PFAS Environmental Pollution and Antioxidant Responses: An Overview of the Impact on Human Field


 and
and 
Abstract
:1. Introduction
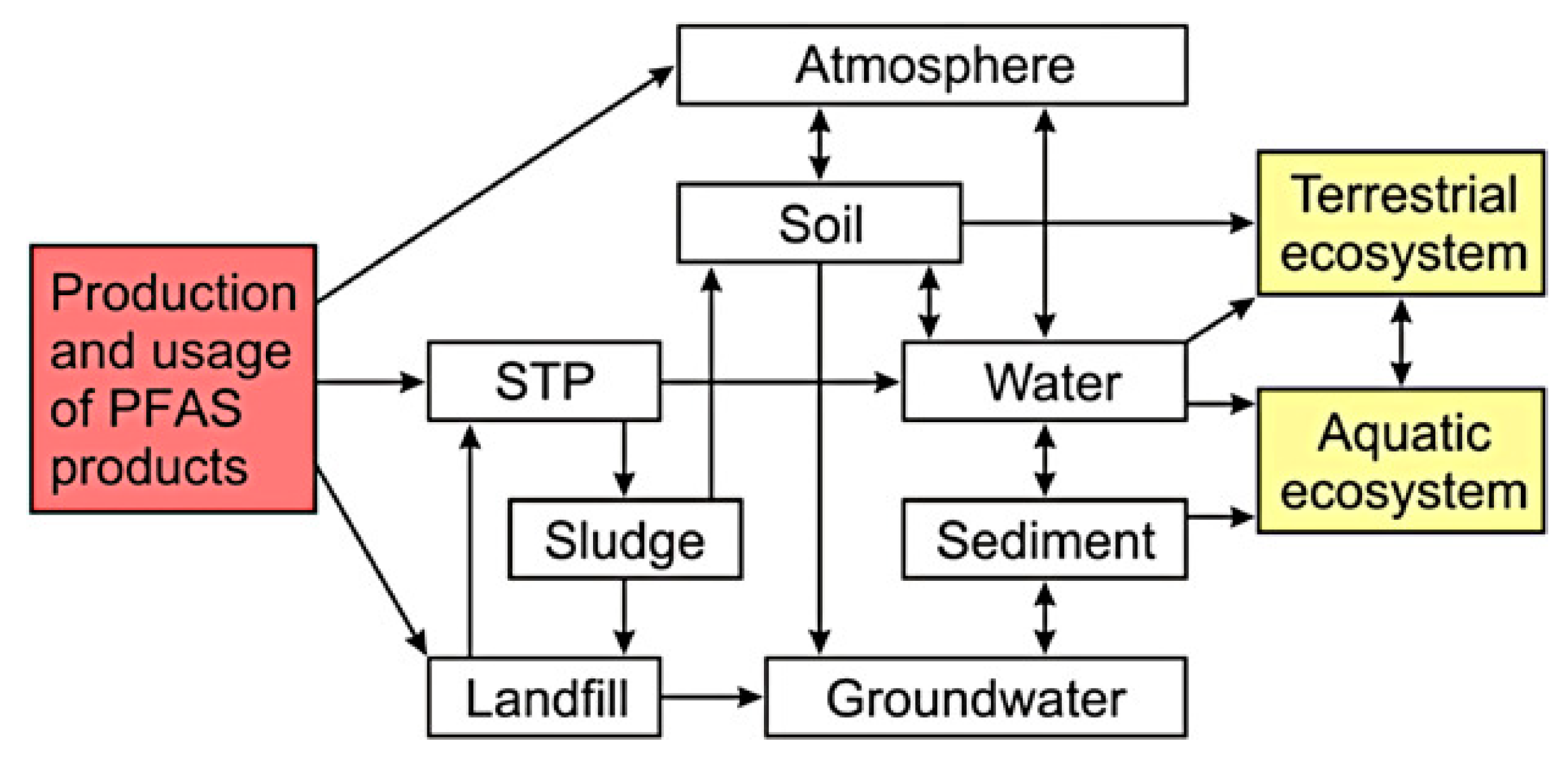
2. PFAS and Their Distribution in the Environment
2.1. First Case: The USA
2.2. European Situation
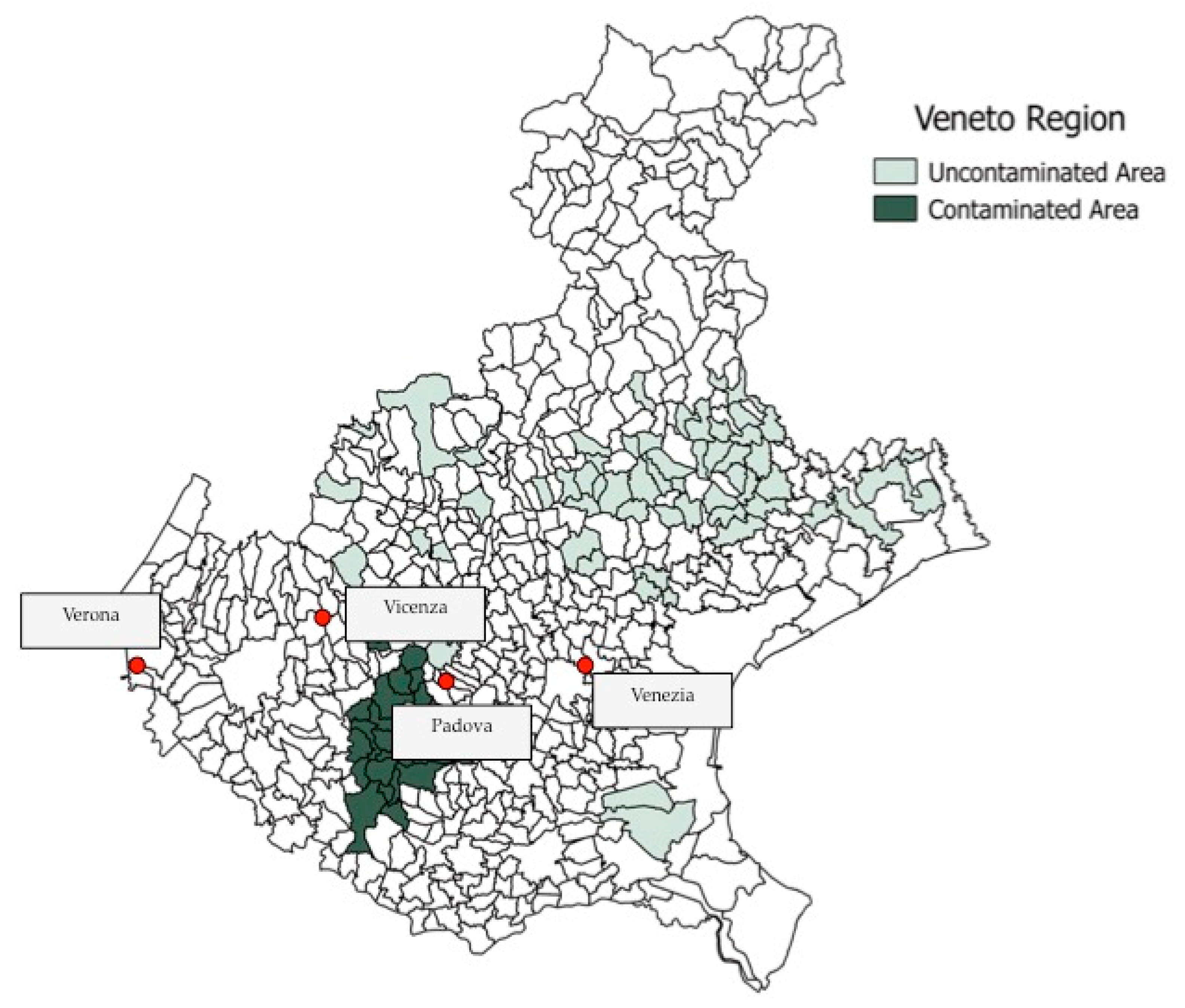
2.3. Veneto Region
3. Epidemiological Studies
3.1. The C8 Health Project
- Helsingør Statement on Poly- and Perfluorinated Alkyl substances (PFAS) [137];
- The Madrid Statement on Poly- and Perfluoroalkyl Substances (PFAS) [110];
- Zürich Statement on Future Actions on Per- and Polyfluoroalkyl Substances (PFAS) [138].
- These statements increased the consciousness of researchers, managers and communities on the PFAS issues.
3.2. Studies after the C8 Health Project and Panel
4. Mammals: In Vitro and In Vivo Studies Correlating PFAS Levels and Oxidative Stress
4.1. PFAS Affect Oxidative Stress Biomarkers In Vitro
4.2. PFAS Affect Oxidative Stress Biomarkers In Vivo
5. Oxidative Stress in Human Health Due to PFAS Pollution
5.1. Male Infertility
5.2. Female Infertility
5.3. Pregnant Women and Developmental Consequences
5.4. Kidney and Thyroid Disorders
5.5. Cholesterol
5.6. Diabetes
5.7. Platelets and Cardiovascular Desease
6. PFAS and Oxidative Stress in Other Vertebrates
6.1. Chemical Data
6.2. Biochemical Data
6.3. Biomolecular Data

{kind=link}
{kind=link}
| Species | PFAS | Effects | References |
|---|---|---|---|
| Carassius auratus | PFOA | Increased ROS formation 1, lipid peroxidation 2, GSH decrease 3, SOD induction 4, CAT induction 4, GPX inhibition 4, GR induction 4 | [184] |
| Danio rerio | PFOA | CAT inhibition 7, cat down-regulation 8, prdx down-regulation 8 | [281] |
| Danio rerio | PFOS | Increased ROS formation 1, lipid peroxidation 2, SOD induction 4, CAT induction 4, GPX induction 4, nrf2 up-regulation 5 | [307] |
| Danio rerio | PFOS | Increased ROS formation 1, lipid peroxidation 2, SOD induction 4, CAT inhibition 4, GPX induction 4, sod down-regulation 5, cat down-regulation 5, gpx down-regulation 5 | [308] |
| Danio rerio | PFOS | Increased ROS formation 1, SOD induction 4, GPX induction 4 | [309] |
| Danio rerio | PFNA | Increased ROS formation 1 | [310] |
| Danio rerio | PFOA, PFNA | Lipid peroxidation 6, gpx up-regulation 4, prdx up-regulation 5 | [314] |
| Danio rerio | PFPiAs | gpx up-regulation 5, prdx up-regulation 5, msr up-regulation 5 | [331] |
| Gallus gallus domesticus | PFOS | SOD induction 4 | [319] |
| Gobiocypris rarus | PFOA | gpx down-regulation 5, prdx up-regulation 5, msr up-regulation 5 | [329] |
| Gobiocypris rarus | PFOA | gpx up-regulation 5 | [330] |
| Homo sapiens | PFOA, PFOS | Increased ROS formation 1, apoptosis 11, SOD induction 4, CAT induction 4, GR induction 4, GPX inhibition 4 | [192] |
| Homo sapiens | PFHxS, PFOA, PFOS, PFNA, PFDA, PFUnA, PFDoA | Increased ROS formation 1, increased DNA damage 9, antioxidant defense inhibition 10 | [196] |
| Homo sapiens | PFOA, PFOS | Estrogenicity and anti-estrogenicity 12 | [202] |
| Homo sapiens | PFOA, PFOS, PFNA | Estradiol production 13, progesterone 13 production, testosterone production 13 | [203] |
| Homo sapiens | PFOA, PFOS | Thyroid diseases | [213] |
| Homo sapiens | PFOA, PFOS | Chronic kidney diseases | [216] |
| Homo sapiens | PFHxS, PFOA, PFOS, PFNA | Decreased eGFR 14 | [217] |
| Homo sapiens | PFHxs, PFOA, PFOS | Increased cholesterol outcomes 15 | [218] |
| Homo sapiens | PFOA, PFOS, PFNA | Hyperglycemia 16, increased serum HDL cholesterol 16, increased blood insulin 17 | [219] |
| Homo sapiens | PFOA, PFOS | Increased blood levels of glucocorticoid 17, increased blood levels of androgenic hormones 18 | [222] |
| Homo sapiens | PFOA, PFOS | Increased total adiponectin levels 19, body weight decrease | [223] |
| Homo sapiens | PFHxS, PFHxA, PFHpA, PFOA, PFOS, PFNA, PFDA, PFUnDA, PFDoDA, PFTrDA, PFTeDA | Allergic diseases | [224] |
| Homo sapiens | PFOA, PFOS | Polyunsuturated fatty acids levels decrease 20, birth weight decrease | [225] |
| Homo sapiens | PFOA | Up-regulation of progesterone activated genes 5 | [242] |
| Homo sapiens | PFOA, PFOS | Increased T4 levels 21 | [260] |
| Homo sapiens | PFOA | Reduced fetal growth | [261] |
| Homo sapiens | PFOA, PFOS | Reduced birth weight | [262] |
| Homo sapiens | PFOA, PFOS | Reduced cord serum concentrations and birth weight size | [265] |
| Homo sapiens | PFOA, PFOS | Affected growth of organs and the skeleton | [267] |
| Homo sapiens | PFBS, PFHxA, PFHpA, PFHxS, PFOA, PFOS, PFNA, PFDA, PFTA, PFDoA | Asthma | [268] |
| Homo sapiens | PFHxS, PFOA, PFOS, PFNA | Asthma | [270] |
| Homo sapiens | PFOA, PFOS | High insulin and trygliceride concentrations | [272] |
| Homo sapiens | PFOA, PFOS, PFDeA | Changes in lipid profile 16, changes in ALT levels 16 | [274] |
| Homo sapiens | PFOA, PFOS, PFNA, PFHxS, PFDeA, PFOSA, Et-PFOSA, Me-PFOSA | Increased TSH22, increased eGFR 14 | [278] |
| Homo sapiens | PFOA, PFOS | Reduced birth weight | [284] |
| Homo sapiens | PFOA, PFOS | Increased cholesterol outcomes 16 | [287] |
| Homo sapiens | PFOA, PFOS, PFNA, PFHxS | High serum cholesterol 16 | [290] |
| Homo sapiens | PFOA, PFOS, PFNA, PFHxS | High serum cholesterol 16 | [291] |
| Homo sapiens | PFOA | HNF4α inhibition 23, hnf4α down-regulation 5 | [296] |
| Homo sapiens | PFOS, PFOA, PFHxS, PFNA, PFDA | Potential diabetogenic effect | [300] |
| Homo sapiens | PFOA, PFOS | Potential adiposity, decreased beta-cell function | [301] |
| Homo sapiens | PFOA | Impaired platelet aggregation and increased cardiovascular risk | [305] |
| Homo sapiens, Mus musculus | PFPA, PFHpA, PFOA, PFUnA, PFDoA | Increased PPARα activity 24 | [65] |
| Homo sapiens, Mus musculus | PFOA, PFOS, PFHxS, PFHxA, PFNA, PFDA, PFBA, PFBS | Increased PPARα activity 24 | [280] |
| Homo sapiens, Mus musculus, Rattus norvegicus | PFOA, PFOS | Increased PPARα activity 24, increased PPARβ activity 24, increased PPARγ activity 24 | [211] |
| Homo sapiens, Mus musculus, Rattus norvegicus | PFOA, PFOS | Reduced birth weight | [266] |
| Homo sapiens, Rattus norvegicus | PFOA, PFOS | Up-regulation of multiple nuclear receptor genes 5 | [285] |
| Micropterus salmoides | PFOS | gpx up-regulation 8, sod down-regulation 8, cat up-regulation 8, prdx down-regulation 8 | [332] |
| Mus musculus | PFOA | Lipid peroxidation 25, amylase induction, lipase induction, sod1 up-regulation 5, sod2 up-regulation5, gpx2 up-regulation 5, nqo1 up-regulation 5 | [204] |
| Mus musculus | PFOS | Increased in peroxisomal fatty acid beta-oxidation 4, increased peroxisomal catalase activity 4 | [208] |
| Mus musculus | PFOA | pparα up-regulation 5, pparγ up-regulation 5 | [209] |
| Mus musculus | PFOS | Serum testosterone decrease 19, epididymal sperm counts decrease, down-regulation of genes encoding testicular receptors for gonadotropin, growth hormone, insulin-like growth factor 1 and steroidogenic enzymes 5 | [234] |
| Mus musculus | PFOA | Damaged seminiferous tubules, reduced sperm quality, reduced serum testosterone and progesterone levels 13, INSL3 enzyme decrease 23, cytochrome P450 decrease 23 | [235] |
| Mus musculus | PFOS | Immune response induction 19 | [269] |
| Oncorhynchus mykiss | PFOA | CAT inhibition4 | [324] |
| Oreochromis niloticus | PFOA, PFOS | Increased ROS formation 1, lipid peroxidation 2, GSH decrease 3 | [306] |
| Oryzias latipes | PFOA | SOD inhibition 4, CAT inhibition 4 | [322] |
| Pelophylaxnigro maculatus | PFOA | nrf2 up-regulation 5 | [313] |
| Perca flavescens | PFHxS, PFOS, PFDS, PFECHS, PFOA, PFNA, PFDA, PFUnA, PFDoA, PFTriA, PFTetraA | SOD induction 4, CAT induction 4, cat down-regulation 5 | [321] |
| Pimephales promelas | PFOA, PFOS | Lipid peroxidation 6 | [315] |
| Rattus norvegicus | PFOA | Increased ROS formation 1, increased mithocondrial superoxide formation 25, increased nitric oxide formation 1, apoptosis 19, increased proinflammatory cytokines 19, reduced adenosine triphosphate levels 26, reduced cardiolipin peroxidation 27, reduced cytochrome c release 19 | [200] |
| Rattus norvegicus | PFOA | Increased ACO activity 4, increased PPARα activity 24 | [207] |
| Rattus norvegicus | PFDoA | Serum testosterone decrease 19, serum estradiol decrease 19, down-regulation of genes involved in cholesterol transport and steroid biosynthesis 5 | [233] |
| Rattus norvegicus | PFOA | Up-regulation of genes involved in metabolism of lipids, cell communication, growth, hormone regulatory pathways, proteolysis, peptidolysis and signal transduction 8, down-regulation of genes involved in inflammation and immunity, regulation of hormones, general metabolism and G-protein coupled receptor protein signaling pathways 8 | [294] |
| Salmo salar | PFOA, PFOS | sod up-regulation 5, cat up-regulation 5, gpx up-regulation5 | [328] |
| Ursus marittimus | PFASs | Increased levels of brain steroid hormones 28 | [295] |
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| acyl CoA oxidase | ACO |
| Alkaline Phosphatase | ALP |
| Aminotransferase | ALT |
| Androgen Receptor | AR |
| Bisphenol A | BPA |
| Catalase | CAT |
| Cyclooxygenase 2 | COX–2 |
| Dichlorodiphenyldichloroethylene | DDE |
| Endocrine Disruptors | ED |
| Glutamyl-Cysteine Ligase | GCL |
| Glomerular Filtration Rate | GFR |
| γ–glutamyl transferase | GGT |
| Glutathione peroxidase | GPx |
| Glutathione Reductase | GR |
| Glutathione S-transferase | GST |
| Hepatocyte nuclear factor 4 alpha | HNF4α |
| Health Protection Agency | HPA |
| insulin like factor 3 | INSL3 |
| Luteinizing Hormone | LH |
| Methionine sulfoxide reductases | Msr |
| Normal Granulosa Cells | N–GCs |
| National Health and Nutrition Examination Survey | NHANES |
| Nurses’ Health Study II | NHSII |
| Nitric Oxide | NO |
| Polychlorinated Biphenyls | PCBs |
| Granulosa Cells from Polycystic Ovaries | PCO–GCs |
| Perfluorinated Alkyl Substances | PFAS |
| Perfluorinated Carboxylic Acids | PFCA |
| Perfluorinated compounds | PFCs |
| Perfluorobutanoic acid | PFBA |
| Perfluorobutane sulfonate | PFBS |
| Perfluorodecanoic acid | PFDA |
| Perfluorododecanoic acid | PFDoA |
| Perfluoroheptanoic acid | PFHpA |
| Perfluorohexanoic acid | PFHxA |
| Perfluorohexanesulfonic acid | PFHxS |
| Perfluorononanoic acid | PFNA |
| Perfluorooctanoic acid | PFOA |
| Perfluorooctane sulfonic acid | PFOS |
| Perfluoropentanoic acid | PFPeA |
| Perfluoroalkyl phosphinic acid | PFPiAs |
| Perfluoroundecanoic acid | PFUnA |
| Persistent Mobile Organic Contaminants | PMOC |
| Peroxisome Proliferator-Activated Receptor | PPAR |
| Peroxiredoxins | Prdx |
| Reactive Oxygen Species | ROS |
| Stress Granules | SG |
| Superoxide Dismutase | SOD |
| Type 2 Diabetes | T2D |
| Thyroxine | T4 |
| Total Antioxidant Capacity | TAC |
| Total Cholesterol | TC |
| Tolerable Daily Intake | TDI |
| Triglycerides | TG |
References
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acworth, I.N.; McCabe, D.R.; Maher, T.J. The Analysis of Free Radicals, Their Reaction Products, and Antioxidants. In Oxidants, Antioxidants, and Free Radicals; Baskin, S.I., Salem, H., Eds.; Taylor Francis: Washington, DC, USA, 1997; pp. 23–77. [Google Scholar]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.D.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yi, J. Cancer cell killing via ROS: To increase or decrease, that is a question. Cancer Biol. Ther. 2008, 7, 1875–1884. [Google Scholar] [CrossRef] [PubMed]
- Lodovici, M.; Bigagli, E. Oxidative stress and air pollution exposure. J. Toxicol. 2011, 2011, 487074. [Google Scholar] [CrossRef] [PubMed]
- Federico, A.; Cardaioli, E.; Da Pozzo, P.; Formichi, P.; Gallus, G.N.; Radi, E. Mitochondria, oxidative stress and neurodegeneration. J. Neurol. Sci. 2012, 322, 254–262. [Google Scholar] [CrossRef]
- Zahn, D.; Neuwald, I.J.; Knepper, T.P. Analysis of mobile chemicals in the aquatic environment-current capabilities, limitations and future perspectives. Anal. Bioanal. Chem. 2020, 412, 4763–4784. [Google Scholar] [CrossRef] [PubMed]
- Gorbi, S.; Virno Lamberti, C.; Notti, A.; Benedetti, M.; Fattorini, D.; Moltedo, G.; Regoli, F. An ecotoxicological protocol with caged mussels, Mytilus galloprovincialis, for monitoring the impact of an offshore platform in the Adriatic Sea. Mar. Environ. Res. 2008, 65, 34–49. [Google Scholar] [CrossRef]
- Valko, M.; Jomova, K.; Rhodes, C.J.; Kuča, K.; Musílek, K. Redox- and non-redox-metal-induced formation of free radicals and their role in human disease. Arch. Toxicol. 2016, 90, 1–37. [Google Scholar] [CrossRef]
- Halliwell, B. Antioxidant defence mechanisms: From the beginning to the end (of the beginning). Free Radic. Res. 1999, 31, 261–272. [Google Scholar] [CrossRef]
- Ferro, K.; Ferro, D.; Corrà, F.; Bakiu, R.; Santovito, G.; Kurtz, J. Cu, Zn superoxide dismutase genes in Tribolium castaneum: Evolution, molecular characterisation, and gene expression during immune priming. Front. Immunol. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Chatzidimitriou, E.; Bisaccia, P.; Corrà, F.; Bonato, M.; Irato, P.; Manuto, L.; Toppo, S.; Bakiu, R.; Santovito, G. Copper/zinc superoxide dismutase from the crocodile icefish Chionodraco hamatus: Antioxidant defense at constant sub-zero temperature. Antioxidants 2020, 9, 325. [Google Scholar] [CrossRef] [PubMed]
- Muhtadi, R.; Lorenz, A.; Mpaulo, S.J.; Siebenwirth, C.; Scherthan, H. Catalase T-deficient fission yeast meiocytes show resistance to ionizing radiation. Antioxidants 2020, 9, 881. [Google Scholar] [CrossRef] [PubMed]
- Ferro, D.; Franchi, N.; Bakiu, R.; Ballarin, L.; Santovito, G. Molecular characterization and metal induced gene expression of the novel glutathione peroxidase 7 from the chordate invertebrate Ciona Robusta. Comp. Biochem. Physiol. C 2018, 205, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Al-Asadi, S.; Malik, A.; Bakiu, R.; Santovito, G.; Schuller, K. Characterization of the peroxiredoxin 1 subfamily from Tetrahymena thermophila. Cell. Mol. Life Sci. 2019, 76, 4745–4768. [Google Scholar] [CrossRef] [PubMed]
- Tolomeo, A.M.; Carraro, A.; Bakiu, R.; Toppo, S.; Garofalo, F.; Pellegrino, D.; Gerdol, M.; Ferro, D.; Place, S.P.; Santovito, G. Molecular characterization of novel mitochondrial peroxiredoxins from the Antarctic emerald rockcod and their gene expression in response to environmental warming. Comp. Biochem. Physiol. C 2019, 255, 108580. [Google Scholar] [CrossRef]
- Ricci, F.; Lauro, F.M.; Grzymski, J.J.; Read, R.; Bakiu, R.; Santovito, G.; Luporini, P.; Vallesi, A. The anti-oxidant defense system of the marine polar ciliate Euplotes nobilii: Characterization of the msrB gene family. Biology 2017, 6, 4. [Google Scholar] [CrossRef] [Green Version]
- Formigari, A.; Boldrin, F.; Santovito, G.; Cassidy-Hanley, D.; Clark, T.G.; Piccinni, E. Functional characterization of the 5′-upstream region of MTT5 metallothionein gene from Tetrahymena thermophila. Protist 2010, 161, 71–77. [Google Scholar] [CrossRef]
- Santovito, G.; Boldrin, F.; Irato, P. Metal and metallothionein distribution in different tissues of the Mediterranean clam Venerupis philippinarum during copper treatment and detoxification. Comp. Biochem. Physiol. C 2015, 174–175, 46–53. [Google Scholar] [CrossRef]
- Jakubczyk, K.; Kałduńska, J.; Dec, K.; Kawczuga, D.; Janda, K. Antioxidant properties of small-molecule non-enzymatic compounds. Pol. Merkur. Lekarski 2020, 48, 128–132. [Google Scholar]
- Haug, L. Approaches to integrating the toxicology and epidemiology evidence on PFAS. Environ. Epidemiol. 2019, 3, 210. [Google Scholar] [CrossRef]
- Cousins, I.T.; DeWitt, J.C.; Glüge, J.; Goldenman, G.; Herzke, D.; Lohmann, R.; Miller, M.; Ng, C.A.; Scheringer, M.; Vierke, L.; et al. Strategies for grouping per- and polyfluoroalkyl substances (PFAS) to protect human and environmental health. Environ. Sci. Process Impacts 2020, 22, 1444–1460. [Google Scholar] [CrossRef]
- Hansen, K.J.; Clemen, L.A.; Ellefson, M.; Johnson, H.O. Compound-specific, quantitative characterization of organic fluorochemicals in biological matrices. Environ. Sci. Technol. 2001, 35, 766–770. [Google Scholar] [CrossRef]
- Remde, A.; Debus, R. Biodegradability of fluorinated surfactants under aerobic and anaerobic conditions. Chemosphere 1996, 32, 1563–1574. [Google Scholar] [CrossRef]
- Schröder, H.F. Determination of fluorinated surfactants and their metabolites in sewage sludge samples by liquid chromatography with mass spectrometry and tandem mass spectrometry after pressurised liquid extraction and separation on fluorine-modified reversed-phase sorbents. J. Chromatogr. A 2003, 1020, 131–151. [Google Scholar] [CrossRef] [PubMed]
- Sáez, M.; de Voogt, P.; Parsons, J.R. Persistence of perfluoroalkylated substances in closed bottle tests with municipal sewage sludge. Environ. Sci. Pollut. Res. Int. 2008, 15, 472–477. [Google Scholar] [CrossRef] [Green Version]
- Frömel, T.; Knepper, T.P. Biodegradation of fluorinated alkyl substances. Rev. Environ. Contam. Toxicol. 2010, 208, 161–177. [Google Scholar] [CrossRef] [PubMed]
- Buck, R.C.; Franklin, J.; Berger, U.; Conder, J.M.; Cousins, I.T.; de Voogt, P.; Jensen, A.A.; Kannan, K.; Mabury, S.A.; van Leeuwen, S.P. Perfluoroalkyl and polyfluoroalkyl substances in the environment: Terminology, classification, and origins. Integr. Environ. Assess. Manag. 2011, 7, 513–541. [Google Scholar] [CrossRef] [PubMed]
- Lau, C.; Anitole, K.; Hodes, C.; Lai, D.; Pfahles-Hutchens, A.; Seed, J. Perfluoroalkyl acids: A review of monitoring and toxicological findings. Toxicol. Sci. 2007, 99, 366–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; DeWitt, J.C.; Higgins, C.P.; Cousins, I.T. A never-ending story of per- and polyfluoroalkyl substances (PFAS)? Environ. Sci. Technol. 2017, 51, 2508–2518. [Google Scholar] [CrossRef]
- Ghisi, R.; Vamerali, T.; Manzetti, S. Accumulation of perfluorinated alkyl substances (PFAS) in agricultural plants: A review. Environ. Res. 2019, 169, 326–341. [Google Scholar] [CrossRef]
- Bossi, R.; Riget, F.F.; Dietz, R.; Sonne, C.; Fauser, P.; Dam, M.; Vorkamp, K. Preliminary screening of perfluorooctane sulfonate (PFOS) and other fluorochemicals in fish, birds and marine mammals from Greenland and the Faroe Islands. Environ. Pollut. 2005, 136, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Cai, M.; Zhao, Z.; Yin, Z.; Ahrens, L.; Huang, P.; Cai, M.; Yang, H.; He, J.; Sturm, R.; Ebinghaus, R. Occurrence of perfluoroalkyl compounds in surface waters from the North Pacific to the Arctic Ocean. Environ. Sci. Technol. 2011, 44, 661–668. [Google Scholar] [CrossRef] [PubMed]
- Holmström, K.E.; Järnberg, U.; Bignert, A. Temporal trends of PFOS and PFOA in guillemot eggs from the Baltic Sea, 1968–2003. Environ. Sci. Technol. 2005, 39, 80–84. [Google Scholar] [CrossRef]
- Paul, A.G.; Jones, K.C.; Sweetman, A.J. A first global production, emission, and environmental inventory for perfluorooctane sulfonate. Environ. Sci. Technol. 2008, 43, 386–392. [Google Scholar] [CrossRef]
- Zhao, Y.; Wong, C.; Wong, M. Environmental contamination, human exposure and body loadings of perfluorooctane sulfonate (PFOS), focusing on Asian countries. Chemosphere 2012, 89, 355–368. [Google Scholar] [CrossRef] [PubMed]
- Bytingsvik, J.; Van Leeuwen, S.P.J.; Hamers, T.; Swart, K.; Aars, J.; Lie, E.; Nilsen, E.M.E.; Wiig, O.; Derocher, A.E.; Jenssen, B.M. Perfluoroalkyl substances in polar bear mother-cub pairs: A comparative study based on plasma levels from 1998 and 2008. Environ. Int. 2012, 49, 92–99. [Google Scholar] [CrossRef]
- Muir, D.; Bossi, R.; Carlsson, P.; Evans, M.; De Silva, A.; Halsall, C.; Rauert, C.; Herzke, D.; Hung, H.; Letcher, R.; et al. Levels and trends of poly- and perfluoroalkyl substances in the Arctic environment—An update. Emerg. Contam. 2019, 5, 240–271. [Google Scholar] [CrossRef]
- Ahrens, L.; Xie, Z.; Ebinghaus, R. Distribution of perfluoroalkyl compounds in seawater from Northern Europe, Atlantic Ocean, and Southern Ocean. Chemosphere 2010, 78, 1011–1016. [Google Scholar] [CrossRef]
- Herzke, D.; Huber, S.; Bervoets, L.; D’Hollander, W.; Hajslova, J.; Pulkrabova, J.; Brambilla, G.; De Filippis, S.P.; Klenow, S.; Heinemeyer, G. Perfluorinated alkylated substances in vegetables collected in four European countries; occurrence and human exposure estimations. Environ. Sci. Pollut. Res. 2013, 20, 7930–7939. [Google Scholar] [CrossRef]
- Jin, Y.H.; Liu, W.; Sato, I.; Nakayama, S.F.; Sasaki, K.; Saito, N.; Tsuda, S. PFOS and PFOA in environmental and tap water in China. Chemosphere 2009, 77, 605–611. [Google Scholar] [CrossRef]
- Kowalczyk, J.; Ehlers, S.; Oberhausen, A.; Tischer, M.; Fürst, P.; Schafft, H.; Lahrssen-Wiederholt, M. Absorption, distribution, and milk secretion of the perfluoroalkyl acids PFBS, PFHxS, PFOS, and PFOA by dairy cows fed naturally contaminated feed. J. Agric. Food Chem. 2013, 61, 2903–2912. [Google Scholar] [CrossRef] [PubMed]
- Loos, R.; Gawlik, B.M.; Locoro, G.; Rimaviciute, E.; Contini, S.; Bidoglio, G. EU wide survey of polar organic persistent pollutants in European river waters. Environ. Pollut. 2009, 157, 561–568. [Google Scholar] [CrossRef] [PubMed]
- So, M.; Taniyasu, S.; Yamashita, N.; Giesy, J.; Zheng, J.; Fang, Z.; Im, S.; Lam, P.K. Perfluorinated compounds in coastal waters of Hong Kong, South China, and Korea. Environ. Sci. Technol. 2004, 38, 4056–4063. [Google Scholar] [CrossRef]
- So, M.; Miyake, Y.; Yeung, W.; Ho, Y.; Taniyasu, S.; Rostkowski, P.; Yamashita, N.; Zhou, B.; Shi, X.; Wang, J. Perfluorinated compounds in the Pearl river and Yangtze river of China. Chemosphere 2007, 68, 2085–2095. [Google Scholar] [CrossRef]
- Washington, J.W.; Yoo, H.; Ellington, J.J.; Jenkins, T.M.; Libelo, E.L. Concentrations, distribution, and persistence of perfluoroalkylates in sludge-applied soils near Decatur, Alabama, USA. Environ. Sci. Technol. 2010, 44, 8390–8396. [Google Scholar] [CrossRef] [PubMed]
- Arinaitwe, K.; Koch, A.; Taabu-Munyaho, A.; Marien, K.; Reemtsma, T.; Berger, U. Spatial profiles of perfluoroalkyl substances and mercury in fish from northern Lake Victoria, East Africa. Chemosphere 2020, 260, 127536. [Google Scholar] [CrossRef]
- Banzhaf, S.; Filipovic, M.; Lewis, J.; Sparrenbom, C.J.; Barthel, R. A review of contamination of surface-, ground-, and drinking water in Sweden by perfluoroalkyl and polyfluoroalkyl substances (PFAS). Ambio 2017, 46, 335–346. [Google Scholar] [CrossRef] [Green Version]
- Prevedouros, K.; Cousins, I.T.; Buck, R.C.; Korzeniowski, S.H. Sources, fate and transport of perfluorocarboxylates. Environ. Sci. Technol. 2006, 40, 32–44. [Google Scholar] [CrossRef]
- Wang, Z.; Cousins, I.T.; Scheringer, M.; Buck, R.C.; Hungerbu¨hler, K. Global emission inventories for C4-C14 perfluoroalkyl carboxylic acid (PFCA) homologues from 1951 to 2030, part I: Production and emissions from quantifiable sources. Environ. Int. 2014, 70, 62–75. [Google Scholar] [CrossRef]
- Gredelj, A.; Nicoletto, C.; Valsecchi, S.; Ferrario, C.; Polesello, S.; Lava, R.; Zanon, F.; Barausse, A.; Palmeri, L.; Guidolin, L.; et al. Uptake and translocation of perfluoroalkyl acids (PFAA) in red chicory (Cichorium intybus L.) under various treatments with pre-contaminated soil and irrigation water. Sci. Total Environ. 2020, 708, 134766. [Google Scholar] [CrossRef]
- Borg, D.; Lund, B.O.; Lindquist, N.G.; Håkansson, H. Cumulative health risk assessment of 17 perfluoroalkylated and polyfluoroalkylated substances (PFAS) in the Swedish population. Environ. Int. 2013, 59, 112–123. [Google Scholar] [CrossRef]
- Ahrens, L. Polyfluoroalkyl compounds in the aquatic environment: A review of their occurrence and fate. J. Environ. Monit. 2011, 13, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Houde, M.; De Silva, A.O.; Muir, D.C.; Letcher, R.J. Monitoring of perfluorinated compounds in aquatic biota: An updated review. Environ. Sci. Technol. 2011, 45, 7962–7973. [Google Scholar] [CrossRef]
- Kannan, K.; Koistinen, J.; Beckmen, K.; Evans, T.; Gorzelany, J.F.; Hansen, K.J.; Jones, P.D.; Helle, E.; Nyman, M.; Giesy, J.P. Accumulation of perfluorooctane sulfonate in marine mammals. Environ. Sci. Technol. 2011, 35, 1593–1598. [Google Scholar] [CrossRef]
- Kannan, K.; Tao, L.; Sinclair, E.; Pastva, S.D.; Jude, D.J.; Giesy, J.P. Perfluorinated compounds in aquatic organisms at various trophic levels in a Great Lakes food chain. Arch. Environ. Contam. Toxicol. 2005, 48, 559–566. [Google Scholar] [CrossRef]
- Yamashita, N.; Kannan, K.; Taniyasu, S.; Horii, Y.; Petrick, G.; Gamo, T. A global survey of perfluorinated acids in oceans. Mar. Pollut. Bull. 2005, 51, 658–668. [Google Scholar] [CrossRef]
- Fujii, S.; Polprasert, C.; Tanaka, S.; Lien, N.P.; Qui, Y. New POPs in the water environment: Distribution, bioaccumulation and treatment of perfluorinated compounds: A review paper. J. Water Suppl. Res. Technol. AQUA 2007, 56, 313–326. [Google Scholar] [CrossRef]
- Arvaniti, O.S.; Stasinakis, A.S. Review on the occurrence, fate and removal of perfluorinated compounds during wastewater treatment. Sci. Total Environ. 2015, 524–525, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Filipovic, M.; Berger, U. Are perfluoroalkyl acids in waste water treatment plant effluents the result of primary emissions from the technosphere or of environmental recirculation? Chemosphere 2015, 129, 74–80. [Google Scholar] [CrossRef]
- Upham, B.L.; Park, J.S.; Babica, P.; Sovadinova, I.; Rummel, A.M.; Trosko, J.E.; Hirose, A.; Hasegawa, R.; Kanno, J.; Sai, K. Structure-activity-dependent regulation of cell communication by perfluorinated fatty acids using in vivo and in vitro model systems. Environ. Health Perspect. 2009, 117, 545–551. [Google Scholar] [CrossRef] [Green Version]
- Klaunig, J.E.; Shinohara, M.; Iwai, H.; Chengelis, C.P.; Kirkpatrick, J.B.; Wang, Z.; Bruner, R.H. Evaluation of the chronic toxicity and carcinogenicity of perfluorohexanoic acid (PFHxA) in Sprague-Dawley rats. Toxicol. Pathol. 2014, 43, 209–220. [Google Scholar] [CrossRef] [PubMed]
- Rosen, M.B.; Das, K.P.; Wood, C.R.; Wolf, C.J.; Abbott, B.D.; Lau, C. Evaluation of perfluoroalkyl acid activity using primary mouse and human hepatocytes. Toxicology 2013, 308, 129–137. [Google Scholar] [CrossRef]
- Vongphachan, V.; Cassone, C.G.; Wu, D.; Chiu, S.; Crump, D.; Kennedy, S.W. Effects of perfluoroalkyl compounds (PFAS) on mRNA expression levels of thyroid hormone-responsive genes in primary cultures of avian neuronal cells. Toxicol. Sci. 2011, 120, 392–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolf, C.J.; Schmid, J.E.; Lau, C.; Abbott, B.D. Activation of mouse and human peroxisome proliferator-activated receptor-alpha (PPARα) by perfluoroalkyl acids (PFAAs): Further investigation of C4-C12 compounds. Reprod. Toxicol. 2012, 33, 546–551. [Google Scholar] [CrossRef]
- Abbott, B.D.; Wolf, C.J.; Schmid, J.E.; Das, K.P.; Zehr, R.D.; Helfant, L.; Nakayama, S.; Lindstrom, A.B.; Strynar, M.J.; Lau, C. Perfluorooctanoic acid-induced developmental toxicity in the mouse is dependent on expression of peroxisome proliferator- activated receptor-alpha. Toxicol. Sci. 2007, 98, 571–581. [Google Scholar] [CrossRef] [Green Version]
- Kudo, N.; Kawashima, Y. Toxicity and toxicokinetics of perfluorooctanoic acid in humans and animals. J. Toxicol. Sci. 2003, 28, 49–57. [Google Scholar] [CrossRef] [Green Version]
- Wolf, C.J.; Fenton, S.E.; Schmid, J.E.; Calafat, A.M.; Kuklenyik, Z.; Bryant, X.A.; Thibodeaux, J.; Das, K.P.; White, S.S.; Lau, C.S. Developmental toxicity of perfluorooctanoic acid in the CD-1 mouse after cross-foster and restricted gestational exposures. Toxicol. Sci. 2007, 95, 462–473. [Google Scholar] [CrossRef] [Green Version]
- Henderson, W.M.; Smith, M.A. Perfluorooctanoic acid and perfluorononanoic acid in fetal and neonatal mice following in utero exposure to 8-2 fluorotelomer alcohol. Toxicol. Sci. 2007, 95, 452–461. [Google Scholar] [CrossRef] [Green Version]
- Fang, X.; Zhang, L.; Feng, Y.; Zhao, Y.; Dai, J. Immunotoxic effects of perfluorononanoic acid on BALB/c mice. Toxicol. Sci. 2008, 105, 312–321. [Google Scholar] [CrossRef] [PubMed]
- Wolf, C.J.; Zehr, R.D.; Schmid, J.E.; Lau, C.; Abbott, B.D. Developmental effects of perfluorononanoic acid in the mouse are dependent on peroxisome proliferator-activated receptor-alpha. PPAR Res. 2010, 2010, 282896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, M.W.; Birnbaum, L.S. Developmental toxicity of perfluorodecanoic acid in C57BL/6N mice. Toxicol. Sci. 1989, 12, 442–448. [Google Scholar] [CrossRef]
- Heuvel, J.P.V.; Kuslikis, B.I.; Peterson, R.E. Covalent binding of perfluorinated fatty acids to proteins in the plasma, liver and testes of rats. Chem. Biol. Interact. 1992, 82, 317–328. [Google Scholar] [CrossRef]
- Takahashi, M.; Ishida, S.; Hirata-Koizumi, M.; Ono, A.; Hirose, A. Repeated dose and reproductive/developmental toxicity of perfluoroundecanoic acid in rats. J. Toxicol. Sci. 2014, 39, 97–108. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Zhang, H.; Cui, R.; Guo, X.; Wang, D.; Dai, J. Activation of peroxisome proliferator-activated receptor α ameliorates perfluorododecanoic acid-induced production of reactive oxygen species in rat liver. Arch. Toxicol. 2016, 90, 1383–1397. [Google Scholar] [CrossRef]
- Ren, X.M.; Zhang, Y.F.; Guo, L.H.; Qin, Z.F.; Lv, Q.Y.; Zhang, L.Y. Structure-activity relations in binding of perfluoroalkyl compounds to human thyroid hormone T3 receptor. Arch. Toxicol. 2015, 89, 233–242. [Google Scholar] [CrossRef]
- Zhang, H.; Hou, J.; Cui, R.; Guo, X.; Shi, Z.; Yang, F.; Dai, J. Phosphoproteome analysis reveals an important role for glycogen synthase kinase-3 in perfluorododecanoic acid-induced rat liver toxicity. Toxicol. Lett. 2013, 218, 61–69. [Google Scholar] [CrossRef]
- Hu, W.Y.; Jones, P.D.; DeCoen, W.; King, L.; Fraker, P.; Newsted, J.; Giesy, J.P. Alterations in cell membrane properties caused by perfluorinated compounds. Comp. Biochem. Physiol. C 2003, 135, 77–88. [Google Scholar] [CrossRef]
- Ankley, G.T.; Kuehl, D.W.; Kahl, M.D.; Jensen, K.M.; Linnum, A.; Leino, R.L.; Villeneuve, D.A. Reproductive and developmental toxicity and bioconcentration of perfluorooctanesulfonate in a partial life-cycle test with the fathead minnow (Pimephales promelas). Environ. Toxicol. Chem. 2005, 24, 2316–2324. [Google Scholar] [CrossRef]
- Seacat, A.M.; Thomford, P.J.; Hansen, K.J.; Olsen, G.W.; Case, M.T.; Butenhoff, J.L. Subchronic toxicity studies on perfluorooctanesulfonate potassium salt in cynomolgus monkeys. Toxicol. Sci. 2002, 68, 249–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seacat, A.M.; Thomford, P.J.; Hansen, K.J.; Clemen, L.A.; Eldridge, S.R.; Elcombe, C.R.; Butenhoff, J.L. Sub-chronic dietary toxicity of potassium perfluorooctanesulfonate in rats. Toxicology 2003, 183, 117–131. [Google Scholar] [CrossRef]
- Liu, Z.; Lu, Y.; Shi, Y.; Wang, P.; Jones, K.; Sweetman, A.J.; Johnson, A.C.; Zhang, M.; Zhou, Y.; Lu, X.; et al. Crop bioaccumulation and human exposure of perfluoroalkyl acids through multimedia transport from a mega fluorochemical industrial park, China. Environ. Int. 2017, 106, 37–47. [Google Scholar] [CrossRef] [Green Version]
- Guelfo, J.L.; Higgins, C.P. Subsurface transport potential of perfluoroalkyl acids at aqueous film-forming foam (AFFF)-impacted sites. Environ. Sci. Technol. 2013, 47, 4164–4171. [Google Scholar] [CrossRef]
- Blaine, A.C.; Rich, C.D.; Hundal, L.S.; Lau, C.; Mills, M.A.; Harris, K.M.; Higgins, C.P. Uptake of perfluoroalkyl acids into edible crops via land applied biosolids: Field and greenhouse studies. Environ. Sci. Technol. 2013, 47, 14062–14069. [Google Scholar] [CrossRef]
- Blaine, A.C.; Rich, C.D.; Sedlacko, E.M.; Hyland, K.C.; Stushnoff, C.; Dickenson, E.R.V.; Higgins, C.P. Perfluoroalkyl acid uptake in lettuce (Lactuca sativa) and strawberry (Fragaria ananassa) irrigated with reclaimed water. Environ. Sci. Technol. 2014, 48, 14361–14368. [Google Scholar] [CrossRef]
- Felizeter, S.; McLachlan, M.S.; De Voogt, P. Uptake of perfluorinated alkyl acids by hydroponically grown lettuce (Lactuca sativa). Environ. Sci. Technol. 2012, 46, 11735–11743. [Google Scholar] [CrossRef] [PubMed]
- McLachlan, M.S.; Felizeter, S.; Klein, M.; Kotthoff, M.; De Voogt, P. Fate of a perfluoroalkyl acid mixture in an agricultural soil studied in lysimeters. Chemosphere 2019, 223, 180–187. [Google Scholar] [CrossRef]
- Vestergren, R.; Ullah, S.; Cousins, I.T.; Berger, U. A matrix effect-free method for reliable quantification of perfluoroalkyl carboxylic acids and perfluoroalkane sulfonic acids at low parts per trillion levels in dietary samples. J. Chromatogr. A 2012, 1237, 64–71. [Google Scholar] [CrossRef]
- European Food Safety Authority. Perfluoroalkylated substances in food: Occurrence and dietary exposure. EFSA J. 2012, 10, 2743. [Google Scholar] [CrossRef]
- Vestergren, R.; Cousins, I.T. Tracking the pathways of human exposure to perfluorocarboxylates. Environ. Sci. Technol. 2009, 43, 5565–5575. [Google Scholar] [CrossRef]
- Calafat, A.M.; Kuklenyik, Z.; Reidy, J.A.; Caudill, S.P.; Tully, J.S.; Needham, L.L. Serum concentrations of 11 polyfluoroalkyl compounds in the U.S. population: Data from the National Health and Nutrition Examination Survey (NHANES) 1999−2000. Environ. Sci. Technol. 2007, 41, 2237–2242. [Google Scholar] [CrossRef] [PubMed]
- Biegel, L.B.; Hurtt, M.E.; Frame, S.R.; O’Connor, J.C.; Cook, J.C. Mechanisms of extrahepatic tumor induction by peroxisome proliferators in male CD rats. Toxicol. Sci. 2001, 60, 44–55. [Google Scholar] [CrossRef]
- U.S. Environmental Protection Agency. SAB Review of EPA’s Draft Risk Assessment of Potential Human Health Effects Associated with PFOA and Its Salts. EPA-SAB-06-006. Available online: http://yosemite.epa.gov/sab/SABPRODUCT.NSF/acdfe15ab1de42e485256ead006ff442/A3C83648E77252828525717F004B9099/$File/sab_06_006.pdf (accessed on 2 October 2020).
- Lundin, J.I.; Alexander, B.H.; Olsen, G.W.; Church, T.R. Ammonium perfluorooctanoate production and occupational mortality. Epidemiology 2009, 20, 921–928. [Google Scholar] [CrossRef] [PubMed]
- Barry, V.; Winquist, A.; Steenland, K. Per Uorooctanoic Acid (PFOA) exposures and incident cancers among adults living near a chemical plant. Environ. Health Perspect. 2013, 121, 1313–1318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leonard, R.C.; Kreckmann, K.H.; Sakr, C.J.; Symons, J.M. Retrospective cohort mortality study of workers in a polymer production plant including a reference population of regional workers. Ann. Epidemiol. 2008, 18, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Steenland, K.; Woskie, S. Cohort mortality study of workers exposed to perfluorooctanoic acid. Am. J. Epidemiol. 2012, 176, 909–917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frisbee, S.J.; Brooks, A.P., Jr.; Maher, A.; Flensborg, P.; Arnold, S.; Fletcher, T.; Steenland, K.; Shankar, A.; Knox, S.S.; Pollard, C.; et al. The C8 health project: Design, methods, and participants. Environ. Health Perspect. 2009, 117, 1873–1882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pistocchi, A.; Loos, R. A map of European emissions and concentrations of PFOS and PFOA. Environ. Sci. Technol. 2009, 43, 9237–9244. [Google Scholar] [CrossRef]
- Atkinson, C.; Blake, S.; Hall, T.; Kanda, R.; Rumsby, P. Survey of the prevalence of perfluorooctane sulphonate (PFOS), perfluorooctanoic acid (PFOA) and related compounds in drinking water and their sources. Report DEFRA 7585, Drinking Water Inspectorate, Department for Environment, Food and Rural Affairs. Available online: http://www.fwr.org/analysis/dwi0854.htm (accessed on 2 October 2020).
- Lange, F.T.; Wenz, M.; Schmidt, C.K.; Brauch, H.J. Occurrence of perfluoroalkyl sulfonates and carboxylates in German drinking water sources compared to other countries. Water Sci. Technol. 2007, 56, 151–158. [Google Scholar] [CrossRef]
- Gyllenhammar, I.; Berger, U.; Sundstro¨m, M.; McCleaf, P.; Eure’n, K.; Eriksson, S.; Ahlgren, S.; Lignell, S. Influence of contaminated drinking water on perfluoroalkyl acid levels in human serum—A case study from Uppsala, Sweden. Environ. Res. 2015, 140, 673–683. [Google Scholar] [CrossRef]
- Pitter, G.; DaRe, F.; Canova, C.; Barbieri, G.; ZareJeddi, M.; Daprà, F.; Manea, M.; Zolin, R.; Bettega, A.M.; Stopazzolo, G.; et al. Serum levels of perfluoroalkyl substances (PFAS) in adolescents and young adults exposed to contaminated drinking water in the Veneto Region, Italy: A cross-sectional study based on a health surveillance program. Environ. Health Perspect. 2020, 128, 27007. [Google Scholar] [CrossRef] [Green Version]
- Valsecchi, S.; Babut, M.; Mazzoni, M.; Pascariello, S.; Ferrario, C.; De Felice, B.; Bettinetti, R.; Veyrand, B.; Marchand, P.; Polesello, S. Perfluoroalkyl Substances (PFAS) in Fish from European Lakes: Current contamination status, sources, and perspectives for monitoring. Environ. Toxicol. Chem. 2020. [Google Scholar] [CrossRef] [PubMed]
- Brooke, D.; Footitt, A.; Nwaogu, T.A. Environmental Risk Evaluation Report: Perfluorooctanesulphonate (PFOS). United Kingdom: Environment Agency. 2004. Available online: https://assets.publishing.service.gov.u/kgovernment/uploads/system/uploads/attachment_data/file/290857/scho1009brbl-e-e.pdf (accessed on 2 October 2020).
- OECD Report ENV/JM/RD. 2002. Available online: https://www.oecd.org/env/ehs/risk-assessment/2382880.pdf (accessed on 2 October 2020).
- OECD. Results of the 2006 Survey on Production and Use of PFOS, PFAS, PFOA, PFCA, Their Related Substances and Products/Mixtures Containing These Substances; ENV/JM/MONO; OECD: Paris, France, 2006; Volume 36, p. 6. [Google Scholar]
- Stockholm Convention on POPs. POPRC Recommendations for Listing Chemicals. Available online: http://chm.pops.int/Convention/POPs-ReviewCommittee/Chemicals/tabid/243/language/en-US/Default.aspx (accessed on 2 October 2020).
- European Commission. Directive 2006/122/EC of the European Parliament and of the Council of 12 December 2006 amending for the 30th time Council Directive 76/769/EEC on the approximation of the laws, regulations and administrative provisions of the member states relating to restrictions on the marketing and use of certain dangerous substances and preparations (perfluorooctane sulfonates). Off. J. Eur. Commun. 2006, 58, 32–34. Available online: https://eur-lex.europa.eu/LexUriServ/LexUriServ.do?uri=OJ:L:2006:372:0032:0034:EN:PDF (accessed on 2 October 2020).
- Blum, A.; Balan, S.A.; Scheringer, M.; Goldenman, G.; Trier, X.; Cousins, I.T.; Diamond, M.; Higgins, C.; Avery, E.; Peaslee, G.; et al. The Madrid Statement on poly- and perfluoroalkyl substances (PFAS). Environ. Health Perspect. 2015, 123, A107–A111. [Google Scholar] [CrossRef] [PubMed]
- United Nations Environment Programme, Stockholm Convention. Decision SC-9/12. Listing of perfluorooctanoic acid (PFOA), its salts and PFOA related compounds. In Proceedings of the Ninth Meeting of the Conference of the Parties to the Stockholm Convention, Geneva, Switzerland, 29 April–10 May 2019; Available online: http://chm.pops.int/TheConvention/ConferenceoftheParties/Meetings/COP9/tabid/7521/Default.aspx (accessed on 2 October 2020).
- United Nations Environment Programme, Stockholm Convention. Working Document UNEP/POPS/POPRC.13/4. Proposal to list perfluorohexane sulfonic acid (CAS No: 355-46-4, PFHxS), its salts and PFHxS-related compounds in Annexes A, B and/or C to the Stockholm Convention on Persistent Organic Pollutants. In Proceedings of the Thirteenth meeting of the Persistent Organic Pollutants Review Committee (POPRC.13), Rome, Italy, 17–20 October 2017; Available online: https://assets.publishing.service.gov.uk/government/uploads/system/uploads/attachment_data/file/338258/PFOS___PFOA_General_Information_phe_v1.pdf (accessed on 2 October 2020).
- Public Health England, Toxicology Department. PFOS and PFOA General Information. 2009. Available online: http://www.pops.int/Default.aspx?tabid=5965 (accessed on 2 October 2020).
- Geueke, B. FPF Dossier: Per- and polyfluoroalkyl substances (PFAS). Zenodo 2016. [Google Scholar] [CrossRef]
- Trinkwasserkommission (German Drinking Water Commission of the Ministry of Health). Provisional Evaluation of PFT in Drinking Water with the Guide Substances Perfluorooctanoic Acid (PFOA) and Perfluorooctane Sulfonate (PFOS) as Examples. 2006. Available online: https://www.umweltbundesamt.de/sites/default/files/medien/pdfs/pft-in-drinking-water.pdf (accessed on 2 October 2020).
- Veneto Region. Progress and Prospects for Veneto Agriculture. 2014. Available online: http://statistica.regione.veneto.it/ENG/Pubblicazioni/RapportoStatistico2014/pdf/Capitolo07.pdf (accessed on 2 October 2020).
- Nicoletto, C.; Maucieri, C.; Sambo, P. Effects on water management and quality characteristics of ozone application in chicory forcing process: A pilot system. Agronomy 2017, 7, 29. [Google Scholar] [CrossRef]
- WHO. Keeping Our Water Clean: The Case of Water Contamination in the Veneto Region, Italy. World Health Organization. 2017. Available online: https://www.euro.who.int/__data/assets/pdf_file/0019/341074/pfas-report-20170606-h1330-print-isbn.pdf (accessed on 2 October 2020).
- ARPAV. Concentrations of the Perfluoroalkyl Substances in the Waters of Veneto Region, Open Data on PFAS Monitoring, from 02/07/2013 to 20/09/2018. 2018. Available online: http://www.arpa.veneto.it/dati-ambientali/open-dataidrosfera/concentrazione-di-sostanze-perfluoroalchiliche-pfas-nelle-acqueprelevate-da-arpav (accessed on 2 October 2020).
- Mastrantonio, M.; Bai, E.; Uccelli, R.; Cordiano, V.; Screpanti, A.; Crosignani, P. Drinking water contamination from perfluoroalkyl substances (PFAS): An ecological mortality study in the Veneto Region, Italy. Eur. J. Public Health 2018, 28, 180–185. [Google Scholar] [CrossRef]
- Darrow, L.A.; Stein, C.R.; Steenland, K. Serum perfluorooctanoic acid and per- fluoro octane sulfonate concentrations in relation to birth outcomes in the Mid- Ohio Valley, 2005–2010. Environ. Health Perspect. 2013, 121, 1207–1213. [Google Scholar] [CrossRef] [Green Version]
- Webster, E.; Ellis, D.A.; Reid, L.K. Modeling the environmental fate of perfluorooctanoic acid and perfluorooctanoate: An investigation of the role of individual species partitioning. Environ. Toxicol. Chem. 2010, 29, 1466–1475. [Google Scholar] [CrossRef]
- Giesy, J.P.; Kannan, K. Global distribution of perfluorooctane sulfonate in wildlife. Environ. Sci. Technol. 2001, 35, 1339–1342. [Google Scholar] [CrossRef]
- Grandjean, P. Health status of workers exposed to perfluorinated alkylate substances. J. Occup. Environ. Med. 2018, 60, e562. [Google Scholar] [CrossRef]
- Taves, D.R. Evidence that there are two forms of fluoride in human serum. Nature 1968, 217, 1050. [Google Scholar] [CrossRef]
- Taves, D.R. Comparison of “organic” fluoride in human and nonhuman serums. J. Dent. Res. 1971, 50, 783. [Google Scholar] [CrossRef]
- Shen, Y.W.; Taves, D.R. Fluoride concentrations in the human placenta and maternal and cord blood. An. J. Obstet. Gynecol. 1974, 119, 205–207. [Google Scholar] [CrossRef]
- Guy, W.S.; Taves, D.R.; Brey, W.S., Jr. Organic Fluorocompounds in Human Plasma: Prevalence and Characterization. In Biochemistry Involving Carbon-Fluorine Bonds; Filler, R., Ed.; ACS Publications: Washington, DC, USA, 1979; Volume 28, pp. 117–134. [Google Scholar] [CrossRef]
- Ubel, F.A.; Sorenson, S.; Roach, D. Health status of plant workers exposed to fluorochemicals-a preliminary report. Am. Ind. Hyg. Assoc. J. 1980, 41, 584–589. [Google Scholar] [CrossRef] [PubMed]
- Gilliland, F.D. Fluorocarbons and Human Health: Studies in an Occupational Cohort. Ph.D. Thesis, University of Minnesota, Minneapolis, MN, USA, 1992. [Google Scholar]
- Gilliland, F.D.; Mandel, J.S. Peripheral Blood Lymphocyte Count in Men Occupationally Exposed to Perfluorooctanoic Acid. Personal Communication. 1993. Available online: https://www.ag.state.mn.us/Office/Cases/3M/docs/PTX/PTX2498.pdf (accessed on 2 October 2020).
- Gilliland, F.D.; Mandel, J.S. Mortality among employees of a perfluorooctanoic acid production plant. J. Occup. Med. 1993, 35, 950–954. [Google Scholar] [CrossRef] [PubMed]
- Gilliland, F.D.; Mandel, J.S. Serum perfluorooctanoic acid and hepatic enzymes, lipoproteins and cholesterol: A study of occupationally exposed men. Am. J. Ind. Med. 1996, 29, 560–568. [Google Scholar] [CrossRef]
- 3M. Perfluorooctane Sulfonate: Current Summary of Human Sera, Health and Toxicological Data; U.S. EPA Administrative Record AR226-0548; 3M: Saint Paul, MN, USA, 1999. [Google Scholar]
- Calafat, A.M.; Wong, L.Y.; Kuklenyik, Z.; Reidy, J.A.; Needham, L.L. Polyfluoroalkyl chemicals in the U.S. population: Data from the National Health and Nutrition Examination Survey (NHANES) 2003–2004 and comparisons with NHANES 1999–2000. Environ. Health Perspect. 2007, 115, 1596–1602. [Google Scholar] [CrossRef]
- Andrews, D.; Walker, B. Poisoned Legacy. Know Your Environment: Protect Your Health. 2015. Available online: https://www.ewg.org/research/poisoned-legacy (accessed on 2 October 2020).
- Scheringer, M.; Trier, X.; Cousins, I.T.; de Voogt, P.; Fletcher, T.; Wang, Z.; Webster, T.F. Helsingør statement on poly- and perfluorinated alkyl substances (PFAS). Chemosphere 2014, 114, 337–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ritscher, A.; Wang, Z.; Scheringer, M.; Boucher, J.M.; Ahrens, L.; Berger, U.; Bintein, S.; Bopp, S.K.; Borg, D.; Buser, A.M.; et al. Zürich statement on future actions on per- and polyfluoroalkyl substances (PFAS). Environ. Health Perspect. 2018, 126, 84502. [Google Scholar] [CrossRef]
- CDC. Center for Disease Control and Prevention: National Health and Nutrition Examination Survey. 2009–2010 Data Documentation, Codebook, and Frequencies. 2013, Polyfluoroalkyl Chemicals (PFC_F). Available online: http://wwwn.cdc.gov/nchs/nhanes/2009-2010/PFC_F.htm (accessed on 2 October 2020).
- Kato, K.; Wong, L.Y.; Jia, L.T.; Kuklenyik, Z.; Calafat, A.M. Trends in exposure to polyfluoroalkyl chemicals in the U.S. population: 1999–2008. Environ. Sci. Technol. 2011, 45, 8037–8045. [Google Scholar] [CrossRef]
- Axmon, A.; Axelsson, J.; Jakobsson, K.; Lindh, C.H.; Jönsson, B.A. Time trends between 1987 and 2007 for perfluoroalkyl acids in plasma from Swedish women. Chemosphere 2014, 102, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Glynn, A.; Berger, U.; Bignert, A.; Ullah, S.; Aune, M.; Lignell, S.; Darnerud, P.O. Perfluorinated alkyl acids in blood serum from primiparous women in Sweden: Serial sampling during pregnancy and nursing, and temporal trends 1996–2010. Environ. Sci. Technol. 2012, 46, 9071–9079. [Google Scholar] [CrossRef]
- Gebbink, W.A.; Berger, U.; Cousins, I.T. Estimating human exposure to PFOS isomers and PFCA homologues: The relative importance of direct and indirect (precursor) exposure. Environ. Int. 2015, 74, 160–169. [Google Scholar] [CrossRef] [Green Version]
- Nøst, T.H.; Vestergren, R.; Berg, V.; Nieboer, E.; Odland, J.Ø.; Sandanger, T.M. Repeated measurements of per- and polyfluoroalkyl substances (PFAS) from 1979 to 2007 in males from Northern Norway: Assessing time trends, compound correlations and relations to age/birth cohort. Environ Int. 2014, 67, 43–53. [Google Scholar] [CrossRef] [Green Version]
- Bjerregaard-Olsen, C.; Bach, C.C.; Long, M.; Ghisari, M.; Bossi, R.; Bech, B.H.; Nohr, E.A.; Henriksen, T.B.; Olsen, J.; Bonefeld-Jørgensen, E.C. Time trends of perfluorinated alkyl acids in serum from Danish pregnant women 2008–2013. Environ. Int. 2016, 91, 14–21. [Google Scholar] [CrossRef]
- Schröter-Kermani, C.; Müller, J.; Jürling, H.; Conrad, A.; Schulte, C. Retrospective monitoring of perfluorocarboxylates and perfluorosulfonates in human plasma archived by the German Environmental Specimen Bank. Int. J. Hyg. Environ. Health 2016, 6, 633–640. [Google Scholar] [CrossRef]
- Yeung, L.W.Y.; Robinson, S.J.; Koschorreck, J.; Mabury, S.A. Part I. A temporal study of PFCAs and their precursors in human plasma from two German cities 1982–2009. Environ. Sci. Technol. 2013, 47, 3865–3874. [Google Scholar] [CrossRef]
- Yeung, L.W.Y.; Robinson, S.J.; Koschorreck, J.; Mabury, S.A. Part II. A temporal study of PFOS and its precursors in human plasma from two German cities in 1982–2009. Environ. Sci. Technol. 2013, 47, 3875–3882. [Google Scholar] [CrossRef]
- Toms, L.M.L.; Thompson, J.; Rotander, A.; Hobson, P.; Calafat, A.M.; Kato, K.; Ye, X.; Broomhall, S.; Harden, F.; Mueller, J.F. Decline in perfluorooctane sulfonate and perfluorooctanoate serum concentrations in an Australian population from 2002 to 2011. Environ. Int. 2014, 71, 74–80. [Google Scholar] [CrossRef] [Green Version]
- Hölzer, J.; Midasch, O.; Rauchfuss, K.; Kraft, M.; Reupert, R.; Angerer, J.; Kleeschulte, P.; Marschall, N.; Wilhelm, M. Biomonitoring of perfluorinated compounds in children and adults exposed to perfluorooctanoate-contaminated drinking water. Environ. Health Perspect. 2008, 116, 651–657. [Google Scholar] [CrossRef]
- Okada, E.; Kashino, I.; Matsuura, H.; Sasaki, S.; Miyashita, C.; Yamamoto, J.; Ikeno, T.; Ito, Y.M.; Matsumura, T.; Tamakoshi, A.; et al. Temporal trends of perfluoroalkyl acids in plasma samples of pregnant women in Hokkaido, Japan, 2003-2011. Environ. Int. 2013, 60, 89–96. [Google Scholar] [CrossRef]
- Harada, K.H.; Yang, H.R.; Moon, C.S.; Hung, N.N.; Hitomi, T.; Inoue, K.; Niisoe, T.; Watanabe, T.; Kamiyama, S.; Takenaka, K.; et al. Levels of perfluorooctane sulfonate and perfluorooctanoic acid in female serum samples from Japan in 2008, Korea in 1994–2008 and Vietnam in 2007–2008. Chemosphere 2010, 79, 314–319. [Google Scholar] [CrossRef]
- Yeung, L.W.Y.; So, M.K.; Jiang, G.B.; Taniyasu, S.; Yamashita, N.; Song, M.Y.; Wu, Y.N.; Li, J.G.; Giesy, J.P.; Guruge, K.S.; et al. Perfluorooctanesulfonate and related fluorochemicals in human blood samples from China. Environ. Sci. Technol. 2006, 40, 715–720. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.Y.; Shi, Y.L.; Wang, J.; Cai, Y.; Wu, Y. Concentrations of perfluorinated compounds in human blood from twelve cities in China. Environ. Toxicol. Chem. 2010, 29, 2695–2701. [Google Scholar] [CrossRef]
- Ingelido, A.M.; Marra, V.; Abballe, A.; Valentini, S.; Iacovella, N.; Barbieri, P.; Porpora, M.G.; Domenico, A.D.; De Felip, E. Perfluorooctanesulfonate and perfluorooctanoic acid exposures of the Italian general population. Chemosphere 2010, 80, 1125–1130. [Google Scholar] [CrossRef]
- Olsen, G.W.; Church, T.R.; Miller, J.P.; Burris, J.M.; Hansen, K.J.; Lundberg, J.K.; Armitage, J.B.; Herron, R.M.; Medhdizadehkashi, Z.; Nobiletti, J.B.; et al. Perfluorooctanesulfonate and other fluorochemicals in the serum of American Red Cross adult blood donors. Environ. Health Perspect. 2003, 111, 1892–1901. [Google Scholar] [CrossRef] [Green Version]
- Toms, L.M.; Calafat, A.; Kato, K.; Thompson, J.; Harden, F.; Hobson, P.; Sjodin, A.; Jochen, A.; Mueller, F. Polyfluoroalkyl Chemicals in pooled blood serum from infants, children and adults in Australia. Environ. Sci. Technol. 2009, 43, 4194–4199. [Google Scholar] [CrossRef] [PubMed]
- Harada, K.; Saito, N.; Inoue, K.; Yoshinaga, T.; Watanabe, T.; Sasaki, S.; Kamiyama, S.; Koizumi, A. The influence of time, sex and geographic factors on levels of perfluorooctane sulfonate and perfluorooctanoate in human serum over the last 25 years. J. Occup. Health 2004, 46, 141–147. [Google Scholar] [CrossRef] [Green Version]
- Midasch, O.; Schettgen, T.; Angerer, J. Pilot study on the perfluorooctanesulfonate and perfluorooctanoate exposure of the German general population. Int. J. Environ. Health 2006, 209, 489–496. [Google Scholar] [CrossRef]
- Wilhelma, M.; Angererb, J.; Frommec, H.; Holzera, J. Contribution to the evaluation of reference values for PFOA and PFOS in plasma of children and adults from Germany. Int. J. Hyg. Environ. Health 2009, 212, 56–60. [Google Scholar] [CrossRef]
- Ericson, I.; Gómez, M.; Nadal, M.; Van Bavel, B.; Lindström, G.; Domingo, J.L. Perfluorinated chemicals in blood of residents in Catalonia (Spain) in relation to age and gender: A pilot study. Environ. Int. 2007, 33, 616–623. [Google Scholar] [CrossRef] [PubMed]
- Kannan, K.; Corsolini, S.; Falandysz, J.; Fillmann, G.; Kumar, K.S.; Loganathan, B.G.; Mustafa, A.M.; Olivero, J.; Van Wouwe, N.; Yang, J.H.; et al. Perfluorooctanesulfonate and related fluorochemicals in human blood from several countries. Environ. Sci. Technol. 2004, 38, 4489–4495. [Google Scholar] [CrossRef] [PubMed]
- Tomy, G.T.; Tittlemier, S.A.; Palace, V.P.; Budakowski, W.R.; Braekevelt, E.; Brinkworth, L.; Friesen, K. Biotransformation of N-ethyl perfluorooctanesulfonamide by rainbow trout (Onchorhynchus mykiss) liver microsomes. Environ. Sci. Technol. 2004, 38, 758–762. [Google Scholar] [CrossRef]
- EFSA. Perfluorooctane sulfonate (PFOS), perfluorooctanoic acid (PFOA) and their salts. Scientific opinion of the panel on contaminants in the food chain. EFSA J. 2008, 653, 1–131. [Google Scholar] [CrossRef] [Green Version]
- Fromme, H.; Midasch, O.; Twardella, D.; Angerer, J.; Boehmer, S.; Liebl, B. Occurrence of perfluorinated substances in an adult German population in southern Bavaria. Int. Arch. Occup. Environ. Health 2007, 80, 313–319. [Google Scholar] [CrossRef]
- De Felip, E.; Abballe, A.; Albano, F.L.; Battista, T.; Carraro, V.; Conversano, M.; Franchini, S.; Giambanco, L.; Iacovella, N.; Ingelido, A.M.; et al. Current exposure of Italian women of reproductive age to PFOS and PFOA: A human biomonitoring study. Chemosphere 2015, 137, 1–8. [Google Scholar] [CrossRef]
- Li, Y.; Fletcher, T.; Mucs, D.; Scott, K.; Lindh, C.H.; Tallving, P.; Jakobsson, K. Half-lives of PFOS, PFHxS and PFOA after end of exposure to contaminated drinking water. Occup. Environ. Med. 2018, 75, 46–51. [Google Scholar] [CrossRef] [Green Version]
- Polesello, S.; Pagnotta, R.; Marziali, L.; Patrolecco, L.; Rusconi, M.; Stefani, F.; Valsecchi, S. Realizzazione di uno Studio di Valutazione del Rischio Ambientale e Sanitario Associato alla Contaminazione da Sostanze Perfluoro-Alchiliche (PFAS) nel Bacino del Po e nei Principali Bacini Fluviali Italiani. 2013, Relazione Finale. Available online: http://www.minambiente.it/sites/default/files/archivio/allegati/reach/progettoPFAS_ottobre2013.pdf (accessed on 2 October 2020).
- Valsecchi, S.; Rusconi, M.; Mazzoni, M.; Viviano, G.; Pagnotta, R.; Zaghi, C.; Serrini, G.; Polesello, S. Occurrence and sources of perfluoroalkyl acids in Italian river basins. Chemosphere 2015, 129, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Ingelido, A.M.; Abballe, A.; Gemma, S.; Dellatte, E.; Iacovella, N.; DeAngelis, G.; Zampaglioni, F.; Marra, V.; Miniero, R.; Valentini, S.; et al. Biomonitoring of perfluorinated compounds in adults exposed to contaminated drinking water in the Veneto Region, Italy. Environ Int. 2018, 110, 149–159. [Google Scholar] [CrossRef]
- U.S. EPA. Drinking Water Health Advisory for Perfluorooctanoic Acid (PFOA). 2016. Available online: https://www.epa.gov/sites/production/files/2016-05/documents/pfoa_health_advisory_final_508.pdf (accessed on 2 October 2020).
- U.S. EPA. Drinking Water Health Advisory for Perfluorooctane Sulfonate (PFOS). 2016. Available online: https://www.epa.gov/sites/production/files/2016-05/documents/pfos_health_advisory_final_508.pdf (accessed on 2 October 2020).
- Danish Environmental Protection Agency. Perfluoroalkylated Substances: PFOA, PFOS and PFOSA Evaluation of Health Hazards and Proposal of a Health Based Quality Criterion for Drinking Water, Soil and Ground Water. Environmental Project No.1665. 2015, p. 2015. Available online: http://www2.mst.dk/Udgiv/publications/2015/04/978-87-93283-01-5.pdf (accessed on 2 October 2020).
- Health Council of the Netherlands. Perfluorooctanoic Acid and Its Salts. Evaluation of the Carcinogenicity and Genotoxicity. 2013. Publication no. 2013/32. Available online: https://www.gezondheidsraad.nl/sites/default/files/201332Perfluorooctanoic_acid_and_its_salts.pdf (accessed on 2 October 2020).
- Girardi, P.; Merler, E. A mortality study on male subjects exposed to polyfluoroalkyl acids with high internal dose of perfluorooctanoic acid. Environ. Res. 2019, 179, 108743. [Google Scholar] [CrossRef]
- Crebelli, R.; Caiola, S.; Conti, L.; Cordelli, E.; De Luca, G.; Dellatte, E.; Eleuteri, P.; Iacovella, N.; Leopardi, P.; Marcon, F.; et al. Can sustained exposure to PFAS trigger a genotoxic response? A comprehensive genotoxicity assessment in mice after subacute oral administration of PFOA and PFBA. Regul. Toxicol. Pharmacol. 2019, 106, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Costa, G.; Sartori, S.; Consonni, D. Thirty years of medical surveillance in perfluorooctanoic acid production workers. J. Occup. Environ. Med. 2009, 51, 364–372. [Google Scholar] [CrossRef] [PubMed]
- Salihovic, S.; Stubleski, J.; Kärrman, A.; Larsson, A.; Fall, T.; Lind, L. Changes in markers of liver function in relation to changes in perfluoroalkyl substances—A longitudinal study. Environ. Int. 2018, 117, 196–203. [Google Scholar] [CrossRef]
- Gallo, V.; Leonardi, G.; Genser, B.; Lopez-Espinosa, M.-J.; Frisbee, S.J.; Karlsson, L.; Ducatman, A.M.; Fletcher, T. Serum perfluorooctanoate (PFOA) and perfluorooctane sulfonate (PFOS) concentrations and liver function biomarkers in a population with elevated PFOA exposure. Environ. Health Perspect. 2012, 120. [Google Scholar] [CrossRef] [Green Version]
- Darrow, L.A.; Groth, A.C.; Winquist, A.; Shin, H.-M.; Bartell, S.M.; Steenland, K. Modeled perfluorooctanoic acid (PFOA) exposure and liver function in a Mid-Ohio valley community. Environ. Health Perspect. 2016, 124. [Google Scholar] [CrossRef] [Green Version]
- Post, G.B.; Cohn, P.D.; Cooper, K.R. Perfluorooctanoic acid (PFOA), an emerging drinking water contaminant: A critical review of recent literature. Environ. Res. 2012, 116, 93–117. [Google Scholar] [CrossRef] [PubMed]
- IARC Working Group on the Evaluation of Carcinogenic Risks to Humans. Polychlorinated Biphenyls and Polybrominated Biphenyls; IARC Monographs on the Evaluation of Carcinogenic Risks to Humans, No. 107; International Agency for Research on Cancer: Lyon, France, 2016. [Google Scholar]
- Abdellatif, A.; Al-Tonsy, A.H.; Awad, M.E.; Roberfroid, M.; Khan, M.N. Peroxisomal enzymes and 8-hydroxydeoxyguanosine in rat liver treated with perfluorooctanoic acid. Dis. Markers 2003, 19, 19–25. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Yu, K.; Shi, X.; Wang, J.; Lam, P.K.; Wu, R.S.; Zhou, B. Induction of oxidative stress and apoptosis by PFOS and PFOA in primary cultured hepatocytes of freshwater tilapia (Oreochromis niloticus). Aquat. Toxicol. 2007, 82, 135–143. [Google Scholar] [CrossRef]
- Panaretakis, T.; Shabalina, I.G.; Grandér, D.; Shoshan, M.C.; DePierre, J.W. Reactive oxygen species and mitochondria mediate the induction of apoptosis in human hepatoma HepG2 cells by the rodent peroxisome proliferator and hepatocarcinogen, perfluorooctanoic acid. Toxicol. Appl. Pharmacol. 2001, 173, 56–64. [Google Scholar] [CrossRef]
- Yao, X.; Zhong, L. Genotoxic risk and oxidative DNA damage in HepG2 cells exposed to perfluorooctanoic acid. Mutat. Res. 2005, 587, 38–44. [Google Scholar] [CrossRef]
- Wang, X.; Liu, L.; Zhang, W.; Zhang, J.; Du, X.; Huang, Q.; Tian, M.; Shen, H. Serum metabolome biomarkers associate low-level environmental perfluorinated compound exposure with oxidative /nitrosative stress in humans. Environ. Pollut. 2017, 229, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Pandey, K.B.; Rizvi, S.I. Markers of oxidative stress in erythrocytes and plasma during aging in humans. Oxid. Med. Cell. Longev. 2010, 3, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Barnham, K.J.; Masters, C.L.; Bush, A.I. Neurodegenerative diseases and oxidative stress. Nat. Rev. Drug Discov. 2004, 3, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Visconti, R.; Grieco, D. New insights on oxidative stress in cancer. Curr. Opin. Drug Discov. 2009, 12, 240–245. [Google Scholar]
- Tsutsui, H.; Kinugawa, S.; Matsushima, S. Oxidative stress and heart failure. Am. J. Physiol. Heart Circ. Physiol. 2011, 301, H2181–H2190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, X.Z.; Hu, D.C. Effects of perfluorooctanoate and perfluorooctane sulfonate exposure on hepatoma Hep G2 cells. Arch. Toxicol. 2009, 83, 851–861. [Google Scholar] [CrossRef]
- Harman, D. Origin and evolution of the free radical theory of aging: A brief personal history. Biogerontology 2009, 10, 773–781. [Google Scholar] [CrossRef] [PubMed]
- Mates, J.M.; Segura, J.A.; Alonso, F.J.; Márquez, J. Roles of dioxins and heavy metals in cancer and neurological diseases using ROS-mediated mechanisms. Free Radic. Biol. Med. 2010, 49, 1328–1341. [Google Scholar] [CrossRef]
- Santovito, G.; Trentin, E.; Gobbi, I.; Bisaccia, P.; Tallandini, I.; Irato, P. Non-enzymatic antioxidant responses of Mytilus galloprovincialis: Insights into the physiological role against metal-induced oxidative stress risk. Comp. Biochem. Physiol. C 2020, 240, 108909. [Google Scholar] [CrossRef]
- Wielsøe, M.; Long, M.; Ghisari, M.; Bonefeld-Jørgensen, E.C. Perfluoroalkylated substances (PFAS) affect oxidative stress biomarkers in vitro. Chemosphere 2015, 129, 239–245. [Google Scholar] [CrossRef]
- Zeng, Z.; Song, B.; Xiao, R.; Zeng, G.; Gong, J.; Chen, M.; Xu, P.; Zhang, P.; Shen, M.; Yi, H. Assessing the human health risks of perfluorooctane sulfonate by in vivo and in vitro studies. Environ. Int. 2019, 126, 598–610. [Google Scholar] [CrossRef] [PubMed]
- Aruoma, O.K. Free radicals, oxidative stress, and antioxidants in human health and disease. J. Am. Oil Chem. Soc. 1998, 75, 199–212. [Google Scholar] [CrossRef] [PubMed]
- Cooke, M.S.; Evans, M.D.; Dizdaroglu, M.; Lunec, J. Oxidative DNA damage: Mechanisms, mutation, and disease. FASEB J. 2003, 17, 1195–1214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suh, K.S.; Choi, E.M.; Kim, Y.J.; Hong, S.M.; Park, S.Y.; Rhee, S.Y.; Oh, S.; Kim, S.W.; Pak, Y.K.; Choe, W.; et al. Perfluorooctanoic acid induces oxidative damage and mitochondrial dysfunction in pancreatic β-cells. Mol. Med. Rep. 2017, 15, 3871–3878. [Google Scholar] [CrossRef] [Green Version]
- Watkins, A.M.; Wood, C.R.; Lin, M.T.; Abbott, B.D. The effects of perfluorinated chemicals on adipocyte differentiation in vitro. Mol. Cell. Endocrin. 2015, 400, 90–101. [Google Scholar] [CrossRef]
- Henry, N.D.; Fair, P.A. Comparison of in vitro cytotoxicity, estrogenicity and antiestrogenicity of triclosan, perfluorooctane sulfonate and perfluorooctanoic acid. J. Appl. Toxicol. 2013, 33, 265–272. [Google Scholar] [CrossRef]
- Kraugerud, M.; Zimmer, K.E.; Ropstad, E.; Verhaegen, S. Perfluorinated compounds differentially affect steroidogenesis and viability in the human adrenocortical carcinoma (H295R) in vitro cell assay. Toxicol. Lett. 2011, 205, 62–68. [Google Scholar] [CrossRef]
- Kamendulis, L.M.; Wu, Q.; Sandusky, G.E.; Hocevar, B.A. Perfluorooctanoic acid exposure triggers oxidative stress in the mouse pancreas. Toxicol. Rep. 2014, 1, 513–521. [Google Scholar] [CrossRef] [Green Version]
- Klaunig, J.E.; Babich, M.A.; Baetcke, K.P.; Cook, J.C.; Corton, J.C.; David, R.M.; DeLuca, J.G.; Lai, D.Y.; McKee, R.H.; Peters, J.M.; et al. PPARα agonist-induced rodent tumors: Modes of action and human relevance. Crit. Rev. Toxicol. 2003, 33, 655–780. [Google Scholar] [CrossRef]
- Andersen, M.E.; Butenhoff, J.L.; Chang, S.-C.; Farrar, D.G.; Kennedy, G.L., Jr.; Lau, C.; Olsen, G.W.; Seed, J.; Wallace, K.B. Perfluoroalkyl acids and related chemistries—Toxicokinetics and mode of action. Toxicol. Sci. 2008, 102, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Intrasuksri, U.; Rangwala, S.M.; O’Brien, M.; Noonan, D.J.; Feller, D.R. Mechanisms of peroxisome proliferation by perfluorooctanoic acid and endogenous fatty acids. Gen. Pharmacol. 1998, 31, 187–197. [Google Scholar] [CrossRef]
- Sohlenius, A.K.; Eriksson, A.M.; Högström, C.; Kimland, M.; DePierre, J.W. Perfluorooctane sulfonic acid is a potent inducer of peroxisomal fatty acid b-oxidation and other activities known to be affected by peroxisome proliferators in mouse liver. Pharmacol. Toxicol. 1993, 72, 90–93. [Google Scholar] [CrossRef] [PubMed]
- Rosen, M.B.; Lee, J.S.; Ren, H.; Vallanat, B.; Liu, J.; Waalkes, M.P.; Abbott, B.D.; Lau, C.; Corton, J.C. Toxicogenomic dissection of the perfluorooctanoic acid transcript profile in mouse liver: Evidence for the involvement of nuclear receptors PPARα and CAR. Toxicol. Sci. 2008, 103, 46–56. [Google Scholar] [CrossRef] [Green Version]
- Buhrke, T.; Kibellus, A.; Lampen, A. In vitro toxicological characterization of perfluorinated carboxylic acids with different carbon chain lengths. Toxicol. Lett. 2013, 218, 97–104. [Google Scholar] [CrossRef]
- Vanden Heuvel, J.P.; Thompson, J.T.; Frame, S.R.; Gillies, P.J. Differential activation of nuclear receptors by perfluorinated fatty acid analogs and natural fatty acids: A comparison of human, mouse, and rat peroxisome proliferator-activated receptor-a, b, and c, liver X receptor-b, and retinoid X receptor-a. Toxicol. Sci. 2006, 92, 476–489. [Google Scholar] [CrossRef] [Green Version]
- Berger, J.; Moller, D.E. The mechanisms of action of PPARs. Annu. Rev. Med. 2002, 53, 409–435. [Google Scholar] [CrossRef] [Green Version]
- Melzer, D.; Rice, N.; Depledge, M.H.; Henley, W.E.; Galloway, T.S. Association between serum perfluorooctanoic acid (PFOA) and thyroid disease in the U.S. National Health and Nutrition Examination Survey. Environ. Health Perspect. 2010, 118, 686–692. [Google Scholar] [CrossRef] [Green Version]
- Olsen, G.W.; Burlew, M.; Burris, J.; Mandel, J. A Cross-Sectional Analysis of Serum Perfluorooctanesulfonate (PFOS) and Perfluorooctanoate (PFOA) in Relation to Clinical Chemistry, Thyroid Hormone, Hematology and Urinalysis Results from Male and Female Employee Participants of the 2000 Antwerp and Decatur Fluorochemical Medical Surveillance Program 3M Company; Final Report; AR-226-1087; U.S. Environmental Protection Agency: Washington, DC, USA, 2001.
- Wen, L.-L.; Lin, L.-Y.; Su, T.-C.; Chen, P.-C.; Lin, C.-Y. Association between serum perfluorinated chemicals and thyroid function in U.S. adults: The national health and nutrition examination survey 2007–2010. J. Clin. Endocrin. Metab 2013, 98, E1456–E1464. [Google Scholar] [CrossRef]
- Shankar, A.; Xiao, J.; Ducatman, A. Perfluoroalkyl chemicals and chronic kidney disease in us adults. Am. J. Epidem. 2011, 174, 893–900. [Google Scholar] [CrossRef]
- Watkins, D.J.; Josson, J.; Elston, B.; Bartell, S.M.; Shin, H.-M.; Vieira, V.M.; Savitz, D.A.; Fletcher, T.; Wellenius, G.A. Exposure to perfluoroalkyl acids and markers of kidney function among children and adolescents living near a chemical plant. Environ. Health Perspect. 2013, 121, 625. [Google Scholar] [CrossRef] [Green Version]
- Fisher, M.; Arbuckle, T.E.; Wade, M.; Haines, D.A. Do perfluoroalkyl substances affect metabolic function and plasma lipids? Analysis of the 2007–2009, Canadian Health Measures Survey (CHMS) cycle 1. Environ. Res. 2013, 121, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.Y.; Chen, P.C.; Lin, Y.C.; Lin, L.Y. Association among serum perfluoroalkyl chemicals, glucose homeostasis, and metabolic syndrome in adolescents and adults. Diabetes Care 2009, 32, 702–707. [Google Scholar] [CrossRef] [Green Version]
- Olsen, G.W.; Gilliland, F.D.; Burlew, M.M.; Burris, J.M.; Mandel, J.S.; Mandel, J.H. An epidemiologic investigation of reproductive hormones in men with occupational exposure to perfluorooctanoic acid. J. Occupat. Environ. Med. 1998, 40, 614–622. [Google Scholar] [CrossRef]
- Itoh, S.; Araki, A.; Mitsui, T.; Miyashita, C.; Goudarzi, H.; Sasaki, S.; Cho, K.; Nakazawa, H.; Iwasaki, Y.; Shinohara, N.; et al. Association of perfluoroalkyl substances exposure in utero with reproductive hormone levels in cord blood in the Hokkaido study on environment children’s health. Environ. Int. 2016, 94, 51–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goudarzi, H.; Araki, A.; Itoh, S.; Sasaki, S.; Miyashita, C.; Mitsui, T.; Nakazawa, H.; Nonomura, K.; Kishi, R. The association of prenatal exposure to perfluorinated chemicals with glucocorticoid and androgenic hormones in cord blood samples: The Hokkaido study. Environ. Health Perspect. 2017, 125, 111–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minatoya, M.; Itoh, S.; Miyashita, C.; Araki, A.; Sasaki, S.; Miura, R.; Goudarzi, H.; Iwasaki, Y.; Kishi, R. Association of prenatal exposure to perfluoroalkyl substances with cord blood adipokines and birth size: The Hokkaido study on environment and children’s health. Environ. Res. 2017, 156, 175–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okada, E.; Sasaki, S.; Kashino, I.; Matsuura, H.; Miyashita, C.; Kobayashi, S.; Itoh, K.; Ikeno, T.; Tamakoshi, A.; Kishi, R. Prenatal exposure to perfluoroalkyl acids and allergic diseases in early childhood. Environ. Int. 2014, 65, 127–134. [Google Scholar] [CrossRef]
- Kishi, R.; Nakajima, T.; Goudarzi, H.; Kobayashi, S.; Sasaki, S.; Okada, E.; Miyashita, C.; Itoh, S.; Araki, A.; Ikeno, T.; et al. The association of prenatal exposure to perfluorinated chemicals with maternal essential and long-chain polyunsaturated fatty acids during pregnancy and the birth weight of their offspring: The Hokkaido study. Environ. Health Perspect. 2015, 123, 1038–1045. [Google Scholar] [CrossRef] [Green Version]
- Pérez, F.; Nadal, M.; Navarro-Ortega, A.; Fàbrega, F.; Domingo, J.L.; Barceló, D.; Farré, M. Accumulation of perfluoroalkyl substances in human tissues. Environ. Int. 2013, 59, 354–362. [Google Scholar] [CrossRef]
- Cargnelutti, F.; Di Nisio, A.; Pallotti, F.; Sabovic, I.; Spaziani, M.; Tarsitano, M.G.; Paoli, D.; Foresta, C. Effects of endocrine disruptors on foetal testis development, male puberty, and transition age. Endocrine 2020. [Google Scholar] [CrossRef]
- Di Nisio, A.; Foresta, C. Water and soil pollution as determinant of water and food quality/contamination and its impact on male fertility. Reprod. Biol. Endocrinol. 2019, 17, 4. [Google Scholar] [CrossRef] [PubMed]
- Hauser, R.; Skakkebaek, N.E.; Hass, U.; Toppari, J.; Juul, A.; Andersson, A.M.; Kortenkamp, A.; Heindel, J.J.; Trasande, L. Male reproductive disorders, diseases and costs of exposure to endocrine-disrupting chemicals in the European Union. J. Clin. Endocrinol. Metab 2015, 100, 1267–1277. [Google Scholar] [CrossRef] [PubMed]
- Darbandi, M.; Darbandi, S.; Agarwal, A.; Sengupta, P.; Durairajanayagam, D.; Henkel, R.; Sadeghi, M.R. Reactive oxygen species and male reproductive hormones. Reprod. Biol. Endocrinol. 2018, 16, 87. [Google Scholar] [CrossRef] [Green Version]
- Beattie, M.C.; Chen, H.; Fan, J.; Papadopoulos, V.; Miller, P.; Zirkin, R.P. Aging and luteinizing hormone effects on reactive oxygen species production and DNA damage in rat Leydig cells. Biol. Reprod. 2013, 88, 1–7. [Google Scholar] [CrossRef]
- Tostes, R.C.; Carneiro, F.S.; Carvalho, M.H.C.; Reckelhoff, J.F. Reactive oxygen species: Players in the cardiovascular effects of testosterone. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2015, 310, R1–R14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Z.; Zhang, H.; Liu, Y.; Xu, M.; Dai, J. Alterations in gene expression and testosterone synthesis in the testes of male rats exposed to perfluorododecanoic acid. Tox. Sci. 2007, 98, 206–215. [Google Scholar] [CrossRef] [Green Version]
- Wan, H.T.; Zhao, Y.G.; Wong, M.H.; Lee, K.F.; Yeung, W.S.B.; Giesy, J.P.; Wong, K.C.K. Testicular signaling is the potential target of perfluorooctanesulfonate-mediated subfertility in male mice. Biol. Reprod. 2011, 84, 1016–1023. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Lu, Y.; Luo, B.; Yan, S.; Guo, X.; Dai, J. Proteomic analysis of muse testis reveals perfluorooctanoic acid-induced reproductive dysfunction via direct disturbance of testicular steroidogenic machinery. J. Prot. Res. 2014, 13, 3370–3385. [Google Scholar] [CrossRef]
- Baker, H.; Burger, H.; de Kretser, D.; Hudson, B.; Santen, R.J.; Swerdloff, R.S. Relative Incidence of Etiologic Disorders in Male Infertility. In Male Reproductive Dysfunction: Diagnosis and Management of Hypogonadism, Infertility and Impotence; Santen, R.J., Swerdloff, R.S., Eds.; Marcel Dekker Inc.: New York, NY, USA, 1986; pp. 341–372. [Google Scholar]
- Bisht, S.; Faiq, M.; Tolahunase, M.; Dada, R. Oxidative stress and male infertility. Nat. Rev. Urol. 2017, 14, 470–485. [Google Scholar] [CrossRef]
- Ramalho-Santos, J.; Varum, S.; Amaral, S.; Mota, P.C.; Sousa, A.P.; Amaral, A. Mitochondrial functionality in reproduction: From gonads and gametes to embryos and embryonic stem cells. Hum. Reprod. Update 2009, 15, 553–572. [Google Scholar] [CrossRef] [Green Version]
- Blaylock, M.G.; Cuthbertson, B.H.; Galley, H.F.; Ferguson, N.R.; Webster, N.R. The effect of nitric oxide and peroxynitrite on apoptosis in human polymorphonuclear leukocytes. Free Radic. Biol. Med. 1998, 25, 748–752. [Google Scholar] [CrossRef]
- Wang, W.; Zhou, W.; Wu, S.; Liang, F.; Li, Y.; Zhang, J.; Cui, L.; Feng, Y.; Wang, Y. Perfluoroalkyl substances exposure and risk of polycystic ovarian syndrome related infertility in Chinese women. Envron. Pollut. 2019, 247, 824–831. [Google Scholar] [CrossRef]
- Masjedi, F.; Keshtgar, S.; Zal, F.; Talaei-Khozani, T.; Sameti, S.; Fallahi, S.; Kazeroni, M. Effects of vitamin D on steroidogenesis, reactive oxygen species production, and enzymatic antioxidant defence in human granulosa cells of normal and polycystic ovaries. J. Ster. Biochem. Mol. Biol. 2020, 197, 105521. [Google Scholar] [CrossRef]
- Di Nisio, A.; Rocca, M.S.; Sabovic, I.; De Rocco, M.; Corsini, P.C.; Guidolin, D.; Zanon, C.; Acquasaliente, L.; Carosso, A.R.; De Toni, L.; et al. Perfluorooctanoic acid alters progesterone activity in human endometrial cells and induces reproductive alterations in young women. Chemosphere 2020, 242, 125208. [Google Scholar] [CrossRef]
- Taketani, T.; Tamura, H.; Takasaki, A.; Lee, L.; Kizuka, F.; Tamura, I.; Taniguchi, K.; Maekawa, R.; Asada, H.; Shimamura, K.; et al. Protective role of melatonin in progesterone production by human luteal cells. J. Pineal. Res. 2011, 51, 207–213. [Google Scholar] [CrossRef]
- Mamsen, L.S.; Jonsson, B.A.G.; Lindh, C.H.; Olesen, R.H.; Larsen, A.; Ernst, E.; Kelsey, T.W.; Andersen, C.Y. Concentration of perfluorinated compounds in cotinine in human foetal organs, placenta, and maternal plasma. Sci. Tot. Environ. 2017, 596–597, 97–105. [Google Scholar] [CrossRef] [Green Version]
- Sagiv, S.K.; Rifas-Shiman, S.L.; Fleisch, A.F.; Webster, T.F.; Calafat, A.M.; Ye, X.; Gillman, M.W.; Oken, E. Early-pregnancy plasma concentrations of perfluoroalkyl substances and birth outcomes in Project Viva: Confounded by pregnancy hemodynamics? Am. J. Epidemiol. 2018, 187, 793–802. [Google Scholar] [CrossRef]
- Spratlen, M.J.; Perera, F.P.; Lederman, S.A.; Robinson, M.; Kannan, K.; Trasande, L.; Herbstman, J. Cord blood perfluoroalkyl substances in mothers exposed to the World Trade Center disaster during pregnancy. Environ. Pollut. 2020, 246, 482–490. [Google Scholar] [CrossRef]
- Midasch, O.; Drexler, H.; Hart, N.; Beckmann, M.W.; Angerer, J. Transplacental exposure of neonates to perfluorooctanesulfonate and perfluorooctanoate: A pilot study. Int. Arch. Occup. Environ. Health 2007, 80, 643–648. [Google Scholar] [CrossRef] [PubMed]
- Fei, C.; McLaughlin, J.K.; Tarone, R.E.; Olsen, J. Perfluorinated chemicals and fetal growth: A study within the danish national birth cohort. Environ. Health Perspect. 2007, 115, 1677–1682. [Google Scholar] [CrossRef] [PubMed]
- Winkens, K.V.R.; Berger, U.; Cousins, I. Early life exposure to per- and polyfluoroalkyl substances (PFAS): A critical review. Emer. Contamin. 2017, 3, 55–68. [Google Scholar] [CrossRef]
- Huo, X.; Zhang, L.; Huang, R.; Feng, L.; Wang, W.; Zhang, J. Perfluoroalkyl substances exposure in early pregnancy and preterm birth in singleton pregnancies: A prospective cohort study. Environ. Health 2020, 19, 60. [Google Scholar] [CrossRef] [PubMed]
- Trier, X.; Granby, K.; Christensen, J.H. Polyfluorinated surfactants (pfs) in paper and board coatings for food packaging. Environ. Sci. Pollut. Res. Int. 2011, 18, 1108–1120. [Google Scholar] [CrossRef]
- Washino, N.; Saijo, Y.; Sasaki, S.; Kato, S.; Ban, S.; Konishi, K.; Ito, R.; Nakata, A.; Iwasaki, Y.; Saito, K.; et al. Correlations between prenatal exposure to perfluorinated chemicals and reduced fetal growth. Environ. Health Perspect. 2009, 117, 660–667. [Google Scholar] [CrossRef] [Green Version]
- Bjerregaard-Olesen, C.; Bach, C.C.; Long, M.; Ghisari, M.; Bech, B.H.; Nohr, E.A.; Henriksen, T.B.; Olsen, J.; Bonefeld-Jørgensen, E.C. Determinants of serum levels of perfluorinated alkyl acids in Danish pregnant women. Int. J. Hyg. Environ. Health 2016, 219, 867–875. [Google Scholar] [CrossRef] [PubMed]
- Brantsaeter, A.L.; Whitworth, K.W.; Ydersbond, T.A.; Haug, L.S.; Haugen, M.; Knutsen, H.K.; Thomsen, C.; Meltzer, H.M.; Becher, G.; Sabaredzovic, A.; et al. Determinants of plasma concentrations of perfluoroalkyl substances in pregnant Norwegian women. Environ. Int. 2013, 54, 74–84. [Google Scholar] [CrossRef] [Green Version]
- Lewin, A.; Arbuckle, T.E.; Fisher, M.; Liang, C.L.; Marro, L.; Davis, K.; Abdelouahab, N.; Fraser, W.D. Univariate predictors of maternal concentrations of environmental chemicals: The MIREC study. Int. J. Hyg. Environ. Health 2017, 220, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Berg, V.; Nost, T.H.; Huber, S.; Rylander, C.; Hansen, S.; Veyhe, A.S.; Fuskevåg, O.M.; Odland, J.Ø.; Sandanger, T.M. Maternal serum concentrations of per-and polyfluoroalkyl substances and their predictors in years with reduced production and use. Environ. Int. 2014, 69, 58–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eriksson, U.; Mueller, J.F.; Toms, L.L.; Hobson, P.; Karrman, A. Temporal trends of PFSAs, PFCAs and selected precursors in Australian serum from 2002 to 2013. Environ. Pollut. 2017, 220, 168–177. [Google Scholar] [CrossRef]
- Wilhelm, M.; Wittsiepe, J.; Volkel, W.; Fromme, H.; Kasper-Sonnenberg, M. Perfluoroalkyl acids in children and their mothers: Association with drinking water and time trends of inner exposures-results of the Duisburg birth cohort and Bochum cohort studies. Int. J. Hyg. Environ. Health 2015, 218, 645–655. [Google Scholar] [CrossRef]
- Tsai, M.S.; Miyashita, C.; Araki, A.; Itoh, S.; Bamai, Y.A.; Goudarzi, H.; Okada, E.; Kashino, I.; Matsuura, H.; Kishi, R. Determinants and temporal trends of perfluoroalkyl substances in pregnant women: The Hokkaido study on Environment and Children’s Health. Int. J. Environ. Res. Public Health 2018, 15, 989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Cock, M.; de Boer, M.R.; Lamoree, M.; Legler, J.; van de Bor, M. Prenatal exposure to endocrine disrupting chemicals in relation to thyroid hormone levels in infants—A Dutch prospective cohort study. Environ. Health 2014, 13, 106. [Google Scholar] [CrossRef] [Green Version]
- Johnson, P.I.; Sutton, P.; Atchley, D.S.; Koustas, E.; Lam, J.; Sen, S.; Robinson, K.A.; Axelrad, D.A.; Woodruff, T.J.; Sutton, P.; et al. The navigation guide-evidence-based medicine meets environmental health: Systematic review of non-human evidence for PFOA effects on fetal growth. Environ. Health Perspect. 2014, 122, 1040–1051. [Google Scholar] [CrossRef] [Green Version]
- Bach, C.C.; Bech, B.H.; Brix, N.; Nohr, E.A.; Bonde, J.P.E.; Henriksen, T.B. Perfluoroalkyl and polyfluoroalkyl substances and human fetal growth: A systematic review. Crit. Rev. Toxicol. 2015, 45, 53–67. [Google Scholar] [CrossRef]
- Liew, Z.; Ritz, B.; Bonefeld-Jorgensen, E.C.; Henriksen, T.B.; Nohr, E.A.; Bech, B.H.; Fei, C.; Bossi, R.; von Ehrenstein, O.S.; Streja, E.; et al. Prenatal exposure to perfluoroalkyl substances and the risk of congenital cerebral palsy in children. Am. J. Epidemiol. 2014, 180, 574–581. [Google Scholar] [CrossRef] [Green Version]
- Jensen, T.K.; Andersen, L.B.; Kyhl, H.B.; Nielsen, F.; Christesen, H.T.; Grandjean, P. Association between perfluorinated compound exposure and miscarriage in Danish pregnant women. PLoS ONE 2015, 10, e0123496. [Google Scholar] [CrossRef]
- Apelberg, B.J.; Witter, F.R.; Herbstman, J.B.; Calafat, A.M.; Halden, R.U.; Needham, L.L.; Goldman, L.R. Cord serum concentrations of perfluorooctane sulfonate (PFOS) and perfluorooctanoate (PFOA) in relation to weight and size at birth. Environ. Health Perspect. 2007, 115, 1670–1676. [Google Scholar] [CrossRef] [Green Version]
- Negri, E.; Metruccio, F.; Guercio, V.; Tosti, L.; Benfenati, E.; Bonzi, R.; La Vecchia, C.; Moretto, A. Exposure to PFOA and PFOS and fetal growth: A critical merging of toxicological and epidemiological data. Crit. Rev. Toxicol. 2017, 47, 482–508. [Google Scholar] [CrossRef]
- Fei, C.; McLaughlin, J.K.; Tarone, R.E.; Olsen, J. Fetal growth indicators and perfluorinated chemicals: A study in the Danish National Birth Cohort. Am. J. Epidemiol. 2008, 168, 66–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, G.H.; Tung, K.Y.; Tsai, C.-H.; Liu, M.M.; Wang, D.; Liu, W.; Jin, Y.-H.; Hsieh, W.-S.; Lee, Y.L.; Chen, P.-C. Serum perfluoroalkyl concentrations, asthma outcomes, and immulogical markers in a case-control study of Taiwanese children. Environ. Health Perspect. 2013, 121, 507–513. [Google Scholar] [CrossRef] [Green Version]
- Dong, G.H.; Liu, M.M.; Wang, D.; Zheng, L.; Liang, Z.F.; Jin, Y.-H. Sub-chronic effect of perfluorooctanesulfonate (PFOS) on the balance of type 1 and type 2 cytokine in adult C57BL6 mice. Arch. Toxicol. 2011, 85, 1235–1244. [Google Scholar] [CrossRef] [PubMed]
- Humblet, O.; Diaz-Ramirez, L.G.; Balmes, J.R.; Pinney, S.M.; Hiatt, R.A. Perfluoroalkyl chemicals and asthma among children 12-19 years of age: NHANES (1999–2008). Environ. Health Perspect. 2014, 122, 1129–1233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gruber, C.; Illi, S.; Lau, S.; Nickel, R.; Forster, J.; Kamin, W.; Bauer, C.-P.; Wahn, V.; Wahn, U. Transient suppression of atopy in early childhood is associated with high vaccination coverage. Pediatrics. 2003, 111, 282–288. [Google Scholar] [CrossRef] [Green Version]
- Timmermann, C.A.; Rossing, L.I.; Grøntved, A.; Ried-Larsen, M.; Dalgård, C.; Andersen, L.B.; Grandjean, P.; Nielsen, F.; Svendsen, K.D.; Scheike, T.; et al. Adiposity and glycemic control in children exposed to perfluorinated compounds. J. Clin. Endocrinol. Metab. 2014, 99, E608–E614. [Google Scholar] [CrossRef] [Green Version]
- Ovsyannikova, I.G.; Dhiman, N.; Jacobson, R.M.; Vierkant, R.A.; Poland, G.A. Frequency of measles virus-specific CD4+ and CD8+ T cells in subjects seronegative or highly seropositive for measles vaccine. Clin. Diagn. Lab. Immunol. 2003, 10, 411–416. [Google Scholar] [CrossRef] [Green Version]
- Mora, A.M.; Fleisch, A.F.; Rifas-Shiman, S.L.; Baidal, J.A.W.; Pardo, L.; Webster, T.F.; Calafat, A.M.; Ye, X.; Oken, E.; Sagiv, S.K. Early life-exposure to per- and polyfluoroalkyl substances and mid-childhood lipid and alanine aminotransferase levels. Environ. Int. 2018, 111, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Olsen, G.W.; Butenhoff, J.L.; Zobel, L.R. Perfluoroalkyl chemicals and human fetal development: An epidemiologic review with clinical and toxicological perspectives. Reprod. Toxicol. 2009, 27, 212–213. [Google Scholar] [CrossRef]
- Kudo, N.; Katakura, M.; Sato, Y.; Kawashima, Y. Sex hormone-regulated renal transport of perfluorooctanoic acid. Chem. Biol. Interact. 2002, 139, 301–316. [Google Scholar] [CrossRef]
- Zhang, W.; Zhang, Y.; Taniyasu, S.; Yeung, L.W.Y.; Lam, P.K.S.; Wang, J.; Li, X.; Yamashita, N.; Dai, J. Distribution and fate of perfluoroalkyl substances in municipal wastewater treatment plants in economically developed areas in China. Environ. Pollut. 2013, 176, 10–17. [Google Scholar] [CrossRef]
- Blake, B.E.; Pinney, S.M.; Hines, E.P.; Fenton, S.E.; Ferguson, K.K. Associations between longitudinal serum perfluoroalkyl substance (PFAS) levels and measures of thyroid hormone, kidney function, and body mass index in the Fernald Community Cohort. Environ. Pollut. 2018, 242, 894–904. [Google Scholar] [CrossRef]
- Han, X.; Nabb, D.L.; Russell, M.H.; Kennedy, G.L.; Rickard, R.W. Renal elimination of perfluorocarboxylates (PFCAs). Chem. Res. Toxicol. 2012, 25, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Wolf, C.J.; Takacs, M.L.; Schmid, J.E.; Lau, C.; Abbott, B.D. Activation of mouse and human peroxisome proliferator-activated receptor alpha by perfluoroalkyl acids of different functional groups and chain lengths. Toxicol. Sci. 2008, 106, 162–171. [Google Scholar] [CrossRef] [Green Version]
- Hagenaars, A.; Vergauwen, L.; Benoot, D.; Laukens, K.; Knapen, D. Mechanistic toxicity study of perfluorooctanoic acid in zebrafish suggests mitochondrial dysfunction to play a key role in PFOA toxicity. Chemosphere 2013, 91, 844–856. [Google Scholar] [CrossRef] [PubMed]
- Stanifer, J.W.; Stapleton, H.M.; Souma, T.; Wittmer, A.; Zhao, X.; Boulware, L.E. Perfluorinated chemicals as emerging environmental threats to kidney health. A scoping review. Clin. J. Am. Soc. Nephrol. 2018, 13, 1479–1492. [Google Scholar] [CrossRef] [Green Version]
- Basu, G.; Mohapatra, A. Interactions between thyroid disorders and kidney disease. Indian J. Endocrinol. Metab. 2012, 16, 204–213. [Google Scholar] [CrossRef]
- Verner, M.A.; Loccisano, A.E.; Morken, N.H.; Yoon, M.; Wu, H.; McDougall, R.; Maisonet, M.; Marcus, M.; Kishi, R.; Miyashita, C.; et al. Associations of perfluoroalkyl substances (PFAS) with lower birth weight: An evaluation of potential confounding by glomerular filtration rate using a physiologically based pharmacokinetic model (PBPK). Environ. Health Perspect. 2015, 123, 1317–1324. [Google Scholar] [CrossRef]
- Bjork, J.; Butenhoff, J.; Wallace, K.J.T. Multiplicity of nuclear receptor activation by PFOA and PFOS in primary human and rodent hepatocytes. Toxicol. 2011, 288, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Rakhshandehroo, M.; Hooiveld, G.; Muller, M.; Kersten, S. Comparative analysis of gene regulation by the transcription factor PPARα between mouse and human. PLoS ONE 2009, 4, e6796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steenland, K.; Tinker, S.; Frisbee, S.; Ducatman, A.; Vaccarino, V. Association of perfluorooctanoic acid and perfluorooctane sulfonate with serum lipids among adults living near a chemical plant. Am. J. Epidemiol. 2009, 170, 1268–1278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olsen, G.W.; Zobel, L.R. Assessment of lipid, hepatic, and thyroid parameters with serum perfluorooctanoate (PFOA) concentrations in fluorochemical production workers. Int. Arch. Occup. Environ. Health 2007, 81, 231–246. [Google Scholar] [CrossRef] [Green Version]
- Steenland, K.; Savitz, T.F.; David, A. Epidemiologic evidence on the health effects of perfluorooctanoic acid (PFOA). Environ. Health Perspect. 2010, 118, 1100–1108. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.; Wang, H.; Yu, Y.Y.; Li, Y.B.; Naidu, R.; Liu, R. Using 2003–2014 U.S. NHANES data to determine the associations between per- and polyfluoroalkyl substances and cholesterol: Trend and implications. Ecotoxicol. Environ. Saf. 2019, 173, 461–468. [Google Scholar] [CrossRef]
- Nelson, J.W.; Hatch, E.E.; Webster, T.F. Exposure to polyfluoroalkyl chemicals and cholesterol, body weight, and insulin resistance in the general U.S. population. Environ. Health Perspect. 2010, 118, 197–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graber, J.M.; Alexander, C.; Laumbach, R.J.; Black, K.; Strickland, P.O.; Georgopoulos, P.G.; Marshall, E.G.; Shendell, D.G.; Alderson, D.; Mi, Z.; et al. Per and polyfluoroalkyl substances (PFAS) blood levels after contamination of a community water supply and comparison with 2013-2014 NHANES. J. Expo. Environ. Epidemiol. 2019, 29, 172–182. [Google Scholar] [CrossRef]
- Jain, R.B.; Ducatman, A. Roles of gender and obesity in defining correlations between perfluoroalkyl substances and lipid/lipoproteins. Sci. Tot. Environ. 2019, 653, 74–81. [Google Scholar] [CrossRef]
- Guruge, K.S.; Yeung, L.W.; Yamanaka, N.; Miyazaki, S.; Lam, P.K.; Giesy, J.P.; Jones, P.D.; Yamashita, N. Gene expression profiles in rat liver treated with perfluorooctanoic acid (PFOA). Toxicol. Sci. 2006, 89, 93–107. [Google Scholar] [CrossRef]
- Pedersen, K.E.; Letcher, R.J.; Sonne, C.; Dietz, R.; Styrishave, B. Per- and polyfluoroalkyl substances (PFAS)-new endocrine disruptors in polar bears (Ursus maritimus)? Environ. Int. 2016, 96, 180–189. [Google Scholar] [CrossRef]
- Scharmach, E.; Buhrke, T.; Lichtenstein, D.; Lampen, A. Perfluorooctanoic acid affects the activity of the hepatocyte nuclear factor 4 alpha (HNF4a). Toxicol. Lett. 2012, 212, 106–112. [Google Scholar] [CrossRef]
- Hayhurst, G.P.; Lee, Y.H.; Lambert, G.; Ward, J.M.; Gonzalez, F.J. Hepatocyte nuclear factor 4a (nuclear receptor 2A1) is essential for maintenance of hepatic gene expression and lipid homeostasis. Mol. Cell. Biol. 2001, 21, 1393–1403. [Google Scholar] [CrossRef] [Green Version]
- Rhee, J.; Inoue, Y.; Yoon, J.C.; Puigserver, P.; Fan, M.; Gonzalez, F.J.; Spiegelman, B.M. Regulation of hepatic fasting response by PPARγ coactivator-1a (PGC-1): Requirement for hepatocyte nuclear factor 4a in gluconeogenesis. Proc. Natl. Acad. Sci. USA 2003, 100, 4012–4017. [Google Scholar] [CrossRef] [Green Version]
- Ohguchi, H.; Tanaka, T.; Uchida, A.; Magoori, K.; Kudo, H.; Kim, I.; Daigo, K.; Sakakibara, I.; Okamura, M.; Harigae, H.; et al. Hepatocyte nuclear factor 4a contributes to thyroid hormone homeostasis by cooperatively regulating the type 1 iodothyronine deiodinase gene with GATA4 and Krüppel like transcription factor 9. Mol. Cell. Biol. 2008, 28, 3917–3931. [Google Scholar] [CrossRef] [Green Version]
- Sun, Q.; Zong, G.; Valvi, D.; Nielsen, F.; Coull, B.; Grandjean, P. Plasma concentrations of perfluoroalkyl substances and risk of Type 2 diabetes: A prospective investigation among U.S. women. Environ. Health Perspect. 2018, 126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domazet, S.L.; Grøntved, A.; Timmermann, A.G.; Nielsen, F.; Jensen, T.K. Longitudinal associations of exposure to perfluoroalkylated substances in childhood and adolescence and indicators of adiposity and glucose metabolism 6 and 12 years later: The European Youth Heart Study. Diabetes Care 2016, 39, 1745–1751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, T.C.; Kuo, C.C.; Hwang, J.J.; Lien, G.W.; Chen, M.F.; Chen, P.C. Serum perfluorinated chemicals, glucose homeostasis and the risk of diabetes in working-aged Taiwanese adults. Environ. Int. 2016, 88, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Lind, L.; Zethelius, B.; Salihovic, S.; van Bavel, B.; Lind, P.M. Circulating levels of perfluoroalkyl substances and prevalent diabetes in the elderly. Diabetologia 2014, 57, 473–479. [Google Scholar] [CrossRef] [PubMed]
- Cardenas, A.; Gold, D.R.; Hauser, R.; Kleinman, K.P.; Hivert, M.F.; Calafat, A.M.; Ye, X.; Webster, T.F.; Horton, E.S.; Oken, E. Plasma concentrations of per- and polyfluoroalkyl substances at baseline and associations with glycemic indicators and diabetes incidence among high-risk adults in the diabetes prevention program trial. Environ. Health Perspect. 2017, 125, 107001. [Google Scholar] [CrossRef] [Green Version]
- De Toni, L.; Radu, C.M.; Sabovic, I.; Di Nisio, A.; Dall’Acqua, S.; Guidolin, D.; Spampinato, S.; Campello, E.; Simioni, P.; Foresta, C. Increased cardiovascular risk associated with chemical sensitivity to perfluoro-octanoic acid: Role of impaired platelet aggregation. Int. J. Mol. Sci. 2020, 21, 399. [Google Scholar] [CrossRef] [Green Version]
- Tang, J.; Lu, X.; Chen, F.; Ye, X.; Zhou, D.; Yuan, J.; He, J.; Chen, B.; Shan, X.; Jiang, J.; et al. Effects of perfluorooctanoic acid on the associated genes expression of autophagy signaling pathway of Carassius auratus lymphocytes in vitro. Front. Physiol. 2018, 9, 1748. [Google Scholar] [CrossRef]
- Shi, X.; Zhou, B. The role of Nrf2 and MAPK pathways in PFOS-induced oxidative stress in zebrafish embryos. Toxicol. Sci. 2010, 115, 391–400. [Google Scholar] [CrossRef] [Green Version]
- Du, J.; Cai, J.; Wang, S.; You, H. Oxidative stress and apotosis to zebrafish (Danio rerio) embryos exposed to perfluorooctane sulfonate (PFOS) and ZnO nanoparticles. Int. J. Occup. Med. Environ. Health 2017, 30, 213–229. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Men, B.; He, Y.; Xu, H.; Liu, M.; Wang, D. Effect of single-wall carbon nanotubes on bioconcentration and toxicity of perfluorooctane sulfonate in zebrafish (Danio rerio). Sci. Total Environ. 2017, 607–608, 509–518. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Sheng, N.; Zhang, W.; Dai, J. Toxic effects of perfluorononanoic acid on the development of Zebrafish (Danio rerio) embryos. J. Environ. Sci. 2015, 32, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Shi, G.; Xie, Y.; Guo, Y.; Dai, J. 6:2 fluorotelomer sulfonamide alkylbetaine (6:2 FTAB), a novel perfluorooctane sulfonate alternative, induced developmental toxicity in zebrafish embryos. Aquat. Toxicol. 2018, 195, 24–32. [Google Scholar] [CrossRef]
- Wu, Y.; Huang, J.; Deng, M.; Jin, Y.; Yang, H.; Liu, Y.; Cao, Q.; Mennigen, J.A.; Tu, W. Acute exposure to environmentally relevant concentrations of Chinese PFOS alternative F-53B induces oxidative stress in early developing zebrafish. Chemosphere 2019, 235, 945–951. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.W.; Choi, K.; Park, K.; Seong, C.; Yu, S.D.; Kim, P. Adverse effects of perfluoroalkyl acids on fish and other aquatic organisms: A review. Sci. Total Environ. 2020, 707, 135334. [Google Scholar] [CrossRef] [PubMed]
- Rainieri, S.; Conlledo, N.; Langerholc, T.; Madorran, E.; Sala, M.; Barranco, A. Toxic effects of perfluorinated compounds at human cellular level and on a model vertebrate. Food Chem. Toxicol. 2017, 104, 14–25. [Google Scholar] [CrossRef]
- Oakes, K.D.; Sibley, P.K.; Solomon, K.R.; Mabury, S.A.; Van der Kraak, G.J. Impact of perfluorooctanoic acid on fathead minnow (Pimephales promelas) fatty acyl-CoA oxidase activity, circulating steroids, and reproduction in outdoor microcosms. Environ. Toxicol. Chem. 2004, 23, 1912–1919. [Google Scholar] [CrossRef]
- Garofalo, F.; Santovito, G.; Amelio, D. Morpho-functional effects of heat stress on the gills of Antarctic T. bernacchii and C. hamatus. Mar. Poll. Bull. 2019, 141, 194–204. [Google Scholar] [CrossRef]
- Wu, Y.; Deng, M.; Jin, Y.; Liu, X.; Mai, Z.; You, H.; Mu, X.; He, X.; Alharthi, R.; Kostyniuk, D.J.; et al. Toxicokinetics and toxic effects of a Chinese PFOS alternative F-53B in adult zebrafish. Ecotoxicol. Environ. Saf. 2019, 171, 460–466. [Google Scholar] [CrossRef]
- Wu, Y.; Deng, M.; Jin, Y.; Mu, X.; He, X.; Luu, N.T.; Yang, C.; Tu, W. Uptake and elimination of emerging polyfluoroalkyl substance F-53B in zebrafish larvae: Response of oxidative stress biomarkers. Chemosphere 2019, 215, 182–188. [Google Scholar] [CrossRef]
- Briels, N.; Ciesielski, T.M.; Herzke, D.; Jaspers, V.L.B. Developmental toxicity of perfluorooctanesulfonate (PFOS) and its chlorinated polyfluoroalkyl ether sulfonate alternative F-53B in the domestic chicken. Environ. Sci. Technol. 2018, 52, 12859–12867. [Google Scholar] [CrossRef] [PubMed]
- Cole, S.P.; Deeley, R.G. Transport of glutathione and glutathione conjugates by MRP1. Trends Pharmacol. Sci. 2006, 27, 438–446. [Google Scholar] [CrossRef]
- Houde, M.; Giraudo, M.; Douville, M.; Bougas, B.; Couture, P.; De Silva, A.O.; Spencer, C.; Lair, S.; Verreault, J.; Bernatchez, L.; et al. A multi-level biological approach to evaluate impacts of a major municipal effluent in wild St. Lawrence River yellow perch (Perca flavescens). Sci. Total Environ. 2014, 497–498, 307–318. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.H. Perfluorooctanoic acid induces peroxisomal fatty acid oxidation and cytokine expression in the liver of male Japanese medaka (Oryzias latipes). Chemosphere 2010, 81, 548–552. [Google Scholar] [CrossRef] [PubMed]
- Ferro, D.; Bakiu, R.; De Pittà, C.; Boldrin, F.; Cattalini, F.; Pucciarelli, S.; Miceli, C.; Santovito, G. Cu,Zn superoxide dismutases from Tetrahymena thermophila: Molecular evolution and gene expression of the first line of antioxidant defenses. Protist 2015, 166, 131–145. [Google Scholar] [CrossRef] [PubMed]
- Tilton, S.C.; Orner, G.A.; Benninghoff, A.D.; Carpenter, H.M.; Hendricks, J.D.; Pereira, C.B.; Williams, D.E. Genomic profiling reveals an alternate mechanism for hepatic tumor promotion by perfluorooctanoic acid in rainbow trout. Environ. Health Perspect. 2008, 116, 1047–1055. [Google Scholar] [CrossRef] [Green Version]
- Costantini, D.; Blévin, P.; Herzke, D.; Moe, B.; Gabrielsen, G.W.; Bustnes, J.O.; Chastel, O. Higher plasma oxidative damage and lower plasma antioxidant defences in an Arctic seabird exposed to longer perfluoroalkyl acids. Environ. Res. 2019, 168, 278–285. [Google Scholar] [CrossRef] [PubMed]
- Ferro, D.; Bakiu, R.; Pucciarelli, S.; Miceli, C.; Vallesi, A.; Irato, P.; Santovito, G. Molecular characterization, protein-protein interaction network, and evolution of four glutathione peroxidases from Tetrahymena thermophila. Antioxidants 2020, 9, 949. [Google Scholar] [CrossRef]
- Franchi, N.; Ferro, D.; Ballarin, L.; Santovito, G. Transcription of genes involved in glutathione biosynthesis in the solitary tunicate Ciona intestinalis exposed to metals. Aquat. Toxicol. 2012, 114–115, 14–22. [Google Scholar] [CrossRef]
- Arukwe, A.; Mortensen, A.S. Lipid peroxidation and oxidative stress responses of salmon fed a diet containing perfluorooctane sulfonic- or perfluorooctane carboxylic acids. Comp. Biochem. Physiol. C 2011, 154, 288–295. [Google Scholar] [CrossRef]
- Wei, Y.; Chan, L.L.; Wang, D.; Zhang, H.; Wang, J.; Dai, J. Proteomic analysis of hepatic protein profiles in rare minnow (Gobiocypris rarus) exposed to perfluorooctanoic acid. J. Proteome Res. 2008, 7, 1729–1739. [Google Scholar] [CrossRef]
- Wei, Y.; Liu, Y.; Wang, J.; Tao, Y.; Dai, J. Toxicogenomic analysis of the hepatic effects of perfluorooctanoic acid on rare minnows (Gobiocypris rarus). Toxicol. Appl. Pharmacol. 2008, 226, 285–297. [Google Scholar] [CrossRef]
- Kim, S.; Stroski, K.M.; Killeen, G.; Smitherman, C.; Simcik, M.F.; Brooks, B.W. 8:8 Perfluoroalkyl phosphinic acid affects neurobehavioral development, thyroid disruption, and DNA methylation in developing zebrafish. Sci. Total Environ. 2020, 736, 139600. [Google Scholar] [CrossRef]
- Collí-Dulá, R.C.; Martyniuk, C.J.; Streets, S.; Denslow, N.D.; Lehr, R. Molecular impacts of perfluorinated chemicals (PFAS) in the liver and testis of male largemouth bass (Micropterus salmoides) in Minnesota Lakes. Comp. Biochem. Physiol. D 2016, 19, 129–139. [Google Scholar] [CrossRef]
- Lavut, A.; Raveh, D. Sequestration of highly expressed mRNAs in cytoplasmic granules, P-bodies, and stress granules enhances cell viability. PLoS Genet. 2012, 8, e1002527. [Google Scholar] [CrossRef] [Green Version]
- Olszewska, M.; Bujarski, J.J.; Kurpisz, M. P-bodies and their functions during mRNA cell cycle: Mini-review. Cell Biochem. Funct. 2012, 30, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Nicorelli, E.; Gerdol, M.; Buonocore, F.; Pallavicini, A.; Scapigliati, G.; Guidolin, L.; Irato, P.; Corrà, F.; Santovito, G. First evidence of T cell restricted intracellular antigen (TIA) protein gene expression in Antarctic fish. Invert. Surviv. J. 2018, 15, 127. [Google Scholar]
- Drago, L.; Ballarin, L.; Santovito, G. Stress granules in Ciona robusta: Molecular evolution of TIAR and TTP and early evidence of their gene expression under stress conditions induced by metals. Invert. Surviv. J. 2020, 17, 12–13. [Google Scholar]
- Tolomeo, A.M.; Carraro, A.; Bakiu, R.; Toppo, S.; Gerdol, M.; Irato, P.; Pellegrino, D.; Garofalo, F.; Bisaccia, P.; Corrà, F.; et al. Too warm or not too warm... Is the antioxidant system of Antarctic fish ready to face climate changes? Invert. Surviv. J. 2020, 17, 11. [Google Scholar]
- Wang, Z.; Cousins, I.T.; Scheringer, M.; Hungerbühler, K. Fluorinated alternatives to long-chain perfluoroalkyl carboxylic acids (PFCAs), perfluoroalkane sulfonic acids (PFSAs) and their potential precursors. Environ. Int. 2013, 60, 242–248. [Google Scholar] [CrossRef]
- Wang, S.; Huang, J.; Yang, Y.; Hui, Y.; Ge, Y.; Larssen, T.; Yu, G.; Deng, S.; Wang, B.; Harman, C. First report of a Chinese PFOS alternative overlooked for 30 years: Its toxicity, persistence, and presence in the environment. Environ. Sci. Technol. 2013, 47, 10163–10170. [Google Scholar] [CrossRef]
- Munoz, G.; Liu, J.; Duy, S.V.; Sauvé, S. Analysis of F-53B, Gen-X, ADONA, and emerging fluoroalkylether substances in environmental and biomonitoring samples: A review. Trends Environ. Anal. Chem. 2019, 23, e00066. [Google Scholar] [CrossRef]
- Xiao, F. Emerging poly-and perfluoroalkyl substances in the aquatic environment: A review of current literature. Water Res. 2017, 124, 482–495. [Google Scholar] [CrossRef] [PubMed]
- Szilagyi, J.T.; Avula, V.; Fry, R.C. Perfluoroalkyl Substances (PFAS) and Their Effects on the Placenta, Pregnancy, and Child Development: A Potential Mechanistic Role for Placental Peroxisome Proliferator-Activated Receptors (PPARs). Curr. Environ. Health Rep. 2020, 7, 222–230. [Google Scholar] [CrossRef] [PubMed]


| PFAS | Use | Structure | Molecular Weight | Confirmed Toxic Effects |
|---|---|---|---|---|
| PFBA | synthetic chemistry |  | 214.04 | unknown |
| PFPeA | breakdown product of stain and grease-proof coatings on food packaging, couches, carpets |  | 264.05 | yes [61] |
| PFHxA | unknown |  | 314.05 | yes [62] |
| PFHpA | breakdown product of stain and grease-proof coatings on food packaging, couches, carpets |  | 364.06 | yes [63,64,65] |
| PFOA | water and oil repellant in fabrics and textiles, food packaging |  | 414.07 | yes [66,67,68] |
| PFNA | surfactant for synthesis of textiles and polymers |  | 464.08 | yes [61,69,70,71] |
| PFDA | breakdown product of stain and grease-proof coatings on food packaging, couches, carpets |  | 514.08 | yes [72,73] |
| PFUnA | unknown |  | 564.09 | yes [74] |
| PFDoA | unknown |  | 614.1 | yes [75,76,77] |
| PFBS | stain repellant |  | 300.1 | yes [78] |
| PFHxS | surfactant for textiles |  | 400.11 | unknown |
| PFOS | firefighting foam, textiles |  | 500.13 | yes [79,80,81] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bonato, M.; Corrà, F.; Bellio, M.; Guidolin, L.; Tallandini, L.; Irato, P.; Santovito, G. PFAS Environmental Pollution and Antioxidant Responses: An Overview of the Impact on Human Field. Int. J. Environ. Res. Public Health 2020, 17, 8020. https://doi.org/10.3390/ijerph17218020
Bonato M, Corrà F, Bellio M, Guidolin L, Tallandini L, Irato P, Santovito G. PFAS Environmental Pollution and Antioxidant Responses: An Overview of the Impact on Human Field. International Journal of Environmental Research and Public Health. 2020; 17(21):8020. https://doi.org/10.3390/ijerph17218020
Chicago/Turabian StyleBonato, Marco, Francesca Corrà, Marta Bellio, Laura Guidolin, Laura Tallandini, Paola Irato, and Gianfranco Santovito. 2020. "PFAS Environmental Pollution and Antioxidant Responses: An Overview of the Impact on Human Field" International Journal of Environmental Research and Public Health 17, no. 21: 8020. https://doi.org/10.3390/ijerph17218020
APA StyleBonato, M., Corrà, F., Bellio, M., Guidolin, L., Tallandini, L., Irato, P., & Santovito, G. (2020). PFAS Environmental Pollution and Antioxidant Responses: An Overview of the Impact on Human Field. International Journal of Environmental Research and Public Health, 17(21), 8020. https://doi.org/10.3390/ijerph17218020