Aldehyde Dehydrogenase 2 Ameliorates Chronic Alcohol Consumption-Induced Atrial Fibrillation through Detoxification of 4-HNE

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Generation of ALDH2*2 Knock-In (KI) Mice
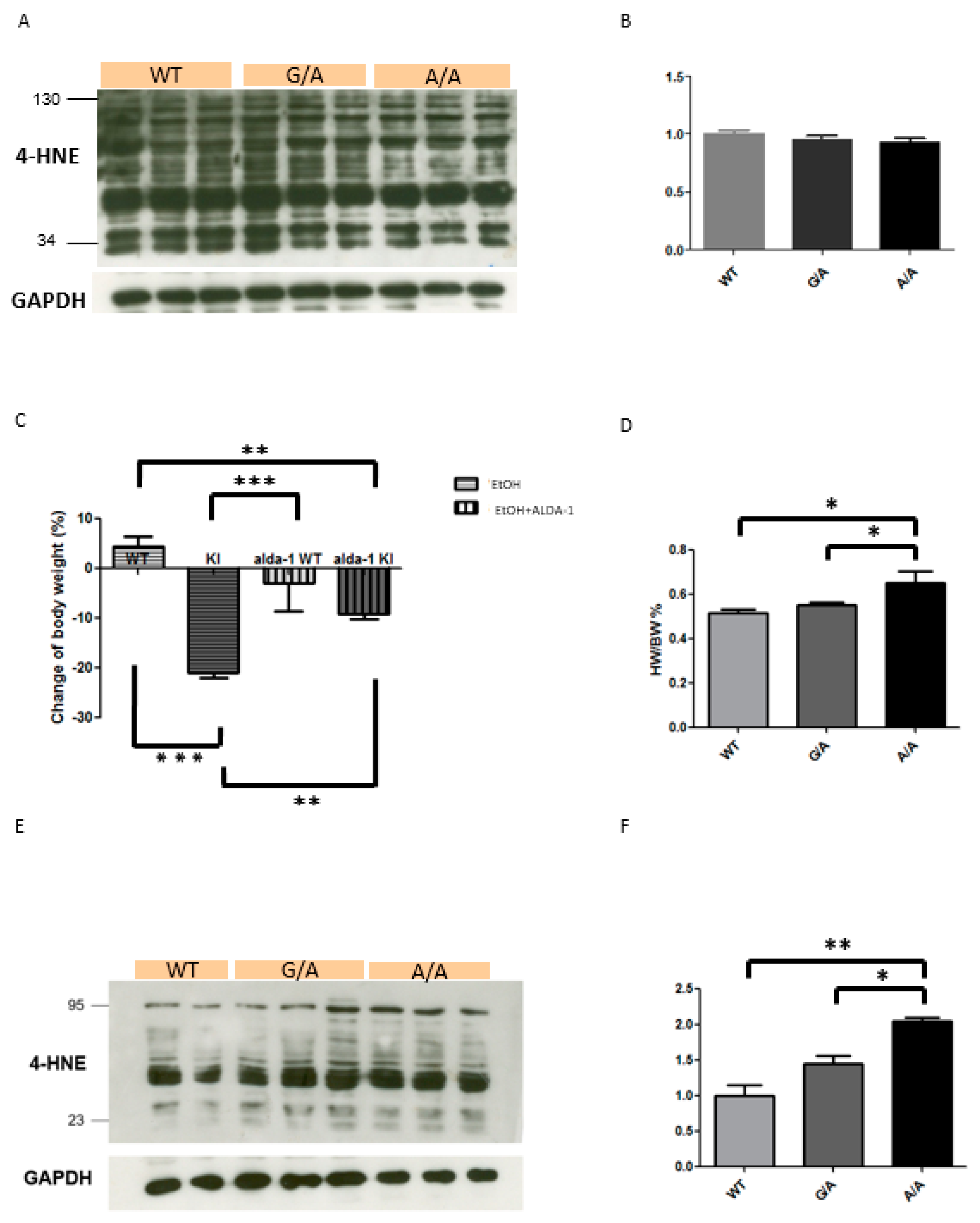
2.2. Increased 4-HNE Production in the ALDH2*2 KI Mice with Chronic Ethanol Intoxication
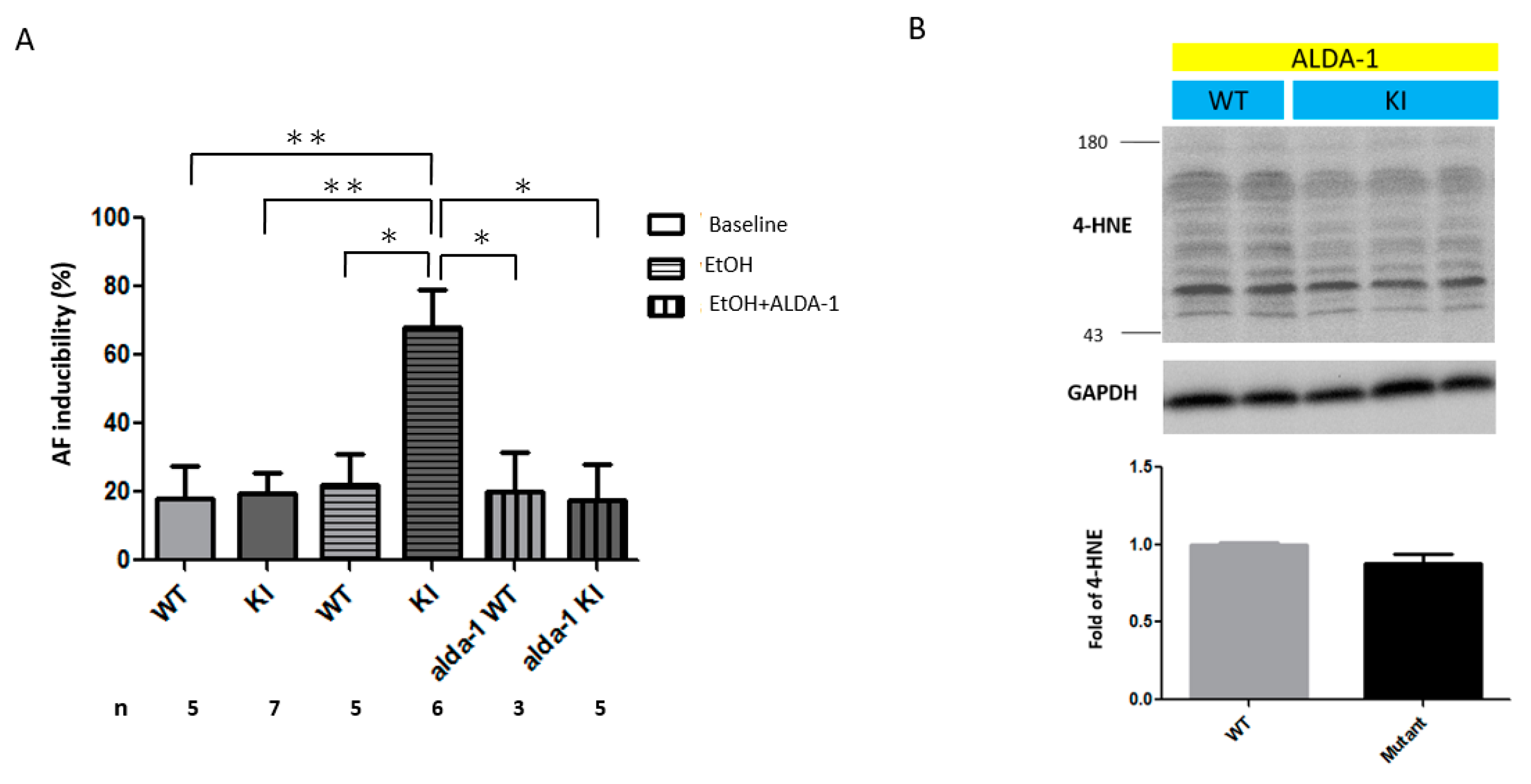
2.3. Increased AF Inducibility in the ALDH2*2 KI Mice with Chronic Ethanol Intoxication
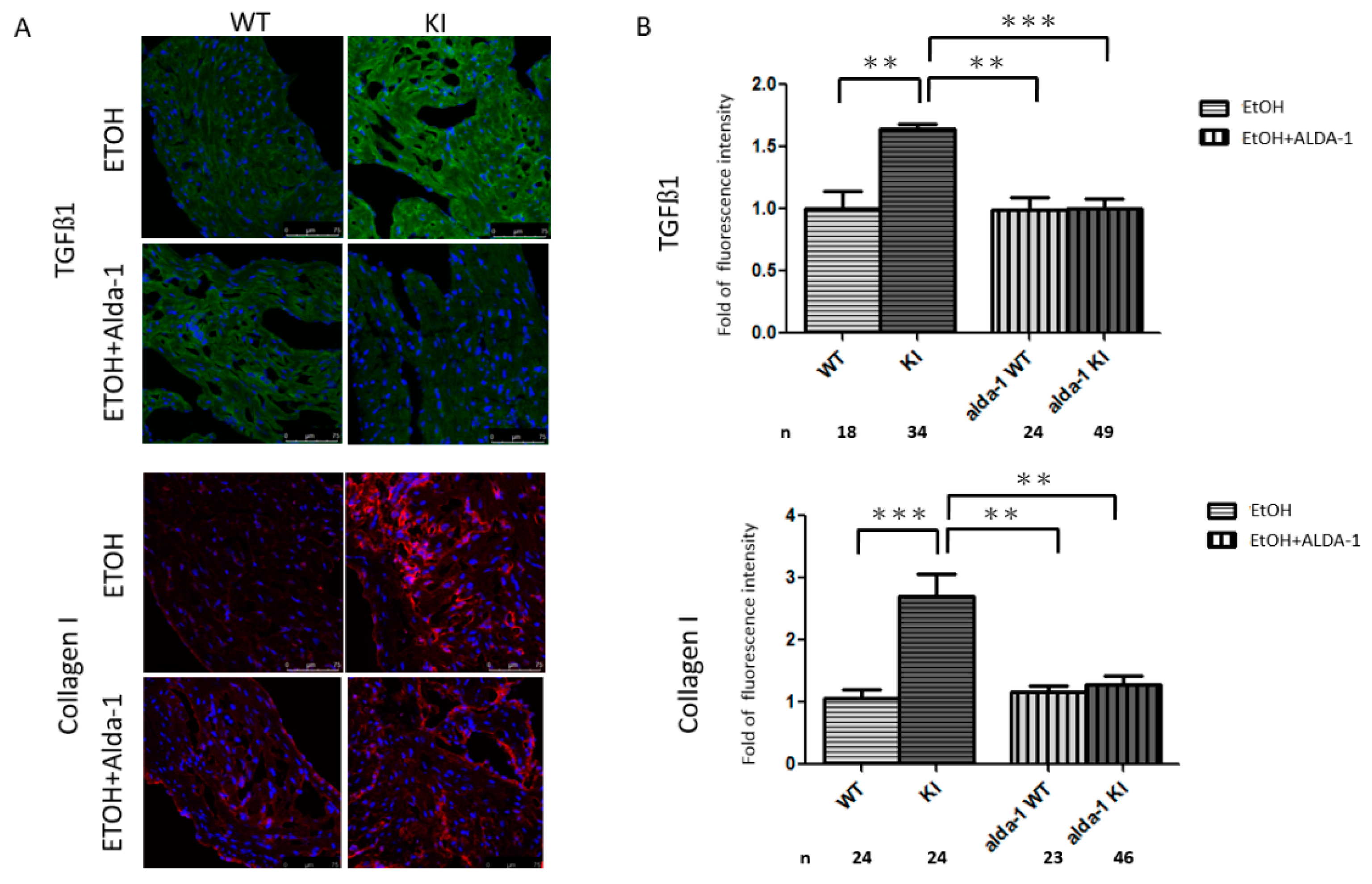
2.4. Increased Atrial Fibrosis in the ALDH2*2 KI Mice with Chronic Ethanol Consumption
2.5. In Vivo Treatment with Alda-1 (ALDH2 Activator)
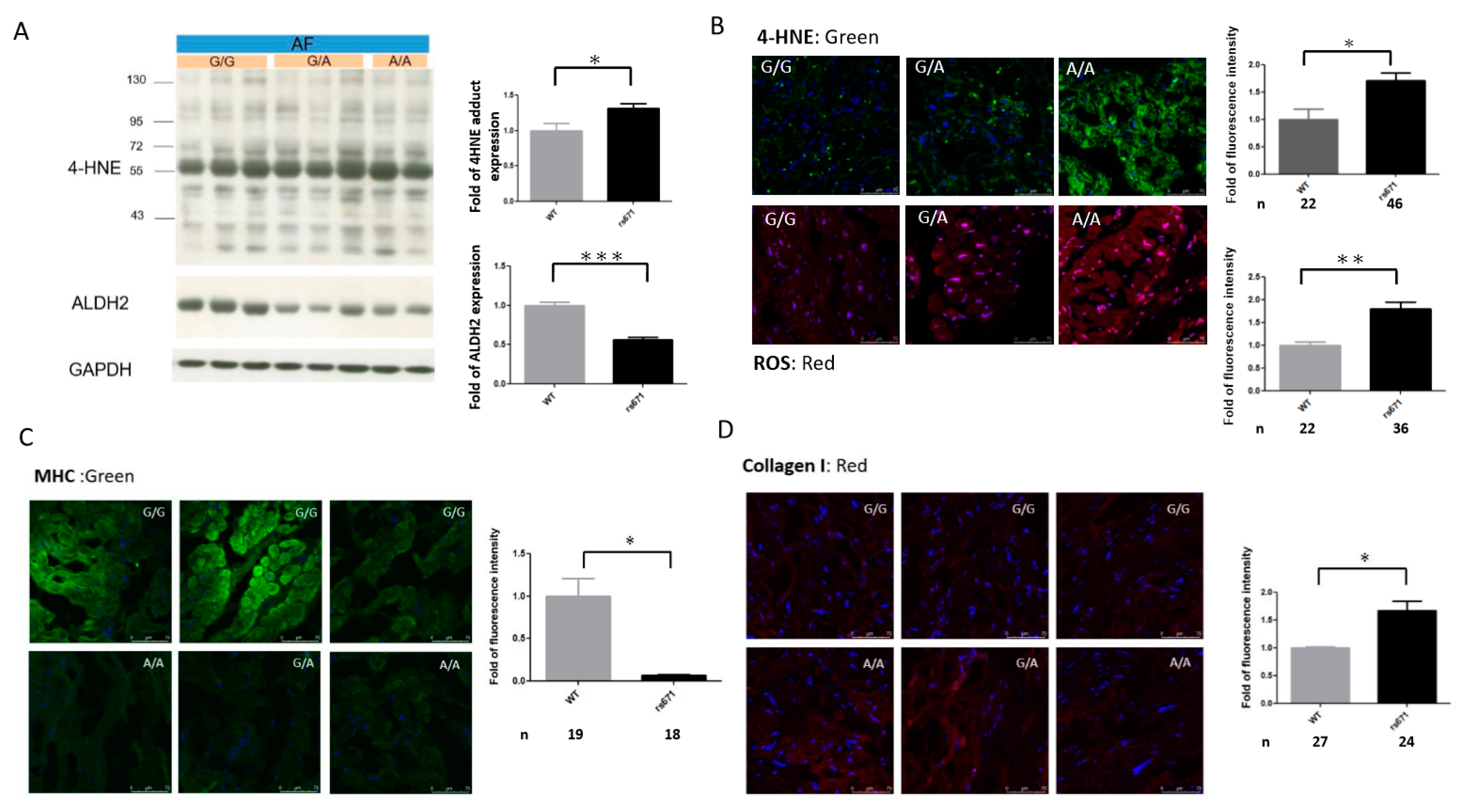
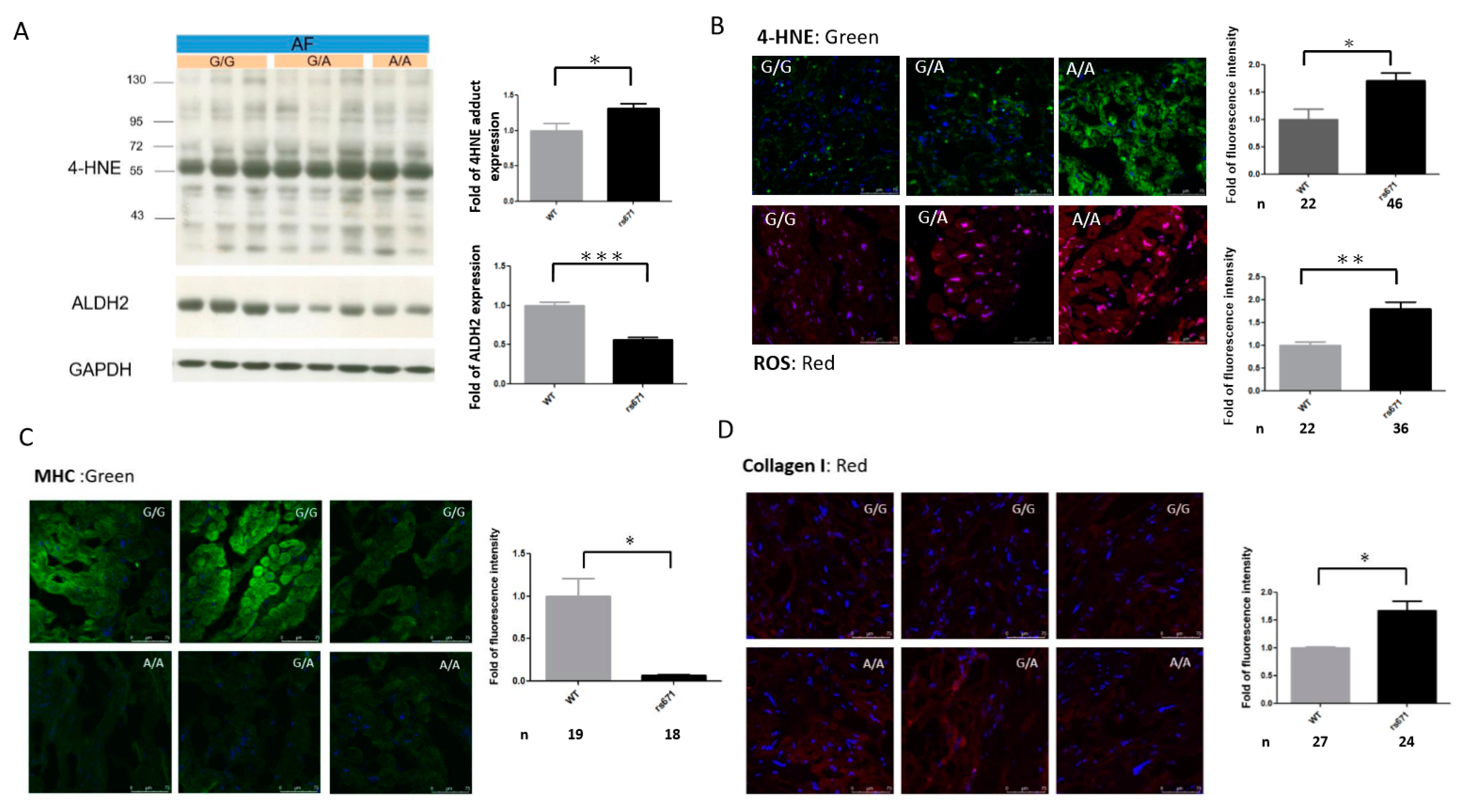
2.6. Association of the ALDH2*2 Polymorphism with Oxidative Stress and Structural Remodeling in Human AF Tissues
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Human Samples
4.3. Generation of ALDH2*2 KI Mice Using CRISPR/CAS9
4.4. Mice Maintenance
4.5. Ethanol Challenge
4.6. ALDH2 Enzymatic Activity
4.7. Western Blot Analysis
4.8. Programmed Electrical Stimulation
4.9. Histology and Immunohistochemistry
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Chugh, S.S.; Havmoeller, R.; Narayanan, K.; Singh, D.; Rienstra, M.; Benjamin, E.J.; Gillum, R.F.; Kim, Y.H.; McAnulty, J.H., Jr.; Zheng, Z.J.; et al. Worldwide epidemiology of atrial fibrillation: A global burden of disease 2010 study. Circulation 2014, 129, 837–847. [Google Scholar] [CrossRef] [Green Version]
- Korantzopoulos, P.; Kolettis, T.M.; Galaris, D.; Goudevenos, J.A. The role of oxidative stress in the pathogenesis and perpetuation of atrial fibrillation. Int. J. Cardiol. 2007, 115, 135–143. [Google Scholar] [CrossRef]
- Budas, G.R.; Disatnik, M.H.; Mochly-Rosen, D. Aldehyde dehydrogenase 2 in cardiac protection: A new therapeutic target? Trends Cardiovasc. Med. 2009, 19, 158–164. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.H.; Budas, G.R.; Churchill, E.N.; Disatnik, M.H.; Hurley, T.D.; Mochly-Rosen, D. Activation of aldehyde dehydrogenase-2 reduces ischemic damage to the heart. Science 2008, 321, 1493–1495. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, S.; Chandra, A.; Wang, L.F.; Seifert, W.E., Jr.; DaGue, B.B.; Ansari, N.H.; Srivastava, S.K.; Bhatnagar, A. Metabolism of the lipid peroxidation product, 4-hydroxy-trans-2-nonenal, in isolated perfused rat heart. J. Biol. Chem. 1998, 273, 10893–10900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esterbauer, H.; Schaur, R.J.; Zollner, H. Chemistry and biochemistry of 4-hydroxynonenal, malonaldehyde and related aldehydes. Free Radic. Biol. Med. 1991, 11, 81–128. [Google Scholar] [CrossRef]
- Mali, V.R.; Palaniyandi, S.S. Regulation and therapeutic strategies of 4-hydroxy-2-nonenal metabolism in heart disease. Free Radical Res. 2014, 48, 251–263. [Google Scholar] [CrossRef] [PubMed]
- Stewart, M.J.; Malek, K.; Crabb, D.W. Distribution of messenger RNAs for aldehyde dehydrogenase 1, aldehyde dehydrogenase 2, and aldehyde dehydrogenase 5 in human tissues. J. Investig. Med. 1996, 44, 42–46. [Google Scholar]
- Yoshida, A.; Huang, I.Y.; Ikawa, M. Molecular abnormality of an inactive aldehyde dehydrogenase variant commonly found in Orientals. Proc. Natl. Acad. Sci. USA 1984, 81, 258–261. [Google Scholar] [CrossRef] [Green Version]
- Goedde, H.W.; Agarwal, D.P.; Harada, S.; Meier-Tackmann, D.; Ruofu, D.; Bienzle, U.; Kroeger, A.; Hussein, L. Population genetic studies on aldehyde dehydrogenase isozyme deficiency and alcohol sensitivity. Am. J. Hum. Genet. 1983, 35, 769–772. [Google Scholar]
- Larso, H.N.; Weiner, H.; Hurley, T.D. Disruption of the coenzyme binding site and dimer interface revealed in the crystal structure of mitochondrial aldehyde dehydrogenase ‘Asian’ variant. J. Biol. Chem. 2005, 280, 30550–30556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seitz, H.K.; Matsuzaki, S.; Yokoyama, A.; Homann, N.; Vakevainen, S.; Wang, X.D. Alcohol and cancer. Alcohol Clin. Exp. Res. 2001, 25, 137–143. [Google Scholar] [CrossRef]
- Chang, J.S.; Hsiao, J.R.; Chen, C.H. ALDH2 polymorphism and alcohol-related cancers in Asians: A public health perspective. J. Biomed. Sci. 2017, 24, 19–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Zhang, D.; Jin, W.; Shao, C.; Yan, P.; Xu, C.; Sheng, H.; Liu, Y.; Yu, J.; Xie, Y.; et al. Mitochondrial aldehyde dehydrogenase-2 (ALDH2) Glu504Lys polymorphism contributes to the variation in efficacy of sublingual nitroglycerin. J. Clin. Investig. 2006, 116, 506–511. [Google Scholar] [CrossRef] [Green Version]
- Mizuno, Y.; Harada, E.; Morita, S.; Kinoshita, K.; Hayashida, M.; Shono, M.; Morikawa, Y.; Murohara, T.; Nakayama, M.; Yoshimura, M.; et al. East Asian Variant of Aldehyde Dehydrogenase 2 Is Associated with Coronary Spastic Angina: Possible Roles of Reactive Aldehydes and Implications of Alcohol Flushing Syndrome. Circulation 2015, 131, 1665–1673. [Google Scholar] [CrossRef] [Green Version]
- Jo, S.A.; Kim, E.K.; Park, M.H.; Han, C.; Park, H.Y.; Jang, Y.; Song, B.J.; Jo, I. A Glu487Lys polymorphism in the gene for mitochondrial aldehyde dehydrogenase 2 is associated with myocardial infarction in elderly Korean men. Clin. Chim. Acta 2007, 382, 43–47. [Google Scholar] [CrossRef]
- Takagi, S.; Iwai, N.; Yamauchi, R.; Kojima, S.; Yasuno, S.; Baba, T.; Terashima, M.; Tsutsumi, Y.; Suzuki, S.; Morii, I.; et al. Aldehyde dehydrogenase 2 gene is a risk factor for myocardial infarction in Japanese men. Hypertens. Res. 2002, 25, 677–681. [Google Scholar] [CrossRef] [Green Version]
- Amamoto, K.; Okamura, T.; Tamaki, S.; Kita, Y.; Tsujita, Y.; Kadowaki, T.; Nakamura, Y.; Ueshima, H. Epidemiologic study of the association of low-Km mitochondrial acetaldehyde dehydrogenase genotypes with blood pressure level and the prevalence of hypertension in a general population. Hypertens. Res. 2002, 25, 857–864. [Google Scholar] [CrossRef] [Green Version]
- Sung, Y.F.; Lu, C.C.; Lee, J.T.; Hung, Y.J.; Hu, C.J.; Jeng, J.S.; Chiou, H.Y.; Peng, G.S. Homozygous ALDH2*2 Isan Independent Risk Factor for Ischemic Stroke in Taiwanese Men. Stroke 2016, 47, 2174–2179. [Google Scholar] [CrossRef] [Green Version]
- Marchitti, S.A.; Deitrich, R.A.; Vasiliou, V. Neurotoxicity and metabolism of the catecholamine derived 3,4-dihydroxyphenylacetaldehyde and 3,4-dihydroxyphenylglycolaldehyde: The role of aldehyde dehydrogenase. Pharmacol. Rev. 2007, 59, 125–150. [Google Scholar] [CrossRef]
- Larsson, S.C.; Drca, N.; Wolk, A. Alcohol consumption and risk of atrial fibrillation: A prospective study and dose-response meta-analysis. J. Am. Coll. Cardiol. 2014, 64, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Q.; Weiner, H.; Crabb, D.W. The mutation in the mitochondrial aldehyde dehydrogenase (ALDH2) gene responsible for alcohol-induced flushing increases turnover of the enzyme tetramers in a dominant fashion. J. Clin. Investig. 1996, 98, 2027–2032. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.; Chen, J.; Chen, L.; Histen, G.; Lin, Z.; Gross, S.; Hixon, J.; Chen, Y.; Kung, C.; Chen, Y.; et al. ALDH2 (E487K) mutation increases protein turnover and promotes murine hepatocarcinogenesis. Proc. Natl. Acad. Sci. USA 2015, 112, 9088–9093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ki, S.H.; Park, O.; Zheng, M.; Morales-Ibanez, O.; Kolls, J.K.; Bataller, R.; Gao, B. Interleukin-22 Treatment Ameliorates Alcoholic Liver Injury in a Murine Model of Chronic-Binge Ethanol Feeding: Role of Signal Transducer and Activator of Transcription 3. Hepatology 2010, 52, 1291–1300. [Google Scholar] [CrossRef] [Green Version]
- Purohit, V.; Brenner, D.A. Mechanisms of alcohol-induced hepatic fibrosis: A summary of the Ron Thurman Symposium. Hepatology 2006, 43, 872–878. [Google Scholar] [CrossRef] [PubMed]
- Zambelli, V.O.; Gross, E.R.; Chen, C.H.; Gutierrez, V.P.; Cury, Y.; Mochly-Rosen, D. Aldehyde dehydrogenase-2 regulates nociception in rodent models of acute inflammatory pain. Sci. Transl. Med. 2014, 6, 251ra118. [Google Scholar] [CrossRef] [Green Version]
- Voskoboinik, A.; Kalman, J.M.; De Silva, A.; Nicholls, T.; Costello, B.; Nanayakkara, S.; Prabhu, S.; Stub, D.; Azzopardi, S.; Vizi, D.; et al. Alcohol Abstinence in Drinkers with Atrial Fibrillation. N. Engl. J. Med. 2020, 382, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Ettinger, P.O.; Wu, C.F.; De La Cruz, C., Jr.; Weisse, A.B.; Ahmed, S.S.; Regan, T.J. Arrhythmias and the “Holiday Heart”: Alcohol associated cardiac rhythm disorders. Am. Heart J. 1978, 95, 555–562. [Google Scholar] [CrossRef]
- Sengul, C.; Cevik, C.; Ozveren, O.; Sunbul, A.; Oduncu, V.; Akgun, T.; Can, M.M.; Semiz, E.; Dindar, I. Acute alcohol consumption is associated with increased interatrial electromechanical delay in healthy men. Cardiol. J. 2011, 18, 682–686. [Google Scholar] [CrossRef]
- George, A.; Figueredo, V.M. Alcohol and arrhythmias: A comprehensive review. J. Cardiovasc. Med. 2010, 11, 221–228. [Google Scholar] [CrossRef] [Green Version]
- Piano, M.R. Alcoholic cardiomyopathy: Incidence, clinical characteristics, and pathophysiology. Chest 2002, 121, 1638–1650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcus, G.M.; Smith, L.M.; Whiteman, D.; Tseng, Z.H.; Badhwar, N.; Lee, B.K.; Lee, R.J.; Scheinman, M.M.; Olgin, J.E. Alcohol intake is significantly associated with atrial flutter in patients under 60 years of age and a shorter right atrial effective refractory period. Pacing Clin. Electrophysiol. 2008, 31, 266–272. [Google Scholar] [CrossRef] [PubMed]
- Mandyam, M.C.; Vedantham, V.; Scheinman, M.M.; Tseng, Z.H.; Badhwar, N.; Lee, B.K.; Lee, R.J.; Gerstenfeld, E.P.; Olgin, J.E.; Marcus, G.M. Alcohol and vagal tone as triggers for paroxysmal atrial fibrillation. Am. J. Cardiol. 2012, 110, 364–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Youn, J.Y.; Zhang, J.; Zhang, Y.; Chen, H.; Liu, D.; Ping, P.; Weiss, J.N.; Cai, H. Oxidative stress in atrial fibrillation: An emerging role of NADPH oxidase. J. Mol. Cell Cardiol. 2013, 62, 72–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nattel, S. Molecular and Cellular Mechanisms of Atrial Fibrosis in Atrial Fibrillation. JACC Clin. Electrophysiol. 2017, 3, 425–435. [Google Scholar] [CrossRef]
- Nakano, Y.; Ochi, H.; Onohara, Y.; Sairaku, A.; Tokuyama, T.; Matsumura, H.; Tomomori, S.; Amioka, M.; Hironomobe, N.; Motoda, C.; et al. Genetic Variations of Aldehyde Dehydrogenase 2 and Alcohol Dehydrogenase 1B Are Associated with the Etiology of Atrial Fibrillation in Japanese. J. Biomed. Sci. 2016, 23, 89. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.C.; Chiu, Y.F.; Lee, I.T.; Ho, L.T.; Hung, Y.J.; Hsiung, C.A.; Quertermous, T.; Donlon, T.; Lee, W.J.; Lee, P.C.; et al. Common ALDH2 Genetic Variants Predict Development of Hypertension in the SAPPHIRe Prospective Cohort: Gene-Environmental Interaction with Alcohol Consumption. BMC Cardiovasc. Disord. 2012, 12, 58. [Google Scholar] [CrossRef] [Green Version]
- Schotten, U.; Verheule, S.; Kirchhof, P.; Goette, A. Pathophysiological mechanisms of atrial fibrillation: A translational appraisal. Physiol. Rev. 2011, 91, 265–325. [Google Scholar] [CrossRef]
- Yang, Z.; Shen, W.; Rottman, J.N.; Wilswo, J.P.; Marray, K.T. Rapid stimulation causes electrical remodeling in cultured atrial myocytes. J. Mol. Cell Cardiol. 2005, 38, 299–308. [Google Scholar] [CrossRef]
- Mace, L.C.; Yermalitskaya, L.V.; Yi, Y.; Yang, Z.; Morgan, A.M.; Murray, K.T. Transcriptional remodeling of rapidly stimulated HL-1 atrial myocytes exhibits concordance with human atrial fibrillation. J. Mol. Cell Cardiol. 2009, 47, 485–492. [Google Scholar] [CrossRef] [Green Version]
- Yeh, Y.H.; Kuo, C.T.; Chan, T.H.; Chang, G.J.; Qi, X.Y.; Tsai, F.; Nattel, S.; Chen, W.J. Transforming growth factor-beta and oxidative stress mediate tachycardia-induced cellular remodeling in cultured atrial-derived myocytes. Cardiovasc. Res. 2011, 91, 62–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, T.; Liu, S.; Ma, T.; Jia, Z.; Zhang, Z.; Wang, A. Aldehyde dehydrogenase2 protects against oxidative stress associated with pulmonary arterial hypertension. Redox. Biol. 2017, 11, 286–296. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Xu, D.; Wang, S.; Fu, H.; Wang, K.; Zou, Y.; Sun, A.; Ge, J. Inhibition of aldehyde dehydrogenase 2 activity enhances antimycin-induced rat cardiomyocytes apoptosis through activation of MAPK signaling pathway. Biomed. Pharmacother. 2011, 65, 590–593. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Guo, R.; Yu, L.; Zhang, Y.; Ren, J. Aldehyde dehydrogenase 2 (ALDH2) rescues myocardial ischaemia/reperfusion injury: Role of autophagy paradox and toxic aldehyde. Eur. Heart J. 2011, 32, 1025–1038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, S.H.; Hsu, L.A.; Tsai, H.Y.; Yeh, Y.H.; Lu, C.Y.; Chen, P.C.; Wang, J.C.; Chiu, Y.L.; Lin, C.Y.; Hsu, Y.J. Aldehyde Dehydrogenase 2 Protects Against Abdominal Aortic Aneurysm Formation by Reducing Reactive Oxygen Species, Vascular Inflammation, and Apoptosis of Vascular Smooth Muscle Cells. FASEB J. 2020. [Google Scholar] [CrossRef] [PubMed]
- Sung, Y.H.; Jin, Y.; Kim, S.; Lee, H.W. Generation of knockout mice using engineered nucleases. Methods 2014, 69, 85–93. [Google Scholar] [CrossRef]
- Verheule, S.; Sato, T.; Everett, T.; Engle, S.K.; Otten, D.; Rubart-Von Der Lohe, M.; Nakajima, H.O.; Nakajima, H.; Field, L.J.; Olgin, J.E. Increased vulnerability to atrial fibrillation in transgenic mice with selective atrial fibrosis caused by overexpression of TGF-beta1. Circ. Res. 2004, 94, 1458–1465. [Google Scholar] [CrossRef] [Green Version]
- Yeh, Y.H.; Hsu, L.A.; Chen, Y.H.; Kuo, C.T.; Chang, G.J.; Chen, W.J. Protective Role of Heme oxygenase-1 in Atrial Remodeling. Basic Res. Cardiol. 2016, 111, 58. [Google Scholar] [CrossRef]

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsu, L.-A.; Tsai, F.-C.; Yeh, Y.-H.; Chang, C.-J.; Kuo, C.-T.; Chen, W.-J.; Tsai, H.-Y.; Chang, G.-J. Aldehyde Dehydrogenase 2 Ameliorates Chronic Alcohol Consumption-Induced Atrial Fibrillation through Detoxification of 4-HNE. Int. J. Mol. Sci. 2020, 21, 6678. https://doi.org/10.3390/ijms21186678
Hsu L-A, Tsai F-C, Yeh Y-H, Chang C-J, Kuo C-T, Chen W-J, Tsai H-Y, Chang G-J. Aldehyde Dehydrogenase 2 Ameliorates Chronic Alcohol Consumption-Induced Atrial Fibrillation through Detoxification of 4-HNE. International Journal of Molecular Sciences. 2020; 21(18):6678. https://doi.org/10.3390/ijms21186678
Chicago/Turabian StyleHsu, Lung-An, Feng-Chun Tsai, Yung-Hsin Yeh, Chi-Jen Chang, Chi-Tai Kuo, Wei-Jan Chen, Hsin-Yi Tsai, and Gwo-Jyh Chang. 2020. "Aldehyde Dehydrogenase 2 Ameliorates Chronic Alcohol Consumption-Induced Atrial Fibrillation through Detoxification of 4-HNE" International Journal of Molecular Sciences 21, no. 18: 6678. https://doi.org/10.3390/ijms21186678
APA StyleHsu, L.-A., Tsai, F.-C., Yeh, Y.-H., Chang, C.-J., Kuo, C.-T., Chen, W.-J., Tsai, H.-Y., & Chang, G.-J. (2020). Aldehyde Dehydrogenase 2 Ameliorates Chronic Alcohol Consumption-Induced Atrial Fibrillation through Detoxification of 4-HNE. International Journal of Molecular Sciences, 21(18), 6678. https://doi.org/10.3390/ijms21186678