3. Results and Discussion
Figure 1 shows the DICM images of the surface morphology of the (000
) facet of the two-inch diameter (a–1) nominally undoped (boule A), (b) conventionally nitrogen-doped (boule B), and (c) heavily nitrogen-doped 4H-SiC boules (boule C) examined in this study. The figure reveals that the growth front of the boules comprised two distinct morphological regions: the (000
) facet and the non-facetted region.
On the (000
) facet of the nominally undoped boule (boule A), large macrosteps of height of sub-μm range were observed, and the macrosteps tended to be arranged in petal shapes with a hexagonal symmetry. The height of the macrosteps became larger as their location approached the edge of the (000
) facet, as confirmed by LVSEM (see
Figure 1(a–2)), and a giant slope was found at the boundary between the (000
) facet and the non-facetted region. Between the macrosteps, fairly flat, almost featureless terraces extended. For sample B, the (000
) facet also appeared petal-shaped, but no macrosteps of sub-μm height were observed on the facet. The boundary between the (000
) facet and the non-facetted region was relatively smooth, and an intermediate region was present between the facet and the non-facetted region [
11]. The facet surface of boule B showed hexagonal symmetry, comprising six vicinal (000
) surfaces slightly tilted toward <1
00> directions, i.e., prism plane directions, accompanied by six crystallographically-equivalent ridges extending along the <11
0> directions, and the vicinal (000
) surface exhibited a slightly undulated morphology. On the other hand, the (000
) facet surface of the heavily nitrogen-doped 4H-SiC boule (boule C) was extremely flat. The surface was almost featureless (with no macrosteps and undulations), and the boundary between the facet and the non-facetted region was obscure. As seen in the figure, a number of very small dots was observed on the almost featureless facet surface of sample C. They are hillocks nucleated during the cooling stage of PVT growth process [
17]. Their formation only occurs on the (000
) facet of heavily nitrogen-doped 4H-SiC boules and is thought to be related to the stacking fault formation in heavily nitrogen-doped 4H-SiC crystals [
17].
To reveal the microscopic origin of the observed macroscopic surface morphologies on the (000
) facet of 4H-SiC boules, we conducted AFM observations of the step structure on the facet. First, we examined the step structure in the edge region of the (000
) facet of three differently nitrogen-doped 4H-SiC boules (boules A, B, and C). The locations of the AFM observations are indicated as Points A, B, and C in
Figure 1 for boules A, B, and C, respectively. The locations were determined in such a way that the tilt angles from the basal plane of each location, which was measured by CLSM, were almost the same (0.3°–0.8°).
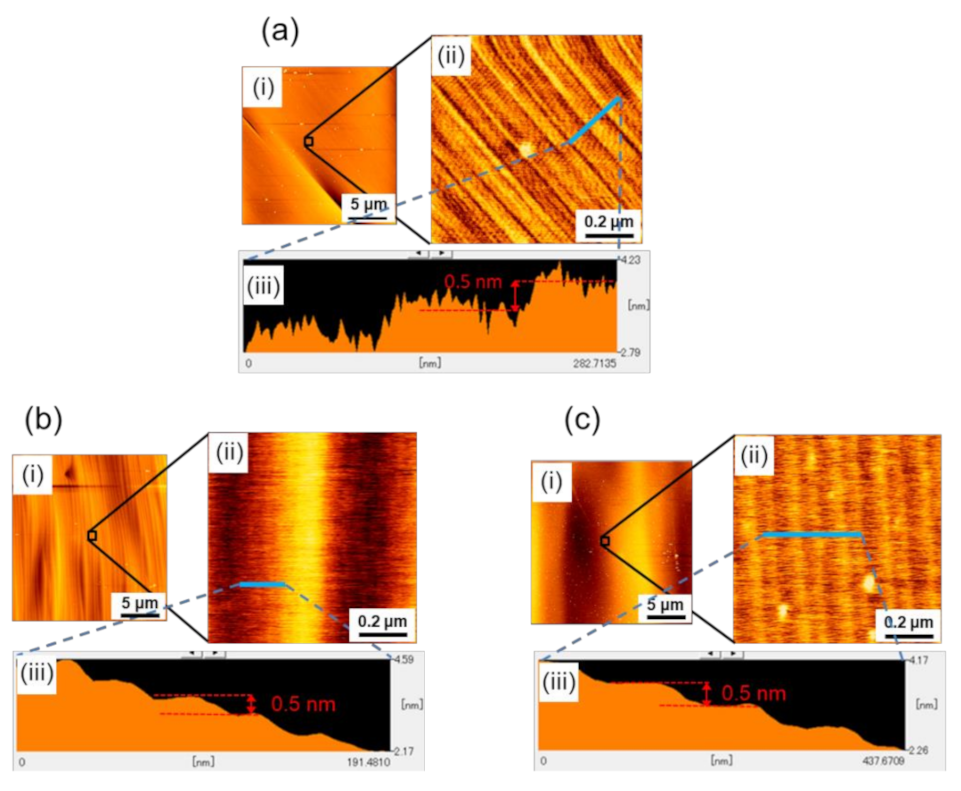
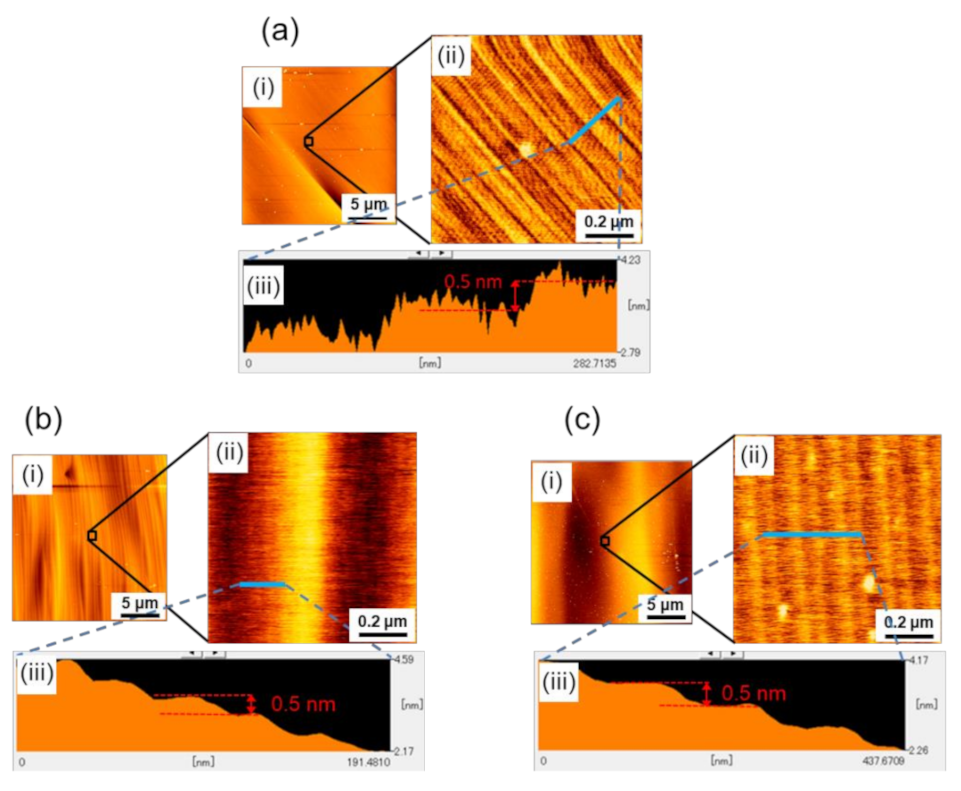
Figure 2a–c show the results of the AFM observations at Points A, B, and C, respectively. In each figure, (i) a wide-area AFM image (20-μm square area), (ii) a small-area image (0.9-μm square area), and (iii) the height profile are shown, where the small area AFM image was taken from the area indicated by an open square in (i), and the height profile was taken along the blue line indicated in (ii). As seen in the figures, microscopically, the facet surfaces of all three boules were covered with steps of half unit-cell height (0.5 nm), but their configurations were slightly different; the white spots observed in some of the AFM images are particles of, what was presumed to be dust. Difference was observed in the non-uniformity of the step separation (terrace width); there was a characteristic undulation of step separation for each boule. Examples of such an undulation of step separation are clearly demonstrated in
Figure 2(bii,ci), where periodically corrugated structures of the step trains were observed. The small area AFM images (
Figure 2(bii,cii)) revealed that these corrugated structures consisted of half unit-cell height steps and were caused by their separation modulation.
As seen in the figures, the wavelength of the undulation was different among the boules, indicating that the wavelength of the undulation was dependent on the doping concentration of the nitrogen donors in the boules.
Table 1 summarizes the nitrogen doping concentration dependence of the undulation wavelength of the step separation, and
Figure 3 shows schematic figures of the separation-undulated step trains on a vicinal surface, where the undulation wavelength of step separation (terrace width) increases from
Figure 3a–c.
We could not observe any undulation in the step separation on the (000) facet edge of boule A (the nominally undoped boule), to which we assigned an undulation wavelength of <70 nm, comparable to or less than the step separation. As shown in the table, as the nitrogen doping concentration increased (from boule A to boule B to boule C), the undulation wavelength significantly increased.
To shed more light on the observed undulated step structure near the facet edge, we examined the step structure in the central region (near a spiral growth center) of the (000
) facet.
Figure 4a–c show the AFM images of the step structures observed at Points A’, B’, and C’ on the facets of boules A, B, and C, respectively, shown in
Figure 1. In each figure, a set of wide-area AFM images (50-μm square or 30-μm square area) and small-area images (25-μm square or 10-μm square area) are shown; the location of the small-area AFM image is indicated by a blue-line open square shown in the wide-area image. As shown in the figures, the facet surface in the central region was also covered with half unit-cell height step trains, which were also undulated at a certain wavelength. The step separation and the undulation wavelength of the step separation in the central region of the (000
) facet of boules A, B, and C are summarized in
Table 1, where the wavelength of boule A could not be measured, since the step separation was significantly large. For boule B, undulated step structures were observed in very limited areas in the central region of the facet. We carefully examined the central region of the facet of boule B and found that undulated step structures were rarely observed in the region. As seen in the table, the undulation wavelength of the step separation was very similar for boules B and C and appeared not to be dependent on the nitrogen doping concentration.
As Yamaguchi et al. [
11] pointed out, on the (000
) facet of 4H-SiC crystals, the step separation is larger in the central region and becomes smaller toward the facet edge, according to the variation of undercooling over the facet. This trend was also observed for boules A, B, and C examined in this study, as shown in
Table 1. They contained different concentrations of nitrogen donors but exhibited a similar variation of step separation from the central to the edge regions of the (000
) facet. The step separation was similar among boules A, B, and C except for the central region of the facet for boule A (N: mid-10
17 cm
−3), which showed a step separation approximately 30 times larger than those observed for boules B and C. The large step separation in the central region of boule A would have resulted from the formation of macrosteps of sub-μm height on the facet.
In
Table 1, we also present the meandering wavelength of the half unit-cell height steps observed in the central region of the (000
) facet of boules A, B, and C. On the facet of boule C, the half unit-cell height steps were fairly straight; thus, the meandering of the surface steps was assumed to be fully suppressed in the central region of boule C.
We assume that the observed undulation of step separation is a precursor state of step bunching on a vicinal surface [
18]. Step bunching is generally thought to have a kinematical origin, and the most widely accepted mechanism is the asymmetric incorporation kinetics of adatoms at surface steps. A well-known form of asymmetric incorporation kinetics is the so-called Ehrlich-Schwoebel (ES) effect, which assumes the preferential incorporation of adatoms to steps from the lower terrace [
19,
20]; a large kinetic barrier is assumed for adatoms to descend the step edge to the lower terrace. This form of asymmetric kinetics, however, causes step bunching only for negative crystal growth, i.e., the etching of crystal. Hence, to induce step bunching for crystal growth, an inverse ES effect needs to be considered [
21,
22,
23], in which a larger kinetic barrier exists for the adatom incorporation from the lower terrace; thus, adatoms are preferentially incorporated to steps from the upper terrace.
The inverse ES effect has already been proposed to explain the coexistence of step bunching and meandering on the 4H-SiC(0001) surface [
21]. However, its validity for the 4H-SiC(000
) surface is yet to be proven. In this regard, an important requirement to justify the model is that the model can explain the experimental results regarding the undulation wavelength of step separation on the 4H-SiC (000
) facet. One important conclusion for the observed undulation of step separation (step bunching) on the (000
) facet of nitrogen-doped 4H-SiC boules is that the undulation wavelength became longer when the nitrogen doping concentration increased.
As described earlier in this section, the nitrogen doping concentration also affects the line profile of surface steps. The AFM images shown in
Figure 4 indicate that the meandering wavelength of the half unit-cell height steps became shorter as the nitrogen doping concentration increased, except for in boule C; we discuss the reason why fairly straight steps were observed on the (000
) facet of boule C below.
Step meandering generally occurs through the competition between the kinematical (destabilizing) and energetic (stabilizing) effects on the step morphology [
24]; the former induces step meandering, whereas the latter stabilizes the straight-line morphology of the surface steps. Here, an important parameter for the energetic effect is the line tension of the steps, i.e., the step stiffness. The step stiffness is the measure of resistance against the kinematical driving force for step meandering and determines the meandering wavelength of the surface steps [
24]; the larger the step stiffness, the longer the meandering wavelength. Hence, the results of the AFM observations shown in
Figure 4 indicate that by some mechanism, nitrogen doping of 4H-SiC crystals reduces the step stiffness on the (000
) surface, making the meandering wavelength shorter as the nitrogen doping concentration increases. The macroscopic facet morphologies observed for boules A, B, and C lend support to this conclusion. As shown in
Figure 1, the facet morphology of the nitrogen-doped 4H-SiC crystals became more isotropic and smoother as the nitrogen doping concentration increased, indicating that energetics (step stiffness), which usually featured a preferred step flow direction reflecting the crystal symmetry, did not greatly influence the facet morphology at a high nitrogen doping concentration.
Usually, a small step stiffness results in a largely meandering step morphology on the growing crystal surface; however, the half unit-cell height steps observed on the (000
) facet of boule C, which were assumed to have a small step stiffness, showed a fairly straight step morphology. This was due to the enhanced diffusion length of the adatoms on the (000
) facet of boule C. As we discuss later in this study, heavy nitrogen doping modified the bonding structure of the 4H-SiC (000
) surface, leading to the enhancement of the diffusion length of the surface adatoms on the growing crystal surface and, consequently, suppressing the step meandering in spite of the small step stiffness [
24].
The influence of the step stiffness on the step bunching behavior was investigated by Sato and Uwaha [
25]. They theoretically investigated the instability of step trains during negative crystal growth (sublimation), assuming an ES-type asymmetric incorporation kinetics of adatoms to the steps. Their calculation took into consideration the step stiffness through the step repulsive interaction. A larger step stiffness gives rise to a larger elastic repulsion interaction between surface steps. They successfully demonstrated step bunching (undulation of step separation) with an asymmetric incorporation kinetics, and their results indicated that the larger the step repulsive interaction, the longer the undulation wavelength. This trend is completely opposite to our experimental results, according to which the undulation wavelength became longer when the step interaction (step stiffness) was reduced by nitrogen doping.
To address this problem, we need to consider another mechanism that causes step bunching during crystal growth. A plausible mechanism is the so-called “two-particle” model proposed by Pimpinelli et al. [
26,
27,
28,
29,
30,
31], which is based on the coupling between the surface densities of diffusing precursor molecules α and of adatoms β, in the presence of the ES barrier at the step edges. The adatoms β are constituents of the crystal and contribute directly to the crystal growth. The precursor molecules α dissociate (crack) on the growing crystal surface, yielding the adsorbed adatoms β on the surface, which contribute indirectly to crystal growth. This is the situation occurring during the PVT growth of SiC; crystal growth occurs through the cracking of precursor molecules such as SiC
2 [
32]. Therefore, the model would fit the PVT growth of SiC. Pimpinelli et al. showed that the presence of the ES barrier for the precursor molecules α causes adatom flows in the down-step direction, leading to the conversion of equidistant step trains into separation-undulated step trains (step bunching), even when the adatoms β possess a larger kinetic barrier to descend the steps to the lower terrace compared to when ascending the steps. The two-particle model is schematically illustrated in
Figure 5, where the precursor molecules α are depicted as a combination of red and green spheres, whereas the adatoms β are depicted as red spheres, which are generated through the cracking of the precursor molecules α on the growing crystal surface.
If the adatoms do not show asymmetric kinetics for their incorporation to the steps, their distribution is symmetrical on the terraces, as in the distribution schematically illustrated by the red solid line in
Figure 5a. The two-particle model assumes the presence of a large ES barrier for the precursor molecules α to descend the steps from the upper terrace, while they could ascend the steps from the lower terrace relatively easily. This asymmetrical form of migration kinetics significantly modifies the distribution of the precursor molecules on the terrace, and the molecules are preferentially found near the terrace edge in the down-step direction, as schematically illustrated by the green solid line in
Figure 5a. In the two-particle model, the adatoms β, which constitute the crystal, are provided by the cracking of the precursor molecules; thus, the asymmetrical distribution of the precursor molecules induces the asymmetrical distribution of the adatoms β. The distributions of the precursor molecules and adatoms on the terrace with consideration of the cracking of the precursor molecules are indicated by the red and green dashed lines, respectively, in
Figure 5b. The resulting asymmetric distribution of the adatoms (red dashed line in
Figure 5b) induces a nominal flow of adatoms from the upper to the lower terrace, even when they feature a larger kinetic barrier for incorporation to the steps from the upper terrace compared to the incorporation from the lower terrace [
26,
27,
28,
29,
30,
31]; this nominal adatom flow toward the down-step direction induces step bunching during crystal growth.
The two-particle model fits the PVT growth of 4H-SiC crystals and could explain the step bunching observed on the (000
) facet of the crystals. Pimpinelli et al. also showed that in the two-particle model, the step stiffness modifies the step bunching behavior. An important conclusion in their study is that the undulation wavelength of the step separation depends on the magnitude of the step stiffness; the weaker the step stiffness, the longer the undulation wavelength. This behavior is in accordance with our experimental results, summarized in
Table 1, according to which the undulation wavelength increases with the increase in the nitrogen doping concentration.
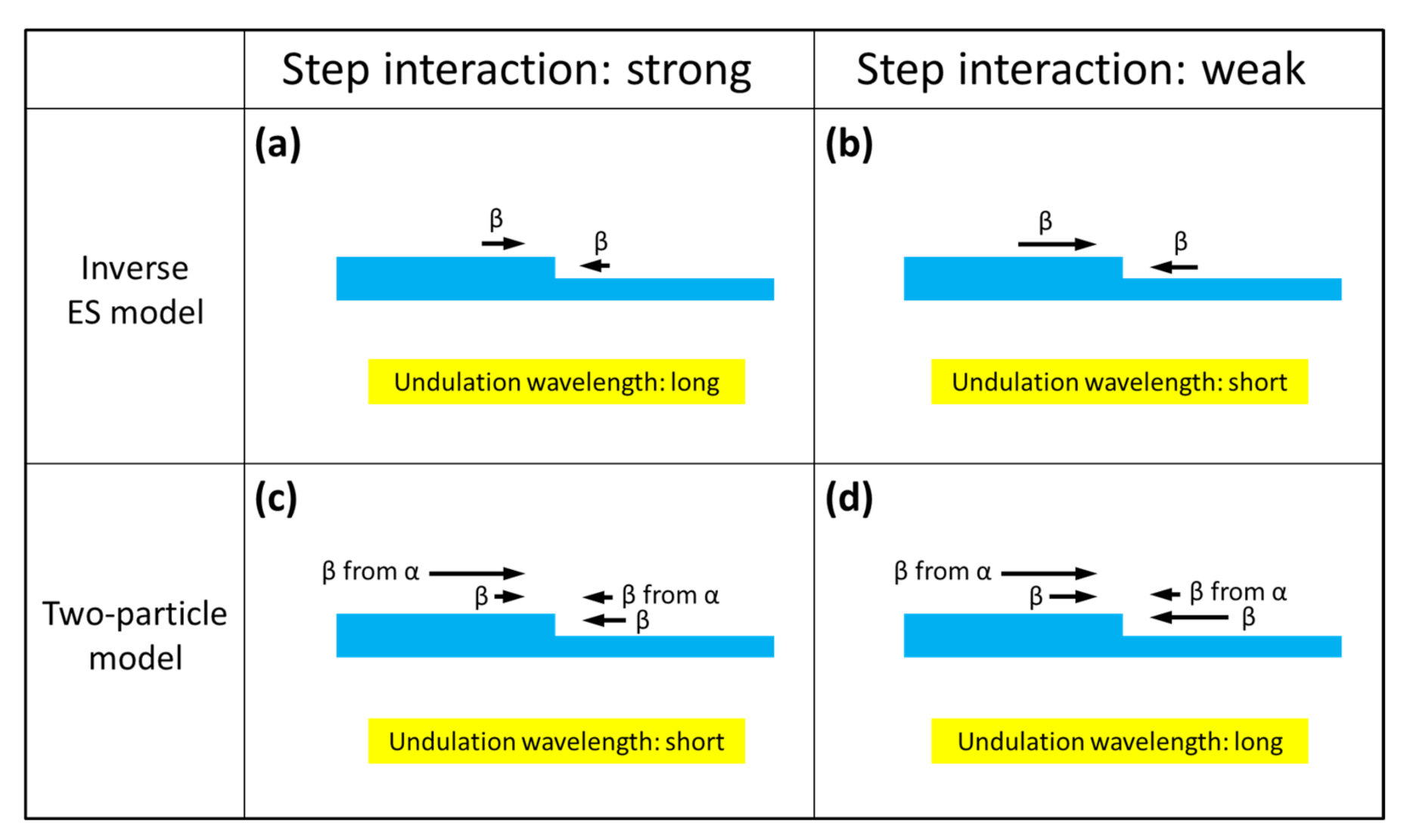
Figure 6 schematically represents the effect of the step repulsive interaction (step stiffness) on the undulation wavelength of step separation for the two models, i.e., the inverse ES model and the two-particle model.
In the figure, the direction and magnitude of the adatom flows toward the steps on a vicinal crystal surface are indicated by the black arrows, and in each section of the figure, the influence of the step repulsive interaction on the adatom flows is schematically illustrated; the adatom flows toward the steps, which express the amount of adatoms incorporated into the steps from the upper and lower terraces, are reduced by the step repulsive interaction. In the inverse ES model, the adatom flows from the upper terraces toward the steps are always larger than those from the lower terraces (
Figure 6a), and this asymmetry in the adatom flow is a kinematical driving force for step bunching. Therefore, when the step interaction (step stiffness) decreases, as the nitrogen doping concentration increases, the asymmetry (difference in the adatom flows from the upper and lower terraces) becomes larger, as schematically shown in
Figure 6b, making the kinematical driving force for the step bunching larger, resulting in a shorter undulation wavelength of step separation.
By contrast, in the two-particle model, the adatom flows from the upper terraces to steps never exceed those from the lower terraces, due to the ES barrier (
Figure 6c). In the model, however, the additional adatom flows produced by the cracking of the precursor molecules α are taken into account, making the total adatom flows from the upper terraces larger than those from the lower terraces, thus leading to step bunching during crystal growth (
Figure 6c). When the step interaction decreases in this model, the adatom flows from both terraces are enhanced, as is the case with the inverse ES model; however, the additional adatom flows produced by the cracking of the precursor molecules remain almost constant, since the distribution of the precursor molecules on the terraces is not greatly affected by the step interaction. Therefore, the difference in the adatom flows from the upper and lower terraces is reduced (
Figure 6d), giving rise to a longer undulation wavelength of step separation, when the step interaction becomes weaker as the nitrogen doping concentration increases.
As shown in
Table 1, the undulation wavelength of the step separation in the central region of the (000
) facet was nearly independent on the nitrogen doping concentration, whereas the dependence was prominent in the edge region. We assume that the largely different step separation in the two regions played a key role in this phenomenon. Firstly, the larger step separation significantly reduces the elastic step interaction, which makes the influence of nitrogen doping less effective, resulting in the undulation wavelength independent of the nitrogen doping concentration. Secondly, as already described, step bunching is generally driven by kinematical interaction between steps via adatom diffusion on the crystal surface; thus, a larger step separation tends to suppress the development of the undulated step structure. In fact, undulation of the step separation was never or rarely observed in the central region of the (000
) facet of boules A and B. By contrast, for boule C, the undulation of the step separation well developed even in the central region of the (000
) facet. This was due to the enhanced diffusion length of the adatoms on the (000
) surfaces of the heavily nitrogen-doped 4H-SiC crystals through the increased effective C/Si ratio due to heavy nitrogen doping [
33,
34]. Borysiuk et al. [
34] reported, based on density functional theory (DFT) calculations, that when the 4H-SiC (000
) surface features excess carbon atoms, they tend to convert a topmost carbon layer to sp
2 bonded configuration, which makes the (000
) surface less active in terms of adatoms and enhances their diffusion length on the surface. Nitrogen atoms, which substitute the carbon site in SiC crystals, play a similar role at the growing crystal surface; heavy nitrogen doping enriches the growing crystal surface with nitrogen atoms, which induces the sp
2 bonded configuration at the growing crystal surface and enhances the adatom diffusion length on the surface during the PVT growth of heavily nitrogen-doped 4H-SiC crystals.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}