
Kisspeptin-8 Induces Anxiety-Like Behavior and Hypolocomotion by Activating the HPA Axis and Increasing GABA Release in the Nucleus Accumbens in Rats

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Housing Conditions
2.2. Intracerebroventricular Cannulation
2.3. Peptide Synthesis
2.4. Treatment
2.5. Behavioral Tests
2.5.1. Elevated Plus Maze Test
2.5.2. Computerized Open Field Test
2.5.3. Marble Burying Test
2.6. Serum Corticosterone, Luteinizing Hormone and Total Protein Concentration Measurement
2.7. Ex Vivo Superfusion
2.8. Statistical Analysis
3. Results
3.1. Behavioral Tests
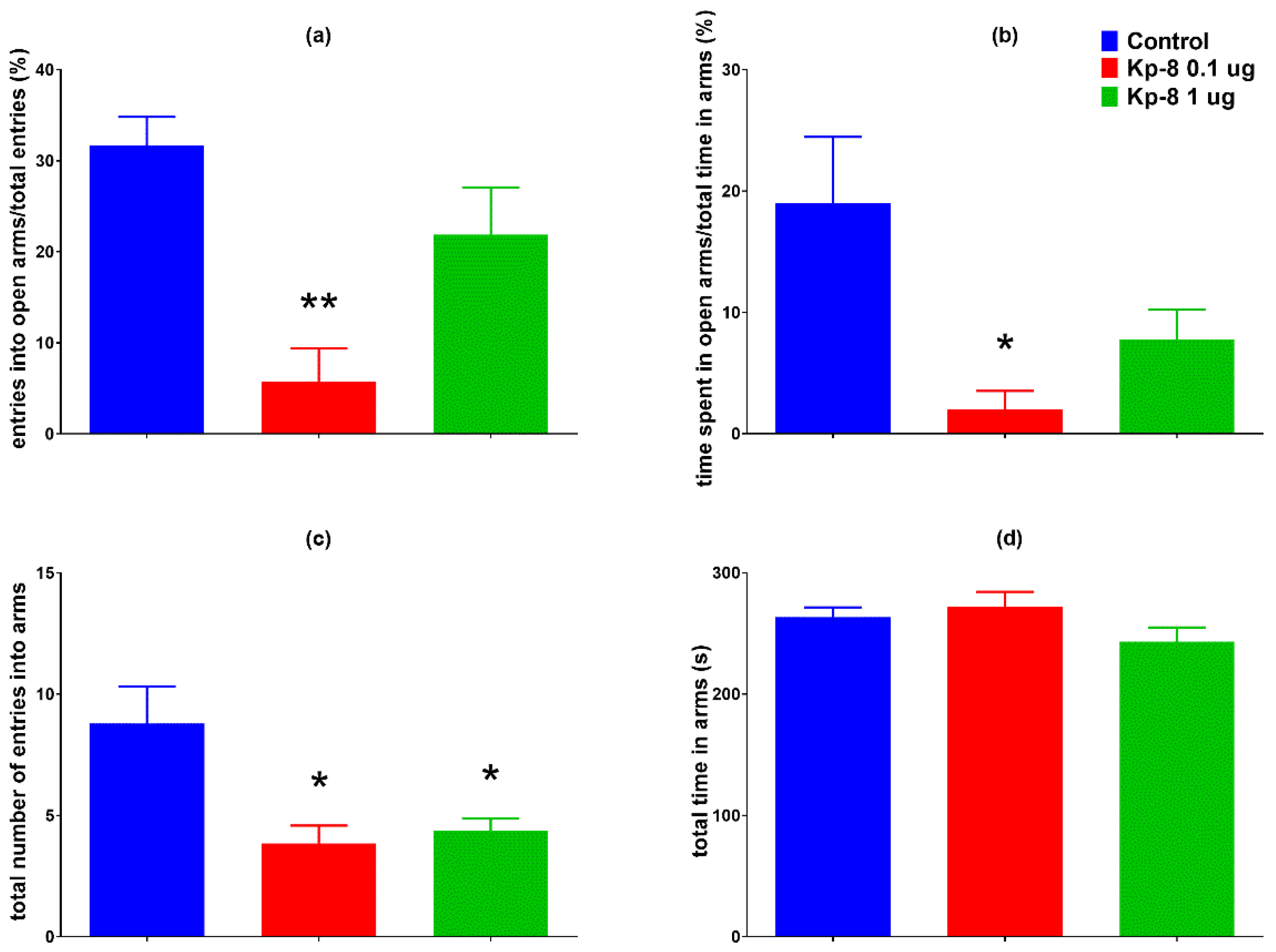
3.1.1. Elevated Plus Maze
3.1.2. Computerized Open Field Test
3.1.3. Marble Burying Test
3.2. Serum Corticosterone, LH and Total Protein
3.3. Ex Vivo Superfusion
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
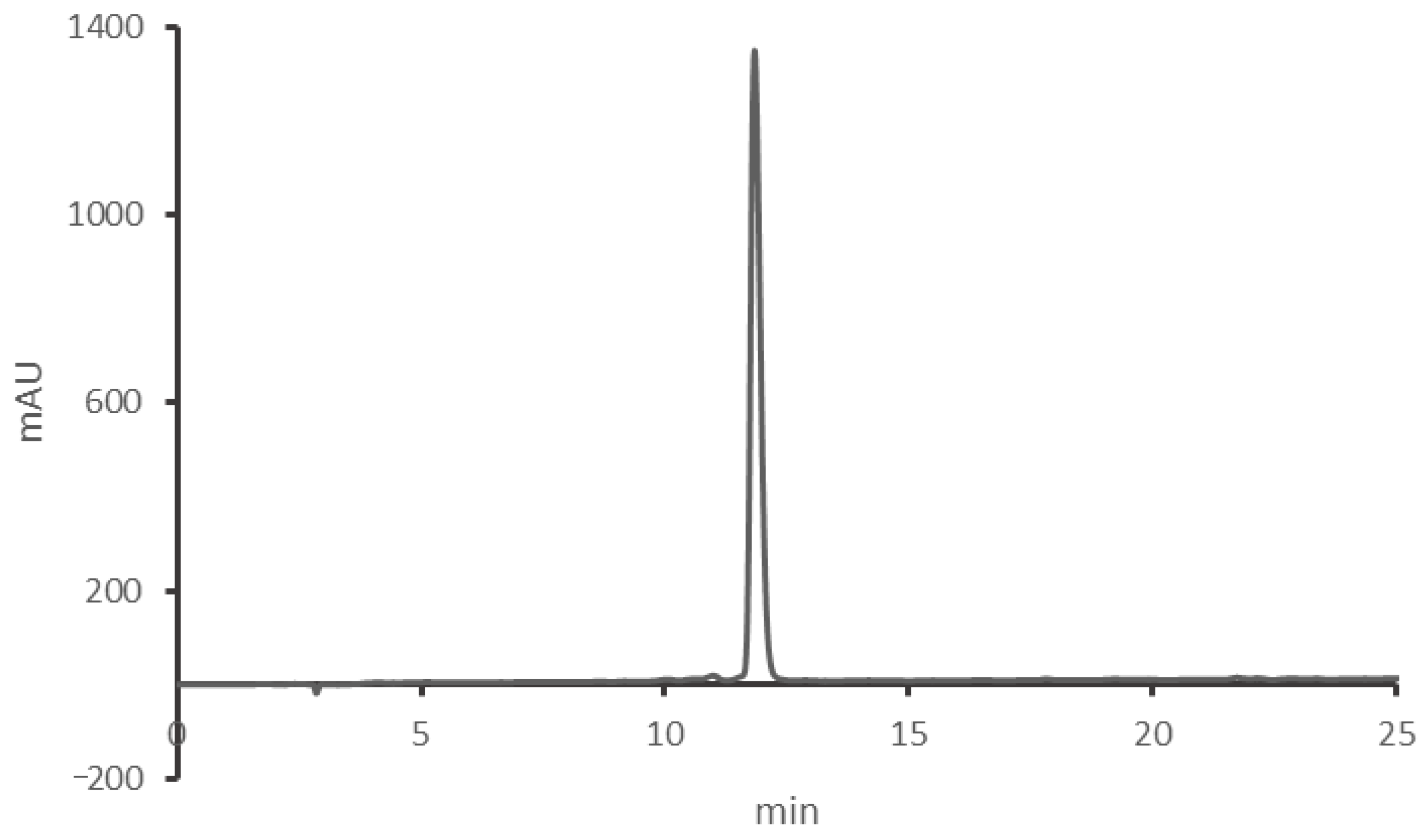

Appendix B

References
- Lee, J.; Miele, M.E.; Hicks, D.J.; Karen, K.; Trent, J.; Weissman, B.; Welch, D.R. KiSS-1, a Novel Human Malignant Melanoma Metastasis-Suppressor Gene. J. Natl. Cancer Inst. 1996, 88, 1731–1737. [Google Scholar] [CrossRef] [PubMed]
- Kotani, M.; Detheux, M.; Vandenbogaerde, A.; Communi, D.; Vanderwinden, J.M.; Le Poul, E.; Brézillon, S.; Tyldesley, R.; Suarez-Huerta, N.; Vandeput, F.; et al. The Metastasis Suppressor Gene KiSS-1 Encodes Kisspeptins, the Natural Ligands of the Orphan G Protein-coupled Receptor GPR54. J. Biol. Chem. 2001, 276, 34631–34636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quillet, R.; Ayachi, S.; Bihel, F.; Elhabazi, K.; Ilien, B.; Simonin, F. RF-amide neuropeptides and their receptors in Mammals: Pharmacological properties, drug development and main physiological functions. Pharmacol. Ther. 2016, 160, 84–132. [Google Scholar] [CrossRef] [PubMed]
- Franssen, D.; Tena-Sempere, M. The kisspeptin receptor: A key G-protein-coupled receptor in the control of the reproductive axis. Best Pract. Res. Clin. Endocrinol. Metab. 2018, 32, 107–123. [Google Scholar] [CrossRef]
- Lee, D.K.; Nguyen, T.; O’Neill, G.P.; Cheng, R.; Liu, Y.; Howard, A.D.; Coulombe, N.; Tan, C.P.; Tang-Nguyen, A.-T.; George, S.R.; et al. Discovery of a receptor related to the galanin receptors. FEBS Lett. 1999, 446, 103–107. [Google Scholar] [CrossRef]
- Ohtaki, T.; Shintani, Y.; Honda, S.I.; Matsumoto, H.; Hori, A.; Kanehashi, K.; Terao, Y.; Kumano, S.; Takatsu, Y.; Masuda, Y.; et al. Metastasis suppressor gene KiSS-1 encodes peptide ligand of a G-protein-coupled receptor. Nature 2001, 411, 613–617. [Google Scholar] [CrossRef]
- Cargnello, M.; Roux, P.P. Activation and Function of the MAPKs and Their Substrates, the MAPK-Activated Protein Kinases. Microbiol. Mol. Biol. Rev. 2011, 75, 50–83. [Google Scholar] [CrossRef] [Green Version]
- Peng, J.; Tang, M.; Zhang, B.P.; Zhang, P.; Zhong, T.; Zong, T.; Yang, B.; Kuang, H. Bin Kisspeptin stimulates progesterone secretion via the Erk1/2 mitogen-activated protein kinase signaling pathway in rat luteal cells. Fertil. Steril. 2013, 99, 1436–1443.e1. [Google Scholar] [CrossRef]
- Taylor, J.; Pampillo, M.; Bhattacharya, M.; Babwah, A.V. Kisspeptin/KISS1R signaling potentiates extravillous trophoblast adhesion to type-I collagen in a PKC- and ERK1/2-dependent manner. Mol. Reprod. Dev. 2014, 81, 42–54. [Google Scholar] [CrossRef]
- Bowe, J.E.; Chander, A.; Liu, B.; Persaud, S.J.; Jones, P.M. The permissive effects of glucose on receptor-operated potentiation of insulin secretion from mouse islets: A role for ERK1/2 activation and cytoskeletal remodelling. Diabetologia 2013, 56, 783–791. [Google Scholar] [CrossRef]
- Oishi, S.; Misu, R.; Tomita, K.; Setsuda, S.; Masuda, R.; Ohno, H.; Ieda, N.; Tsukamura, H.; Inoue, N.; Hirasawa, A.; et al. Activation of Neuropeptide FF Receptors by Kisspeptin Receptor Ligands. ACS Med. Chem. Lett. 2010, 2, 53–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brailoiu, G.C.; Dun, S.L.; Ohsawa, M.; Yin, D.; Yang, J.; Jaw, K.C.; Brailoiu, E.; Dun, N.J. KiSS-1 expression and metastin-like immunoreactivity in the rat brain. J. Comp. Neurol. 2005, 481, 314–329. [Google Scholar] [CrossRef] [PubMed]
- Overgaard, A.; Tena-Sempere, M.; Franceschini, I.; Desroziers, E.; Simonneaux, V.; Mikkelsen, J.D. Comparative analysis of kisspeptin-immunoreactivity reveals genuine differences in the hypothalamic Kiss1 systems between rats and mice. Peptides 2013, 45, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Higo, S.; Honda, S.; Iijima, N.; Ozawa, H. Mapping of Kisspeptin Receptor mRNA in the Whole Rat Brain and its Co-Localisation with Oxytocin in the Paraventricular Nucleus. J. Neuroendocrinol. 2016, 28, 1–8. [Google Scholar] [CrossRef]
- Bonini, J.A.; Jones, K.A.; Adham, N.; Forray, C.; Artymyshyn, R.; Durkin, M.M.; Smith, K.E.; Tamm, J.A.; Boteju, L.W.; Lakhlani, P.P.; et al. Identification and characterization of two G protein-coupled receptors for neuropeptide FF. J. Biol. Chem. 2000, 275, 39324–39331. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.H.; Tao, P.L.; Huang, E.Y.K. Distribution of neuropeptide FF (NPFF) receptors in correlation with morphine-induced reward in the rat brain. Peptides 2010, 31, 1374–1382. [Google Scholar] [CrossRef]
- Guzman, S.; Brackstone, M.; Radovick, S.; Babwah, A.V.; Bhattacharya, M.M. KISS1/KISS1R in Cancer: Friend or Foe? Front. Endocrinol. 2018, 9, 1–9. [Google Scholar] [CrossRef]
- Seminara, S.B.; Messager, S.; Dixon, J.; Slaugenhaupt, S.A.; O’Rahilly, S.; Carlton, M.B.L.; Bo-Abbas, Y.; Schwinof, K.M.; Chatzidaki, E.E.; Gusella, J.F.; et al. The GPR54 Gene as a Regulator of Puberty. N. Engl. J. Med. 2003, 349, 1614–1627. [Google Scholar] [CrossRef] [Green Version]
- Trevisan, C.M.; Montagna, E.; De Oliveira, R.; Christofolini, D.M.; Barbosa, C.P.; Crandall, K.A.; Bianco, B. Kisspeptin/GPR54 System: What Do We Know about Its Role in Human Reproduction? Cell. Physiol. Biochem. 2018, 870, 1259–1276. [Google Scholar] [CrossRef]
- Putteeraj, M.; Soga, T.; Ubuka, T.; Parhar, I.S. A “timed” kiss is essential for reproduction: Lessons from mammalian studies. Front. Endocrinol. 2016, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Pinilla, L.; Aguilar, E.; Dieguez, C.; Millar, R.P.; Tena-Sempere, M. Kisspeptins and Reproduction: Physiological Roles and Regulatory Mechanisms. Physiol. Rev. 2012, 92, 1235–1316. [Google Scholar] [CrossRef] [PubMed]
- Manfredi-Lozano, M.; Roa, J.; Tena-Sempere, M. Connecting metabolism and gonadal function: Novel central neuropeptide pathways involved in the metabolic control of puberty and fertility. Front. Neuroendocrinol. 2018, 48, 37–49. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, Y.; Ikegami, K.; Ishigaki, R.; Ieda, N.; Uenoyama, Y.; Maeda, K.I.; Tsukamura, H.; Inoue, N. Enhancement of the luteinising hormone surge by male olfactory signals is associated with anteroventral periventricular Kiss1 cell activation in female rats. J. Neuroendocrinol. 2017, 29, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Yeo, S.H.; Colledge, W.H. The role of Kiss1 neurons as integrators of endocrine, metabolic, and environmental factors in the hypothalamic-pituitary-gonadal axis. Front. Endocrinol. 2018, 9, 188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Padilla, S.L.; Perez, J.G.; Ben-hamo, M.; Bussi, I.L.; Palmiter, R.D.; Iglesia, H.O. De Kisspeptin Neurons in the Arcuate Nucleus of the Hypothalamus Orchestrate Circadian Rhythms and Metabolism. Curr. Biol. 2019, 29, 592–604.e4. [Google Scholar] [CrossRef] [Green Version]
- Ayachi, S.; Simonin, F. Involvement of Mammalian RF-Amide Peptides and Their Receptors in the Modulation of Nociception in Rodents. Front. Endocrinol. 2014, 5, 158. [Google Scholar] [CrossRef] [Green Version]
- Elhabazi, K.; Humbert, J.P.; Bertin, I.; Schmitt, M.; Bihel, F.; Bourguignon, J.J.; Bucher, B.; Becker, J.A.J.; Sorg, T.; Meziane, H.; et al. Endogenous mammalian RF-amide peptides, including PrRP, kisspeptin and 26RFa, modulate nociception and morphine analgesia via NPFF receptors. Neuropharmacology 2013, 75, 164–171. [Google Scholar] [CrossRef]
- Csabafi, K.; Bagosi, Z.; Dobó, É.; Szakács, J.; Telegdy, G.; Szabó, G. Kisspeptin modulates pain sensitivity of CFLP mice. Peptides 2018, 105, 21–27. [Google Scholar] [CrossRef]
- Clarkson, J.; d’Anglemont de Tassigny, X.; Colledge, W.H.; Caraty, A.; Herbison, A.E. Distribution of kisspeptin neurones in the adult female mouse brain. J. Neuroendocrinol. 2009, 21, 673–682. [Google Scholar] [CrossRef]
- Herbison, A.E.; De Tassigny, X.D.A.; Doran, J.; Colledge, W.H. Distribution and postnatal development of Gpr54 gene expression in mouse brain and gonadotropin-releasing hormone neurons. Endocrinology 2010, 151, 312–321. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, M.; Csabafi, K.; Telegdy, G. Neurotransmissions of antidepressant-like effects of kisspeptin-13. Regul. Pept. 2013, 180, 1–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Comninos, A.N.; Wall, M.B.; Demetriou, L.; Thomas, S.A.; Wilson, S.R.; Jayasena, C.N.; Bloom, S.R.; Bassett, P.; Hönigsperger, C.; Mehta, A.; et al. Kisspeptin modulates sexual and emotional brain processing in humans. J. Clin. Investig. 2017, 127, 709–719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakker, J.; Pierman, S.; González-Martínez, D. Effects of aromatase mutation (ArKO) on the sexual differentiation of kisspeptin neuronal numbers and their activation by same versus opposite sex urinary pheromones. Horm. Behav. 2010, 57, 390–395. [Google Scholar] [CrossRef] [PubMed]
- Hellier, V.; Brock, O.; Candlish, M.; Desroziers, E.; Aoki, M.; Mayer, C.; Piet, R.; Herbison, A.; Colledge, W.H.; Prévot, V.; et al. Female sexual behavior in mice is controlled by kisspeptin neurons. Nat. Commun. 2018, 9, 1–2. [Google Scholar] [CrossRef]
- Kinsey-Jones, J.S.; Li, X.F.; Knox, A.M.I.; Wilkinson, E.S.; Zhu, X.L.; Chaudhary, A.A.; Milligan, S.R.; Lightman, S.L.; O’Byrne, K.T. Down-regulation of hypothalamic kisspeptin and its receptor, Kiss1r, mRNA expression is associated with stress-induced suppression of luteinising hormone secretion in the female rat. J. Neuroendocrinol. 2009, 21, 20–29. [Google Scholar] [CrossRef]
- Rao, Y.S.; Mott, N.N.; Pak, T.R. Effects of kisspeptin on parameters of the HPA axis. Endocrine 2011, 39, 220–228. [Google Scholar] [CrossRef]
- Csabafi, K.; Jászberényi, M.; Bagosi, Z.; Lipták, N.; Telegdy, G. Effects of kisspeptin-13 on the hypothalamic-pituitary-adrenal axis, thermoregulation, anxiety and locomotor activity in rats. Behav. Brain Res. 2013, 241, 56–61. [Google Scholar] [CrossRef] [Green Version]
- Delmas, S.; Porteous, R.; Bergin, D.H.; Herbison, A.E. Altered aspects of anxiety-related behavior in kisspeptin receptor-deleted male mice. Sci. Rep. 2018, 8, 2–8. [Google Scholar] [CrossRef] [Green Version]
- Ogawa, S.; Parhar, I.S. Biological significance of kisspeptin-Kiss 1 receptor signaling in the habenula of teleost species. Front. Endocrinol. 2018, 9, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Adekunbi, D.A.; Li, X.F.; Lass, G.; Shetty, K.; Adegoke, O.A.; Yeo, S.H.; Colledge, W.H.; Lightman, S.L.; O’Byrne, K.T. Kisspeptin neurones in the posterodorsal medial amygdala modulate sexual partner preference and anxiety in male mice. J. Neuroendocrinol. 2018, 30, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Thomson, E.L.; Patterson, M.; Murphy, K.G.; Smith, K.L.; Dhillo, W.S.; Todd, J.F.; Ghatei, M.A.; Bloom, S.R. Central and peripheral administration of kisspeptin-10 stimulates the hypothalamic-pituitary-gonadal axis. J. Neuroendocrinol. 2004, 16, 850–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogawa, S.; Sivalingam, M.; Anthonysamy, R.; Parhar, I.S. Distribution of Kiss2 receptor in the brain and its localization in neuroendocrine cells in the zebrafish. Cell Tissue Res. 2020, 379, 349–372. [Google Scholar] [CrossRef] [PubMed]
- Tolson, K.P.; Garcia, C.; Yen, S.; Simonds, S.; Stefanidis, A.; Lawrence, A.; Smith, J.T.; Kauffman, A.S. Impaired kisspeptin signaling decreases metabolism and promotes glucose intolerance and obesity. J. Clin. Investig. 2014, 124, 3075–3079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desroziers, E.; Mikkelsen, J.; Simonneaux, V.; Keller, M.; Tillet, Y.; Caraty, A.; Franceschini, I. Mapping of kisspeptin fibres in the brain of the pro-oestrous rat. J. Neuroendocrinol. 2010, 22, 1101–1112. [Google Scholar] [CrossRef] [PubMed]
- Mogenson, G.J.; Wu, M. Effects of administration of dopamine D2 agonist quinpirole on exploratory locomotion. Brain Res. 1991, 551, 216–220. [Google Scholar] [CrossRef]
- Yael, D.; Tahary, O.; Gurovich, B.; Belelovsky, K.; Bar-Gad, I. Disinhibition of the nucleus accumbens leads to macro-scale hyperactivity consisting of micro-scale behavioral segments encoded by striatal activity. J. Neurosci. 2019, 39, 5897–5909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boekhoudt, L.; Omrani, A.; Luijendijk, M.C.M.; Wolterink-Donselaar, I.G.; Wijbrans, E.C.; van der Plasse, G.; Adan, R.A.H. Chemogenetic activation of dopamine neurons in the ventral tegmental area, but not substantia nigra, induces hyperactivity in rats. Eur. Neuropsychopharmacol. 2016, 26, 1784–1793. [Google Scholar] [CrossRef]
- Szeliga, A.; Podfigurna, A.; Bala, G.; Meczekalski, B. Kisspeptin and neurokinin B analogs use in gynecological endocrinology: Where do we stand? J. Endocrinol. Investig. 2020, 43, 555–561. [Google Scholar] [CrossRef]
- Asami, T.; Nishizawa, N.; Ishibashi, Y.; Nishibori, K.; Nakayama, M.; Horikoshi, Y.; Matsumoto, S.I.; Yamaguchi, M.; Matsumoto, H.; Tarui, N.; et al. Serum stability of selected decapeptide agonists of KISS1R using pseudopeptides. Bioorg. Med. Chem. Lett. 2012, 22, 6391–6396. [Google Scholar] [CrossRef]
- Rather, M.A.; Basha, S.H.; Bhat, I.A.; Sharma, N.; Nandanpawar, P.; Badhe, M.; Gireesh-Babu, P.; Chaudhari, A.; Sundaray, J.K.; Sharma, R. Characterization, molecular docking, dynamics simulation and metadynamics of kisspeptin receptor with kisspeptin. Int. J. Biol. Macromol. 2017, 101, 241–253. [Google Scholar] [CrossRef]
- Walf, A.A.; Frye, C.A. The use of the elevated plus maze as an assay of anxiety-related behavior in rodents. Nat. Protoc. 2007, 2, 322–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Brouwer, G.; Wolmarans, D.W. Back to basics: A methodological perspective on marble-burying behavior as a screening test for psychiatric illness. Behav. Process. 2018, 157, 590–600. [Google Scholar] [CrossRef] [PubMed]
- Schneider, T.; Popik, P. Attenuation of estrous cycle-dependent marble burying in female rats by acute treatment with progesterone and antidepressants. Psychoneuroendocrinology 2007, 32, 651–659. [Google Scholar] [CrossRef] [PubMed]
- Heffner, T.G.; Hartman, J.A.; Seiden, L.S. A rapid method for the regional dissection of the rat brain. Pharmacol. Biochem. Behav. 1980, 13, 453–456. [Google Scholar] [CrossRef]
- Salvatore, M.F.; Pruett, B.S.; Dempsey, C.; Fields, V. Comprehensive profiling of dopamine regulation in substantia nigra and ventral tegmental area. J. Vis. Exp. 2012, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Jászberényi, M.; Bagosi, Z.; Thurzó, B.; Földesi, I.; Szabó, G.; Telegdy, G. Endocrine, behavioral and autonomic effects of neuropeptide AF. Horm. Behav. 2009, 56, 24–34. [Google Scholar] [CrossRef]
- Kushikata, T.; Yoshida, H.; Kudo, M.; Salvadori, S.; Calo, G.; Hirota, K. The effects of neuropeptide S on general anesthesia in rats. Anesth. Analg. 2011, 112, 845–849. [Google Scholar] [CrossRef]
- Gyires, K.; Zádori, Z.S. Brain neuropeptides in gastric mucosal protection. Curr. Opin. Pharmacol. 2014, 19, 24–30. [Google Scholar] [CrossRef] [Green Version]
- Kastin, A.J.; Pan, W. Peptides and hormesis. Crit. Rev. Toxicol. 2008, 38, 629–631. [Google Scholar] [CrossRef]
- Calabrese, E.J. Hormetic mechanisms. Crit. Rev. Toxicol. 2013, 43, 580–606. [Google Scholar] [CrossRef]
- Rouméas, L.; Humbert, J.P.; Schneider, S.; Doebelin, C.; Bertin, I.; Schmitt, M.; Bourguignon, J.J.; Simonin, F.; Bihel, F. Effects of systematic N-terminus deletions and benzoylations of endogenous RF-amide peptides on NPFF1R, NPFF2R, GPR10, GPR54 and GPR103. Peptides 2015, 71, 156–161. [Google Scholar] [CrossRef] [PubMed]
- Palotai, M.; Telegdy, G.; Tanaka, M.; Bagosi, Z.; Jászberényi, M. Neuropeptide AF induces anxiety-like and antidepressant-like behavior in mice. Behav. Brain Res. 2014, 274, 264–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaewwongse, M.; Takayanagi, Y.; Onaka, T. Effects of RFamide-Related Peptide (RFRP)-1 and RFRP-3 on oxytocin release and anxiety-related behaviour in rats. J. Neuroendocrinol. 2011, 23, 20–27. [Google Scholar] [CrossRef]
- Aguilera, G. The Hypothalamic–Pituitary–Adrenal Axis and Neuroendocrine Responses to Stress. In Handbook of Neuroendocrinology; Elsevier: London, UK, 2012; pp. 175–196. [Google Scholar]
- Mills, E.G.A.; O’Byrne, K.T.; Comninos, A.N. The Roles of the Amygdala Kisspeptin System. Semin. Reprod. Med. 2019, 37, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Herman, J.P.; McKlveen, J.M.; Ghosal, S.; Kopp, B.; Wulsin, A.; Makinson, R.; Scheimann, J.; Myers, B. Regulation of the Hypothalamic-Pituitary-Adrenocortical Stress Response. In Comprehensive Physiology; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2016; Volume 6, pp. 603–621. [Google Scholar]
- Packard, A.E.B.; Egan, A.E.; Ulrich-Lai, Y.M. HPA axis interactions with behavioral systems. Compr. Physiol. 2016, 6, 1897–1934. [Google Scholar] [CrossRef]
- Seibenhener, M.L.; Wooten, M.C. Use of the Open Field Maze to measure locomotor and anxiety-like behavior in mice. J. Vis. Exp. 2015, e52434. [Google Scholar] [CrossRef] [Green Version]
- Vendruscolo, L.F.; Takahashi, R.N.; Brüske, G.R.; Ramos, A. Evaluation of the anxiolytic-like effect of NKP608, a NK1-receptor antagonist, in two rat strains that differ in anxiety-related behaviors. Psychopharmacology 2003, 170, 287–293. [Google Scholar] [CrossRef]
- Ramos, A. Animal models of anxiety: Do I need multiple tests? Trends Pharmacol. Sci. 2008, 29, 493–498. [Google Scholar] [CrossRef]
- Ramos, A.; Mellerin, Y.; Mormède, P.; Chaouloff, F. A genetic and multifactorial analysis of anxiety-related behaviours in Lewis and SHR intercrosses. Behav. Brain Res. 1998, 96, 195–205. [Google Scholar] [CrossRef]
- File, S.E.; Lippa, A.S.; Beer, B.; Lippa, M.T. Animal Tests of Anxiety. In Current Protocols in Pharmacology; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2005; Volume 31, pp. 241–251. [Google Scholar]
- Dixit, P.V.; Sahu, R.; Mishra, D.K. Marble-burying behavior test as a murine model of compulsive-like behavior. J. Pharmacol. Toxicol. Methods 2020, 102, 106676. [Google Scholar] [CrossRef]
- Thomas, A.; Burant, A.; Bui, N.; Graham, D.; Yuva-Paylor, L.A.; Paylor, R. Marble burying reflects a repetitive and perseverative behavior more than novelty-induced anxiety. Psychopharmacology 2009, 204, 361–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Londei, T.; Valentini, A.M.V.; Leone, V.G. Investigative burying by laboratory mice may involve non-functional, compulsive, behaviour. Behav. Brain Res. 1998, 94, 249–254. [Google Scholar] [CrossRef]
- Gyertyán, I. Analysis of the marble burying response: Marbles serve to measure digging rather than evoke burying. Behav. Pharmacol. 1995, 6, 24–31. [Google Scholar]
- Han, S.Y.; McLennan, T.; Czieselsky, K.; Herbison, A.E. Selective optogenetic activation of arcuate kisspeptin neurons generates pulsatile luteinizing hormone secretion. Proc. Natl. Acad. Sci. USA 2015, 112, 13109–13114. [Google Scholar] [CrossRef] [Green Version]
- Pheng, V.; Uenoyama, Y.; Homma, T.; Inamoto, Y.; Takase, K.; Yoshizawa-Kumagaye, K.; Isaka, S.; Watanabe, T.X.; Ohkura, S.; Tomikawa, J.; et al. Potencies of centrally-or peripherally-injected full-length kisspeptin or its C-terminal decapeptide on LH release in intact male rats. J. Reprod. Dev. 2009, 55, 378–382. [Google Scholar] [CrossRef] [Green Version]
- Pineda, R.; Garcia-Galiano, D.; Sanchez-Garrido, M.A.; Romero, M.; Ruiz-Pino, F.; Aguilar, E.; Dijcks, F.A.; Blomenröhr, M.; Pinilla, L.; Van Noort, P.I.; et al. Characterization of the inhibitory roles of RFRP3, the mammalian ortholog of GnIH, in the control of gonadotropin secretion in the rat: In vivo and in vitro studies. Am. J. Physiol. Endocrinol. Metab. 2010, 299, 9–12. [Google Scholar] [CrossRef]
- Hu, K.L.; Chang, H.M.; Li, R.; Yu, Y.; Qiao, J. Regulation of LH secretion by RFRP-3—From the hypothalamus to the pituitary. Front. Neuroendocrinol. 2019, 52, 12–21. [Google Scholar] [CrossRef]
- Lyubimov, Y.; Engstrom, M.; Wurster, S.; Savola, J.M.; Korpi, E.R.; Panula, P. Human kisspeptins activate neuropeptide FF2 receptor. Neuroscience 2010, 170, 117–122. [Google Scholar] [CrossRef]
- Cador, M.; Marco, N.; Stinus, L.; Simonnet, G. Interaction between neuropeptide FF and opioids in the ventral tegmental area in the behavioral response to novelty. Neuroscience 2002, 110, 309–318. [Google Scholar] [CrossRef]
- Kotlinska, J.; Pachuta, A.; Dylag, T.; Silberring, J. The role of neuropeptide FF (NPFF) in the expression of sensitization to hyperlocomotor effect of morphine and ethanol. Neuropeptides 2007, 41, 51–58. [Google Scholar] [CrossRef]
- Runegaard, A.H.; Sørensen, A.T.; Fitzpatrick, C.M.; Jørgensen, S.H.; Petersen, A.V.; Hansen, N.W.; Weikop, P.; Andreasen, J.T.; Mikkelsen, J.D.; Perrier, J.F.; et al. Locomotor- and reward-enhancing effects of cocaine are differentially regulated by chemogenetic stimulation of Gi-signaling in dopaminergic neurons. eNeuro 2018, 5, e0345-17.2018. [Google Scholar] [CrossRef] [PubMed]
- Pielecka-Fortuna, J.; Moenter, S.M. Kisspeptin increases γ-aminobutyric acidergic and glutamatergic transmission directly to gonadotropin-releasing hormone neurons in an estradiol-dependent manner. Endocrinology 2010, 151, 291–300. [Google Scholar] [CrossRef] [PubMed]
- Di Giorgio, N.P.; Bizzozzero-Hiriart, M.; Libertun, C.; Lux-Lantos, V. Unraveling the connection between GABA and kisspeptin in the control of reproduction. Reproduction 2019, 157, R225–R233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.; de Jong, J.W.; Tak, Y.; Peck, J.; Bateup, H.S.; Lammel, S. Nucleus Accumbens Subnuclei Regulate Motivated Behavior via Direct Inhibition and Disinhibition of VTA Dopamine Subpopulations. Neuron 2018, 97, 434–449.e4. [Google Scholar] [CrossRef] [PubMed]
- Gottsch, M.L.; Cunningham, M.J.; Smith, J.T.; Popa, S.M.; Acohido, B.V.; Crowley, W.F.; Seminara, S.; Clifton, D.K.; Steiner, R.A. A role for kisspeptins in the regulation of gonadotropin secretion in the mouse. Endocrinology 2004, 145, 4073–4077. [Google Scholar] [CrossRef] [Green Version]
- Rønnekleiv, O.K.; Qiu, J.; Kelly, M.J. Arcuate Kisspeptin Neurons Coordinate Reproductive Activities with Metabolism. Semin. Reprod. Med. 2019, 37, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Murase, T.; Arima, H.; Kondo, K.; Oiso, Y. Neuropeptide FF reduces food intake in rats. Peptides 1996, 17, 353–354. [Google Scholar] [CrossRef]
- Desprat, C.; Zajac, J.M. Hypothermic effects of neuropeptide FF analogues in mice. Pharmacol. Biochem. Behav. 1997, 58, 559–563. [Google Scholar] [CrossRef]
- Moulédous, L.; Barthas, F.; Zajac, J.M. Opposite control of body temperature by NPFF1 and NPFF2 receptors in mice. Neuropeptides 2010, 44, 453–456. [Google Scholar] [CrossRef]
- Ten, S.C.; Gu, S.Y.; Niu, Y.F.; An, X.F.; Yan, M.; He, M. Central administration of kisspeptin-10 inhibits water and sodium excretion of anesthetized male rats and the involvement of arginine vasopressin. Endocr. Res. 2010, 35, 128–136. [Google Scholar] [CrossRef]
- Tsunematsu, T.; Fu, L.-Y.; Yamanaka, A.; Ichiki, K.; Tanoue, A.; Sakurai, T.; van den Pol, A.N. Vasopressin Increases Locomotion through a V1a Receptor in Orexin/Hypocretin Neurons: Implications for Water Homeostasis. J. Neurosci. 2008, 28, 228–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arai, A.C.; Orwig, N. Factors that regulate KiSS1 gene expression in the hippocampus. Brain Res. 2008, 1243, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Krizo, J.A.; Moreland, L.E.; Rastogi, A.; Mou, X.; Prosser, R.A.; Mintz, E.M. Regulation of Locomotor activity in fed, fasted, and food-restricted mice lacking tissue-type plasminogen activator. BMC Physiol. 2018, 18, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ibos, K.E.; Bodnár, É.; Bagosi, Z.; Bozsó, Z.; Tóth, G.; Szabó, G.; Csabafi, K. Kisspeptin-8 Induces Anxiety-Like Behavior and Hypolocomotion by Activating the HPA Axis and Increasing GABA Release in the Nucleus Accumbens in Rats. Biomedicines 2021, 9, 112. https://doi.org/10.3390/biomedicines9020112
Ibos KE, Bodnár É, Bagosi Z, Bozsó Z, Tóth G, Szabó G, Csabafi K. Kisspeptin-8 Induces Anxiety-Like Behavior and Hypolocomotion by Activating the HPA Axis and Increasing GABA Release in the Nucleus Accumbens in Rats. Biomedicines. 2021; 9(2):112. https://doi.org/10.3390/biomedicines9020112
Chicago/Turabian StyleIbos, Katalin Eszter, Éva Bodnár, Zsolt Bagosi, Zsolt Bozsó, Gábor Tóth, Gyula Szabó, and Krisztina Csabafi. 2021. "Kisspeptin-8 Induces Anxiety-Like Behavior and Hypolocomotion by Activating the HPA Axis and Increasing GABA Release in the Nucleus Accumbens in Rats" Biomedicines 9, no. 2: 112. https://doi.org/10.3390/biomedicines9020112