Petronia, Maria Giordano, Giampaolo Raimondi, Francesco Napolitano, Emilio Di Stasio, Marios C. Kyriacou, Maria Isabella Sifola, and Youssef Rouphael. 2020. "Physiological and Nutraceutical Quality of Green and Red Pigmented Lettuce in Response to NaCl Concentration in Two Successive Harvests" Agronomy 10, no. 9: 1358. https://doi.org/10.3390/agronomy10091358
APA StyleCarillo, P., Giordano, M., Raimondi, G., Napolitano, F., Di Stasio, E., Kyriacou, M. C., Sifola, M. I., & Rouphael, Y. (2020). Physiological and Nutraceutical Quality of Green and Red Pigmented Lettuce in Response to NaCl Concentration in Two Successive Harvests. Agronomy, 10(9), 1358. https://doi.org/10.3390/agronomy10091358