
Identification of New Dioxygenases Able to Recognize Polycyclic Aromatic Hydrocarbons with High Aromaticity



Abstract
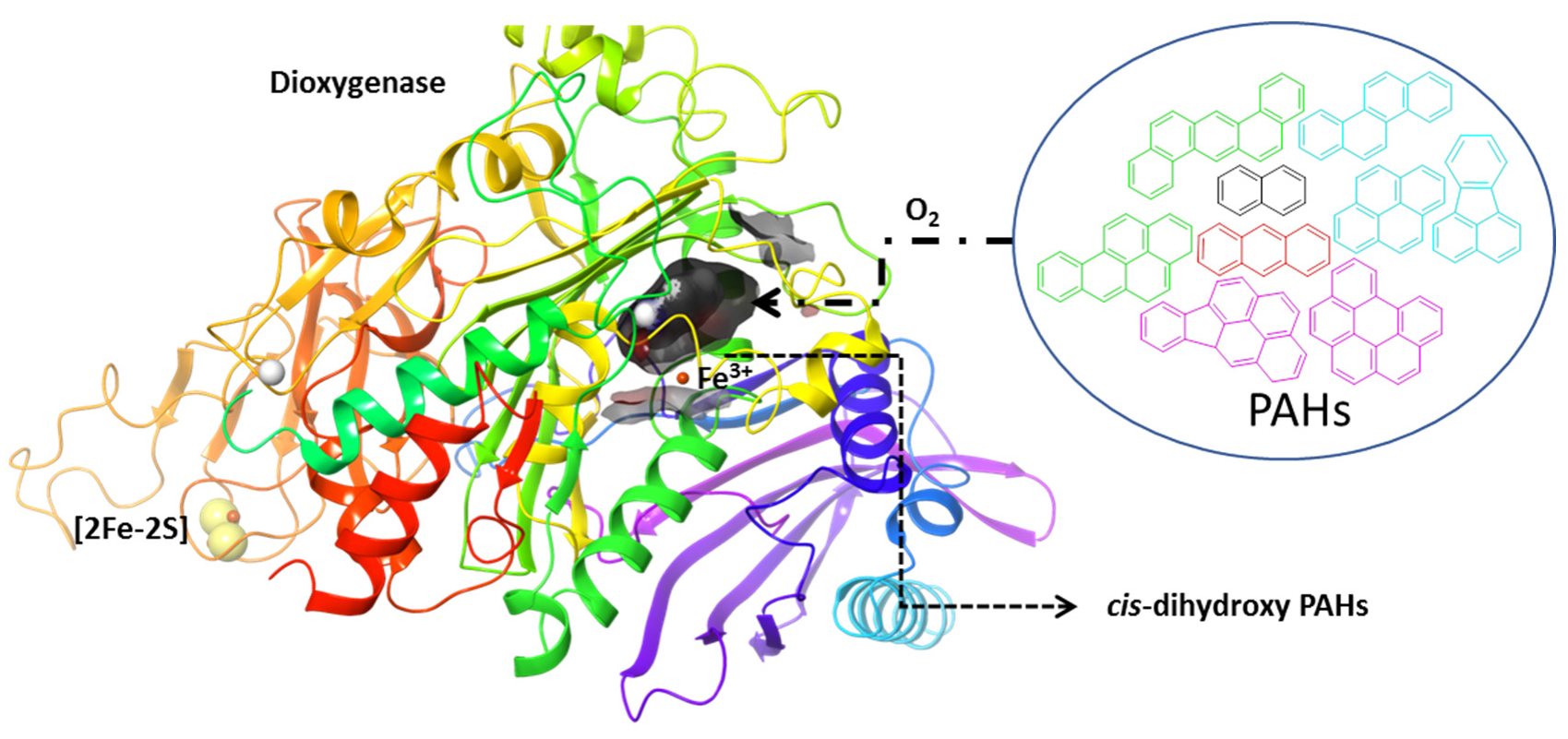
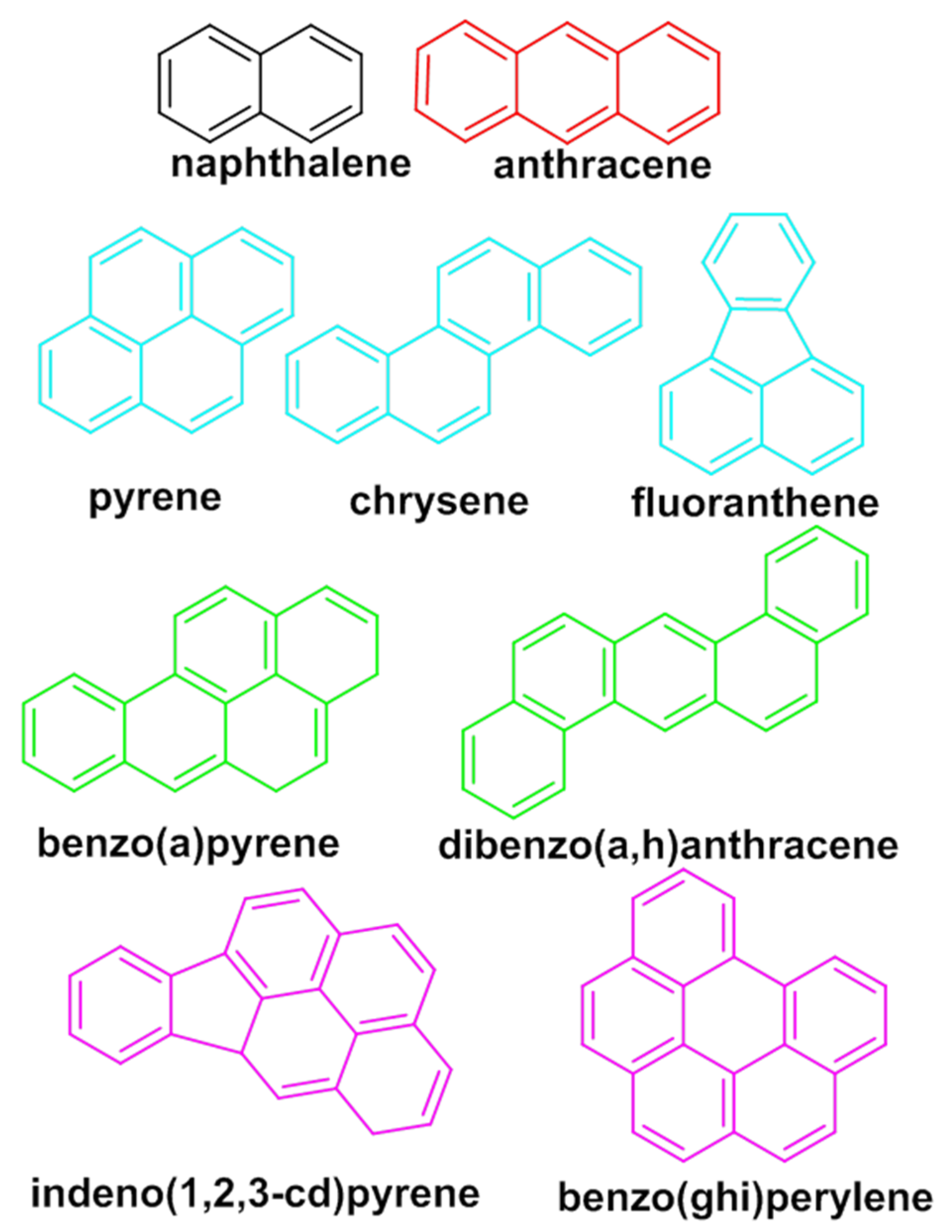
:1. Introduction
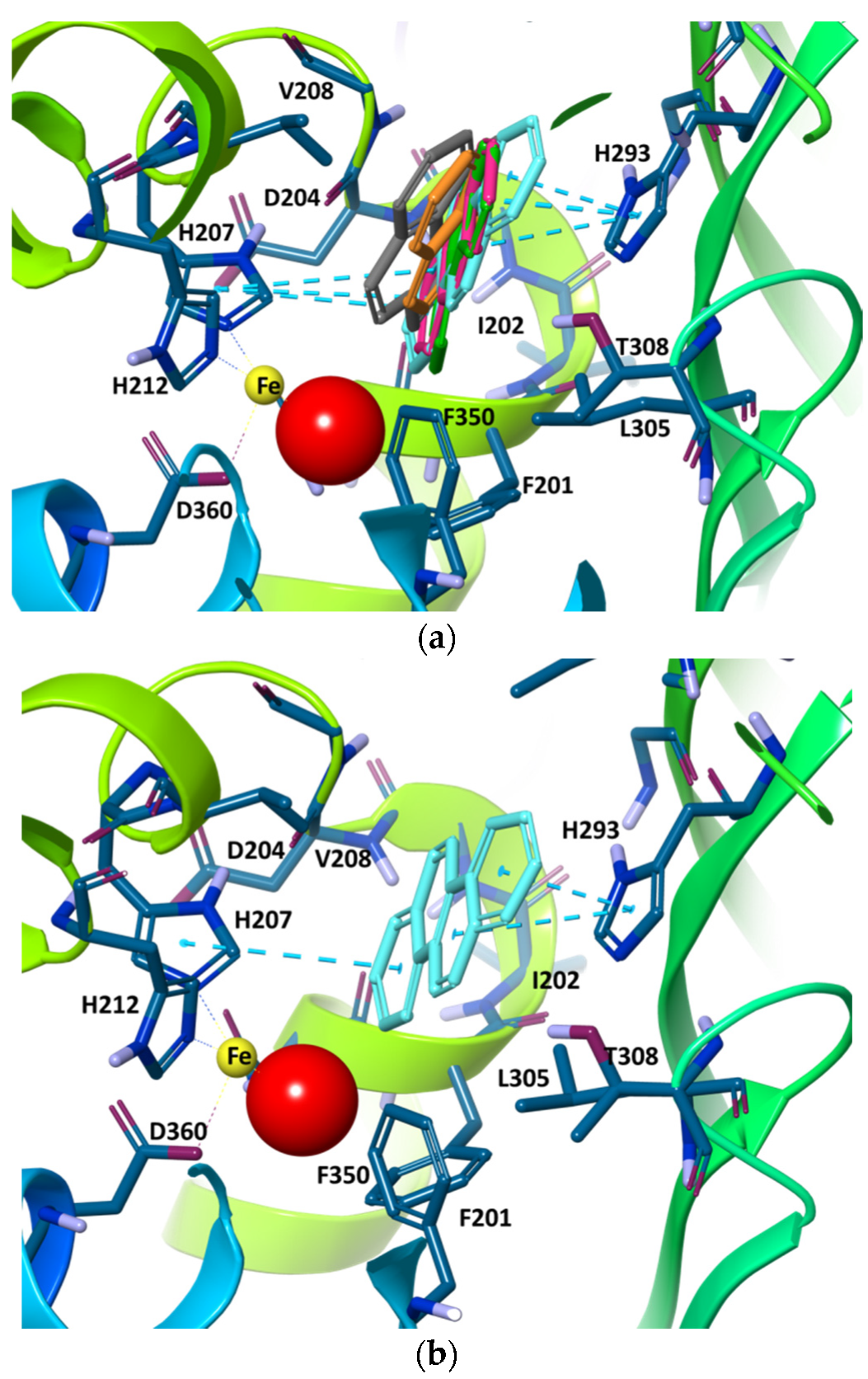
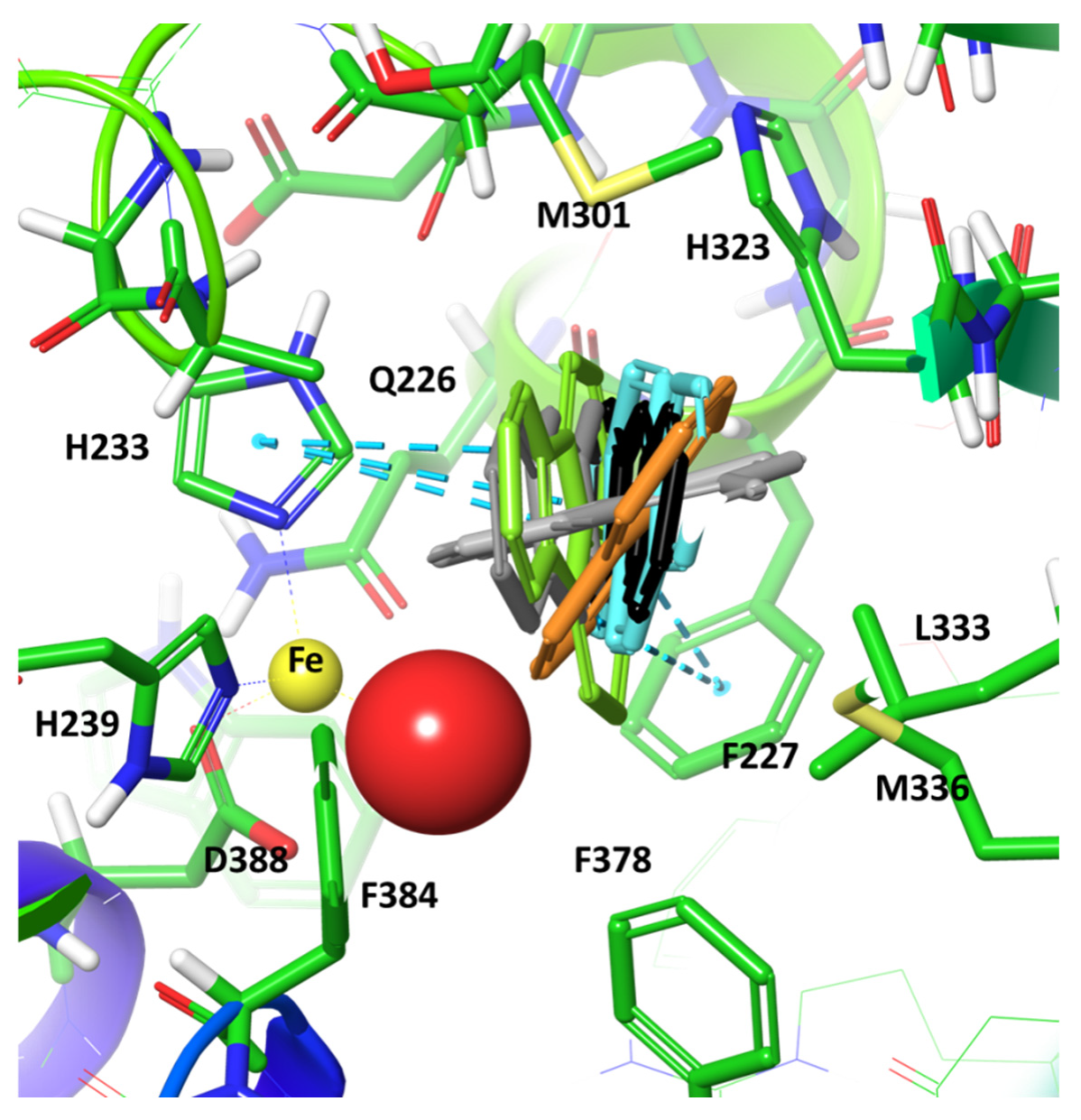
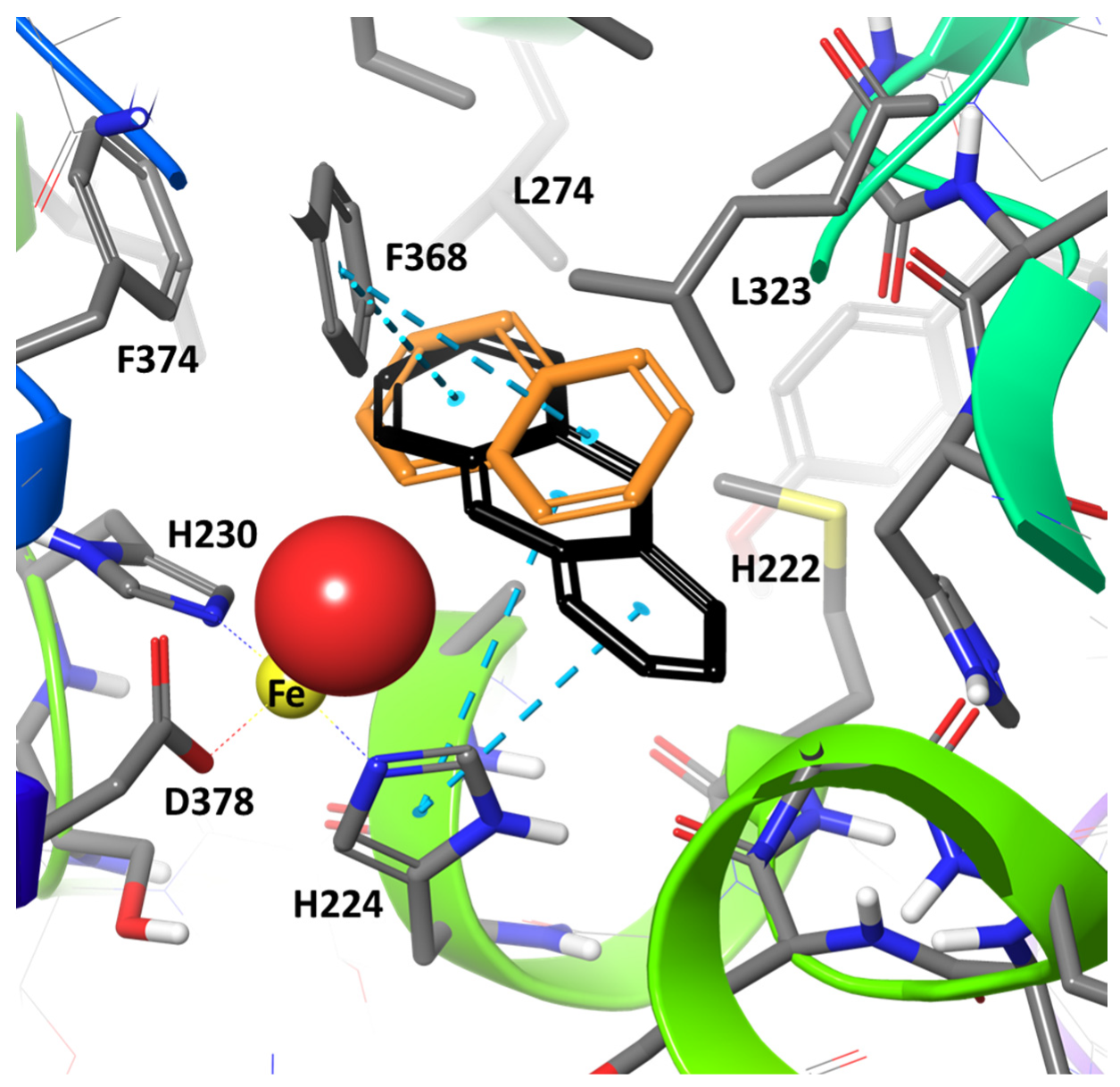
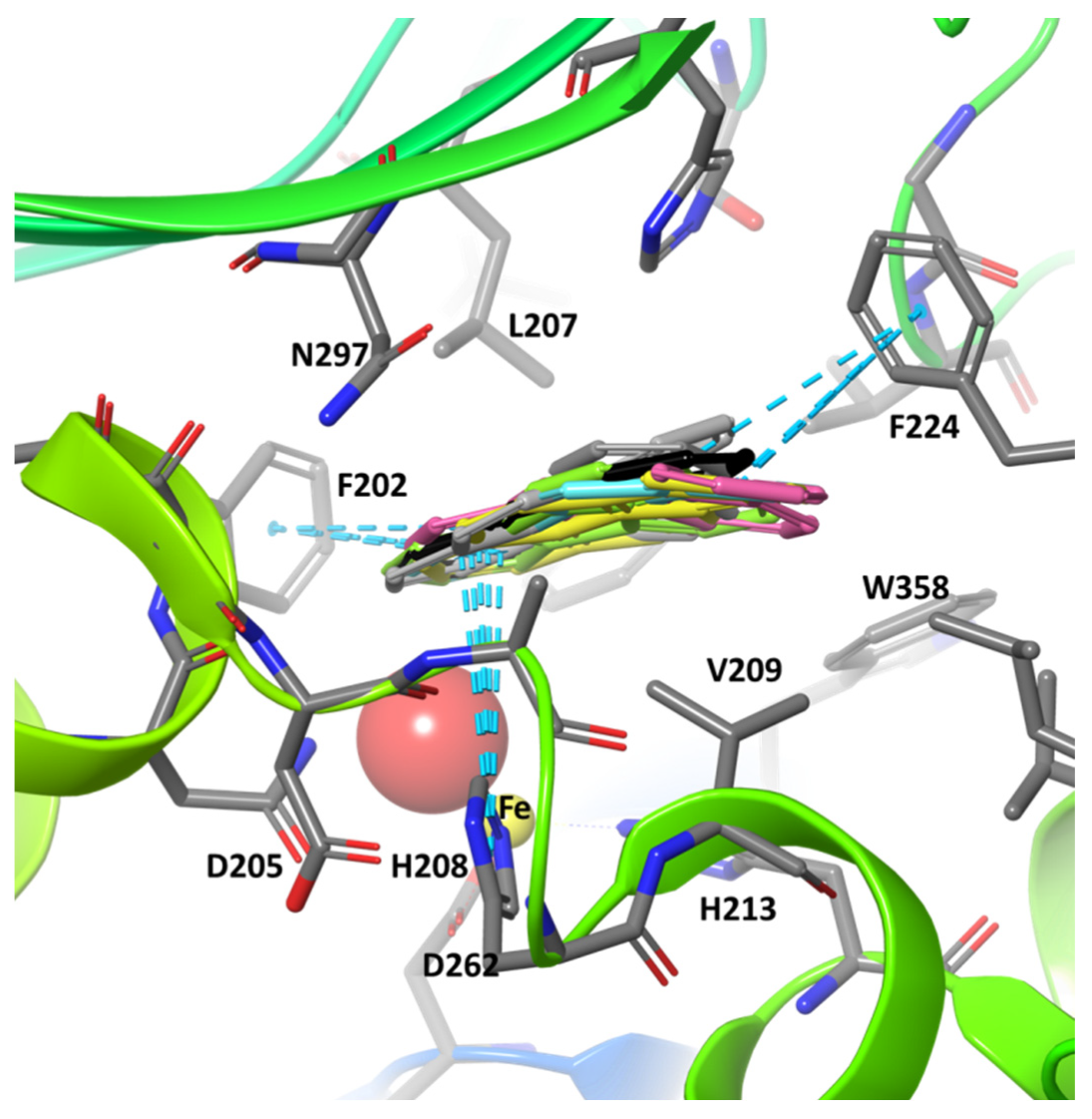
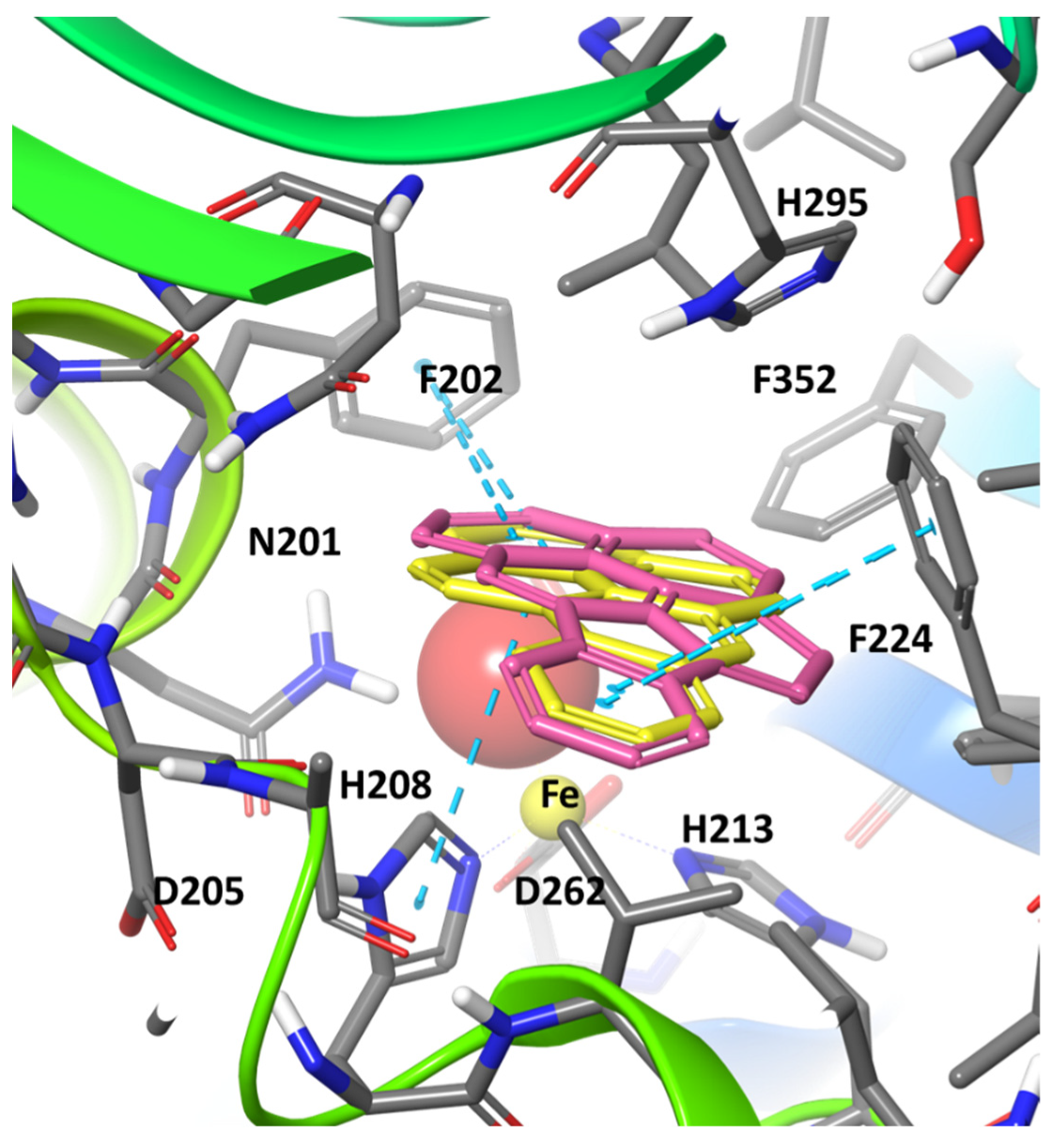
2. Results and Discussion
3. Materials and Methods
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Sahoo, B.M.; Ravi Kumar, B.V.V.; Banik, B.K.; Borah, P. Polyaromatic Hydrocarbons (PAHs): Structures, Synthesis and their Biological Profile. Curr. Org. Synth. 2020, 17, 625–640. [Google Scholar] [CrossRef]
- Patel, A.B.; Shaikh, S.; Jain, K.R.; Desai, C.; Madamwar, D. Polycyclic Aromatic Hydrocarbons: Sources, Toxicity, and Remediation Approaches. Front. Microbiol. 2020, 11, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Shafy, H.I.; Mansour, M.S.M. A review on polycyclic aromatic hydrocarbons: Source, environmental impact, effect on human health and remediation. Egypt. J. Pet. 2016, 25, 107–123. [Google Scholar] [CrossRef] [Green Version]
- Cerniglia, C.E. Biodegradation of polycyclic aromatic hydrocarbons. Biodegradation 1992, 3, 351–368. [Google Scholar] [CrossRef]
- Juhasz, A.L.; Naidu, R. Bioremediation of high molecular weight polycyclic aromatic hydrocarbons: A review of the microbial degradation of benzo[a]pyrene. Int. Biodeterior. Biodegrad. 2000, 45, 57–88. [Google Scholar] [CrossRef]
- Viñas, M.; Solanas, A. Biorremediación de Suelos Contaminados por Hidrocarburos: Caracterización Microbiológica, Química y Ecotoxicológica. Ph.D. Thesis, Universitat de Barcelona, Barcelona, Spain, 2005. [Google Scholar]
- Desai, A.M.; Autenrieth, R.L.; Dimitriou-Christidis, P.; McDonald, T.J. Biodegradation kinetics of select polycyclic aromatic hydrocarbon (PAH) mixtures by Sphingomonas paucimobilis EPA505. Biodegradation 2008, 19, 223–233. [Google Scholar] [CrossRef]
- Vaidya, S.; Devpura, N.; Jain, K.; Madamwar, D. Degradation of chrysene by enriched bacterial consortium. Front. Microbiol. 2018, 9, 1–14. [Google Scholar] [CrossRef]
- Rodríguez-gonzales, A.; Zárate-villarroel, S.G.; Bastida-codina, A. Biodiversidad Bacteriana Presente en Suelos Contaminados con Hidrocarburos Para Realizar Biorremediación; Universidad Nacional de Costa Rica: Heredia, Costa Rica, 2022; Volume 56, ISBN 0000000251419. [Google Scholar]
- Roszak, M.; Jabłó Nska, J.; Stachurska, X.; Dubrowska, K.; Kajdanowicz, J.; Goł˛ Ebiewska, M.; Kiepas-Kokot, A.; Osí Nska, B.; Augustyniak, A.; Karakulska, J. Development of an Autochthonous Microbial Consortium for Enhanced Bioremediation of PAH-Contaminated Soil. Int. J. Mol. Sci. Artic. 2021, 22, 13469. [Google Scholar] [CrossRef]
- Thomas, S.; Veettil, N.T.; Subbiah, K. Isolation, characterization and optimization of chrysene degradation using bacteria isolated from oil-contaminated water. Water Sci. Technol. 2021, 84, 2737–2748. [Google Scholar] [CrossRef]
- Morillo, E.; Madrid, F.; Lara-Moreno, A.; Villaverde, J. Soil bioremediation by cyclodextrins. A review. Int. J. Pharm. 2020, 591, 1–16. [Google Scholar] [CrossRef]
- Jouanneau, Y.; Meyer, C.; Duraffourg, N. Dihydroxylation of four- and five-ring aromatic hydrocarbons by the naphthalene dioxygenase from Sphingomonas CHY-1. Appl. Microbiol. Biotechnol. 2016, 100, 1253–1263. [Google Scholar] [CrossRef] [PubMed]
- Mutnuri, S.; Vasudevan, N.; Matthias, K. Degradation of anthracene and pyrene supplied by microcrystals and non-aqueous-phase liquids. Appl. Microbiol. Biotechnol. 2005, 67, 569–576. [Google Scholar] [CrossRef] [PubMed]
- Kanaly, R.A.; Harayama, S. Biodegradation of high-molecular-weight polycyclic aromatic hydrocarbons by bacteria. J. Bacteriol. 2000, 182, 2059–2067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakoncic, J.; Jouanneau, Y.; Meyer, C.; Stojanoff, V. The catalytic pocket of the ring-hydroxylating dioxygenase from Sphingomonas CHY-1. Biochem. Biophys. Res. Commun. 2007, 352, 861–866. [Google Scholar] [CrossRef] [Green Version]
- Wackett, L.P. Mechanism and applications of Rieske non-heme iron dioxygenases. Enzym. Microb. Technol. 2002, 31, 577–587. [Google Scholar] [CrossRef]
- Boyd, D.R.; Sharma, N.D.; Brannigan, I.N.; McGivern, C.J.; Nockemann, P.; Stevenson, P.J.; McRoberts, C.; Hoering, P.; Allen, C.C.R. Cis-Dihydroxylation of Tricyclic Arenes and Heteroarenes Catalyzed by Toluene Dioxygenase: A Molecular Docking Study and Experimental Validation. Adv. Synth. Catal. 2019, 361, 2526–2537. [Google Scholar] [CrossRef] [Green Version]
- Kauppi, B.; Lee, K.; Carredano, E.; Parales, R.E.; Gibson, D.T.; Eklund, H.; Ramaswamy, S. Structure of an aromatic-ring-hydroxylating dioxygenasenaphthalene 1,2-dioxygenase. Structure 1998, 6, 571–586. [Google Scholar] [CrossRef] [Green Version]
- Mahto, J.K.; Neetu, N.; Waghmode, B.; Kuatsjah, E.; Sharma, M.; Sircar, D.; Sharma, A.K.; Tomar, S.; Eltis, L.D.; Kumar, P. Molecular insights into substrate recognition and catalysis by phthalate dioxygenase from Comamonas testosteroni. J. Biol. Chem. 2021, 297, 101416. [Google Scholar] [CrossRef]
- Ferraro, D.J.; Okerlund, A.L.; Mowers, J.C.; Ramaswamy, S. Structural basis for regioselectivity and stereoselectivity of product formation by naphthalene 1,2-dioxygenase. J. Bacteriol. 2006, 188, 6986–6994. [Google Scholar] [CrossRef] [Green Version]
- Ferraro, D.J.; Gakhar, L.; Ramaswamy, S. Rieske business: Structure-function of Rieske non-heme oxygenases. Biochem. Biophys. Res. Commun. 2005, 338, 175–190. [Google Scholar] [CrossRef]
- Vila, M.A.; Umpiérrez, D.; Seoane, G.; Rodríguez, S.; Carrera, I.; Veiga, N. Computational insights into the oxidation of mono- and 1,4 disubstituted arenes by the Toluene Dioxygenase enzymatic complex. J. Mol. Catal. B Enzym. 2016, 133, S410–S419. [Google Scholar] [CrossRef]
- Jouanneau, Y.; Micoud, J.; Meyer, C. Purification and characterization of a three-component salicylate 1-hydroxylase from Sphingomonas sp. strain CHY-1. Appl. Environ. Microbiol. 2007, 73, 7515–7521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuler, L.; Jouanneau, Y.; Ní Chadhain, S.M.; Meyer, C.; Pouli, M.; Zylstra, G.J.; Hols, P.; Agathos, S.N. Characterization of a ring-hydroxylating dioxygenase from phenanthrene-degrading Sphingomonas sp. strain LH128 able to oxidize benz[a]anthracene. Appl. Microbiol. Biotechnol. 2009, 83, 465–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goris, J.; De Vos, P.; Caballero-Mellado, J.; Park, J.; Falsen, E.; Quensen, J.F.; Tiedje, J.M.; Vandamme, P. Classification of the biphenyl- and polychlorinated biphenyl-degrading strain LB400T and relatives as Burkholderia xenovorans sp. nov. Int. J. Syst. Evol. Microbiol. 2004, 54, 1677–1681. [Google Scholar] [CrossRef] [Green Version]
- Auffret, M.; Labbé, D.; Thouand, G.; Greer, C.W.; Fayolle-Guichard, F. Degradation of a mixture of hydrocarbons, gasoline, and diesel oil additives by Rhodococcus aetherivorans and Rhodococcus wratislaviensis. Appl. Environ. Microbiol. 2009, 75, 7774–7782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olowomofe, T.O.; Oluyege, J.O.; Aderiye, B.I.; Oluwole, O.A. Degradation of poly aromatic fractions of crude oil and detection of catabolic genes in hydrocarbon-degrading bacteria isolated from Agbabu bitumen sediments in Ondo State. AIMS Microbiol. 2019, 5, 308–323. [Google Scholar] [CrossRef]
- Labute, P. Protonate3D: Assignment of ionization states and hydrogen coordinates to macromolecular structures. Proteins Struct. Funct. Bioinform. 2009, 75, 187–205. [Google Scholar] [CrossRef] [Green Version]
- Madhavi Sastry, G.; Adzhigirey, M.; Day, T.; Annabhimoju, R.; Sherman, W. Protein and ligand preparation: Parameters, protocols, and influence on virtual screening enrichments. J. Comput. Aided. Mol. Des. 2013, 27, 221–234. [Google Scholar] [CrossRef]
- Friesner, R.A.; Murphy, R.B.; Repasky, M.P.; Frye, L.L.; Greenwood, J.R.; Halgren, T.A.; Sanschagrin, P.C.; Mainz, D.T. Extra precision glide: Docking and scoring incorporating a model of hydrophobic enclosure for protein-ligand complexes. J. Med. Chem. 2006, 49, 6177–6196. [Google Scholar] [CrossRef] [Green Version]
- Warren, G.L.; Andrews, C.W.; Capelli, A.M.; Clarke, B.; LaLonde, J.; Lambert, M.H.; Lindvall, M.; Nevins, N.; Semus, S.F.; Senger, S.; et al. A critical assessment of docking programs and scoring functions. J. Med. Chem. 2006, 49, 5912–5931. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code (PDB) | Organism | Dioxygenase/ Uniprot No. | Ions/Ligand |
|---|---|---|---|
| 2ckf 2gbw 2gbx 2i7f | Sphingomonas sp. CHY-1 (Gram+) Sphingobium yanoikuyae (Gram+) | Naphthalene 1,2- Q65AT1 Biphenyl 2,3- /A2TC87 | Fe2S2, Fe3+ ---- Biphenyl |
| 2b1x 2b24 | Rhodococcus sp. NCIMB 12038 (Gram-) | Naphthalene Q9 × 3r9 | Fe2S2, Fe3+ Indol |
| 1ulj 1uli | Rhodococcus jostii RHA1 (Gram-) | Biphenyl 2,3 | Fe2S2, Fe3+ Biphenyl |
| 1eg9 1ndo 1o7n 1o7p 3en1 3eqq | Pseudomonas putida (Gram-) | Naphthalene 1,2 P0A110 Toluene 2,3-/A5W4F2 | Fe2S2, Fe3+, SO4− Indole Toluene ---- |
| 4hjl + 4hkv | Pseudomonas sp. C18 (Gram-) | Naphthalene 1,2-/P0A111 | Fe2S2, Fe3+, SO4− Naphthalene Benzamide |
| 2xsh 2xrx 2xso 2xsh | Paraburkholderia xenovorans LB400 (Gram-) | Biphenyl/ P37,334 | Fe2S2, Fe3+, SO4− Biphenyl --- |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodríguez, A.; Zárate, S.G.; Bastida, A. Identification of New Dioxygenases Able to Recognize Polycyclic Aromatic Hydrocarbons with High Aromaticity. Catalysts 2022, 12, 279. https://doi.org/10.3390/catal12030279
Rodríguez A, Zárate SG, Bastida A. Identification of New Dioxygenases Able to Recognize Polycyclic Aromatic Hydrocarbons with High Aromaticity. Catalysts. 2022; 12(3):279. https://doi.org/10.3390/catal12030279
Chicago/Turabian StyleRodríguez, Apolonia, Sandra G. Zárate, and Agatha Bastida. 2022. "Identification of New Dioxygenases Able to Recognize Polycyclic Aromatic Hydrocarbons with High Aromaticity" Catalysts 12, no. 3: 279. https://doi.org/10.3390/catal12030279
APA StyleRodríguez, A., Zárate, S. G., & Bastida, A. (2022). Identification of New Dioxygenases Able to Recognize Polycyclic Aromatic Hydrocarbons with High Aromaticity. Catalysts, 12(3), 279. https://doi.org/10.3390/catal12030279